

Abstract
Cognitive behavioral therapy (CBT) is “gold standard” psychotherapy for social anxiety disorder (SAD). Cognitive models posit that preferential processing of threat mediates excessive forms of anxiety, which is supported by exaggerated amygdala, insula, and cortical reactivity to threatening socio-emotional signals in SAD. However, little is known about neural predictors of CBT success or the mechanisms by which CBT exerts its therapeutic effects. Functional magnetic resonance imaging (fMRI) was conducted during responses to social signals of threat (fearful/angry faces) against positive signals (happy faces) in 14 patients with SAD before and after 12 weeks of CBT. For comparison, 14 healthy control (HC) participants also underwent two fMRI scans, 12 weeks apart. Whole-brain voxel-wise analyses showed therapeutic success was predicted by enhanced pre-treatment activation to threatening faces in higher-order visual (superior and middle temporal gyrus), cognitive, and emotion processing areas (dorsal anterior cingulate cortex, dorsomedial prefrontal cortex). Moreover, a group by time interaction was revealed in prefrontal regions (dorsomedial, medial gyrus) and insula. The interaction was driven by relatively greater activity during threat processing in SAD, which significantly reduced after CBT but did not significantly predict response to CBT. Therefore, pre-treatment cortical hyperactivity to social threat signals may serve as a prognostic indicator of CBT success in SAD. Collectively, CBT-related brain changes involved a reduction in activity in insula, prefrontal, and extrastriate regions. Results are consistent with cognitive models, which associate decreases in threat processing bias with recovery.
Keywords: generalized social anxiety, fMRI, treatment, emotional faces, brain imaging
1. Introduction
Generalized social anxiety disorder (gSAD), also known as social phobia, is one of the most common anxiety disorders in the United States (Kessler et al., 2005). It is characterized by excessive fear and avoidance in a range of interpersonal situations that involve potential scrutiny by others (American Psychiatric Association, 2000). It begins early in life (Otto et al., 2001) and leads to severe impairment, substantially undermining educational attainment, employment, and relationship opportunities (Hambrick et al., 2003; Safren et al., 1996/1997; Schneier et al., 1994). The disorder often precedes and is co-morbid with other psychiatric illnesses such as depression, substance abuse, and additional anxiety disorders (Schneier et al., 1992).
Cognitive theories of anxiety posit preferential attention to threat-relevant signals is a core factor in the development and maintenance of gSAD and other excessive forms of anxiety (Beck and Clark, 1997; Beck et al., 2005; Eysenck, 1992; MacLeod et al., 2002; Wells and Matthews, 1994; Williams et al., 1988). Threat bias occurs throughout stages of information processing (Beck and Clark, 1997); accordingly, enhanced processing of threat cues during goal-directed attention (Pessoa et al., 2002) is an index of bias. Functional magnetic resonance imaging (fMRI) studies indicate attentional bias to threat in gSAD is subserved in part by exaggerated activation to threatening facial expressions in the amygdala and/or insula, which are key limbic/paralimbic structures involved in emotion generation and processing (Adolphs et al., 1995; Craig, 2009; Critchley, 2009; Davidson, 2000; Davis and Whalen, 2001; Jones et al., 2010). Exaggerated cortical response has also been observed in gSAD both to, and in anticipation of, threat (Etkin and Wager, 2007; Freitas-Ferrari et al., 2010). While not as consistently demonstrated as subcortical hyperactivity, exaggerated activity has also been shown in the anterior cingulate cortex (ACC) in gSAD (Amir et al., 2005; Blair et al., 2008; Goldin et al., 2009; Phan et al., 2006) a component of the anterior attention system involved in the detection of salient cues (Posner and Petersen, 1990). Other prefrontal cortex (PFC) regions serving executive functions (Bush et al., 2000; Etkin et al., 2011) have been implicated in gSAD such as dorsomedial, dorsolateral, and medial orbitofrontal cortex (Goldin et al., 2009; Stein et al., 2002; Tillfors et al., 2002) as well as higher-level visual regions (inferior occipital, fusiform gyrus, superior temporal sulcus; Evans et al., 2008; Goldin et al., 2009; Straube et al., 2004) engaged in social cognition (Adolphs 1999; Fusar-Poli et al., 2009). Collectively, evidence is building to show that emotional dysregulation in gSAD is mediated by exaggerated reactivity to signals of socio-emotional threat distributed among limbic/paralimbic, prefrontal, and extrastriate regions.
Despite advances in delineating the pathophysiology of gSAD, little is known about baseline neural correlates that predict treatment response or brain-based changes following cognitive-behavioral therapy (CBT), first-line psychotherapy for gSAD and other anxiety disorders (Heimberg, 2002; Hofmann and Smits, 2008). CBT aims to reduce anxiety symptoms through the modification of negative beliefs (e.g., cognitive restructuring) and behavior (e.g., exposure to fears; Beck et al., 2005). Greater response to treatment is associated with greater reductions in anxiety symptoms, negative beliefs, impairment (Brown et al., 1995; Hofmann and Smits, 2008; Hope et al., 1995), and attentional bias to threat stimuli (Mattia et al., 1993; Pishyar et al., 2008). Findings suggest that therapeutic response is associated with enhanced implicit emotion regulation, though it is not a direct target of treatment per se. Therefore, activity in regions implicated in emotional perception (e.g., evaluation of stimulus) and response to threat signals may predict recovery or change with treatment.
Successful CBT treatment in gSAD has been shown to be predicted by exaggerated pre-treatment activation to angry faces in dorsal and ventral occipitotemporal regions (Doehrmann et al., 2013), secondary visual areas involved in the decoding and processing of cues relevant to motivational state (Lang and Bradley 2010; Sabatinelli et al., 2013). Heightened activation in these higher-level visual areas has been proposed to correspond with emotion regulation capacity, which CBT is intended to enhance. Consequently, patients with less ability to effectively regulate emotions prior to undergoing treatment may experience reduced benefit from CBT (Doehrmann et al., 2013). Other baseline biomarkers of recovery in anxiety are ACC and amygdala. In a study of post-traumatic stress disorder (Bryant et al., 2008), CBT success was predicted by reduced ACC engagement and reduced amygdala response to subliminal fearful faces (i.e., masked). Thus, less reactivity to fear-evoking stimuli in this population is a good prognostic indicator of recovery with CBT (Bryant et al., 2008).
With regard to social anxiety and treatment-related changes in brain activity, a positron emission tomography study showed responders, regardless of treatment type (CBT or medication), exhibited reduced amygdala and ACC activity during symptom provocation involving a public speaking challenge (Furmark et al., 2002). Would analogous changes in brain activity be obtained outside of symptom elicitation, specifically, when merely processing threatening social signals? To our knowledge, this has yet to be investigated with regard to CBT in gSAD. However, fMRI studies of specific phobia comprising threat-relevant images provide clues. For example, in a study by Straube et al. (2006), baseline insula and ACC hyperactivity to spider images in phobics was significantly reduced in patients who completed CBT but not in a wait-list group of patients. Pre-treatment dorsolateral prefrontal response to spider stimuli has also been shown to decrease in patients after completing CBT (Paquette et al., 2003). Though findings have not always been replicated (Schienle et al., 2007), the extant data suggest that CBT exerts an attenuating effect on limbic-prefrontal reactivity to threat stimuli.
