

Abstract
Atherosclerosis, the leading death in the United State, is a disease in which a plaque builds up inside the arteries. As the plaque continues to grow, the shear force of the blood flow through the decreasing cross section of the lumen increases. This force may eventually cause rupture of the plaque, resulting in the formation of thrombus, and possibly heart attack. It has long been recognized that the formation of a plaque relates to the cholesterol concentration in the blood. For example, individuals with LDL above 190 mg/dL and HDL below 40 mg/dL are at high risk, while individuals with LDL below 100 mg/dL and HDL above 50 mg/dL are at no risk. In this paper, we developed a mathematical model of the formation of a plaque, which includes the following key variables: LDL and HDL, free radicals and oxidized LDL, MMP and TIMP, cytockines: MCP-1, IFN-γ, IL-12 and PDGF, and cells: macrophages, foam cells, T cells and smooth muscle cells. The model is given by a system of partial differential equations with in evolving plaque. Simulations of the model show how the combination of the concentrations of LDL and HDL in the blood determine whether a plaque will grow or disappear. More precisely, we create a map, showing the risk of plaque development for any pair of values (LDL,HDL).
Introduction
Atherosclerosis, hardening of the arteries, is the leading cause of death in the United States, and worldwide. The disease triggers heart attack or stroke, with total annual death of 900,000 in the United States [1] and 13 million worldwide [2].
Atherosclerosis is a disease in which a plaque builds up inside the arteries. A plaque contains low density lipoprotein (LDL), macrophages, smooth muscle cells (SMCs), platelets, and debris. The plaque constricts the lumen of the blood vessel thereby increasing the shear force of blood flow [3], [4]. As the plaque continues to grow, the increased shear force may cause rupture of the plaque, possibly resulting in the formation of thrombus (blood clot) [3], [5], ischemic stroke, and heart attack [3]–[5].
The process of plaque development begins with a lesion in the endothelial layer, allowing LDL, to move from the blood into the intima and becoming oxidized LDL (ox-LDL) by free radicals (FRs). FRs are oxidative agents continuously released by bio-chemical reactions within the body, including the intima [6]–[8]. Endothelial cells, sensing the presence of ox-LDL, secrete monocyte chemoattractant protein (MPC-1) [9], [10], which triggers recruitment of monocytes into the intima [11]. After entering the intima, monocytes differentiate into macrophages, which have an affinity for the ox-LDL [12]–[14]. The ingestion of large amounts of ox-LDL transforms the fatty macrophages into foam cells [12], [15]. Foam cells secrete chemokines which attract more macrophages [10], [12], [13]. SMCs from the media move into the intima by chemotactic forces due to MCP-1 [9], [10], and platelet-derived growth factor (PDGF) [10], [16], as well as by haptotaxis by the extracellular matrix (ECM). PDGF is secreted by macrophages, foam cells and SMCs [16], [17]. ECM is remodeled by matrix metalloproteinase (MMP) produced by a variety of cell types including SMCs [18], and is inhibited by tissue inhibitor of metalloproteinase (TIMP) produced by macrophages and SMCs [19]. Interleukin IL-12, secreted by macrophages and foam cells [10], [12], [20], contribute to the growth of a plaque by activating T cells [9], [20], [21]. Indeed, the activated T cells secrete interferon IFN-γ, which in turn activates macrophage in the intima [13], [21], [22]. At the same time that LDL enters the intima, high density lipoprotein (HDL) also enters into the intima, and becomes oxidized by free radicals [7], [8]. However, oxidized HDL (ox-HDL) is not ingested by macrophages. HDL helps prevent atherosclerosis by removing cholesterol from foam cells, and by the limiting inflammatory processes that underline atherosclerosis [23]. Furthermore, HDL takes up free radicals that are otherwise available to LDL. Some of the key players in the atherosclerosis process are shown in Fig. 1.
Figure 1. Atherosclerosis schematics: the presence of ox-LDL in the intima causes monocytes to migrate from the lumen into the intima.

Monocytes differentiate into macrophages which endocytose ox-LDL and become foam cells. SMCs are attracted from the media into intima by chemotaxis and haptotaxis. Cytokines released by macrophages, foam cells and SMCs activate T cells. T cells enhance activation of macrophages. HDL helps prevent atherosclerosis.
It has long been recognized that the cholesterol concentrations in the blood are indicators of the probability that a plaque will develop: higher LDL and lower HDL concentrations indicate a higher probability of plaque development. Public health guidelines in the U.S. specify what levels of LDL are low risk and what levels are high risk; they also specify what levels of HDL are poor and what levels are near ideal [24], [25]. However, what is more relevant is to specify the risk associated with combined levels of LDL and HDL, and this is what the present paper addresses. A schematic of the network of atherosclerosis is given in Fig. 2. In this paper, we developed a mathematical model of plaque formation by a system of partial differential equations based on Fig. 2. The aim of the model is to determine the risk of plaque formation for combined levels of LDL and HDL. In particular, we created a “risk-map” for plaque development in the LDL-HDL coordinate plane, where the first quadrant of the plane was divided into regions of high risk, low risk and no risk. Anti-cholesterol drugs are aimed at lowering high levels of LDL, but some drugs are known to also increase the level of HDL [24]. Hence such a risk-map may be important when evaluating the extend to which an anti-cholesterol drug can reduce the risk of atherosclerosis for particular individuals.
Figure 2. Schematic network of atherosclerosis.

LDL and HDL are oxidized by free radicals, and become ox-LDL and ox-HDL respectively. Ox-LDL recruits macrophages to intima. By ingesting ox-LDL, macrophages are transformed to foam cells. SMCs are attracted into the intima by MCP-1 (secreted by endothelial cells) and PDGF (secreted by macrophages and foam cells). Macrophages, foam cells and SMCs secrete IL-12, which activates T cells. IFN-γ secreted by T cells enhance the activity of macrophages which contributes the plaque built-up.
Materials and Methods
Mathematical model
In this paper, we present a mathematical model based on the network shown in Fig. 2. The model includes the variables listed in Table 1. We assume that all cells are moving with a common velocity u; the velocity is the result of movement of macrophages, T cells and SMCs into the intima. We also assume that all species are diffusing with appropriate diffusion coefficients. The equation for each species of cells X has a form
where the expression on the left-hand side includes advection and diffusion, and FX accounts for various growth factors, bio-chemical reactions, chemotaxis and haptotaxis. The equation for the chemical species are the same but without the advection term. Fig. 3 shows a 2D cross section of a blood vessel with plaque Ω, and a planar cross section of a plaque in the direction along a blood vessel.
Table 1. The variables of the model: concentrations and densities are in units of g/cm 3.
| L: | concentration of LDL | H: | concentration of HDL |
| Lox: | concentration of ox-LDL | r: | concentration of free radicals |
| P: | concentration of MCP-1 | Iγ: | concentration of IFN-γ |
| I 12: | concentration of IL-12 | G: | concentration of PDGF |
| Q: | concentration of MMPs | Qr: | concentration of TIMP |
| M: | density of macrophages | T: | density of T cells |
| S: | density of SMCs | ρ: | density of ECM |
| F: | density of foam cell | σ: | pressure (in g cm 2/day) |
| u: | fluid velocity (in cm/day) |
Figure 3. Two 2D cross sections of a plaque.
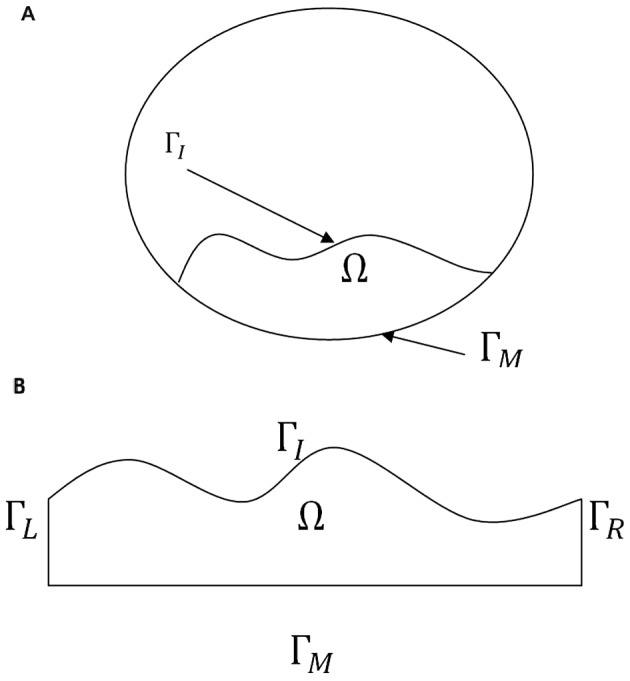
ΓM is the boundary of the intima in contact with the media, and ΓI is the boundary of the intima in contact with the lumen. In (B) ΓL and ΓR are parts of the intima.
Equations for lipoproteins [LDL (L), HDL (H), ox-LDL (Lox)] and free radical (r)
The distribution of LDL, HDL, ox-LDL and free radicals in the intima are described using reaction-diffusion equations [7],
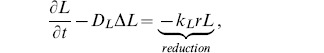 |
(1) |
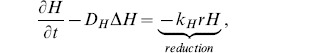 |
(2) |
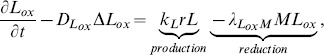 |
(3) |
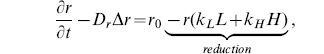 |
(4) |
where kL and kH are reaction rates of oxidization, and 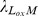 is the reduction rate of ox-LDL due to ingestion by macrophages. Eqs. (1) and (2) model the evolution of LDL and HDL concentrations. It is assumed that LDL and HDL are lost by reaction of oxidation with free radicals. Equation (3) models the production of ox-LDL due to LDL oxidation by reaction with the radicals (first term on right-hand side) and a reduction of ox-LDL through ingestion by macrophages (second term on right-hand side). Equation (4) models the evolution of free radicals concentration with baseline growth r
0.
is the reduction rate of ox-LDL due to ingestion by macrophages. Eqs. (1) and (2) model the evolution of LDL and HDL concentrations. It is assumed that LDL and HDL are lost by reaction of oxidation with free radicals. Equation (3) models the production of ox-LDL due to LDL oxidation by reaction with the radicals (first term on right-hand side) and a reduction of ox-LDL through ingestion by macrophages (second term on right-hand side). Equation (4) models the evolution of free radicals concentration with baseline growth r
0.
