

Abstract
Anoxia and apoptosis are both implicated in chronic tendon pathology, however the influence of anoxia on the viability of tendon cells is not known. The objectives of the current study were to (i) investigate the effect of oxygen withdrawal on the viability of porcine Achilles tendon cells (ATCs), and (ii) examine the ability of IGF-I, a factor with known regenerative properties in tendon, to prevent ATC death. Cultured ATCs were enclosed in an anaerobic chamber. The mechanism of cell death was examined by flow cytometry of ATCs double labeled with Annexin-V and propidium iodide (PI). Caspase activity was determined by a fluorometric assay, and nuclear morphology was examined by Hoechst staining. The cell death induced by anoxia was time-dependent, and was characterized by phosphatidylserine exposure on the outer membrane, caspase activation and DNA fragmentation. Death was inhibited by the addition of IGF-I in a dose-dependent manner. The ability of IGF-I to activate the pro-survival PKB pathway in ATCs was inhibited by LY294002, indicating the importance of PI3K in the response of ATCs to IGF-I. These data suggest that cell death induced by lack of oxygen is predominantly apoptotic and can be prevented by pro-survival IGF-I signaling. This mechanism may contribute to the beneficial effect of IGF-I on tendon.
Keywords: Achilles tendon, anoxia, IGF-I, apoptosis
Introduction
Achilles tendon injury is an important clinical problem in both athletes and non-athletes (24, 27). The histopathology of ruptured tendons shows evidence of pre-existing pathology that predisposes to rupture (35). Of various patterns of pathology including hypoxic, mucoid, fatty, and calcific tendinopathies, hypoxic tendinopathy was the most common in a series of 891 tendon ruptures (23). Although the histology of hypoxic tendinopathy is well described (including regions of hypocellularity, pyknotic nuclei, necrosis, mitochondrial abnormalities, and collagen thinning and disarray) (23),the underlying cellular processes are poorly understood.
Apoptosis, or programmed cell death, is associated with chronic tendon pathology (46, 47). In patients with supraspinatus rupture and with tennis elbow, apoptosis has been reported based on TUNEL staining, DNA laddering and transmission electron microscopy (20, 47). Other features of apoptosis that distinguish it from classic cell death (necrosis) include the activation of pro-death proteases – caspases – and the exposure of phosphatidylserine on the outer plasma membrane that serves as a signal for phagocytosis by macrophages (4, 18, 25). Although apoptosis plays important homeostatic and physiologic roles (for example during regression of embryonic structures) it also plays negative roles in degenerative conditions of bone, cartilage, muscle and nerve. In general, the cell’s decision to survive or to commit to apoptosis appears to result from a balance of pro-survival and death-inducing signals (8).
The events leading to tendon cell death or survival are beginning to be explored (46, 47) but the influences of hypoxia and growth factors on tendon cell viability have not been examined. In other cell types, hypoxia and anoxia induce necrosis, or apoptosis via transcription pathways involving p53 and p38 (13, 40). Opposing such damage following injury is IGF-I a prosurvival factor released by platelets, macrophages, and fibroblasts (19, 37–39). By binding to the IGF-I receptor, IGF-I enacts a number of pro-survival signaling events, including activation of protein kinase B (PKB) which prevents cell death by phosphorylating various cytoplasmic and nuclear targets (44). The effects of IGF-I on tendon cells are of special interest given its promise as a factor that can accelerate biochemical and biomechanical recovery after tendon injury (1–3, 9, 14, 26, 29).
Given that reduced oxygen may play a role in acute or chronic Achilles tendinopathies (12, 21, 33) and that IGF-I has documented anabolic and regenerative effects in tendon (1–3, 9, 14, 26) we evaluated the influence of anoxia and IGF-I on death and survival signal pathways in Achilles tendon cells. The specific objectives were to determine (i) the extent and type of cell death induced by anoxia, and (ii) the ability of IGF-I to inhibit anoxia-induced cell death and to activate the PI3K/PKB pathway.
Materials and Methods
Reagents
Type I collagenase (clostridiopeptidase A), IGF-I, and avidin-flourescein isothiocyanate (FITC) were purchased from Sigma (St Louis, MO). Heat-inactivated fetal bovine serum (FBS) was purchased from Invitrogen (Carlsbad, CA). Annexin-V conjugated to biotin was obtained from Pharmingen (San Diego, CA). Anaerobic chambers, pouches and indicator strips were supplied by BD (Gaspak, Franklin Lakes, NJ). A primary rabbit polyclonal antibody detecting the phosphorylated form of PKB was from Stressgen (Victoria, Canada). P85 antibody was obtained from Cell Signaling Technologies (Beverly, MA).