The objective of the present study was to evaluate neural predictors of CBT response and treatment-related brain-based changes to threat-relevant stimuli in gSAD. Accordingly, we used a perceptual face processing paradigm validated to isolate brain response to signals of threat (Hariri et al., 2002, 2005) before and after 12 weeks of manualized CBT (Hope et al., 2006). For comparison and to control for the effects of re-exposure to threat stimuli with repeated scanning, we enrolled a group of healthy control (HC) volunteers who were also scanned twice, 12 weeks apart.
Based on the literature and cognitive theory, we hypothesized symptom improvement would be predicted by greater pre-treatment activity in higher-level visual areas (e.g., occipitotemporal cortex) and less ACC and amygdala activity. Regarding brain changes, we hypothesized CBT would reduce exaggerated reactivity to threat faces in limbic/paralimbic areas (i.e., amygdala and insula), visual cortex, ACC and other medial PFC areas (e.g., dorsomedial, orbitofrontal) given their role in appraisal, response expression, and regulation (Bush et al., 2000; Etkin et al., 2011).
Furthermore, we hypothesized changes in activation would match clinical response to treatment such that post- versus pre-treatment changes in brain activation would correlate with changes in gSAD symptom severity.
2. Method
2.1. Ethics statements
All participants provided written informed consent in accordance with the Declaration of Helsinki and as approved by the Institutional Review Boards of the University of Michigan Medical School.
2.2 Participants
Fourteen individuals diagnosed with gSAD, who were not in psychotherapy, were identified through local community advertisement and via referrals from an outpatient psychiatric clinic. Fourteen demographically-matched HC were recruited through community advertisements. All participants completed the Structured Clinical Interview for DSM-IV (SCID; First et al., 1997) conducted by licensed clinicians in conjunction with measures of symptoms and negative mood such as the Liebowitz Social Anxiety Scale (LSAS) (Liebowitz, 1987) and Beck Depression Inventory (BDI) (Beck et al., 1996). Regarding symptoms, an LSAS total score cut off of 55 was used to capture levels of symptom severity that ranged from the moderate to the severe (Heimberg et al., 1999). Clinical response was measured with the Clinical Global Impression-Improvement (CGI-I) scale, a clinician-based rating that takes into consideration symptom severity and changes in a patient’s functioning over time. All participants were free of psychotropic medications except for two individuals with gSAD, who were on a stable dose of bupropion for at least 8 weeks before the study with no changes in medication during the study. None of the gSAD participants had a current major depressive episode, severe depression symptoms (i.e., BDI score of 30 or greater; Beck et al., 1996), recent substance abuse/dependence (within 6 months of study), or any history of major psychiatric illness (e.g., bipolar, psychotic disorder). All participants were between 18 and 55 years of age, right-handed, and free of current and past major medical or neurologic illness, as confirmed by a Board Certified physician. None of the participants tested positive for alcohol or illegal substances. Table 1 details the demographic and clinical characteristics of the participants.
Table 1.
Demographic and Clinical Characteristics of Patients and Control Subjects
| gSAD (N=14) | HC (N=14) | |||||
|---|---|---|---|---|---|---|
|
|
||||||
| Mean | SD | Mean | SD | t | p | |
| Age (years) | 28.07 | 8.62 | 23.29 | 5.44 | 1.76 | 0.10 |
| Liebowitz Social Anxiety Scale* | 71.21 | 9.61 | 9.71 | 6.46 | 19.87 | <0.0001 |
| Spielberger State Anxiety | 42.21 | 9.02 | 21.55a | 2.21 | 8.26 | <0.0001 |
| Spielberger Trait Anxiety | 48.07 | 8.43 | 24.73a | 9.76 | 7.23 | <0.0001 |
| Beck Depression Inventory | 10.43 | 8.02 | 1.18a | 1.25 | 4.25 | <0.001 |
|
|
||||||
| N | % | N | % | χ2 | p | |
|
|
||||||
| Gender | 0.15 | 0.70 | ||||
| Male | 5 | 35.7 | 6 | 42.9 | ||
| Female | 9 | 64.3 | 8 | 57.1 | ||
| Race | 3.47 | 0.33 | ||||
| Caucasian | 8 | 57.1 | 12 | 85.7 | ||
| Asian | 2 | 14.3 | 1 | 7.1 | ||
| African American | 2 | 14.3 | 1 | 7.1 | ||
| Hispanic | 2 | 14.3 | ||||
gSAD=generalized social anxiety disorder; HC=healthy control. Spielberger Anxiety Inventory and Beck Depression Inventory measures were not obtained in some HC subjects (aHC n = 11).
Total score calculated by summing total fear and total avoidance subscales.
Patients received 12 weeks of manualized individual CBT (Hope et al., 2006), which consisted of one 60-minute session per week, conducted by a licensed clinical psychologist under the supervision of a licensed clinical psychologist with expertise in CBT and clinical trial investigations involving CBT to ensure adherence to treatment. CBT comprised psychoeducation, cognitive restructuring, in vivo exposures, and relapse prevention (Hope et al., 2006).
2.3 fMRI Task
During scanning, all participants performed a modified emotional face matching task designed to isolate key subcortical emotion processing areas (e.g., amygdala, insula) to signals of threat (i.e., angry, fearful faces) from non-threatening stimuli (happy faces). Prior evidence from our laboratory (Phan et al., 2006, 2013) and others (Stein et al., 2002) has shown that patients with gSAD differ significantly in their fronto-limbic reactivity to threatening (angry/fearful) but not non-threat (happy) signals, whereas this differential response is less evident in healthy volunteers (Fitzgerald et al., 2006). To maximize the activation ‘signal’ for CBT effects, we contrasted threatening against happy faces, rather than neutral faces, fixation crosses or shapes, as this contrast results in more robust and consistent findings of exaggerated limbic reactivity in gSAD based on our (Phan et al., 2006, 2013) and others’ previous work (Etkin and Wager, 2007; Freitas-Ferrari et al., 2010). Moreover, we contrasted angry versus happy faces (AvH) and fearful against happy faces (FvH) allowing us to examine potential separate effects for the type of threat signal based on evidence that angry and fear differentially perturb emotion processing circuitry (Fusar-Poli et al., 2009; Whalen et al., 2001), which may influence treatment-related brain changes. For example, in patients with gSAD, brain changes in response to a selective-serotonin reuptake-inhibitor related to angry and fearful faces occur in different ways, such that, effective treatment attenuated amygdala reactivity to FvH (not AvH) and enhanced ventral medial prefrontal cortex activity to AvH (not FvH) (Phan et al., 2013).
The task comprised photographs from a validated set of face stimuli (Gur et al., 2002) presented in a block-design, during which participants viewed a trio of faces (one target at top, two probes on bottom) and selected by right-handed button press which probe matched the target facial expression. The target and congruent probe faces displayed one of three expressions (fearful, angry, or happy), while the incongruent probe face always displayed a neutral expression. The paradigm consisted of 18 experimental 20-second blocks: 9 blocks of matching emotional faces with each target expression of angry, fearful, or happy interleaved with 9 blocks of matching shapes (triangles, circles, squares) as a sensorimotor control condition, which were counterbalanced across 2 runs for a total task time of 6 minutes. Each task block contained 4 sequential matching trials, 5 seconds each. Therefore, the inter-stimulus interval for faces, the main blocks of interest, was 20 seconds.