Equation for macrophages (M)
The evolution of macrophage density is modeled by
 |
(5) |
Here the first term on right-hand side accounts for recruitment of macrophages by MCP-1 [9], and the second term accounts for the activation of macrophages by IFN-γ [13], [21], [22].
Equation for MCP-1 (P)
The MCP-1 equation is given by
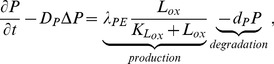 |
(6) |
where the first term on the right-hand side is the production of MCP-1 by endothelial cells, whose density is assumed to be constant, under the influence of ox-LDL [9].
Equation for T cells (T)
The density of T cells, which are primarily CD4+ T cells [20], satisfies the equation
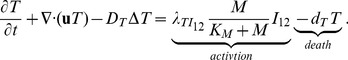 |
(7) |
In this equation, we assume that T cells are activated by IL-12 in conjunction with MHC-II (major histocompatibility complex, class II). Actually, T cells are also activated by IL-1 and IL-6 produced by macrophages and SMCs [10], [12], [13]. However, because of lack of experimental data, we do not include the IL-1 and IL-6 explicitly but instead consider their effect implicitly in estimating the parameter 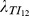 . For simplicity, we include the anti-inflammatory effect of IL-10 produced by macrophages only implicitly, by the factor
. For simplicity, we include the anti-inflammatory effect of IL-10 produced by macrophages only implicitly, by the factor 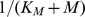 .
.
Equation for IFN-γ (Iγ)
The dynamics of IFN-γ concentration is modeled by
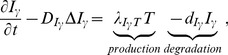 |
(8) |
where the first term on right-hand side represents production of Iγ by T cells [21].
Equation for SMCs (S)
The equation of the SMCs density is given by
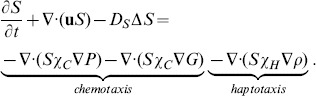 |
(9) |
The first two terms on right-hand side account for chemotaxis by MCP-1 [9], [10], and PDGF [10], [16], and the last term accounts for haptotaxis by ECM.
Equation for IL-12 (I 12)
The concentration of IL-12 is modeled by
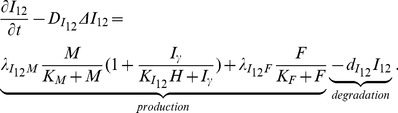 |
(10) |
The first term of right-hand side is the production of I
12 by macrophages enhanced by Iγ and resisted by HDL [23]. The production of I
12 by macrophages is resisted by I
10 (which, for simplicity, is accounted by the factor 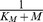 ) [10], [12]. The second term represents the production of I
12 by foam cells [20].
) [10], [12]. The second term represents the production of I
12 by foam cells [20].
Equations for PDGF (G), MMP (Q) and TIMP (Qr)
We have the following sets of reaction diffusion equations for the chemokines (G, Q and Qr):
 |
(11) |
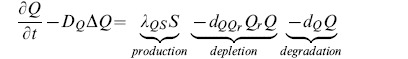 |
(12) |
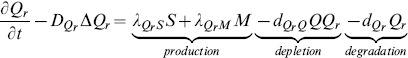 |
(13) |
In equation (11), PDGF is produced by macrophages, foam cells, and SMCs [16], [17]. In equation (12), MMP is secreted by SMCs [18] (first term on right-hand side), and is lost by binding with TIMP (second term). In equation (13), TIMP is produced by SMCs and macrophages [19].
Equation for foam cells (F)
Macrophages that have ingested a large amount of ox-LDL become foam cells [7], [12], [15], so we have
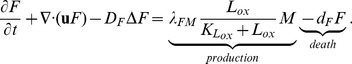 |
(14) |
Equations for ECM (ρ) and pressure (σ)
We assume that the intima has the constituency of a porous medium. Then, by Darcy's law, the velocity u of the cells is given by
| (15) |
where σ is the pressure. We also assume that the total density of all the cells plus the concentration of ρ is constant. This constant should be smaller than the average density of a plaque, 1.22±0.03 g/cm 3 [26], because plaques contain some debris, which are not included in our model. We take the constant to be 1 g/cm 3, i.e.,
| (16) |
We assume that all cells are approximately of the same volume and surface area, so that the diffusion coefficients of the all cells have the same coefficient, D. By adding Eqs. (5), (7), (9) and (14), we get
| (17) |
where
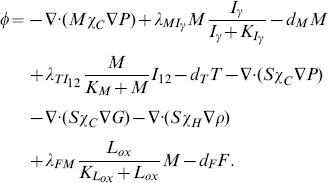 |
(18) |
Eq. (17) gives a relation between ρ and σ. We next derive an equation for ρ. The ECM is degraded by MMP [27], and is remodeled by macrophages and SMCs [18], [27]. For simplicity, we take the remodeling rate to be a constant, λρ, as in [28]. Then the equation of the density of ECM is given by
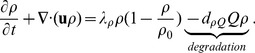 |
(19) |
Since  this equation can be written in the form
this equation can be written in the form
where
| (20) |
Adding this equation and (17), we get our final equation for σ:
| (21) |
The equation for 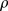 can then be written as
can then be written as
| (22) |
Boundary conditions
For simplicity, we consider only 2-dimensional plaques as in Fig. 3. Then the boundary of the plaque consists of i) ΓM, in contact with media; ii) a free boundary ΓI, inside the lumen, and iii) two more vertical boundaries ΓL and ΓR of the intima in the case of Fig. 3(B).
Boundary conditions on ΓI
We assume flux boundary conditions of the form
| (23) |
for X = L, H, M, T, and non-flux boundary conditions for all other variables,
| (24) |
where 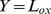 , r, P, Iγ, S, I, I
12, G, Q, Qr, F. The boundary values for ρ are determined by Eq. (16). The coefficient
, r, P, Iγ, S, I, I
12, G, Q, Qr, F. The boundary values for ρ are determined by Eq. (16). The coefficient 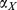 is a constant except for M, and
is a constant except for M, and 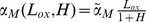 , since ox-LDL attracts monocytes [11], while HDL limits the inflammation process[23]. Note that L
0 and H
0 are the LDL and HDL concentrations in the blood, so we shall be interested to see how these concentrations determine whether a small plaque will grow or shrink.
, since ox-LDL attracts monocytes [11], while HDL limits the inflammation process[23]. Note that L
0 and H
0 are the LDL and HDL concentrations in the blood, so we shall be interested to see how these concentrations determine whether a small plaque will grow or shrink.
As in [29]–[31], we assume that the free boundary ΓI is held together by cell-to-cell adhesion forces so that
where κ is the mean curvature of the surface ΓI. (If ΓI is circular, then κ is the reciprocal of the radius) Furthermore, the continuity condition 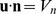 , where n is the outward normal and Vn is the velocity of the free boundary ΓI in the direction n, yields the relation
, where n is the outward normal and Vn is the velocity of the free boundary ΓI in the direction n, yields the relation
| (25) |
Boundary conditions on ΓM
We assume non-flux boundary conditions for all variables except ρ and S on ΓM: For S, we have
where S
0 is SMCs density in the media, and for simplicity we take 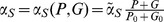 since MCP-1 and PDGF attract SMCs from the media. As in the case of ΓI, the boundary values of ρ are determined by Eq. (16).
since MCP-1 and PDGF attract SMCs from the media. As in the case of ΓI, the boundary values of ρ are determined by Eq. (16).
Boundary conditions on ΓL and ΓR
We assume the periodic boundary conditions on ΓL and ΓR.
Parameter estimation
Table 2 lists the range of molecular weights of proteins and Table 3 lists their range of concentration. In the second columns in Tables 2 and 3, we indicate the (intermediate) values used in the simulations. The Tables 2 and 3 are used to estimate some of the model parameters. A summary of all the model parameters is given in Tables 4 and 5.
Table 2. Molecular weights.
| Protein | Weight (kda) | Explanation |
| LDL | 549 | Over 95% of the LDL protein mass is apolipoprotein |
| B-100 (apo B-100, 549 kDa (1000 g/mol)) [63]. | ||
| HDL | 105 | The range of weight of HDL is 105–130 [64]. |
| Free radical | 0.51 kda | Free radicals include DPPH (0.39 kda), |
| ABTS (0.51 kda) and superoxide anion (0.81 kda) [65]. | ||
| IFN-γ | 17 | IFN-γ is described as a 17 kDa peptide [66]. |
| PDGF | 35 | There are two PDGF polypeptides: |
| PDGF-I with a molecular weight of about 35 kda, and | ||
| PDGF-II with a molecular weight of about 32 kda [67]. | ||
| MCP-1 | 8.9 | [68] |
| IL-12 | 70 | [69] |
| MMP | 52 | MMP-1 has two major species of molecular |
| mass, 57 kDa and 52 kDa [48]. | ||
| TIMP | 25 | The molecular weights of TIMP-1, TIMP-2 and TIMP-3 |
| are 28.5 kDa 21 kDa and 27 kDa respectively [48]. |
Table 3. Concentrations of proteins and cells.
| Proteins & cells | Concentration (gcm −3) | Explanation |
| LDL | 7×10−4–1.9×10−3 | Range is 70–190 mg/dl [25], [70]. |
| HDL | 4×10−4–6×10−4 | Range is 40–60 mg/dl [25], [70]. |
| IFN-γ | 10−9 | Range is of 0.1–10.0 ng/mL [10]. |
| PDGF | 1.5×10−8 | Range in normal humans blood |
| 17.5±3.1 ng/mL [72]. | ||
| MCP-1 | 3×10−10 | 300 pg/ml [73] |
| IL-12 | 5×10−10 | Range 200–800 pg/ml [20]. |
| MMP | 3×10−8 | Range in plasma is 10∼60 ng/ml [74]. |
| TIMP | 3×10−8 | Range in plasma is 10∼60 ng/ml [74] |
| SMC | 6×10−3 | Range 7,500,000–10,000,000 cells per ml [38]. |
| Monocyte | 5×10−5 | Range from 20,000 to 100,000 cells per ml [75]. |
| T cell | 1×10−3 | Range of CD4+ T cells in healthy normal adult |
| of 500,000 to 1,500,000 cells per ml [49]. |
Table 4. Parameters' description and value.