Achilles tendon cell culture
Porcine Achilles tendon tissue, harvested aseptically from animals being euthanized for university teaching purposes, was placed immediately into sterile PBS. Adherent muscle and peritendinous tissue was removed. The sample was then washed repeatedly in sterile PBS, cut into 0.5 mm pieces with a sterile blade and forceps and digested for 24–48 hours in DMEM containing 0.5mg/ml collagenase, 10% FBS, (v/v) 20 mM Hepes, pH 7.4, 2% (v/v) penicillin-streptomycin, and 0.1% (v/v) gentamycin on an agitator at 37°C. The tendon fibroblasts released by this procedure were plated, along with any undigested collagenous tissue, on plasma-treated polystyrene tissue culture dishes. Tendon cell cultures were maintained in DMEM with 10% (v/v) FBS in a 5% CO2 incubator at 37°C. Cells were used in experiments up to the fifth passage.
To examine the effects of anoxia, tendon cells were seeded at 500,000 cells per 60mm plate and allowed to adhere for 48–72 hours, at which point the FBS concentration was reduced to 1%; cells were then maintained as usual in the 5% CO2 incubator (normoxia) or placed at 37°C in an anaerobic chamber (GasPak system) for 24–96 hours (anoxia). Anoxia was confirmed by using methylene-blue indicator strips which become colorless when oxygen is absent. All experiments were performed in triplicate.
Cell viability assay
After indicated timepoints of normoxia or anoxia, cells were harvested by trypsinization and scraping, washed in PBS, fixed for 1h in ice-cold 70% ethanol, then placed for 30 minutes in 50μg/ml propidium iodide (PI) in PBS with 0.1% glucose and 100μg/ml RNaseA. A histogram representing DNA content was generated from each sample using the FL3 channel on a flow cytometer (Coulter Epics XL-MCL); the percentage of apoptotic cells (with sub-diploid DNA) was quantified using the same gate for all samples.
Apoptosis assay
To analyze the mechanism of tendon cell death, normoxic and hypoxic (anoxic) cells were harvested and washed as above, then resuspended in 500 μl of 10mM Hepes, 140 mM NaCl, 2.5 mM CaCl2, pH 7.4 to which 5μl of biotinylated Annexin V was added. After 15 minutes incubation in the dark, 5μl of Avidin-FITC (50μg/ml) and 5μl of PI (10mg/ml) were added. After 30 minutes, the samples were analyzed on a flow cytometer simultaneously on the FL1 and FL3 channels after appropriate compensation with single-stained and unstained tendon cells. Each channel was gated, yielding a scatter plot with four quadrants representing cells that were viable, in early or late apoptosis, or necrosis (46).
Caspase activation assay
Caspase activation was examined in live and dying cells using a flourometric assay kit according to the manufacturer’s instructions (Immunochemistry). Briefly, a fluorescent probe (FAM-VAD-FMK FLICA) was added to anoxic or normoxic tendon cells harvested as above. The probe diffuses freely across the plasma membrane and reacts broadly with active enzymes of the caspase family by binding covalently to a reactive cysteine residue on the active caspase (43). After several washes in PBS, the cells were analyzed by flow cytometry and on a fluorescent microscope (Zeiss Axioplan) after a 10 minute incubation with Hoechst 33342, a nuclear dye which is more readily taken up by apoptotic cells (10).
Western blot analysis for PKB activation
Tendon cells were harvested and 500,000 cells were allowed to adhere in 60mm plates overnight. To render cells quiescent prior to IGF-I stimulation, serum was withdrawn for 12 hours, then IGF-I was added to the medium for varying concentrations and durations. Cells were released from the plate by scraping, centrifuged and lysed in ice-cold solubilization buffer containing 50mM Tris-HCL, 1% Triton X-100, 10% glycerol, 100mM NaCl, 25mM EDTA, 10mM NaF, 1 mM sodium vanadate, 1 mM dithiothreitol, 1 μM phenylmethylsulfonyl fluoride, 10 μg/ml aprotinin, and 10 μg/ml leupeptin. Lysates were sonicated on ice for 10 seconds, centrifuged at 14,000 rpm for 10 min, boiled in SDS and beta-mercaptoethanol, then loaded onto a 9% SDS-PAGE separating gel. Gels were calibrated using prestained SDS-PAGE low molecular weight standards. Proteins were transferred electrophoretically to nitrocellulose membranes, blocked for 1 h in Tris-buffered saline (TBS) containing 3% skim milk, then incubated overnight with the primary anti-PKB antibody at room temperature. After three washes with TBS and 0.1% Tween 20, membranes were incubated with horseradish peroxidase-conjugated secondary antibody at 1:5000 dilution for 1 h. Bands were visualized by using enhanced chemiluminescence. Equal loading was confirmed by reprobing the membranes with antibodies recognizing the p85 subunit of PI 3-kinase.