2.4 fMRI Data Acquisition
Functional imaging was performed with blood-oxygen-level-dependent (BOLD) sensitive whole-brain fMRI on a 3.0 Tesla GE Signa System (General Electric; Milwaukee, WI) using a 4-channel GE Quadrature sending and receiving head coil. Images were acquired with 30 axial, 5-millimeter (mm) thick slices using a standard T2*-sensitive gradient echo reverse spiral acquisition sequence (2 second repetition time; 25 millisecond (ms) echo time; 64 × 64 matrix; 24 centimeter field of view; flip angle 77°; 3.75 × 3.75 × 5 mm final voxel size). A high-resolution, T1-weighted volumetric anatomical scan was also acquired in the axial plane (9 ms repetition time, 1.8 ms echo time; 256 × 256 matrix; 256 mm field of view; flip angle 15°; 124 slices; 1.2 mm slice thickness) at the same position as the functional images for anatomical localization.
2.5 fMRI Data Preprocessing
Data from all participants were visually examined for movement during the scan and participants with moderate to extreme motion (movement greater than 3 mm in any direction or 3 degrees of displacement across each functional run) were excluded and the first 4 volumes from each run were discarded to allow for T1 equilibration effects. Conventional preprocessing steps were used in Statistical Parametric Mapping (SPM8) software package (Wellcome Trust Centre for Neuroimaging, London www.fil.ion.ucl.ac.uk/spm). Briefly, images were temporally corrected to account for differences in slice time collection, spatially realigned to the first image of the first run, normalized to a Montreal Neurological Institute (MNI) template, and smoothed with an 8 mm isotropic Gaussian kernel.
Using a box-car model, contrasts of interest (AvH and FvH) were generated for each participant and then entered into a second-level general linear model treating participants as a random effect (i.e., a random effects analysis). To evaluate predictors of treatment response, contrasts of interest from gSAD pre-treatment scans were entered into a whole-brain analysis of covariance, regressing LSAS change (ΔPreTx - PostTx) while initial severity (LSASPreTx) was controlled for as a regressor of no interest. Consistent with prior fMRI studies of gSAD concerning whole-brain analysis (Amir et al., 2005; Evans et al., 2008; Gentili et al., 2008; Phan et al., 2013), significance was set at p<0.001 (uncorrected) with a cluster extent threshold of greater than 10 contiguous voxels (volume > 80 mm3). This type of joint intensity and cluster size threshold is within the recommended threshold to attain a suitable balance between Type I and Type II errors (Lieberman and Cunningham, 2009).
To examine changes in activation, a whole-brain analysis of variance (ANOVA) using Group (gSAD, HC) and Time (pre-treatment/scan 1, post-treatment/scan 2) as between- and within-subjects factors, respectively, was conducted to test for main effects of Group and Time, as well as for Group x Time interactions. Significance was again set at p<0.001 (uncorrected) with a cluster extent threshold of greater than 10 contiguous voxels.
To identify anatomical labels for whole-brain findings in SPM8, we used the Automated Anatomical Labeling (AAL) atlas based on the Talairach Daemon database (Lancaster et al., 1997, 2000; Tzourio-Mazoyer et al., 2002). To illustrate the magnitude and direction of activation, parameter estimates of peak activation (βweights, arbitrary units [a.u.]) were extracted from spherical (10-mm diameter) regions of interest from each participant and submitted to post hoc scatter plots of the regression analyses, t-tests for ANOVA findings, and examination of associations between ANOVA findings and symptom severity in the Statistical Package for the Social Sciences (SPSS) (Chicago, IL version 18).
3. Results
3.1 Behavioral Performance
Probe identification accuracy averaged 93% within two seconds of the trial duration indicating participants followed instructions. A 2 (Group: gSAD, HC) × 2 (Time: Scan 1, Scan 2) x 3 (Emotion: Angry, Fearful, Happy) ANOVA with repeated measures on the last two factors showed a main effect of Emotion [F(2, 52) 35.22, p<0.001] but no main effect of Group, Time, or Group x Time interaction (all ps > 0.05). Simple effects analysis revealed that all participants were more accurate at matching happy (mean accuracy ± standard deviation %: 97.2 ± 7.9%) than angry faces (86.6 ± 10.0%; p<0.001) and more accurate at matching fearful (96.0 ± 6.3%) than angry faces (p<0.001). No difference emerged between happy and fearful faces (p=0.30). See Figure 1(A) for accuracy per trial type.
Figure 1.


A) Mean responses as a percentage of correctly matched angry, fearful, and happy faces at Time 1 and Time 2 in gSAD and HC. B) Mean reaction times in milliseconds for correctly matched angry, fearful, and happy faces at Time 1 and Time 2 in gSAD and HC. gSAD, Generalized Social Anxiety Disorder; Healthy Control (HC)
Regarding response times for accurate trials, an ANOVA showed main effects of Emotion [F(2, 52) 27.51, p<0.001] and Time [F(1, 26) 6.16, p<0.020]. There was no main effect of Group or Group x Time interaction (all ps > 0.05). Simple effects showed all participants were faster at matching happy (mean reaction time ± standard deviation: 1355 ± 326 ms) than angry faces (1553 ± 338 ms; p<0.001) and faster for fearful (1404 ± 326 ms) than angry faces (p<0.001). Response times were similar for happy and angry faces (p=0.12). See Figure 1(B) for reaction times per trial type.
3.2 Treatment Effects on Social Anxiety Severity
After 12 weeks of individual CBT, gSAD symptom severity, as assessed with LSAS, significantly decreased from an average of 71.2 ± 9.6 to 49.9 ± 20.6 (t=4.01, p<0.001). Based on the CGI-I, about 70% of the gSAD group (10 of 14 patients) were considered to be ‘Responders’ as they were rated to be ‘very much improved’ or ‘much improved’ (CGI-I score of 1 or 2) while 4 of 14 patients had a CGI-I score of >2 post-treatment and were, thus, considered ‘Non-Responders’.
3.3 fMRI Results
3.3.1. Brain predictors
Controlling for initial severity (LSASPreTx), greater pre-treatment response to fearful faces (vs. happy) corresponded with greater symptom improvement following CBT (ΔPreTx - PostTx) in visual (e.g., superior temporal region comprising angular gyrus) (Figure 2) and prefrontal regions (e.g., medial orbitofrontal) (Figure 3). This analysis did not reveal significant covariation with treatment response to activity in the amygdala or insula.
Figure 2.

A) Regressing LSAS change (ΔPreTx - PostTx) while initial severity (LSASPreTx) is controlled for as a regressor of no interest; brain map depicts whole-brain analysis of covariance showing enhanced superior temporal gyrus reactivity to fearful faces in gSAD displayed on statistical t-map at p<0.001. B) Scatter plot of the regression analyses depicting extracted parameter estimates of activation from superior temporal gyrus region of interest showing greater response to CBT in gSAD is predicted by enhanced superior temporal gyrus reactivity to fearful faces.
LSAS, Liebowitz Social Anxiety Scale; CBT, Cognitive Behavioral Therapy; gSAD, Generalized Social Anxiety Disorder.
Figure 3.