| Parameter | Description | Value |
| kL | reaction rate of LDL + Radical→ox-LDL | 2.35×10−4 g −1 cm 3 day−1 [7], [34], [35] |
| kH | reaction rate of HDL + Radical→ox-HDL | 5.29×10−6 g −1 cm 3 day−1 [7], [34] |
| DL | diffusion coefficient of LDL | 29.89 cm 2 day−1 [33], [34], [37] & estimated |
| DH | diffusion coefficient of HDL | 3.93 cm 2 day−1 [33], [34], [37] & estimated |
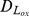
|
diffusion coefficient of oxidized LDL | 29.89 cm 2 day−1 [33], [34], [37] & estimated |
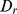
|
diffusion coefficient of radicals | 2.05×10−1 cm 2 day−1 [33], [34], [37] & estimated |
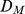
|
diffusion coefficient of macrophage | 8.64×10−7 cm 2 day−1 [28], [36] |
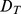
|
diffusion coefficient of T-cell | 8.64×10−7 cm 2 day−1 [28], [36] |

|
diffusion coefficient of IFN-γ | 1.08×102 cm 2 day−1 [76] |
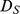
|
diffusion coefficient of SMCs | 8.64×10−7 cm 2 day−1 [28], [36] |
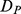
|
diffusion coefficient of MCP-1 | 17.28 cm 2 day−1 [44] |
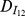
|
diffusion coefficient of IL-12 | 1.08×102 cm 2 day−1 [76] |
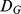
|
diffusion coefficient of PDGF | 8.64×10−2 cm 2 day−1 [42] |
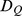
|
diffusion coefficient of MMP | 4.32×10−2 cm 2 day−1 [38] |
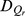
|
diffusion coefficient for TIMPs | 4.32×10−2 cm 2 day−1 [33], [37], [38] & estimated |
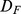
|
diffusion coefficient of foam cells | 8.64×10−7 cm 2 day−1 [28], [36] |
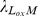
|
rate of ox-LDL ingestion by macrophages | 10 gcm −3 day−1 [7] |
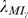
|
activation rate of macrophages by IFN-γ | 0.005 day−1 [39] & estimated |
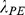
|
production rate of MCP-1 | 8.65×10−10 gcm −3 day−1 [44] & estimated |
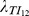
|
activation rate of T cells by IL-12 | 1×106 day−1 [39], [40], [49], [74] & estimated |
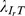
|
production rate of IFN-γ by T cells | 0.066 day−1 [45], [77] |
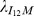
|
production rate of IL-12 by macrophages | 3×10−7 gcm −3 day−1 [45] |
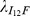
|
production rate of IL-12 by foam cells | 1×10−7 gcm −3 day−1 [45] & estimated |
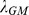
|
production rate of PDGF by macrophages | 0.1 day−1 [41], [42] & estimated |
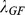
|
production rate of PDGF by foam cells | 0.033 day−1 [41], [42] & estimated |
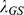
|
production rate of PDGF by SMCs | 0.5 day−1 [41], [42] & estimated |
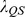
|
production rate of MMP by SMCs | 3×10−4 day−1 [28] |
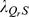
|
production rate of TIMP by SMCs | 3×10−5 day−1 [28] & estimated |
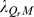
|
production rate of TIMP by macrophages | 6×10−5 day−1 [28] & estimated |
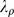
|
remodeling rate of ECM | 0.432 day−1 [28] |
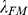
|
activation rate of foam cells | 0.12 day−1 [39] & estimated |
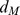
|
death rate of macrophage | 0.015 day−1 [39] |
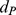
|
degradation rate of MCP-1 | 1.73 day−1 [44] |
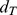
|
death rate of T cell | 0.33 day−1 [39], [40] |
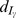
|
degradation rate of IFN-γ | 0.69 day−1 [37] |
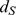
|
death rate of SMC | 0.86 day−1 [7] |
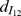
|
degradation rate of IL-12 | 1.188 day−1 [39], [40] |
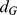
|
degradation rate of PDGF | 3.84 day−1 [42] |
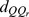
|
binding rate of MMP to TIMP | 4.98×108 cm 3 g −1 day−1 [44], [48] & estimated |
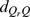
|
binding rate of TIMP to MMP | 1.04×109 cm 3 g −1 day−1 [44], [48] & estimated |
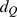
|
degradation rate of MMP | 4.32 day−1 [37] |
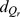
|
degradation rate of TIMP | 21.6 day−1 [46] & estimated |
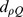
|
degradation rate of ECM due to MMP | 2.59×107 cm 3 g −1 day−1 [36] |
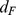
|
death rate of foam cell | 0.03 day−1 [39] & estimated |
Table 5. Parameters' description and value.
| Parameter | Description | Value |
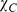
|
chemotactic sensitivity parameter |
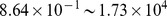 cm
5
g
−1 day−1
[36], [37] (10)*
cm
5
g
−1 day−1
[36], [37] (10)*
|
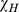
|
haptotaxis parameter |
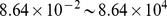 cm
5
g
−1 day−1
[36], [37] (102)*
cm
5
g
−1 day−1
[36], [37] (102)*
|
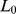
|
source/influx of LDL in blood |
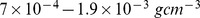 [25]
[25]
|
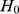
|
source/influx of HDL in blood |
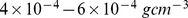 [25]
[25]
|
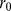
|
source/influx of free radical into intima | 0.26 gcm −3 day−1 [34] |
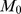
|
source/influx of macrophages from blood |
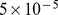
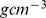 [75]
[75]
|
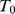
|
source/influx of T cells into intima |
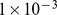
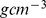 [49]
[49]
|
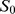
|
source/influx of SMCs into intima |
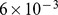
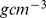 [71]
[71]
|
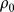
|
ECM density |
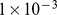
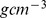 [28]
[28]
|
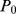
|
MCP-1 concentration |
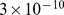 [73]
[73]
|
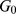
|
PDGF concentration |
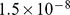 [72]
[72]
|
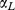
|
influx rate of LDL into intima | 1.0 cm −1 estimated |
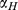
|
influx rate of HDL into intima | 1.0 cm −1 estimated |
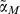
|
influx rate of macrophage into intima | 0.2 cm −1 estimated |
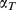
|
influx rate of T cells into intima | 0.05 cm −1 estimated |
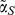
|
influx rate of of SMCs into intima | 0.2 cm −1 estimated |
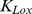
|
ox-LDL saturation for production of MCP-1 | 0.5 gcm −3 [64] & estimated |
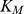
|
macrophages saturation for activation of T cells | 2.5×10−5 gcm −3 [75] & estimated |
| and production of IL-12 | ||
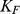
|
foam cells saturation for production of IL-12 | 2.5×10−5 gcm −3 [75] & estimated |
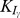
|
IFN-γ saturation for activation of macrophages | 1×10−11 gcm −3 [39] |
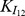
|
IFN-γ saturation for production of IL-12 | 7×10−11 gcm −3 [45] |
* Values chosen in the simulation.
Reaction rates
To estimate some of the parameters in the equations for proteins, we shall use the concept of “accessible surface area” [32], [33] of a protein p, or briefly “area,” 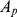 , which is roughly the minimum surface area of the smooth shapes containing the protein. It was estimated in [34], that
, which is roughly the minimum surface area of the smooth shapes containing the protein. It was estimated in [34], that 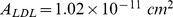 , and
, and 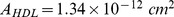 , so that their ratio is
, so that their ratio is
Accordingly, the corresponding reaction rates of the oxidation, kL and kH, are related by 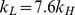 . Moreover, the reaction rate of oxidation of LDL by free radicals is
. Moreover, the reaction rate of oxidation of LDL by free radicals is 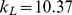
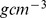 day−1
[7], [34], [35], so that
day−1
[7], [34], [35], so that 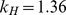 g cm
−3 day−1.
g cm
−3 day−1.
Diffusion coefficients
We assume that the diffusion coefficients of all the cells are the same, and take them to be 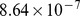 cm
2 day−1
[28], [36]. In order to estimate the diffusion coefficients of the various proteins, we assume that the diffusion coefficient of protein p, Dp, is proportional to its area Ap, i.e.,
cm
2 day−1
[28], [36]. In order to estimate the diffusion coefficients of the various proteins, we assume that the diffusion coefficient of protein p, Dp, is proportional to its area Ap, i.e., 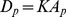 , where we take K to be the same for all small molecules. For glucose, which is a monomeric globular protein,
, where we take K to be the same for all small molecules. For glucose, which is a monomeric globular protein, 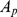 can be computed in terms of the molecular weight
can be computed in terms of the molecular weight 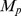 , by the formula
, by the formula 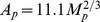 [33], and
[33], and 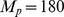 dalton [28],
dalton [28], 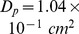 day−1
[37]. Hence, for glucose, K is determined by
day−1
[37]. Hence, for glucose, K is determined by
 |
(26) |
We can now compute 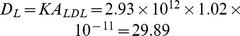
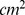 day−1 and
day−1 and 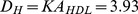
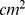 day−1. Free radicals are monomeric globular proteins (average weight is 500 da, Table 2). Hence
day−1. Free radicals are monomeric globular proteins (average weight is 500 da, Table 2). Hence
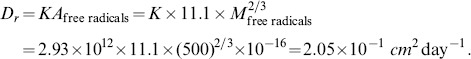 |
The diffusion coefficient of MMP is 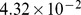
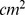 day−1
[38]. We assume that the diffusion coefficient of TIMP is same as that of MMP.
day−1
[38]. We assume that the diffusion coefficient of TIMP is same as that of MMP.
Production rates
We assume that, in Eq. (7), 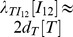 , where
, where 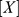 denotes the average concentration of X. We take
denotes the average concentration of X. We take 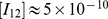 gcm−3,
gcm−3, 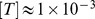 gcm−3 from Table 3, and
gcm−3 from Table 3, and 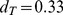 day−1
[39], [40]. Then
day−1
[39], [40]. Then 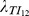 is estimated by
is estimated by 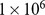 day−1.
day−1.
PDGF is produced by SMCs, and likely also by endothelial cells and macrophages [41]. In wound healing, macrophages produce PDGF at rate of 5.76 day−1
[42]. Since the plaque formation is a much slower process, we take this rate 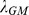 to be much smaller, i.e.,
to be much smaller, i.e., 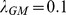 day−1. Since SMCs production rate of PDGF is higher than that by macrophages [41], we take
day−1. Since SMCs production rate of PDGF is higher than that by macrophages [41], we take 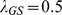 day−1.
day−1.