Statistical analysis
All values in the text and figures are expressed as mean ± S.D. of 3 observations. Statistical comparison between experimental group and control was performed using a repeat measures ANOVA. The confidence limit was predetermined at an α level of 0.05.
Results
Oxygen withdrawal induces apoptosis of Achilles tendon cells
Lack of oxygen resulted in a progressive increase in the rate of cell death. However, the rate of cell death did not reach substantial levels until after 48 hours (Figure 1). Morphologically, the majority of dying cells demonstrated features of apoptosis (shrunken cytoplasm, membrane blebbing) rather than necrosis (swollen cytoplasm, membrane disruption). In cells kept under anoxic conditions but in the presence of added IGF-1, there was no evidence of an increase in cell death over the time period examined (up to 96 hours). Flow cytometry of cells double labeled with Annexin-V and PI confirmed that the predominant type of cell death induced by anoxia was apoptosis (Figure 2A&B). Among the dead cells (PI positive), the majority were Annexin positive (apoptotic), with few being Annexin negative (necrotic).
Figure 1.
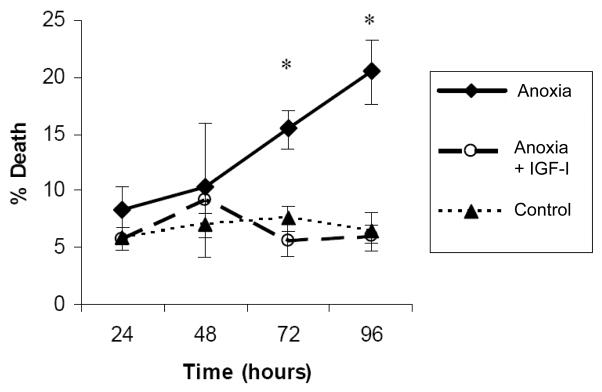
Pro-survival effect of IGF-I under anoxic conditions. Cells were maintained under normoxic or anoxic conditions for the indicated time periods and cell death was indicated by the number of cells having sub-diploid DNA content. Incubation with IGF-I restored the level of tendon cell death to control levels at all time periods examined. * p < 0.01.
Figure 2.

Anoxia-induced tendon cell death is predominantly apoptotic. Cells were kept at normoxia (A, C, E) or anoxia (B, D, F) for 72 hours. A & B: Cells were trypsinized and double-labeled with PI and Annexin V. The bottom left quadrant represents viable cells that are negative for both probes. The top two quadrants represent early (left) and late (right) apoptotic cells, whereas cells at the bottom right are necrotic (PI+, Annexin−). C & D: Tenocytes were incubated with a fluorescent probe recognizing active caspases. In each panel, the largest peak represents background fluorescence from unstained cells. The right-most peak in the anoxic tencoytes (D) is due to positive labeling of tendon cells containing active caspases. E & F: Tendon cells with active caspases (green) were common in the 72 hour anoxic cultures. These same cells frequently demonstrated increased uptake of Hoechst 33342 and fragmented nuclei, indicating that they were undergoing apoptosis.
Caspase activation in anoxic tendon cells
As some authors refer to a “caspase-independent” form of programmed cell death (“aponecrosis”)(4), we undertook experiments to confirm that cell death in these experiments was accompanied by caspase activation. Exposure of tendon cells to anoxia increased caspase activity 3- to 4-fold compared to cells treated identically but kept at normal oxygen tension, as evidenced by flow cytometry (Figure 2C&D). The finding of caspase activation was further validated by labeling anoxic tendon cells (72 hours) simultaneously with the same fluorometric caspase probe and with Hoechst 33342. Tendon cells that demonstrated green fluorescence in the cytoplasm, indicating active caspases, frequently showed increased uptake of Hoechst and fragmented nuclear morphology (Figure 2F).
IGF-I prevents anoxic cell death
ATCs in which IGF-I was added at the same time that oxygen was withdrawn demonstrated levels of cell death equivalent to control values up to 96 hours. The dose-response effect of IGF-I on cell survival was tested using concentrations of IGF-1 up to 500ng/ml (Figure 3). A maximal survival effect was observed at a concentration of 250 ng/ml.
Figure 3.