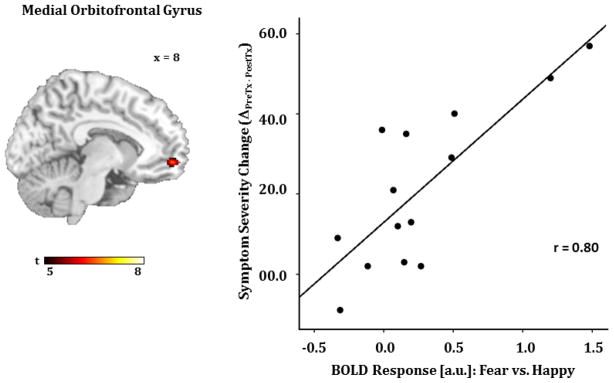
A) Regressing LSAS change (ΔPreTx - PostTx) while initial severity (LSASPreTx) is controlled for as a regressor of no interest; brain map depicts whole-brain analysis of covariance showing increased medial orbitofrontal gyrus reactivity to fearful faces in gSAD displayed on statistical t-map at p<0.001. B) Scatter plot of the regression analyses depicting extracted parameter estimates of activation from medial orbitofrontal region of interest showing greater response to CBT in gSAD is predicted by greater medial orbitofrontal gyrus reactivity to fearful faces.
LSAS, Liebowitz Social Anxiety Scale; CBT, Cognitive Behavioral Therapy; gSAD, Generalized Social Anxiety Disorder.
With regard to angry faces (vs. happy), symptom improvement was predicted by enhanced baseline activity in visual areas (i.e., middle temporal and angular gyri). Analogous to fearful faces, no significant effects involved amygdala or insula activity. See Table 2 for all significant regression results comprising both fearful and angry expressions.
Table 2.
Pre-Treatment to Post-Treatment Decrease in Social Anxiety Severity, Controlling for Pre-Treatment Severity, and Change in Brain Activation: Whole-brain Voxel-wise Regressiona
| Region | MNI Coordinates | Volume (mm3) | Z | |||
|---|---|---|---|---|---|---|
| x | y | z | ||||
|
Fearful > Happy
|
||||||
|
Positive Correlation
|
Superior Medial Frontal Gyrus | −18 | 68 | 16 | 1632 | 4.57 |
| −4 | 36 | 58 | 656 | 3.76 | ||
| Angular Gyrus | −58 | −66 | 28 | 568 | 4.55 | |
| Medial Orbitofrontal Gyrus | 8 | 56 | −8 | 712 | 4.03 | |
| Precentral Gyrus | −36 | 6 | 50 | 1624 | 3.99 | |
| Middle Temporal Gyrus | −68 | −42 | 4 | 536 | 3.86 | |
| Anterior Cingulum | −8 | 34 | 26 | 1064 | 3.81 | |
| 8 | 38 | 24 | 712 | 3.57 | ||
| Inferior Orbital Frontal Gyrus | −38 | 18 | −12 | 80 | 3.66 | |
| Cuneus | −4 | −84 | 40 | 88 | 3.66 | |
| Thalamus | −8 | −18 | 2 | 336 | 3.59 | |
| Middle Frontal Gyrus | −28 | 30 | 38 | 272 | 3.53 | |
| Precuneus | 10 | −64 | 18 | 104 | 3.50 | |
| Superior Temporal Gyrus | −62 | −36 | 20 | 856 | 3.41 | |
| Cerebellum | 20 | −64 | −48 | 248 | 3.38 | |
| Negative Correlation | No significant clusters | |||||
|
Angry > Happy
|
||||||
| Positive Correlation | Middle Temporal Gyrus | 66 | −10 | −24 | 112 | 3.91 |
| Angular Gyrus | −44 | −68 | 42 | 96 | 3.36 | |
| Negative Correlation | Cerebellum | 44 | −58 | −32 | 104 | 3.75 |
All listed clusters significant at p<0 .001 (uncorrected) with a cluster extent threshold of greater than 10 contiguous voxels. MNI, Montreal Neurological Institute; Z, Z-score.
3.3.2. Brain changes
Whole-brain voxel-wise ANOVAs for fearful faces (vs. happy) and angry faces (vs. happy), respectively, were used to evaluate global brain changes, which were followed up with simple effects analyses in SPSS. For fearful faces (vs. happy), a main effect of Group for fusiform gyrus was revealed but no main effect of Time or Group x Time interaction was evident. Follow-up analysis showed gSAD relative to HC exhibited heightened fusiform response to fear.
ANOVA results for angry (vs. happy) faces revealed a main effect of Group but not Time. The Group finding was driven by activity in the parahippocampal gyrus, which was enhanced in HC compared to gSAD. Importantly, there was a significant Group x Time interaction for right insula and prefrontal regions, specifically right medial orbitofrontal and right dorsomedial PFC (e.g., superior medial frontal gyrus in AAL) to angry (vs. happy) faces (Table 3, Figures 4 and 5).
Table 3.
Brain Activation to Social Signals of Threat: Whole-brain Voxel-wise ANOVAa
| Region | MNI Coordinates | Volume (mm3) | F | |||
|---|---|---|---|---|---|---|
| x | y | z | ||||
|
Fearful > Happy
|
||||||
| Main Effect of Group | Fusiform Gyrus | 44 | −34 | −24 | 80 | 16.23 |
| Main Effect of Time | No significant clusters | |||||
| Group x Time Interaction | No significant clusters | |||||
|
Angry > Happy
|
||||||
| Main Effect of Group | Parahippocampal Gyrus | −14 | −24 | −16 | 176 | 19.57 |
| Main Effect of Time | No significant clusters | |||||
| Group x Time Interaction | Medial Orbitofrontal Gyrus | 12 | 56 | −4 | 144 | 18.71 |
| Superior Medial Frontal Gyrus | 6 | 62 | 20 | 192 | 16.51 | |
| Hippocampus | −24 | −40 | 0 | 216 | 16.26 | |
| Superior Temporal Gyrus | −46 | −4 | −10 | 160 | 16.03 | |
| Insula | 34 | −6 | 14 | 80 | 14.47 | |
All listed clusters significant at p<0 .001 (uncorrected) with a cluster extent threshold of greater than 10 contiguous voxels. MNI, Montreal Neurological Institute; F, F-score.
Figure 4.
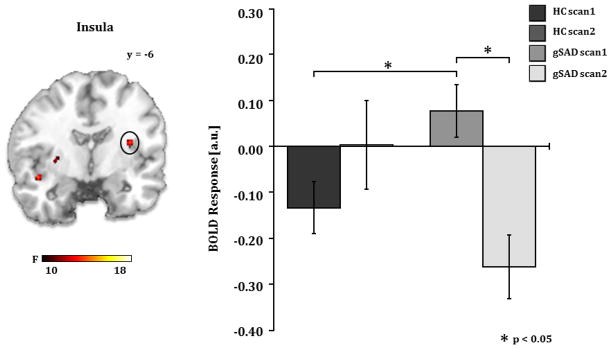
A) Brain map depicting whole-brain voxel-wise ANOVA F-map at p<0.001 showing a significant Group x Time interaction in the insula in response to angry faces. B) Bar graph depicts extracted BOLD signal change from insula region of interest showing insula reactivity to angry faces is greater in gSAD than the HC group at pre-treatment (gSADPre-Tx > HCScan 1, p<0.02) and is reduced by treatment (gSADPre-Tx > gSADPost-Tx, p<0.002). gSAD, Generalized Social Anxiety Disorder; Healthy Control (HC)
Figure 5.