The production rate of MMP by tumor cells was estimated in [28] to be 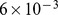 day−1. We assume that SMCs produce MMP at a much lower rate, namely,
day−1. We assume that SMCs produce MMP at a much lower rate, namely, 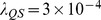 day−1. Since SMCs produce MMP to enable them move into the intima by haptotaxis, we assume that they produce TIMP at a lower rate than MMP, and take
day−1. Since SMCs produce MMP to enable them move into the intima by haptotaxis, we assume that they produce TIMP at a lower rate than MMP, and take 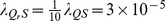 day−1. As macrophages produce most of the TIMP [43], we take the production rate of TIMP by macrophages to be
day−1. As macrophages produce most of the TIMP [43], we take the production rate of TIMP by macrophages to be 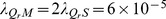 day−1.
day−1.
We assume that the production rate of MCP-1 by endothelial cells, 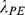 , is twice that of
, is twice that of 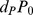 , where
, where 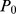 is the concentration of MCP in the blood, which is equal to
is the concentration of MCP in the blood, which is equal to 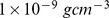 [44]. We assume that
[44]. We assume that 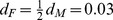 day−1
[39], and that, in Eq. (14),
day−1
[39], and that, in Eq. (14), 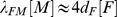 and
and 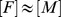 , so that
, so that 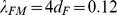 day−1.
day−1.
By [45], 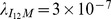 day−1. We assume that foam cells have lower production rates of I
12 and PDGF than macrophages, and take
day−1. We assume that foam cells have lower production rates of I
12 and PDGF than macrophages, and take 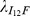 and
and 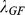 to be one third of the values of
to be one third of the values of 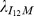 and
and 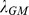 , respectively, so that
, respectively, so that 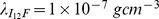 day−1, and
day−1, and 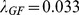 day−1.
day−1.
Degradation rates
The degradation rate of MMP is 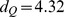 day−1
[37]. Since TIMP has a short half life compared to MMP [46], we take its degradation rate to be
day−1
[37]. Since TIMP has a short half life compared to MMP [46], we take its degradation rate to be 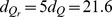 day−1.
day−1.
In [47], the binding rate of MMP and TIMP is reported to be 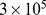
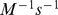 , where
, where 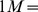 the mass per mole, and the molecular weights of MMP and TIMP are 52 kda and 25 kda, respectively [48]. Accordingly, we derive the binding rate per Molar per second (by same formula as in [44]),
the mass per mole, and the molecular weights of MMP and TIMP are 52 kda and 25 kda, respectively [48]. Accordingly, we derive the binding rate per Molar per second (by same formula as in [44]),
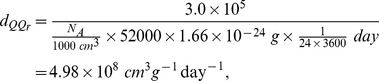 |
and
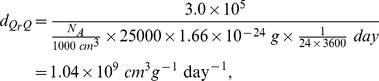 |
where NA is called the Avogadro number, and is the number of molecular per dm
3. 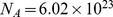 , and
, and 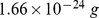 is the mass of a proton for atomic mass unit.
is the mass of a proton for atomic mass unit.
Other parameters
The range of macrophages in the blood is 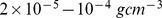 [75]; we take
[75]; we take 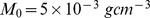 . The range of T cells in the blood is
. The range of T cells in the blood is 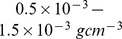 [49]; we take
[49]; we take 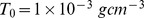 . The range of SMCs is
. The range of SMCs is 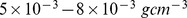 [38]; we take
[38]; we take 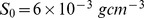 . We assume that
. We assume that 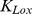 is half of
is half of 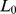 in Eq. (6), and similarly,
in Eq. (6), and similarly, 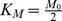 in Eq. (7), and
in Eq. (7), and 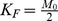 in Eq. (10). We assume that the influx of LDL and HDL into the intima is larger than the influx of macrophages and SMCs, and take
in Eq. (10). We assume that the influx of LDL and HDL into the intima is larger than the influx of macrophages and SMCs, and take 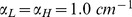 , and
, and 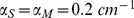 . The influx of T cells is assumed to be smaller than that of macrophages, and we take
. The influx of T cells is assumed to be smaller than that of macrophages, and we take 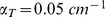 .
.
Numerical methods
Finite element implementation
In order to illustrate our numerical method, we consider the following diffusion equation with Robin boundary conditions:
| (27) |
where 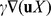 is an advection term, and either
is an advection term, and either 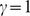 or
or 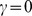 (no advection), and
(no advection), and 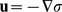 . Multiplying the differential equation by an arbitrary function
. Multiplying the differential equation by an arbitrary function  , and performing integration by parts using the boundary conditions, we get
, and performing integration by parts using the boundary conditions, we get
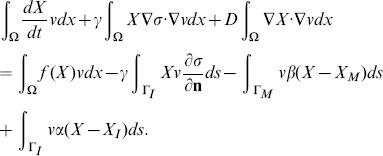 |
(28) |
This is an equivalent formulation of the system (27), which is better suited for simulation.
Similarly, Eq. (21) for σ has the equivalent form:
| (29) |
for an arbitrary function  .
.
Galerkin discretization
The standard Galerkin discretization method uses finite dimensional subspaces 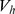 to approximate the solution X. Let
to approximate the solution X. Let 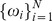 be a basis of
be a basis of 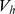 , where N is the number of nodes within the triangulation K. Let
, where N is the number of nodes within the triangulation K. Let 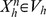 denote the numerical approximation of X at time
denote the numerical approximation of X at time 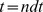 , where dt is the time step,
, where dt is the time step, 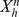 is written as a sum
is written as a sum
| (30) |
for coefficient 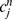 to be determined. If
to be determined. If 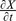 is approximated by
is approximated by 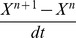 , then (28) is equivalent to
, then (28) is equivalent to
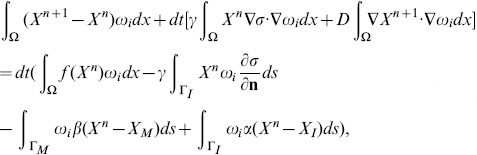 |
(31) |
or,
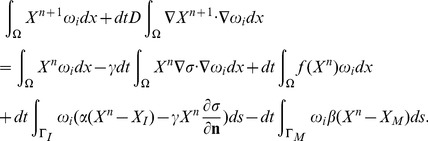 |
(32) |
Recalling (30), we can rewrite the system (32) as a linear system of equations
| (33) |
where 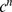 is the vector of
is the vector of 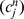 , and the coefficient matrix
, and the coefficient matrix 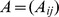 and the right-hand side
and the right-hand side 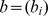 are defined by
are defined by
and
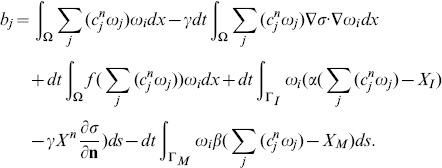 |
Similarly, setting 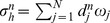 , where
, where 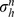 is a numerical approximation of σ at time ndt, Eq. (29) can be written as follows
is a numerical approximation of σ at time ndt, Eq. (29) can be written as follows
| (34) |
where 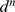 is the vector
is the vector 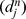 , B is a matrix
, B is a matrix 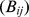 ,
, 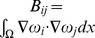 , and
, and 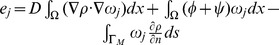 .
.
Outline of the procedure
Suppose the domain Ω(t) has polygonal boundaries ΓI(t) and ΓO(t). Then we can cover the closure 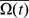 of
of 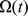 by a regular triangulation
by a regular triangulation 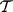 of triangles, i.e.,
of triangles, i.e.,  where each T is a closed triangle. The triangular mesh, which is a basic thing that Finite Elements requires, is generated by distmesh [50], which is a mesh generation tool implemented in MATLAB, and our algorithm is outlined in
Algorithm S1
. For the detailed implementations, such as: construct basis functions over the triangulation, assemble the stiffness matrix, etc, see references [51], [52].
where each T is a closed triangle. The triangular mesh, which is a basic thing that Finite Elements requires, is generated by distmesh [50], which is a mesh generation tool implemented in MATLAB, and our algorithm is outlined in
Algorithm S1
. For the detailed implementations, such as: construct basis functions over the triangulation, assemble the stiffness matrix, etc, see references [51], [52].
Results
Numerical simulation is initialized by a small formed plaque. (see Figs. 4, 5, 6). Five combined levels of LDL and HDL (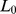 and
and 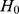 ) are tested for 300 days:
) are tested for 300 days:
Figure 4. Simulations for the atherosclerosis model of 300 days after an initial plaque is formed with H 0 = 40 mg/dL and L 0 = 190 mg/dL.
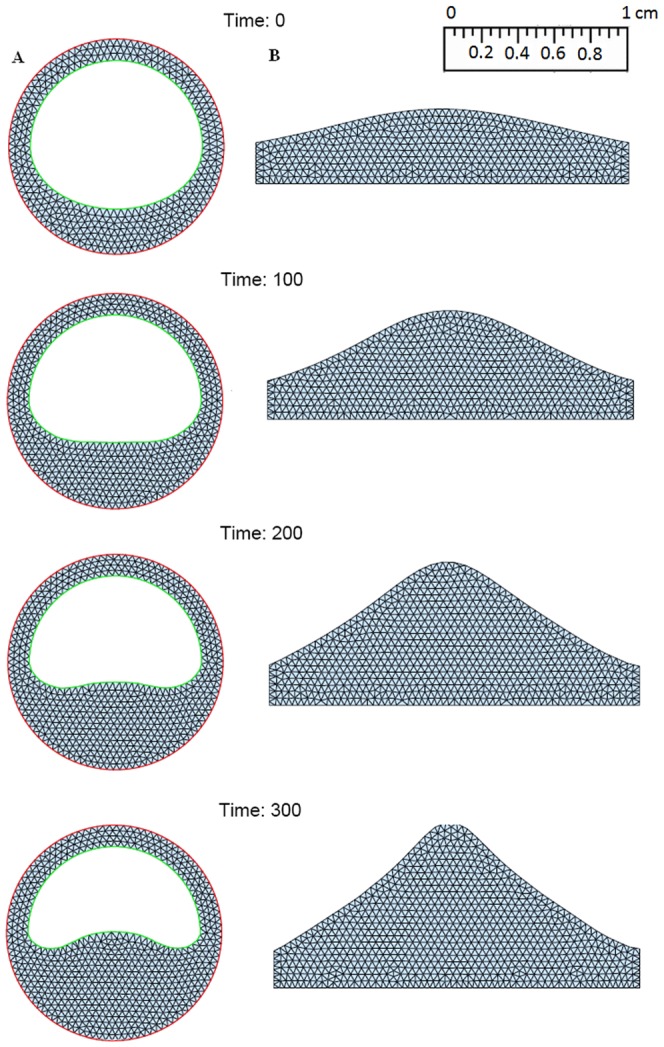
(A: Cross sections of a blood vessel, B:Cross sections along the blood vessel).