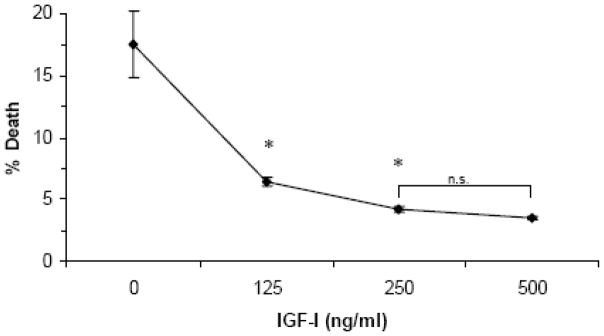
Dose-response of IGF-I protection from apoptosis during anoxia (72 hours). * indicates significant difference from controls and from each other, p < 0.001. N.S; not significantly different from one another.
IGF-I activates PKB through a PI3K dependent manner
To investigate one of the likely mechanisms of anti-apoptotic signaling by IGF-I in tendon cells, a phospho-specific antibody was employed that recognizes PKB when phosphorylated at the Ser473 activation site. Immunoblotting revealed a rapid increase in phosphorylation of PKB, which was elevated by 2′, peaked at 5 minutes and had nearly disappeared by 20 minutes (Figure 4). The phosphorylated form of PKB was not detectable in cells that were pre-incubated with LY294002, a known PI3K inhibitor. LY294002 (25μm) also caused substantial cell death when added to normoxic, quiescent cell cultures indicating that ongoing survival of ATCs is dependent on the PI3K – PKB pathway (data not shown).
Figure 4.
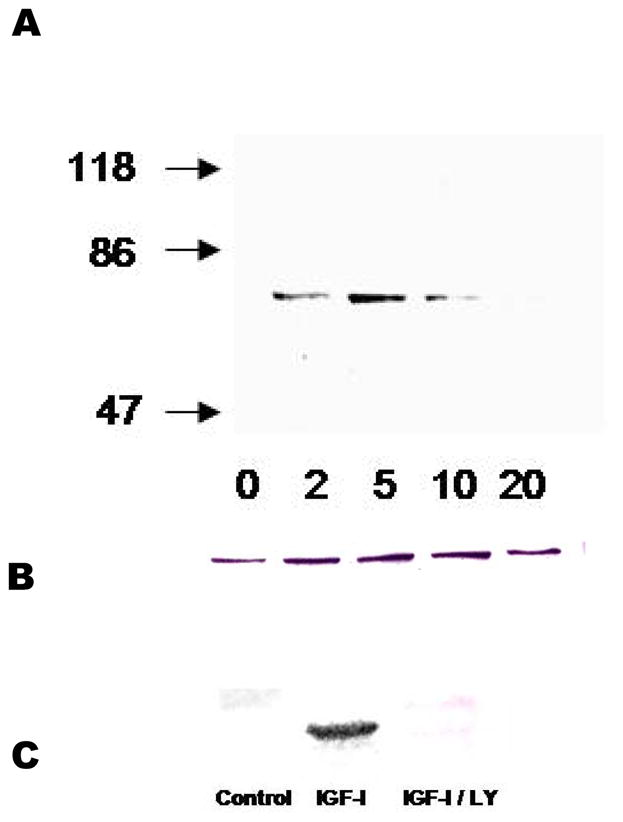
Activation of PKB by IGF-I. A: Phosphorylation at Ser473 showed a rapid onset and decay. B: The same membrane probed with P85 demonstrating equal loading. C: Inhibitory effect of LY (25μm) on PKB activation.
Discussion
In response to a lack of oxygen, Achilles tendon cells undergo either apoptosis or necrosis, with apoptosis representing the predominant mode of death. Apoptosis has not previously been well characterized in tendon cells. This study showed that it occurs in a manner that is characteristic of apoptosis in other cell types that includes exposure of phosphatidylserine on the outer plasma membrane, caspase activation, and nuclear fragmentation (4).
We examined the effects of complete anoxia on tendon cells rather than gradations of PO2, in order to induce a large effect that would allow us to determine whether oxygen withdrawal would induce apoptosis. In young rats, the mean PO2 of tendons was 78.6 mmHg (34), but the PO2 of aged, injured, or hypovascular regions of Achilles and other tendons are not known. Because oxygen supply is known to be a key determinant of a successful healing response (42), future studies to determine actual PO2 in regions of tendinosis before or after rupture are required. Further experiments should also examine the effects of different hypoxic conditions with gradations of PO2.