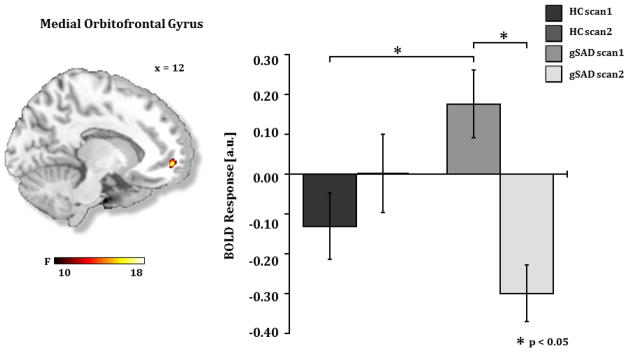
A) Brain map depicting whole-brain voxel-wise ANOVA F-map at p<0.001 showing a significant Group x Time interaction in the medial orbitofrontal gyrus in response to angry faces. B) Bar graph depicts extracted BOLD signal change from medial orbitofrontal region of interest showing medial orbitofrontal reactivity to angry faces is greater in gSAD than the HC group at pre-treatment (gSADPre-Tx > HCScan 1, p<0.02) and is reduced by treatment (gSADPre-Tx > gSADPost-Tx, p<0.001). gSAD, Generalized Social Anxiety Disorder; Healthy Control (HC)
Follow-up analysis within these regions of interest showed reactivity to angry (vs. happy) faces was greater in the gSAD group than in healthy controls (HC) at pre-treatment (gSADPreTx > HCScan1, p<0.05) and was significantly reduced after treatment (gSADPreTx > gSADPostTx, p<0.05). Within group analysis revealed that pre-treatment activity in insula, medial frontal gyrus, and dorsomedial PFC significantly decreased after treatment while activity in these regions between Scan 1 and Scan 2 did not significantly change in the HC group.
3.3.3. Correlation between brain changes and symptom severity
Pearson correlational analyses between treatment change (ΔPreTx - PostTx) in social anxiety severity scores (i.e., LSAS) and BOLD signal treatment change (ΔPreTx - PostTx) evident in ANOVA findings regarding angry (vs. happy) faces, specifically, insula, medial frontal gyrus, and dorsomedial PFC, did not reveal any further significant relationships (ps>0.05).
4. Discussion
To our knowledge this is the first cognitive behavioral therapy (CBT) study in patients with generalized social anxiety disorder (gSAD) to use fMRI to evaluate both neural predictors of treatment response and the general effects of CBT on brain activity to social threat signals. We observed that the majority of patients benefitted from CBT and, as hypothesized, greater symptom reduction was predicted by greater pre-treatment activity to threat (i.e., fearful faces) in higher-level visual regions; namely, superior and middle temporal gyri and inferior frontal gyrus, implicated in the detection and appraisal of socio-emotional cues (Adolphs, 1999; Fusar-Poli et al., 2009). Amygdala activity did not emerge as a marker of recovery as was demonstrated in post-traumatic stress disorder to subliminal threat cues (Bryant et al., 2008). Moreover, while anterior cingulate cortex (ACC) was apparent, it was increased, not decreased, pre-treatment activity that corresponded with CBT success. Methodological differences may have contributed to inconsistent findings. In addition to bilateral dorsal ACC (i.e., anterior cingulum in Table 2), symptom improvement was predicted by baseline hyperactivity to fearful faces in dorsomedial prefrontal and orbitofrontal regions, implicated in appraisal, fear response, and/or regulatory functions (Etkin et al., 2011; Northoff et al., 2006; Rothkirch et al., 2012). Though the pattern of pre-treatment activity predicting symptom improvement was mainly for fearful faces, activity in the middle temporal gyrus for angry faces also positively covaried with recovery suggesting greater reactivity in higher-order visual areas to threat in general predicts CBT success. These results replicate prior evidence that exaggerated activation to threatening faces in extrastriate cortex, but not limbic regions, can foretell CBT success in gSAD (Doehrmann et al., 2013). Together with our prefrontal findings, greater pre-treatment activation to threat faces appears to reflect a hypersensitive system capitalized on by CBT. For example, individuals with neural substrates that drive inflated perceptions of, and/or reactivity to, external threat cues may be more likely to experience reductions in anxiety level or altered perceptions of threat when engaging in cognitive restructuring or exposure exercises in a therapeutic environment.
Regarding changes in brain pathophysiology, patients with gSAD relative to healthy controls (HC) exhibited pre-treatment exaggerated insula, but not amygdala, activity to angry (vs. happy) faces, which significantly decreased in patients after 12 weeks of CBT. No significant changes emerged after 12 weeks in the HC group. The null amygdala result here is somewhat surprising. One possibility is our use of happy faces as a contrast to angry/fearful faces, which we based on fronto-limbic findings that the contrast differentiates gSAD from healthy volunteers (Phan et al., 2006, 2013; Stein et al., 2002). Though not a consistent finding, happy faces can elicit amygdala reactivity in social anxiety (Evans et al., 2008; Straube et al., 2005), potentially due to negative interpretations of the expression (unapproachable, mocking; Campbell et al., 2009; Coles and Heimberg, 2005). Given our small sample size, individual differences in the interpretation of happy may have reduced our ability to find significant amygdala effects. Nevertheless, our hypothesis regarding treatment-related reductions in subcortical activity was partially supported due to insula findings. Insula contributes to the representation of feelings by bringing afferent signals to awareness (Craig, 2009; Critchley et al., 2004) and aversive interoceptive cues or inaccurate interpretations of bodily state are thought to underlie anxiety (Paulus and Stein, 2006, 2010) as evidenced by insula hyperactivity in gSAD (Etkin and Wager, 2007; Freitas-Ferrari et al., 2010). The CBT-related reduction in insula activity to angry faces in gSAD has also been shown in post-CBT spider phobics to negative images (Schienle et al., 2007; Straube 2006) and indicates CBT may help remediate aberrant activity that contributes to emotional awareness by attenuating reactivity during evaluation of threat. Pre-/post-treatment brain changes in the gSAD group to angry faces were also evident in the dorsomedial prefrontal and orbitofrontal areas relative to the HC group. Again, activation significantly decreased in the gSAD group after CBT and no significant changes were demonstrated in the HC group.
Concerning other group effects, there were regions that discriminated activity between our gSAD and HC sample but remained unaffected by CBT. Main effects of Group at baseline showed heightened fusiform response to fearful (vs. happy) faces in gSAD; meanwhile, the HC group demonstrated greater parahippocampal activation for angry (vs. happy) faces. We hesitate to interpret these preliminary, exploratory results, as we did not have a priori hypotheses regarding these regions.
Furthermore, in contrast to our hypothesis, symptom severity did not correlate with brain activation after treatment suggesting effects on brain activity, particularly insula reactivity to emotional faces, may not be related to clinical change. Alternatively, conventional symptom measures may not be an optimal probe of neural modification to salient cues. For example, in a specific phobia study by Schienle et al. (2007), a CBT-related reduction in insula activity to spider pictures correlated with a reduction in somatic panic symptoms as opposed to phobic severity. Thus, an assessment that taps into the function of a region of interest may serve as a more sensitive index of treatment-related brain change.
Our preliminary findings indicate CBT success was predicted by baseline hyper-reactivity to fearful faces in higher-order visual and prefrontal areas. Whereas CBT-related changes were observed as reductions in insula and prefrontal activity to angry faces in gSAD. Fearful expressions signal danger in the environment and its biological relevance pertains to the facilitation of defense mechanisms (Öhman and Mineka, 2001). Unlike angry facial expressions, which convey direct interpersonal aggression, the source of threat communicated by fear is more ambiguous (Ewbank et al., 2009; Fox et al., 2007; Whalen, 1998). Therefore, results could in part be due to an individual’s more elaborative interpretation of fearful faces, for instance, the extent to which it is relevant to oneself. Together, exaggerated response to fearful faces may be a particularly useful prognostic indicator while angry faces may be a more sensitive probe of CBT effects.