Figure 5. Simulations for the atherosclerosis model of 300 days after an initial plaque is formed with H 0 = 50 mg/dL and L 0 = 130 mg/dL.
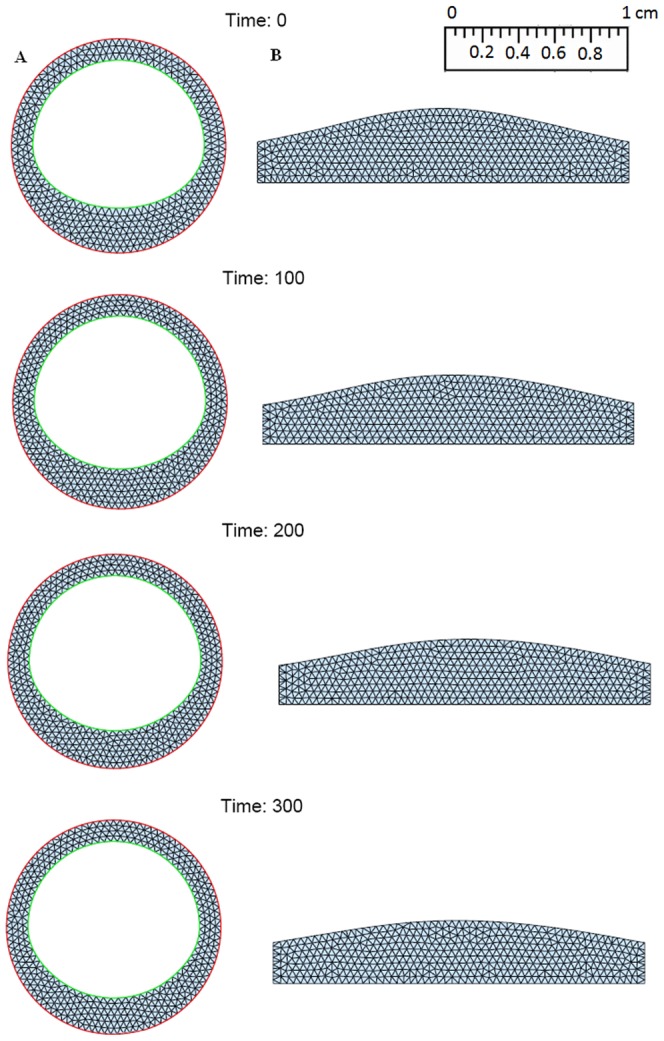
(A: Cross section of a blood vessel; B: Cross section along the blood vessel).
Figure 6. Simulations for the atherosclerosis model of 300 days after an initial plaque is formed with H 0 = 60 mg/dL and L 0 = 70 mg/dL.
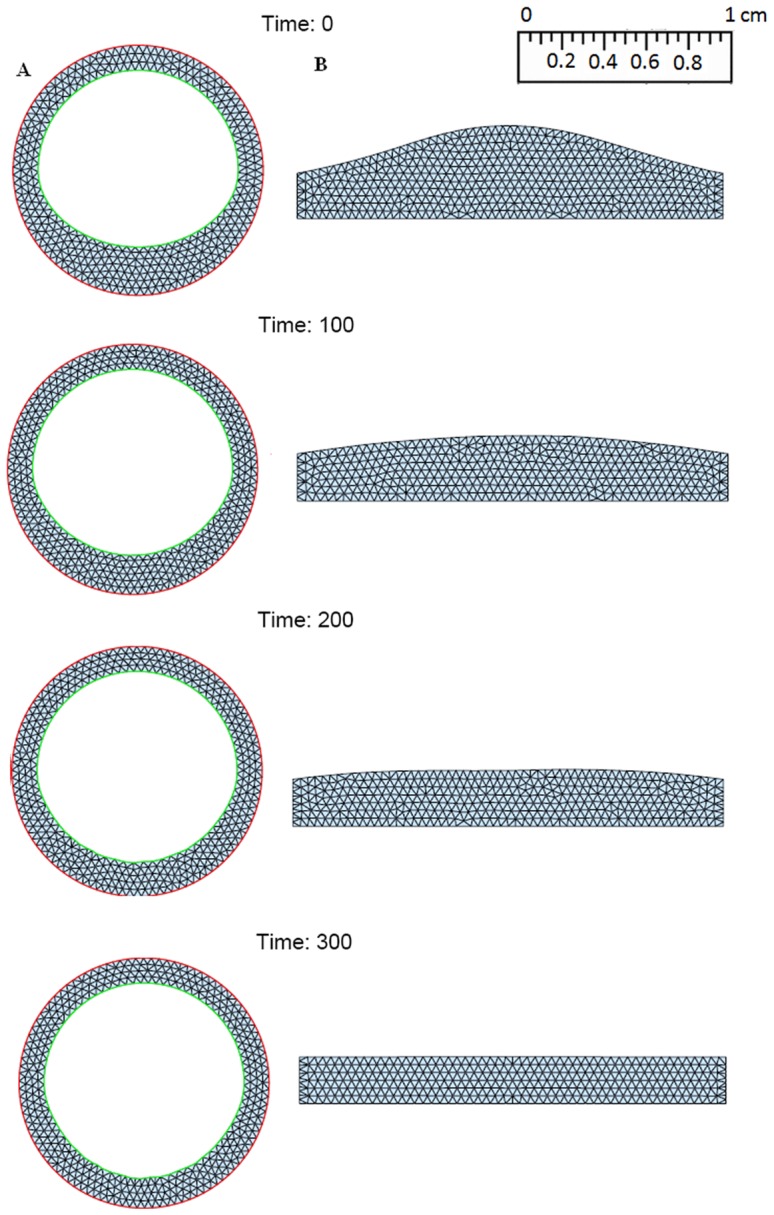
(A: Cross section of a blood vessel; B: Cross section along the blood vessel).
 : a small plaque doubles in size at 300 days;
: a small plaque doubles in size at 300 days;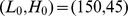 : a small plaque increases approximately 50% at 300 days;
: a small plaque increases approximately 50% at 300 days;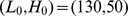 : a small plaque remains small at 300 days;
: a small plaque remains small at 300 days;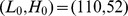 : a small plaque decreases approximately 30% at 300 days;
: a small plaque decreases approximately 30% at 300 days;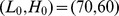 : a small plaque almost disappear at 300 days.
: a small plaque almost disappear at 300 days.
Fig. 4 shows the growth of the plaque in case (a), Fig. 5 shows the shrinkage of the plaque in case (c), and Fig. 6 shows almost no plaque in case (e). In Fig. 7, the weight of the plaque, the summation of total cells, namely, 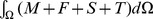 , is plotted for these five scenarios of combined levels of LDL and HDL. Similarly to Fig. 7, we show in supporting information files how the populations of macrophages, SMCs, foam cells and T cells, as well as the concentration of ox-LDL, IFN-γ and IL-12, vary for different levels of LDL and HDL shown in Figs. S1, S2, S3, S4, S5, S6, S7.
, is plotted for these five scenarios of combined levels of LDL and HDL. Similarly to Fig. 7, we show in supporting information files how the populations of macrophages, SMCs, foam cells and T cells, as well as the concentration of ox-LDL, IFN-γ and IL-12, vary for different levels of LDL and HDL shown in Figs. S1, S2, S3, S4, S5, S6, S7.
Figure 7. Plaque weights for different levels of LDL and HDL.
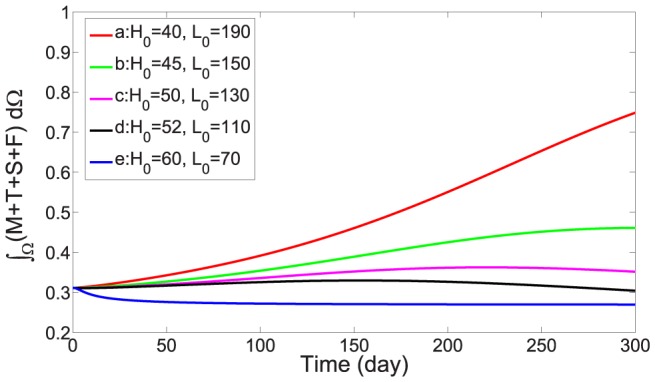
The units of H 0 and L 0 are mg/dL.
Fig. 8 shows a risk-map of plaque development. To create the risk-map, we divided the LDL axis by 121 equidistant points, i.e, 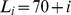 (
( ), and divided the HDL axis by 21 equidistant points, i.e.,
), and divided the HDL axis by 21 equidistant points, i.e., 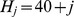 (
(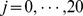 ). For each pair
). For each pair 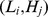 , we computed the weight of the plaque,
, we computed the weight of the plaque, 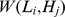 after 100 days on the domain shown in Fig. 3 (B), and formed the risk matrix
after 100 days on the domain shown in Fig. 3 (B), and formed the risk matrix
where 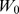 is the initial weight of the plaque. The vertical axis on the right of Fig. 8 shows the legend of the percentage of plaque growth or shrinkage. Accordingly, we divided the LDL-HDL plane into three regions: region I predicts high risk of plaque development, region III predicts no risk, and the intermediate region II predicts low risk.
is the initial weight of the plaque. The vertical axis on the right of Fig. 8 shows the legend of the percentage of plaque growth or shrinkage. Accordingly, we divided the LDL-HDL plane into three regions: region I predicts high risk of plaque development, region III predicts no risk, and the intermediate region II predicts low risk.
Figure 8. Risk map for plaque development: Region I high risk; Region II low risk; Region III no risk.

The five points 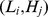 whose plaque's weight was simulated in Fig. 7 over a period of 300 days are indicated by “x”.
whose plaque's weight was simulated in Fig. 7 over a period of 300 days are indicated by “x”.