Although tendon cells express a functional aerobic metabolism (11), they are relatively insensitive to oxygen withdrawal compared to other cell types. Tendon explants maintained in 0% O2 in 10% FBS for 48 hours demonstrated no decline in cell proliferation or proteoglycan production, but collagen production was inhibited; this was in contrast to synovial explants, in which proliferation was significantly reduced, along with proteoglycan and collagen production (31). Likewise, cultured human embryonic Achilles tendon cells whose oxygen was reduced from 20% to 2.7% did not experience a reduction in proliferation, whereas fibroblasts derived from other tissues did (45). Furthermore, the magnitude of cell death we report here for tendon cells is several times lower than that reported in similar experiments with various cell types (7, 17, 30, 40). The relative insensitivity of tendon fibroblasts to hypoxia raises the question of why the relatively low percentage of cells undergoing apoptosis at 72 hours (15–20%) were less resistant than the cells which survived for this duration. From other cell types, hypoxia has been found to irreversibly damage some cells, while surviving cells from the same initial population can exhibit adaptive changes which increase their resistance to hypoxia and other stressful agents (16). Such investigations have yet to be carried out in tendon cells, and may be relevant in understanding their resistance to cell death as shown in the current study.
Chronic Achilles tendon pathology is associated with an angiogenic response (6). Animal studies indicate that angiogenesis represents a late feature of overuse pathology, with neovessels taking several weeks or months to form and penetrate the area of pathology (28). The increased lactate level in Achilles tendinosis (5) suggests that angiogenesis may lag behind oxygen requirements. It is tempting to speculate that compromised blood supply leading to a gradation of hypoxia (with the most severe levels at the center of a tendon lesion) could account for the histological appearance of a central zone of cell death bordered by a peripheral zone of proliferation (45). This pattern is also seen in tumours, which display a central zone of hypoxia, despite an active angiogenic response. Hypoxia could lead not only to cell death, but could also influence transcription in tendon cells resulting in matrix abnormalities such as fibrocartilaginous change, calcification and bony metaplasia (15). These speculations must be tested empirically as the actual conditions within injured tendons are likely to be very complex, with numerous interacting conditions and factors (including mechanical loading, pH, and soluble or matrix-bound substances) determining the outcome.
Chronic hypoxia, one of many stresses that can trigger apoptosis, appears to operate through a pathway that is distinct from most stress-inducing agents (32). Under acute anoxic conditions, the level of hypoxia-inducible factor (HIF-1α) - the “cellular sensor” of oxygen levels - rises, and activates transcription of genes involved in the response to hypoxia, including those encoding TGF-beta and VEGF. If oxygen is chronically low, HIF-1α binds and stabilizes p53, a tumor-suppressor transcription factor capable of inducing cell-cycle arrest or apoptosis. P53 is known to be elevated in experimentally ruptured tendon (22). Hypoxia may also induce apoptosis through other pathways; for example, one report suggested that hypoxia-induced apoptosis requires the development of acidosis (36), but in our experiments apoptosis occurred with no change in the pH of the medium. In vivo, tendons undergo mechanical strain, and static tension held over several minutes reduces tendon oxygen levels (41). It is unlikely that tension-induced hypoxia would induce apoptosis in tendons, as periods of complete hypoxia (anoxia) >24 hours were required to induce substantial amounts of apoptosis in the current study.
In summary, our studies demonstrate that IGF-I activated the PKB pathway in tendon cells in a PI3-kinase dependent manner, and that IGF-I prevented anoxic cell death of ATCs. These findings extend studies conducted using other cell types, in which IGF-I exerts a protective effect from various cellular stresses. Whether other growth factors exert similar pro-survival effects in tendon cells is not yet known. We conclude that future studies should investigate the relative importance of downstream IGF-I signaling, and develop improved methods of achieving prolonged IGF-I delivery to areas of tendon pathology.
Acknowledgments
This work was funded by the Worker’s Compensation Board of B.C. and the B.C. Sports Medicine Foundation. A Scott was supported by the National Science and Engineering Research Council, the Babicki Fellowship in Medical Research and the Michael Smith Foundation for Health Research. V Duronio is a senior scholar of the Michael Smith Foundation for Health Research. K Khan was supported by the Canadian Institutes of Health Research New Investigator programme.