Of note, activity in the medial orbitofrontal gyrus was observed at the level of neural predictors for fearful faces and group effects for angry faces. This region is a network comprising the ventral medial prefrontal cortex and medial division of orbitofrontal cortex (Ongur et al., 2003; Ongur and Price, 2000), which underlie various executive functions (Cardinal et al., 2002; Etkin et al., 2011; Kringelbach and Rolls, 2004; Phillips, 2003; Rolls, 2004; Schoenbaum and Roesch, 2005). Most relevant in the context of our perceptual matching task is its involvement in evaluative functions and emotional response. Insofar as predictors are concerned, the correspondence between exaggerated pre-treatment activity in medial orbitofrontal gyrus to fearful faces and CBT response indicates the greater the capacity to appraise an ambiguous signal of danger, the more likely CBT will be successful. On the other hand, medial orbitofrontal response in gSAD to angry faces may reflect heightened emotional reactivity to cues of aggression, which subsequently decreases with treatment. In summary, threat type may reveal distinguishable medial orbitofrontal functions in the context of neural predictors of CBT success and treatment-related mechanisms of modification.
4.1. Limitations
Our study is not without notable limitations. First, the study design lacked a wait-list control group; therefore, neural and clinical findings cannot be causally attributed to CBT and could be related to a number of factors not related to treatment such as natural course of the illness over the 12-week period, differential regression to the mean in patients and controls, and neural habituation effects. Second, our small sample size may have increased risk for false negatives, and may have contributed to not finding baseline differences or treatment effects in the amygdala. Third, 2 of the 14 participants with gSAD were taking bupropion; though the medication was stable before entering the study and remained unchanged during the study, any influence it may have had on results cannot be ruled out. Additionally, because the majority of patients were considered to have had a positive response to CBT, the study may not have had sufficient power to detect a significant correlation between changes in brain activation with that in symptom severity or to examine differences in brain changes between treatment responders (n=10) versus non-responders (n=4). The pre-treatment exploratory whole-brain regression analysis (Table 2) suggests that brain areas correlating with CBT response may not be localized within the amygdala or insula. Although these observations fit with a broader model incorporating a larger set of brain targets relevant to the pathophysiology of gSAD (Freitas-Ferrari et al., 2010; Hofmann et al., 2012; Shin and Liberzon, 2010), the exploratory nature of these findings, and lack of independent evaluators of treatment fidelity and symptom change, warrants replication and further investigation. Because pre-treatment/post-treatment neural changes were unrelated to social anxiety symptom improvement, we caution against interpreting those changes as being directly related to treatment response. Moreover, results cannot be generalized to other anxiety disorders or to other treatments such as pharmacologic interventions also proven to be effective in treating gSAD. Future studies are needed to determine if the brain effects observed here are specific to CBT or shared across any therapeutic modality as long as the treatment is effective.
5. Conclusions
Our findings provide evidence CBT success is predicted by greater pre-treatment cortical reactivity to threat and that CBT reduces limbic-cortical activity to social signals of threat in patients with generalized social anxiety disorder. The observation that neural predictors and mechanisms of change were modulated by angry and fearful faces suggest type of socio-emotional threat signal should be considered when evaluating neural markers. Future studies that employ a more comprehensive set of threat stimuli with randomized placebo-controlled and/or comparative active treatment designs and larger samples are needed to determine whether CBT effects are mediated by these specific patterns of brain changes so that we can better delineate mechanisms of therapeutic actions of CBT and other effective treatments and predictors of treatment response.
Highlights.
Cognitive behavioral therapy (CBT) is a common psychotherapy for anxiety disorders
Baseline activity in visual, emotion, and cognitive areas correlated with CBT success
CBT-related changes included decreases in emotion processing and regulatory regions
Acknowledgments
This work was supported by grants from the National Institutes of Health, National Institute of Mental Health (MH076198 to KLP and MH093679 to HK).
Footnotes
Declaration of Interest
None.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Adolphs R. Social cognition and the human brain. Trends Cogn Sci. 1999;12:469–479. doi: 10.1016/s1364-6613(99)01399-6. [DOI] [PubMed] [Google Scholar]
- Adolphs R, Tranel D, Damasio H, Damasio AR. Fear and the human amygdala. J Neurosci. 1995;15:5879–5891. doi: 10.1523/JNEUROSCI.15-09-05879.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- American Psychiatric Association. Diagnostic and statistical manual of mental disorders. 4. Washington, DC: American Psychiatric Association; 2000. text revision. [Google Scholar]
- Amir N, Klumpp H, Elias J, Bedwell JS, Yanasak N, Miller LS. Increased activation of the anterior cingulate cortex during processing of disgust faces in individuals with social phobia. Biol Psychiatry. 2005;57:975–981. doi: 10.1016/j.biopsych.2005.01.044. [DOI] [PubMed] [Google Scholar]
- Beck AT, Clark DA. An information processing model of anxiety: automatic and strategic processes. Behav Res Ther. 1997;35:49–58. doi: 10.1016/s0005-7967(96)00069-1. [DOI] [PubMed] [Google Scholar]
- Beck AT, Emery G, Greenberg RL. Anxiety disorders and phobias: a cognitive perspective. New York: Basic Books; 2005. [Google Scholar]
- Beck AT, Steer RA, Brown GK. Manual for the Beck Depression Inventory-II. San Antonio, TX: Psychological Corp; 1996. [Google Scholar]
- Blair K, Shaywitz J, Smith BW, Rhodes R, Geraci M, Jones M, Blair RJR, Pine DS. Response to emotional expressions in generalized social phobia and generalized anxiety disorder: evidence for separate disorders. Am J Psychiatry. 2008;165:1193–1202. doi: 10.1176/appi.ajp.2008.07071060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown EJ, Heimberg RG, Juster HR. Social phobia subtype and avoidant personality disorder: effect on severity of social phobia, impairment, and outcome of cognitive behavioral treatment. Behav Ther. 1995;26:467–486. [Google Scholar]