Sensitivity analysis
In order to support the robustness of the simulation results, we ran sensitivity analysis on parameters which appear in the differential equations and in the boundary conditions. The parameters chosen are those whose baseline was somewhat crudely estimated while at the same time they seem to play an important role in the development of the plaque. Specifically, we chose all the 15 production rate parameters from the third box of Table 4, all the 5 influx rate parameters from the third box of Table 5, and 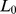 ,
, 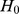 . We list all these parameters with their range, baseline and unit in Table 6.
. We list all these parameters with their range, baseline and unit in Table 6.
Table 6. Parameters chosen for sensitivity analysis.
| Parameter | Range | Baseline | Unit |
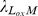
|
[5,20] | 10 |
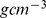 day−1 day−1
|
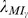
|
[0.002, 0.01] | 0.005 | day−1 |
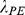
|
[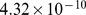 , , 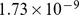 ] ] |
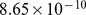
|
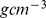 day−1 day−1
|
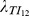
|
[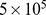 , , 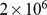 ] ] |
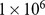
|
day−1 |
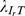
|
[0.033, 0.132] |
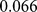
|
day−1 |
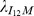
|
[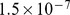 , , 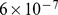 ] ] |
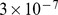
|
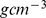 day−1 day−1
|
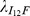
|
[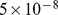 , , 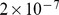 ] ] |
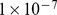
|
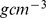 day−1 day−1
|
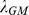
|
[0.05, 0.2] | 0.1 | day−1 |
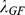
|
[0.016, 0.066] | 0.033 | day−1 |
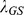
|
[0.25, 1] | 0.5 | day−1 |
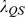
|
[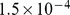 , , 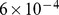 ] ] |
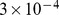
|
day−1 |
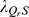
|
[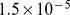 , , 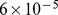 ] ] |
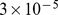
|
day−1 |
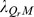
|
[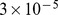 , , 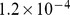 ] ] |
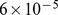
|
day−1 |
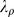
|
[0.266, 0.864] | 0.432 | day−1 |
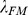
|
[0.06, 0.24] | 0.12 | day−1 |
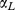
|
[0.5, 2.0] | 1.0 |
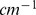
|
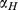
|
[0.5, 2.0] | 1.0 |
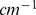
|
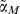
|
[0.1, 0.4] | 0.2 |
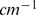
|
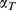
|
[0.025, 0.1] | 0.05 |
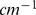
|
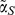
|
[0.1, 0.4] | 0.2 |
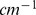
|
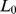
|
[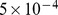 , , 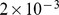 ] ] |
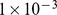
|
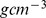
|
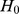
|
[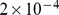 , , 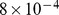 ] ] |
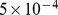
|
|
Following the sensitivity analysis method described in [53], we performed Latin hypercube sampling and generated 100 samples to calculate the partial rank correlation coefficients (PRCC) and p-values with respect to the weight of the plaque after 300 days. The PRCCs are shown in Fig. 9, and all the p-values (not shown here) are less than 0.01. A positive PRCC (i.e., positive correlation) means that an increase in the parameter value will increase the weight of the plaque while a negative PRCC (i.e., negative correlation) means increase in the parameter will decrease the weight of the plaque. We note that 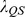 is positively correlated, as it should be. Indeed, if
is positively correlated, as it should be. Indeed, if 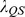 is increased then MMP (Q) is increased (Eq. (12)) so that ECM (ρ) is decreased (Eq. (19)) and hence the plaque weight
is increased then MMP (Q) is increased (Eq. (12)) so that ECM (ρ) is decreased (Eq. (19)) and hence the plaque weight 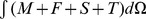 is increased (Eq. (16)). As another example, note that
is increased (Eq. (16)). As another example, note that 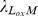 is negatively correlated. Indeed, if
is negatively correlated. Indeed, if 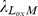 is increased then
is increased then 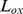 is decreased (Eq. (3)), and
is decreased (Eq. (3)), and 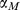 in the boundary condition will decrease, leading to smaller M, and then to smaller T and F. Similar explanation can be given to the other parameters.
in the boundary condition will decrease, leading to smaller M, and then to smaller T and F. Similar explanation can be given to the other parameters.
Figure 9. The PRCC of parameters for sensitivity analysis.
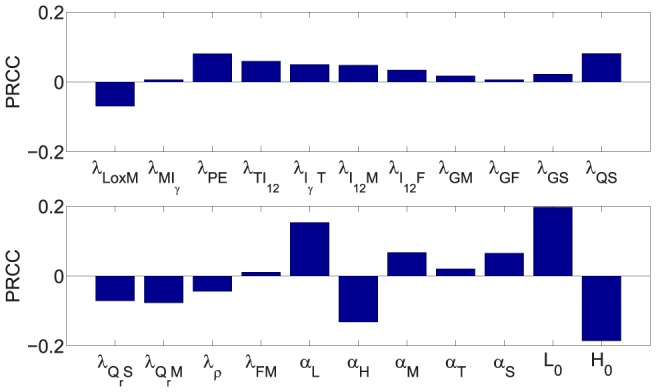
The most significant positively correlated parameters are 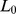 and its influx rate
and its influx rate 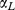 . This is not surprising since LDL initiates the plaque formation. The most significant negatively correlated parameters are
. This is not surprising since LDL initiates the plaque formation. The most significant negatively correlated parameters are 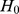 and its influx rate
and its influx rate 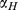 . Indeed, since HDL reduces the availability of free radicals, it plays an important negative role in plaque formation.
. Indeed, since HDL reduces the availability of free radicals, it plays an important negative role in plaque formation.
Discussion
Atherosclerosis is a disease in which a plaque builds up inside an artery. The process of plaque formation begins when, as a result of a lesion in the artery, cholesterols LDL and HDL enter the intima, and LDL becomes oxidized by free radicals. Upon sensing ox-LDL, endothelial cells secrete MCP-1 which attracts monocytes from the blood. As monocytes enter to the intima, they differentiate into macrophages that ingest the ox-LDL and become foam cells. Foam cells attract more macrophages, followed by T cells from the blood, and SMCs from the media. HDL reduces the available free radicals, as well as inflammation within the evolving plaque, thus HDL acts to block plaque growth.
Public health guidelines in the U.S. specify that LDL level of 100–129 mg/dL is near ideal, 130–159 mg/dL is borderline high, and 160–189 mg/dL is very high, whereas concentration of HDL above 60 mg/dL is best, and below 40 mg/dL for men or below 50 mg/dL for women is poor [25]. An important question is how to evaluate the risk of atherosclerosis for a pair of LDL and HDL taken together. This question is addressed in the present paper. We built a mathematical model of plaque development by a system of partial differential equations. The model includes two parameters: 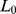 , the level of LDL in the blood, and
, the level of LDL in the blood, and 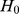 , the the level of HDL in the blood.
, the the level of HDL in the blood.
The model can simulate the evolution of a small plaque for any pair of values of 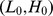 . In Figs. 4, 5, 6, we simulated the plaque evolution over a period of 300 days. For example, one extreme case of
. In Figs. 4, 5, 6, we simulated the plaque evolution over a period of 300 days. For example, one extreme case of 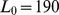 mg/dL,
mg/dL, 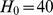 mg/dL, the plaque doubled after 300 days; in another extreme case of
mg/dL, the plaque doubled after 300 days; in another extreme case of 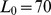 mg/dL,
mg/dL, 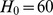 mg/dL, the plaque disappeared after 300 days. We created a risk-map by taking sampling points of LDL and HDL values, and computing the weight of the plaque for each pair
mg/dL, the plaque disappeared after 300 days. We created a risk-map by taking sampling points of LDL and HDL values, and computing the weight of the plaque for each pair 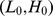 after 100 days. The map shown in Fig. 8, indicates the percentage of plaque growth or shrinkage for any such pair. We accordingly divided the (LDL,HDL) quadrant into three regions: high risk, low risk, and non risk.
after 100 days. The map shown in Fig. 8, indicates the percentage of plaque growth or shrinkage for any such pair. We accordingly divided the (LDL,HDL) quadrant into three regions: high risk, low risk, and non risk.
The need to consider the ratio of LDL/HDL in predicting coronary heart disease was suggested in a case study by [54]. The American Heart Association considers the ratio 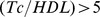 to indicate high risk of heart disease, and the ratio
to indicate high risk of heart disease, and the ratio 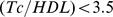 to be risk free [55], where Tc denotes the total cholesterol, which is calculated by the formula
to be risk free [55], where Tc denotes the total cholesterol, which is calculated by the formula
Table 7 shows the National Cholesterol Education Program (NCEP) guidelines associated with plaque buildup [56]. Accordingly, for the five points (a)–(e) in Results we have:
Table 7. National Cholesterol Education Program guidelines.
| LDL Cholesterol Level | Category |
| Less than 100 mg/dL | Optimal |
| 100 to 129 mg/dL | Near or above optimal |
| 130 to 159 mg/dL | Borderline high |
| 160 to 189 mg/dL | High |
| 190 mg/dL and above | Very high |
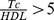 for any value of Tr;
for any value of Tr;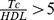 if Tr is above normal;
if Tr is above normal;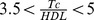 if Tr is not very high;
if Tr is not very high;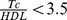 if Tr is normal;
if Tr is normal;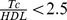 if Tr is normal;.
if Tr is normal;.
According to the NCEP guidelines, (a) and (b) should be in the high risk region; (c) in the low risk region; and (d), (e) in the no risk region, as indeed they are according placed in the risk map in Fig. 8.
Some anti-cholesterol drugs, such as statins, lower LDL and at the same time also increase the HDL [24]. It is important to know which drugs can best achieve the desired risk-free balance between LDL and HDL, that is, bring the individual's (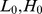 ) into the no risk (or low risk) region. By focusing not on just reducing
) into the no risk (or low risk) region. By focusing not on just reducing 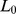 or on just increasing
or on just increasing 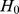 , but on moving the combined (
, but on moving the combined (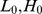 ) to the no risk (or low risk) region in the shortest medically feasible path, we believe one could choose a more personalized medicine from those currently available, which will reduce the risk of atherosclerosis with the lowest amount of doze, thereby also possibly reducing potential negative side effects.
) to the no risk (or low risk) region in the shortest medically feasible path, we believe one could choose a more personalized medicine from those currently available, which will reduce the risk of atherosclerosis with the lowest amount of doze, thereby also possibly reducing potential negative side effects.