References
- 1.Abrahamsson SO, Lohmander S. Differential effects of insulin-like growth factor-I on matrix and DNA synthesis in various regions and types of rabbit tendons. J Orthop Res. 1996;14:370–376. doi: 10.1002/jor.1100140305. [DOI] [PubMed] [Google Scholar]
- 2.Abrahamsson SO, Lundborg G, Lohmander LS. Long-term explant culture of rabbit flexor tendon: effects of recombinant human insulin-like growth factor-I and serum on matrix metabolism. J Orthop Res. 1991;9:503–515. doi: 10.1002/jor.1100090406. [DOI] [PubMed] [Google Scholar]
- 3.Abrahamsson SO, Lundborg G, Lohmander LS. Recombinant human insulin-like growth factor-I stimulates in vitro matrix synthesis and cell proliferation in rabbit flexor tendon. J Orthop Res. 1991;9:495–502. doi: 10.1002/jor.1100090405. [DOI] [PubMed] [Google Scholar]
- 4.Aigner T, Kim HA. Apoptosis and cellular vitality: issues in osteoarthritic cartilage degeneration. Arthritis Rheum. 2002 Aug;46:1986–1996. doi: 10.1002/art.10554. [DOI] [PubMed] [Google Scholar]
- 5.Alfredson H, Bjur D, Thorsen K, Lorentzon R, Sandstrom P. High intratendinous lactate levels in painful chronic Achilles tendinosis. An investigation using microdialysis technique. J Orthop Res. 2002 Sep;20:934–938. doi: 10.1016/S0736-0266(02)00021-9. [DOI] [PubMed] [Google Scholar]
- 6.Alfredson H, Ohberg L, Forsgren S. Is vasculo-neural ingrowth the cause of pain in chronic Achilles tendinosis? An investigation using ultrasonography and colour Doppler, immunohistochemistry, and diagnostic injections. Knee Surg Sports Traumatol Arthrosc. 2003 Sep;11:334–338. doi: 10.1007/s00167-003-0391-6. [DOI] [PubMed] [Google Scholar]
- 7.Araya R, Uehara T, Nomura Y. Hypoxia induces apoptosis in human neuroblastoma SK-N-MC cells by caspase activation accompanying cytochrome c release from mitochondria. FEBS Lett. 1998 Nov 13;439:168–172. doi: 10.1016/s0014-5793(98)01363-5. [DOI] [PubMed] [Google Scholar]
- 8.Banasiak KJ, Xia Y, Haddad GG. Mechanisms underlying hypoxia-induced neuronal apoptosis. Prog Neurobiol. 2000 Oct;62:215–249. doi: 10.1016/s0301-0082(00)00011-3. [DOI] [PubMed] [Google Scholar]
- 9.Banes AJ, Tsuzaki M, Hu P, Brigman B, Brown T, Almekinders L, et al. PDGF-BB, IGF-I and mechanical load stimulate DNA synthesis in avian tendon fibroblasts in vitro. J Biomech. 1995;28:1505–1513. doi: 10.1016/0021-9290(95)00098-4. [DOI] [PubMed] [Google Scholar]
- 10.Belloc F, Dumain P, Boisseau MR, Jalloustre C, Reiffers J, Bernard P, et al. A flow cytometric method using Hoechst 33342 and propidium iodide for simultaneous cell cycle analysis and apoptosis determination in unfixed cells. Cytometry. 1994 Sep 1;17:59–65. doi: 10.1002/cyto.990170108. [DOI] [PubMed] [Google Scholar]
- 11.Birch HL, Rutter GA, Goodship AE. Oxidative energy metabolism in equine tendon cells. Res Vet Sci. 1997 Mar-Apr;62:93–97. doi: 10.1016/s0034-5288(97)90127-2. [DOI] [PubMed] [Google Scholar]
- 12.Birch HL, Smith TJ, Poulton C, Peiffer D, Goodship AE. Do regional variations in flexor tendons predispose to site-specific injuries? Equine veterinary journal. 2002;24:288–292. doi: 10.1111/j.2042-3306.2002.tb05435.x. [DOI] [PubMed] [Google Scholar]
- 13.Chae HJ, Kim SC, Han KS, Chae SW, An NH, Kim HM, et al. Hypoxia induces apoptosis by caspase activation accompanying cytochrome C release from mitochondria in MC3T3E1 osteoblasts. p38 MAPK is related in hypoxia-induced apoptosis. Immunopharmacol Immunotoxicol. 2001 May;23:133–152. doi: 10.1081/iph-100103855. [DOI] [PubMed] [Google Scholar]
- 14.Dahlgren LA, van der Meulen MC, Bertram JE, Starrak GS, Nixon AJ. Insulin-like growth factor-I improves cellular and molecular aspects of healing in a collagenase-induced model of flexor tendinitis. J Orthop Res. 2002;20:910–919. doi: 10.1016/S0736-0266(02)00009-8. [DOI] [PubMed] [Google Scholar]