- Bryant RA, Felmingham K, Kemp A, Das P, Hughes G, Peduto A, Williams L. Amygdala and ventral anterior cingulate activation predicts treatment response to cognitive behaviour therapy for post-traumatic stress disorder. Psychol Med. 2008;38:555–561. doi: 10.1017/S0033291707002231. [DOI] [PubMed] [Google Scholar]
- Bush G, Luu P, Posner MI. Cognitive and emotional influences in anterior cingulate cortex. Trends Cogn Sci. 2000;4:215–222. doi: 10.1016/s1364-6613(00)01483-2. [DOI] [PubMed] [Google Scholar]
- Campbell DW, Sareen J, Stein MB, Kravetsky LB, Paulus MP, Hassard ST, Reiss JP. Happy but not so approachable: the social judgments of individuals with generalized social phobia. Depress Anxiety. 2009;26:419–424. doi: 10.1002/da.20474. [DOI] [PubMed] [Google Scholar]
- Cardinal RN, Parkinson JA, Hall J, Everitt BJ. Emotion and motivation: the role of the amygdala, ventral striatum, and prefrontal cortex. Neurosci Biobehav Rev. 2002;26:321–352. doi: 10.1016/s0149-7634(02)00007-6. [DOI] [PubMed] [Google Scholar]
- Coles ME, Heimberg RG. Recognition bias for critical faces in social phobia: a replication and extension. Behav Res Ther. 2005;43:109–120. doi: 10.1016/j.brat.2003.12.001. [DOI] [PubMed] [Google Scholar]
- Craig ADB. How do you feel--now? The anterior insula and human awareness. Nat Rev Neurosci. 2009;10:59–70. doi: 10.1038/nrn2555. [DOI] [PubMed] [Google Scholar]
- Critchley HD. Psychophysiology of neural, cognitive and affective integration: fMRI and autonomic indicants. Int J Psychophysiol. 2009;73:88–94. doi: 10.1016/j.ijpsycho.2009.01.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Critchley HD, Wiens S, Rotshtein P, Öhman A, Dolan RJ. Neural systems supporting interoceptive awareness. Nat Neurosci. 2004;7:189–195. doi: 10.1038/nn1176. [DOI] [PubMed] [Google Scholar]
- Davidson RJ. Affective style, psychopathology, and resilience: brain mechanisms and plasticity. Am Psychol. 2000;55:1196–1214. doi: 10.1037//0003-066x.55.11.1196. [DOI] [PubMed] [Google Scholar]
- Davis M, Whalen PJ. The amygdala: vigilance and emotion. Mol Psychiatry. 2001;6:13–34. doi: 10.1038/sj.mp.4000812. [DOI] [PubMed] [Google Scholar]
- Doehrmann O, Ghosh SS, Polli FE, Reynolds GO, Horn F, Keshavan A, Triantafyllou C, Saygin ZM, Whitfield-Gabrieli S, Hofmann SG, Pollack M, Gabrieli JD. Predicting treatment response in social anxiety disorder from functional magnetic resonance imaging. JAMA Psychiatry. 2013;70:87–97. doi: 10.1001/2013.jamapsychiatry.5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Etkin A, Egner T, Kalisch R. Emotional processing in anterior cingulate and medial prefrontal cortex. Trends Cogn Sci. 2011;15:85–93. doi: 10.1016/j.tics.2010.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Etkin A, Wager TD. Functional neuroimaging of anxiety: a meta-analysis of emotional processing in PTSD, social anxiety disorder, and specific phobia. Am J Psychiatry. 2007;164:1476–1488. doi: 10.1176/appi.ajp.2007.07030504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Evans KC, Wright CI, Wedig MM, Gold AL, Pollack MH, Rauch SL. A functional MRI study of amygdala responses to angry schematic faces in social anxiety disorder. Depress Anxiety. 2008;25:496–505. doi: 10.1002/da.20347. [DOI] [PubMed] [Google Scholar]
- Ewbank MP, Lawrence AD, Passamonti L, Keane J, Peers PV, Calder AJ. Anxiety predicts a differential neural response to attended and unattended facial signals of anger and fear. Neuroimage. 2009;44:1144–1151. doi: 10.1016/j.neuroimage.2008.09.056. [DOI] [PubMed] [Google Scholar]
- Eysenck MW. Anxiety: The cognitive perspective. Hillsdale, NJ: Lawrence Erlbaum Associates; 1992. [Google Scholar]
- First MB, Spitzer RL, Gibbon M, Williams JBW. Structured Clinical Interview for the DSM-IV Axis I Disorders: SCID-I/P, Version 2.0. Biometrics Research Department, New York State Psychiatric Institute; 1997. [Google Scholar]
- Fitzgerald DA, Angstadt M, Jelsone LM, Nathan PJ, Phan KL. Beyond threat: amygdala reactivity across multiple expressions of facial affect. Neuroimage. 2006;30:1441–1448. doi: 10.1016/j.neuroimage.2005.11.003. [DOI] [PubMed] [Google Scholar]
- Fox E, Calder AJ, Mathews A, Yiend J. Anxiety and sensitivity to gaze direction in emotionally expressive faces. Emotion. 2007;7:478–486. doi: 10.1037/1528-3542.7.3.478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Freitas-Ferrari MC, Hallak JEC, Trzesniak C, Filho AS, Machado-de-Sousa JP, Chagas MHN, Nardi AE, Crippa JAS. Neuroimaging in social anxiety disorder: a systematic review of the literature. Prog Neuropsychopharmacol Biol Psychiatry. 2010;34:565–580. doi: 10.1016/j.pnpbp.2010.02.028. [DOI] [PubMed] [Google Scholar]
- Furmark T, Tillfors M, Marteinsdottir I, Fischer H, Pissiota A, Langstrom B, Fredrikson M. Common changes in cerebral blood flow in patients with social phobia treated with citalopram or cognitive-behavioral therapy. Arch Gen Psychiatry. 2002;59:425–433. doi: 10.1001/archpsyc.59.5.425. [DOI] [PubMed] [Google Scholar]
- Fusar-Poli P, Placentino A, Carletti F, Landi P, Allen P, Surguladze S, Benedetti F, Abbamonte M, Gasparotti R, Barale F, Perez J, McGuire P, Politi P. Functional atlas of emotional faces processing: a voxel-based meta-analysis of 105 functional magnetic resonance imaging studies. J Psychiatry Neurosci. 2009;34:418–432. [PMC free article] [PubMed] [Google Scholar]
- Gentili C, Gobbini MI, Ricciardi E, Vanello N, Pietrini P, Haxby JV, Guazzelli M. Differential modulation of neural activity throughout the distributed neural system for face perception in patients with Social Phobia and healthy subjects. Brain Res Bull. 2008;77:286–292. doi: 10.1016/j.brainresbull.2008.08.003. [DOI] [PubMed] [Google Scholar]
- Goldin PR, Manber T, Hakimi S, Canli T, Gross JJ. Neural bases of social anxiety disorder: emotional reactivity and cognitive regulation during social and physical threat. Arch Gen Psychiatry. 2009;66:170–180. doi: 10.1001/archgenpsychiatry.2008.525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gur RC, Sara R, Hagendoorn M, Marom O, Hughett P, Macy L, Turner T, Bajcsy R, Posner A, Gur RE. A method for obtaining 3-dimensional facial expressions and its standardization for use in neurocognitive studies. J Neurosci Methods. 2002;115:137–143. doi: 10.1016/s0165-0270(02)00006-7. [DOI] [PubMed] [Google Scholar]
- Hambrick JP, Turk CL, Heimberg RG, Schneier FR, Liebowitz MR. The experience of disability and quality of life in social anxiety disorder. Depress Anxiety. 2003;18:46–50. doi: 10.1002/da.10110. [DOI] [PubMed] [Google Scholar]
- Hariri AR, Drabant EM, Munoz KE, Kolachana BS, Mattay VS, Egan MF, Weinberger DR. A susceptibility gene for affective disorders and the response of the human amygdala. Arch Gen Psychiatry. 2005;62:146–152. doi: 10.1001/archpsyc.62.2.146. [DOI] [PubMed] [Google Scholar]