To illustrate this approach, we note that for some drugs the ratio of decrease in LDL to increase in HDL is already known. For example, this ratio is 1/3 for the new experimental drug Evacetrapib. Some anti-cholesterol drugs only decrease LDL (e.g. Colestid) while others only increase HDL (e.g. Lofibra). We can the represent effect of such drugs by unit vectors in the 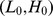 plane: for example, Colestid ←, Lofibra ↑, and Evacetrapid
plane: for example, Colestid ←, Lofibra ↑, and Evacetrapid 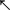 . In Fig. 10. we consider three individuals, A, B, and C in the high risk region. In order to move them to the low risk region with the minimum amount of medication (side effects are ignored), the individual should choose the drug for which the line segment from the individual initial position to the low risk region is the shortest (We assume that the amount of drug is proportional to the length of the line segment). Thus the best drug for A is the one that primarily increases HDL. Similarly, C will do better with a drug that primarily decrease LDL, and B should use a drug with appropriate ratio of decreasing LDL to increasing HDL.
. In Fig. 10. we consider three individuals, A, B, and C in the high risk region. In order to move them to the low risk region with the minimum amount of medication (side effects are ignored), the individual should choose the drug for which the line segment from the individual initial position to the low risk region is the shortest (We assume that the amount of drug is proportional to the length of the line segment). Thus the best drug for A is the one that primarily increases HDL. Similarly, C will do better with a drug that primarily decrease LDL, and B should use a drug with appropriate ratio of decreasing LDL to increasing HDL.
Figure 10. Drug treatment recommended for individuals A, B and C.

Some work has been done on antioxidant therapy for reducing the risk of atherosclerosis, but so far it has had limited success in preventing cardiovascular diseases [57]–[59]. A review of studies in which antioxidant gene therapy has been successfully used is given in [60]. Our model could account for antioxidative medication once we gain a good understanding of how such medication affects the source of free radicals, r 0, in Eq. (4).
Some of the parameters in the differential equations in our model had to be rather crudely estimated, since no data were available, while others may slightly vary depending on the individual. As more data become available, parameter values may be further refined. Our model uses only the values of LDL and HDL as biomarkers. It will be interesting in the future to incorporate also triglycerides into the risk-map. Future work should also explore how other risk factors, such as high blood pressure, smoking and diabetes affect the risk-map.
We did not include in this paper the circulation ox-LDL in the blood, which is elevated only in patients with advanced atherosclerosis [61], [62]. Our model could be extended to include this additional biomarker, but at present there is not enough data on how the level of ox-LDL in the blood correlates to a specific advanced state of the disease.
Supporting Information
Algorithm for finite element implementation of the mathematical model.
(PDF)
Macrophages population for different levels of LDL and HDL.
(PDF)
SMCs population for different levels of LDL and HDL.
(PDF)
T cells population for different levels of LDL and HDL.
(PDF)
Foam cells population for different levels of LDL and HDL.
(PDF)
Concentration of ox-LDL for different levels of LDL and HDL.
(PDF)
Concentration of IFN-γ for different levels of LDL and HDL.
(PDF)
Concentration of IL-12 for different levels of LDL and HDL.
(PDF)
Funding Statement
This research has been supported by the Mathematical Biosciences Institute and the National Science Foundation under grant DMS 0931642 (http://nsf.gov/awardsearch/showAward?AWD_ID=0931642). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Hoyert DL, Xu J (2012) Deaths: Preliminary Data for 2011, National Vital Statistics Reports 61. [PubMed]
- 2.The top 10 causes of death. Available: http://www.who.int/mediacentre/factsheets/fs310/en/. Accessed 2013 Dec. 18.
- 3.Frink RJ (2002) Inflammatory Atherosclerosis, Heart Research Foundation.
- 4. Moreno PR (2010) Vulnerable Plaque: Definition, Diagnosis, and Treatment. Cardiology Clinics 28: 1–30. [DOI] [PubMed] [Google Scholar]
- 5. Libby P, Ridker PM, Maseri A (2002) Inflammation and Atherosclerosis. Circulation 105: 1135–1143. [DOI] [PubMed] [Google Scholar]
- 6. Cohen A, Myerscough MR, Thompson RS (2012) Athero-protective effects of High Density Lipoproteins (HDL): An ODE model of the early stages of atherosclerosis. Preprint [DOI] [PubMed] [Google Scholar]
- 7. McKay C, McKee S, Mottram N, Mulholland T, Wilson S (2005) Towards a model of atherosclerosis. Strathclyde Mathematics Research Report [Google Scholar]
- 8. Ryu BH (2000) Low Density Lipoprotein (LDL), Atherosclerosis and Antioxidants. Biotechnol. Bioprocess Eng 5: 313–319. [Google Scholar]
- 9. Harrington JR (2000) The Role of MCP-1 in Atherosclerosis. Stem Cells 18: 65–66. [DOI] [PubMed] [Google Scholar]
- 10. Reape TJ, Groot P (1999) Chemokines and atherosclerosis. Atherosclerosis 147: 213C225. [DOI] [PubMed] [Google Scholar]
- 11. Osterud B, Bjorklid E (2003) Role of Monocytes in Atherogenesis. Physiol Rev 83: 1069–1112. [DOI] [PubMed] [Google Scholar]
- 12. Gui T, Shimokado A, Sun Y, Akasaka T, Muragaki Y (2012) Diverse Roles ofMacrophages in Atherosclerosis: From Inflammatory Biology to Biomarker Discovery. Mediators of Inflammation 2012: ID693083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Johnson JL, Newby AC (2009) Macrophage heterogeneity in atherosclerotic plaques. Curr Opin Lipidol 20: 370–378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Little MP, Gola A, Tzoulaki I (2009) A Model of Cardiovascular Disease Giving a Plausible Mechanism for the Effect of Fractionated Low-Dose Ionizing Radiation Exposure. PLoS Computational Biology 5: e1000539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Calvez V, Ebde A, Meunier N, Raoult A (2009) Mathematical modelling of the atherosclerotic plaque formation. CEMRACS 2008 - Modelling and Numerical Simulation of Complex Fluids 28: 1–12. [Google Scholar]
- 16. Ross R, Masuda J, Raines EW, Gown AM, Katsuda S, et al. (1990) Localization of PDGF-B protein in macrophages in all phases of atherogenesis. Science 248: 1009–1012. [DOI] [PubMed] [Google Scholar]
- 17. Raines EW, Ross R (1993) Smooth muscle cells and the pathogenesis of the lesions of atherosclerosis. Br Heart J 69: 30–37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Rodriguez JA, Orbe J, Paramo JA (2007) Metalloproteases, Vascular Remodeling, and Atherothrombotic Syndromes. Rev Esp Cardiol 60: 959–967. [DOI] [PubMed] [Google Scholar]
- 19. Fabunmi RP, Sukhova GK, Sugiyama S, Libby P (1998) Expression of Tissue Inhibitor of Metalloproteinases-3 in Human Atheroma and Regulation in Lesion-Associated Cells A Potential Protective Mechanism in Plaque Stability. Circ Res 83: 270–278. [DOI] [PubMed] [Google Scholar]
- 20. King IL, Segal BM (2005) Cutting edge: IL-12 induces CD4+CD25- T cell activation in the presence of T regulatory cells. J Immunol 175: 641–645. [DOI] [PubMed] [Google Scholar]
- 21. Hansson GK, Holm J, Jonasson L (1989) Detection of activated T lymphocytes in the human atherosclerotic plaque, Am. J Pathol 135: 169–175. [PMC free article] [PubMed] [Google Scholar]
- 22. Kosaka C, Masuda J, Shimokado K, Zen K, Yokota T, et al. (1992) Interferon-gamma suppresses PDGF production from THP-1 cells and blood monocyte-derived macrophages. Atherosclerosis 97: 75–87. [DOI] [PubMed] [Google Scholar]
- 23.Barter P (2005) The role of HDL-cholesterol in preventing atherosclerotic disease, European Heart Journal Supplements. 7: : 4–8. [Google Scholar]
- 24. Barter P, Brandrup-Wognsen G, Palmer M, Nicholls S (2010) Effect of statins on HDL-C: a complex process unrelated to changes in LDL-C: analysis of the VOYAGER Database. J Lipid Res 51: 1546–1553. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Mayo Clinic Staff, Cholesterol levels: What numbers should you aim for? Available: http://www.mayoclinic.com/health/cholesterol-levels/CL00001. Accessed 2013 Dec. 12.