- 15.Danis A. Soft tissue ossification: mechanism. Bull Mem Acad R Med Belg. 1992;147:298–306. discussion 306–297. [PubMed] [Google Scholar]
- 16.Dong Z, Wang JZ, Yu F, Venkatachalam MA. Apoptosis-resistance of hypoxic cells: multiple factors involved and a role for IAP-2. Am J Pathol. 2003 Aug;163:663–671. doi: 10.1016/S0002-9440(10)63693-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.El-Saggan AH, Dovinova I, Sulova Z, Barancik M, Hunakova L, Breier A, et al. Hypoxia increases cell death in multidrug-resistant leukemia cells. Differences in viability and ultrastructure between sensitive and multidrug-resistant L1210 mouse leukemic cells under hypoxia. Gen Physiol Biophys. 2003 Jun;22:265–273. [PubMed] [Google Scholar]
- 18.Fan X, Krahling S, Smith D, Williamson P, Schlegel RA. Macrophage surface expression of annexins I and II in the phagocytosis of apoptotic lymphocytes. Mol Biol Cell. 2004 Jun;15:2863–2872. doi: 10.1091/mbc.E03-09-0670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Fortier LA, Mohammed HO, Lust G, Nixon AJ. Insulin-like growth factor-I enhances cell-based repair of articular cartilage. J Bone Joint Surg Br. 2002;84:276–288. doi: 10.1302/0301-620x.84b2.11167. [DOI] [PubMed] [Google Scholar]
- 20.Galliani I, Burattini S, Mariani AR, Riccio M, Cassiani G, Falcieri E. Morpho-functional changes in human tendon tissue. Eur J Histochem. 2002;46:3–12. doi: 10.4081/1649. [DOI] [PubMed] [Google Scholar]
- 21.Jarvinen M, Jozsa L, Kannus P, Jarvinen TL, Kvist M, Leadbetter W. Histopathological findings in chronic tendon disorders. Scand J Med Sci Sports. 1997 Apr;7:86–95. doi: 10.1111/j.1600-0838.1997.tb00124.x. [DOI] [PubMed] [Google Scholar]
- 22.Kakar S, Khan U, McGrouther DA. Differential cellular response within the rabbit tendon unit following tendon injury. J Hand Surg [Br] 1998 Oct;23:627–632. doi: 10.1016/s0266-7681(98)80017-x. [DOI] [PubMed] [Google Scholar]
- 23.Kannus P, Jozsa L. Histopathological changes preceding spontaneous rupture of a tendon. A controlled study of 891 patients. J Bone Joint Surg Am. 1991 Dec;73:1507–1525. [PubMed] [Google Scholar]
- 24.Khan KM, Cook JL, Kannus P, Maffulli N, Bonar SF. Time to abandon the “tendinitis” myth. Bmj. 2002 Mar 16;324:626–627. doi: 10.1136/bmj.324.7338.626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kurosaka K, Takahashi M, Watanabe N, Kobayashi Y. Silent cleanup of very early apoptotic cells by macrophages. J Immunol. 2003 Nov 1;171:4672–4679. doi: 10.4049/jimmunol.171.9.4672. [DOI] [PubMed] [Google Scholar]
- 26.Kurtz CA, Loebig TG, Anderson DD, DeMeo PJ, Campbell PG. Insulin-like growth factor I accelerates functional recovery from Achilles tendon injury in a rat model. Am J Sports Med. 1999;27:363–369. doi: 10.1177/03635465990270031701. [DOI] [PubMed] [Google Scholar]
- 27.Maffulli N, Wong J, Almekinders LC. Types and epidemiology of tendinopathy. Clin Sports Med. 2003 Oct;22:675–692. doi: 10.1016/s0278-5919(03)00004-8. [DOI] [PubMed] [Google Scholar]
- 28.Mehta S, Gimbel JA, Soslowsky LJ. Etiologic and pathogenetic factors for rotator cuff tendinopathy. Clin Sports Med. 2003 Oct;22:791–812. doi: 10.1016/s0278-5919(03)00012-7. [DOI] [PubMed] [Google Scholar]
- 29.Molloy T, Wang Y, Murrell G. The roles of growth factors in tendon and ligament healing. Sports Med. 2003;33:381–394. doi: 10.2165/00007256-200333050-00004. [DOI] [PubMed] [Google Scholar]
- 30.Munkarah AR, Morris R, Baumann P, Deppe G, Malone J, Diamond MP, et al. Effects of prostaglandin E(2) on proliferation and apoptosis of epithelial ovarian cancer cells. J Soc Gynecol Investig. 2002 May-Jun;9:168–173. [PubMed] [Google Scholar]
- 31.Rempel D, Abrahamsson SO. The effects of reduced oxygen tension on cell proliferation and matrix synthesis in synovium and tendon explants from the rabbit carpal tunnel: an experimental study in vitro. J Orthop Res. 2001 Jan;19:143–148. doi: 10.1016/S0736-0266(00)00005-X. [DOI] [PubMed] [Google Scholar]