- 26. Rahdert DA, Sweet WL, Tio FO, Janicki C, Duggan DM (1999) Measurement of density and calcium in human atherosclerotic plaque and implications for arterial brachytherapy. Cardiovasc Radiat Med 1: 358–367. [DOI] [PubMed] [Google Scholar]
- 27. Orbe J, Rodriguez JA, Arias R (2003) Antioxidant vitamins increase the collagen content and reduce MMP-1 in a porcine model of atherosclerosis: Implications for plaque stabilization. Atherosclerosis 167: 45–53. [DOI] [PubMed] [Google Scholar]
- 28. Kim Y, Roh S, Lawler S, Friedman A (2011) miR and AMPK mutual antagonism in glioma cell migration and proliferation: a mathematical model. PLoS One 6: e28293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Byrne HM (1997) The importance of intercellular adhesion in the development of carcinomas. IMA Journal of Mathematics Applied in Medicine & Biology 14: 305–323. [PubMed] [Google Scholar]
- 30. Byrne HM (1999) A weakly nonlinear analysis of a model of avascular solid tumour growth. J Math Biol 39: 59–89. [DOI] [PubMed] [Google Scholar]
- 31. Byrne HM, Chaplain MA (1996) Modelling the role of cell-cell adhesion in the growth and development of carcinomas. Mathematical and Computer Modelling 24: 1–7. [Google Scholar]
- 32. Chothia C (1976) The nature of the accessible and buried surfaces in proteins. J Mol Biol 105: 1–12. [DOI] [PubMed] [Google Scholar]
- 33. Janin J, Chothia C (1978) Role of Hydrophobicity in the Binding of Coenzymes. Biochemistry 17: 2943–2948. [DOI] [PubMed] [Google Scholar]
- 34. Cobbold C, Sherratt J, Maxwell S (2002) Lipoprotein oxidation and its significance for atherosclerosis: a mathematical approach. Bulletin of Mathematical Biology 64: 65–95. [DOI] [PubMed] [Google Scholar]
- 35. Ingold KU, Bowry VW, Stocker R, Walling C (1993) Autoxidation of lipids and antioxidation by a-tocopherol and ubiquinol in homogeneous solution and in aqueous dispersions of lipids: unrecognised consequences of lipid particle size as exemplified by oxidation of human low density lipoprotein. Proc Natl Acad Sci 90: 45–49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Kim Y, Lawler S, Nowicki M, Chiocca E, Friedman A (2009) A mathematical model for pattern formation of glioma cells outside the tumor spheroid core. J Theor Biol 260: 359–371. [DOI] [PubMed] [Google Scholar]
- 37. Kim Y, Friedman A (2010) Interaction of tumor with its micro-environment: A mathematical model. Bull Math Biol 75: 1029–1068. [DOI] [PubMed] [Google Scholar]
- 38. Mercapide J, Cicco R, Castresana J, Klein-Szanto A (2003) 450 Stromelysin-1/matrix metalloproteinase-3 (MMP-3) expression accounts for invasive properties of human astrocytoma cell lines. Int J Cancer 106: 676–682. [DOI] [PubMed] [Google Scholar]
- 39. Friedman A, Turner J, Szomolay B (2008) A model on the influence of age on immunity to infection with Mycobacterium tuberculosis. Exp Gerontol 43: 275–285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Orme IM (1987) Aging and immunity to tuberculosis: increased susceptibility of old mice reflects a decreased capacity to generate mediator T lymphocytes. J Immunol 138: 4414–4418. [PubMed] [Google Scholar]
- 41. Barrett T, Benditt E (1988) Platelet-derived growth factor gene expression in human atherosclerotic plaques and normal artery wall. Proc Natl Acad Sci 85: 2810–2814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Xue C, Friedman A, Sen CK (2009) A mathematical model of ischemic cutaneous wounds. Proc Natl Acad Sci 106: 16782–16787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Dollery CM, Libby P (2006) Atherosclerosis and proteinase activation. Cardiovascular Research 69: 625–635. [DOI] [PubMed] [Google Scholar]
- 44. Chen D, Roda JM, Mash CB, Eubank TD, Friedman A (2012) Hypoxia inducible factors mediated-inhibition of cancer by GM-CSF: A mathematical model. Bulletin of Mathematical Biology 74: 2752–2777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Day J, Friedman A, Schlesinger LS (2009) Modeling the immune rheostat of macrophages in the lung in response to infection. Proc Natl Acad Sci 106: 11246–11251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Yamamoto S, Nguyen JH (2006) TIMP-1/MMP-9 imbalance in brain edema in rats with fulminant hepatic failure. J Surg Res 134: 307–314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Olson MW, Gervasi DC, Mobashery S, Fridman R (1997) Kinetic analysis of the binding of human matrix metalloproteinase-2 and -9 to tissue inhibitor of metalloproteinase (TIMP)-1 and TIMP-2. J Biol Chem 21: 29975–29983. [DOI] [PubMed] [Google Scholar]
- 48. Palosaari H, Yliopisto O (2003) Matrix Metalloproteinases (MMPs) and Their Specific Tissue Inhibitors (TIMPs) in Mature Human Odontoblasts and Pulp Tissue. Oulun yliopisto [DOI] [PubMed] [Google Scholar]
- 49. Bofill M, Janossy G, Lee CA, MacDonald-Burns D, Phillips AN, et al. (1992) Laboratory 475 control values for CD4 and CD8 T lymphocytes Implications for HIV-1 diagnosis. Clin Exp Immunol 88: 243–252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Persson PO, Strang G (2004) A Simple Mesh Generator in MATLAB. SIAM Review 46: 329–345. [Google Scholar]
- 51. Alberty J, Carstensen C, Funken SA (1999) Remarks around 50 lines of Matlab: short finite element implementation. Numerical Algorithms 20: 117–137. [Google Scholar]
- 52.Z Li. The finite element method for two dimensional problems. Chapter 9 Avialable http://www4.ncsu.edu/~zhilin/TEACHING/MA587/chap9.pdf. Accessed 2013 Oct. 10.
- 53. Marino S, Hogue IB, Ray CJ, Kirschner DE (2008) A methodology for performing global uncertainty and sensitivity analysisin systems. J Theor Biol 254: 178–196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Arsenault BJ, Rana JS, Stroes ES, Despres JP, Shah PK, et al. (2009) Beyond low-density lipoprotein cholesterol: respective contributions of non-high-density lipoprotein cholesterol levels, triglycerides, and the total cholesterol/high-density lipoprotein cholesterol ratio to coronary heart disease risk in apparently healthy men and women. J Am Coll Cardiol 55: 35–41. [DOI] [PubMed] [Google Scholar]
- 55.Mayo Clinic Staff, How important is cholesterol ratio? Available: http://www.mayoclinic.org/diseases-conditions/high-blood-cholesterol/expert-answers/cholesterol-ratio/faq-20058006. Accessed 2013 Dec. 20.
- 56.Crestor Managing Choleserol. Available: http://www.crestor.com/c/your-arteries/tools-resources/index.aspx.
- 57. Otani H (2013) Site-Specific Antioxidative Therapy for Prevention of Atherosclerosis and Cardiovascular Disease. Oxid Med Cell Longev [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Ozkanlar S, Akcay F (2012) Antioxidant vitamins in atherosclerosis–animal experiments and clinical studies. Adv Clin Exp Med 21: 115–123. [PubMed] [Google Scholar]
- 59. Tardif J (2005) Antioxidants and atherosclerosis: emerging drug therapies. Curr Atheroscler Rep 7: 71–77. [DOI] [PubMed] [Google Scholar]
- 60. Levonen A, Vahakangas E, Koponen JK, Herttuala S (2008) Antioxidant Gene Therapy for Cardiovascular Disease Current Status and Future Perspectives. Circulation 117: 2142–2150. [DOI] [PubMed] [Google Scholar]
- 61. Ishigaki Y, Oka Y, Katagiri H (2009) Circulating oxidized LDL: a biomarker and 501 a pathogenic factor. Curr Opin Lipidol 20: 363–369. [DOI] [PubMed] [Google Scholar]
- 62. Tsimikas S, Witztum JL (2001) Measuring circulating oxidized low-density lipoprotein to evaluate coronary risk. Circulation 103: 1930–1932. [DOI] [PubMed] [Google Scholar]
- 63. Rajman I, Eacho P, Chowienczyk P, Ritter J (1999) LDL particle size: an important drug target? Br J Clin Pharmacol 48: 125–133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Hokland B, Mendez A, Oram J (1992) Cellular localization and characterization of proteins that bind high density lipoprotein. J Lipid Res 33: 1335–1342. [PubMed] [Google Scholar]
- 65. Mitsuhashi T, Ono K, Fukuda M, Hasegawa Y (2013) Free radical scavenging ability and structure of a 90-kDa protein from the scallop shell. Fisheries Science 79: 495–502. [Google Scholar]
- 66. Garcia-Tunon I, Ricote M, Ruiz A, Fraile B, Paniagua R, et al. (2007) Influence of IFN-gamma and its receptors in human breast cancer. BMC Cancer 7: 158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Antoniades H (1981) Human platelet-derived growth factor (PDGF): purification of PDGF-I and PDGF-II and separation of their reduced subunits. Proc Natl Acad Sci 78: 7314–7317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Yokochi S, Hashimoto H, Ishiwata Y, Shimokawa H, Haino M, et al. (2001) An Anti-Inflammatory Drug, Propagermanium, May Target GPI-Anchored Proteins Associated with an MCP-1 Receptor, CCR2. Journal of Interferon and Cytokine Research 21: 389–398. [DOI] [PubMed] [Google Scholar]
- 69. Hamza T, Barnett J, Li B (2010) Interleukin 12 a Key Immunoregulatory Cytokine in Infection Applications. Int J Mol Sci 11: 789–806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. O'Keefe JH, Cordain L, Harris WH, Moe RM, Vogel R (2004) Optimal low-density lipoprotein is 50 to 70 mg/dl: lower is better and physiologically normal. Journal of the American College of Cardiology 43: 2142–2146. [DOI] [PubMed] [Google Scholar]
- 71. Miller DC, Thapa A, Haberstroh KM, Webster TJ (2004) Endothelial and vascular smooth muscle cell function on poly(lactic-co-glycolic acid) with nano-structured surface features. Biomaterials 25: 53–61. [DOI] [PubMed] [Google Scholar]
- 72. Bowen-Pope DF, Malpass TW, Foster DM, Ross R (1984) Platelet-derived growth factor in 526 vivo: levels, activity, and rate of clearance. Blood 64: 458–469. [PubMed] [Google Scholar]
- 73. Rhodes J, Sharkey J, Andrews P (2009) Serum IL-8 and MCP-1 concentration do not identify patients with enlarging contusions after traumatic brain injury. J Trauma 66: 1591–1597. [DOI] [PubMed] [Google Scholar]
- 74. Gattorno M, Picco P, Vignola S, Stalla F, Buoncompagni A, et al. (1998) Serum interleukin 12 concentration in juvenile chronic arthritis. Ann Rheum Dis 57: 425–428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Tietz NW (1999) Clinical Guide to Laboratory Tests. Philadelphia 3rd Ed.
- 76. Liao KL, Bai XF, Friedman A (2013) The role of CD200-CD200R in tumor immune evasion. J Theor Biol 328: 65–76. [DOI] [PubMed] [Google Scholar]
- 77. Tsukaguchi K, Balaji KN, Boom WH (1995) CD4+ alpha beta T cell and gamma delta T cell responses to Mycobacterium tuberculosis. J Immunol 154: 1786–1796. [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Algorithm for finite element implementation of the mathematical model.
(PDF)
Macrophages population for different levels of LDL and HDL.
(PDF)
SMCs population for different levels of LDL and HDL.
(PDF)
T cells population for different levels of LDL and HDL.
(PDF)
Foam cells population for different levels of LDL and HDL.
(PDF)
Concentration of ox-LDL for different levels of LDL and HDL.
(PDF)
Concentration of IFN-γ for different levels of LDL and HDL.
(PDF)
Concentration of IL-12 for different levels of LDL and HDL.
(PDF)