- 32.Renton A, Llanos S, Lu X. Hypoxia induces p53 through a pathway distinct from most DNA-damaging and stress-inducing agents. Carcinogenesis. 2003 Jul;24:1177–1182. doi: 10.1093/carcin/bgg044. [DOI] [PubMed] [Google Scholar]
- 33.Riley GP, Harrall RL, Constant CR, Chard MD, Cawston TE, Hazleman BL. Tendon degeneration and chronic shoulder pain: changes in the collagen composition of the human rotator cuff tendons in rotator cuff tendinitis. Ann Rheum Dis. 1994 Jun;53:359–366. doi: 10.1136/ard.53.6.359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Rocca M, Giavaresi G, Nicoli Aldini N, Fini M, Marcacci M, Zaffagnini S, et al. pO2 measurement in an experimental model of patellar tendon autograft pro-anterior cruciate ligament. Int J Artif Organs. 1998 Mar;21:174–178. [PubMed] [Google Scholar]
- 35.Sano H, Uhthoff HK, Backman DS, Brunet JA, Trudel G, Pham B, et al. Structural disorders at the insertion of the supraspinatus tendon. Relation to tensile strength. J Bone Joint Surg Br. 1998 Jul;80:720–725. doi: 10.1302/0301-620x.80b4.8203. [DOI] [PubMed] [Google Scholar]
- 36.Schmaltz C, Hardenbergh PH, Wells A, Fisher DE. Regulation of proliferation-survival decisions during tumor cell hypoxia. Mol Cell Biol. 1998 May;18:2845–2854. doi: 10.1128/mcb.18.5.2845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Skottner A, Arrhenius-Nyberg V, Kanje M, Fryklund L. Anabolic and tissue repair functions of recombinant insulin-like growth factor I. Acta Paediatr Scand Suppl. 1990;367:63–66. doi: 10.1111/j.1651-2227.1990.tb11635.x. [DOI] [PubMed] [Google Scholar]
- 38.Skottner A, Kanje M, Jennische E, Sjogren J, Fryklund L. Tissue repair and IGF-I. Acta Paediatr Scand Suppl. 1988;347:110–112. [PubMed] [Google Scholar]
- 39.Steenfos H, Spencer M, Hunt TK. Insulin-like growth factor I has a major role in wound healing. Surgical Forum 1989. 1989;40:68–70. [Google Scholar]
- 40.Stempien-Otero A, Karsan A, Cornejo CJ, Xiang H, Eunson T, Morrison RS, et al. Mechanisms of hypoxia-induced endothelial cell death. Role of p53 in apoptosis. J Biol Chem. 1999 Mar 19;274:8039–8045. doi: 10.1074/jbc.274.12.8039. [DOI] [PubMed] [Google Scholar]
- 41.Takemiya T, Maeda J. The functional characteristics of tendon blood circulation in the rabbit hindlimbs. Jpn J Physiol. 1988;38:361–374. doi: 10.2170/jjphysiol.38.361. [DOI] [PubMed] [Google Scholar]
- 42.Tandara AA, Mustoe TA. Oxygen in wound healing--more than a nutrient. World J Surg. 2004 Mar;28:294–300. doi: 10.1007/s00268-003-7400-2. [DOI] [PubMed] [Google Scholar]
- 43.Veitonmaki N, Cao R, Wu LH, Moser TL, Li B, Pizzo SV, et al. Endothelial cell surface ATP synthase-triggered caspase-apoptotic pathway is essential for k1–5-induced antiangiogenesis. Cancer Res. 2004 May 15;64:3679–3686. doi: 10.1158/0008-5472.CAN-03-1754. [DOI] [PubMed] [Google Scholar]
- 44.Vincent AM, Feldman EL. Control of cell survival by IGF signaling pathways. Growth Horm IGF Res. 2002 Aug;12:193–197. doi: 10.1016/s1096-6374(02)00017-5. [DOI] [PubMed] [Google Scholar]
- 45.Webster DF, Burry HC. The effects of hypoxia on human skin, lung and tendon cells in vitro. Br J Exp Pathol. 1982 Feb;63:50–55. [PMC free article] [PubMed] [Google Scholar]
- 46.Yuan J, Murrell GA, Trickett A, Wang MX. Involvement of cytochrome c release and caspase-3 activation in the oxidative stress-induced apoptosis in human tendon fibroblasts. Biochim Biophys Acta. 2003 Jun 17;1641:35–41. doi: 10.1016/s0167-4889(03)00047-8. [DOI] [PubMed] [Google Scholar]
- 47.Yuan J, Murrell GA, Wei AQ, Wang MX. Apoptosis in rotator cuff tendonopathy. J Orthop Res. 2002 Nov;20:1372–1379. doi: 10.1016/S0736-0266(02)00075-X. [DOI] [PubMed] [Google Scholar]