- Hariri AR, Tessitore A, Mattay VS, Fera F, Weinberger DR. The amygdala response to emotional stimuli: a comparison of faces and scenes. Neuroimage. 2002;17:317–323. doi: 10.1006/nimg.2002.1179. [DOI] [PubMed] [Google Scholar]
- Heimberg RG. Cognitive-behavioral therapy for social anxiety disorder: current status and future directions. Biol Psychiatry. 2002;51:101–108. doi: 10.1016/s0006-3223(01)01183-0. [DOI] [PubMed] [Google Scholar]
- Heimberg RG, Horner KJ, Juster HR, Safren SA, Brown EJ, Schneier FR, Liebowitz MR. Psychometric properties of the Liebowitz Social Anxiety Scale. Psychol Med. 1999;29:199–212. doi: 10.1017/s0033291798007879. [DOI] [PubMed] [Google Scholar]
- Hofmann SG, Ellard KK, Siegle GJ. Neurobiological correlates of cognitions in fear and anxiety: a cognitive-neurobiological information-processing model. Cogn Emot. 2012;26:282–299. doi: 10.1080/02699931.2011.579414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hofmann SG, Smits JAJ. Cognitive-behavioral therapy for adult anxiety disorders: a meta-analysis of randomized placebo-controlled trials. J Clin Psychiatry. 2008;69:621–632. doi: 10.4088/jcp.v69n0415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hope DA, Heimberg RG, Turk CL. Managing social anxiety: a cognitive-behavioral therapy approach. New York: Oxford University Press; 2006. [Google Scholar]
- Hope DA, Herbert JD, White C. Diagnostic subtype, avoidant personality disorder, and efficacy of cognitive-behavioral group therapy for social phobia. Cognit Ther Res. 1995;19:399–417. [Google Scholar]
- Jones CL, Ward J, Critchley HD. The neuropsychological impact of insular cortex lesions. J Neurol Neurosurg Psychiatry. 2010;81:611–618. doi: 10.1136/jnnp.2009.193672. [DOI] [PubMed] [Google Scholar]
- Kessler RC, Chiu WT, Demler O, Walters EE. Prevalence, severity, and comorbidity of 12-Month DSM-IV disorders in the national comorbidity survey replication. Arch Gen Psychiatry. 2005;62:617–627. doi: 10.1001/archpsyc.62.6.617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kringelbach ML, Rolls ET. The functional neuroanatomy of the human orbitofrontal cortex: evidence from neuroimaging and neuropsychology. Prog Neurobiol. 2004;72:341–372. doi: 10.1016/j.pneurobio.2004.03.006. [DOI] [PubMed] [Google Scholar]
- Lancaster JL, Summerln JL, Rainey L, Freitas CS, Fox PT. The Talairach Daemon, a database server for Talairach Atlas Labels. NeuroImage. 1997;5:S633. [Google Scholar]
- Lancaster JL, Woldorff MG, Parsons LM, Liotti M, Freitas CS, Rainey L, Kochunov PV, Nickerson D, Mikiten SA, Fox PT. Automated Talairach atlas labels for functional brain mapping. Hum Brain Mapp. 2000;10:120–131. doi: 10.1002/1097-0193(200007)10:3<120::AID-HBM30>3.0.CO;2-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lang PJ, Bradley MM. Emotion and the motivational brain. Biol Psychol. 2010;84:437–450. doi: 10.1016/j.biopsycho.2009.10.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lieberman MD, Cunningham WA. Type I and Type II error concerns in fMRI research: re-balancing the scale. Soc Cogn Affect Neurosci. 2009;4:423–428. doi: 10.1093/scan/nsp052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liebowitz MR. Social phobia. Mod Probl Pharmacopsychiatry. 1987;22:141–173. doi: 10.1159/000414022. [DOI] [PubMed] [Google Scholar]
- MacLeod C, Rutherford E, Campbell L, Ebsworthy G, Holker L. Selective attention and emotional vulnerability: assessing the causal basis of their association through the experimental manipulation of attentional bias. J Abnorm Psychol. 2002;111:107–123. [PubMed] [Google Scholar]
- Mattia JI, Heimberg RG, Hope DA. The revised Stroop color-naming task in social phobics. Behav Res Ther. 1993;31:305–313. doi: 10.1016/0005-7967(93)90029-t. [DOI] [PubMed] [Google Scholar]
- Northoff G, Heinzel A, de Greck M, Bermpohl F, Dobrowolny H, Panksepp J. Self-referential processing in our brain—a meta-analysis of imaging studies on the self. Neuroimage. 2006;31:440–457. doi: 10.1016/j.neuroimage.2005.12.002. [DOI] [PubMed] [Google Scholar]
- Öhman A, Mineka S. Fears, phobias, and preparedness: toward an evolved module of fear and fear learning. Psychol Rev. 2001;108:483–522. doi: 10.1037/0033-295x.108.3.483. [DOI] [PubMed] [Google Scholar]
- Ongur D, Ferry AT, Price JL. Architectonic subdivision of the human orbital and medial prefrontal cortex. J Comp Neurol. 2003;460:425–449. doi: 10.1002/cne.10609. [DOI] [PubMed] [Google Scholar]
- Ongur D, Price JL. The organization of networks within the orbital and medial prefrontal cortex of rats, monkeys and humans. Cereb Cortex. 2000;10:206–219. doi: 10.1093/cercor/10.3.206. [DOI] [PubMed] [Google Scholar]
- Otto MW, Pollack MH, Maki KM, Gould RA, Worthington JJ, Smoller JW, Rosenbaum JF. Childhood history of anxiety disorders among adults with social phobia: rates, correlates, and comparisons with patients with panic disorder. Depress Anxiety. 2001;14:209–213. doi: 10.1002/da.1068. [DOI] [PubMed] [Google Scholar]
- Paquette V, Lévesque J, Mensour B, Leroux JM, Beaudoin G, Bourgouin P, Beauregarda M. Change the mind and you change the brain: effects of cognitive-behavioral therapy on the neural correlates of spider phobia. Neuroimage. 2003;18:401–409. doi: 10.1016/s1053-8119(02)00030-7. [DOI] [PubMed] [Google Scholar]
- Paulus MP, Stein MB. An insular view of anxiety. Biol Psychiatry. 2006;60:383–387. doi: 10.1016/j.biopsych.2006.03.042. [DOI] [PubMed] [Google Scholar]
- Paulus MP, Stein MB. Interoception in anxiety and depression. Brain Struct Funct. 2010;214:451–463. doi: 10.1007/s00429-010-0258-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pessoa L, Kastner S, Ungerleider LG. Attentional control of the processing of neutral and emotional stimuli. Brain Res Cogn Brain Res. 2002;15:31–45. doi: 10.1016/s0926-6410(02)00214-8. [DOI] [PubMed] [Google Scholar]
- Phan KL, Coccaro EF, Angstadt M, Kreger KJ, Mayberg HS, Liberzon I, Stein MB. Corticolimbic brain reactivity to social signals of threat before and after sertraline treatment in generalized social phobia. Biol Psychiatry. 2013;73:329–336. doi: 10.1016/j.biopsych.2012.10.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phan KL, Fitzgerald DA, Nathan PJ, Tancer ME. Association between amygdala hyperactivity to harsh faces and severity of social anxiety in generalized social phobia. Biol Psychiatry. 2006;59:424–429. doi: 10.1016/j.biopsych.2005.08.012. [DOI] [PubMed] [Google Scholar]
- Phillips ML. Understanding the neurobiology of emotion perception: implications for psychiatry. Br J Psychiatry. 2003;182:190–192. doi: 10.1192/bjp.182.3.190. [DOI] [PubMed] [Google Scholar]
- Pishyar R, Harris LM, Menzies RG. Responsiveness of measures of attentional bias to clinical change in social phobia. Cogn Emot. 2008;22:1209–1227. [Google Scholar]
- Posner MI, Petersen SE. The Attention System of the human brain. Annu Rev Neurosci. 1990;13:25–42. doi: 10.1146/annurev.ne.13.030190.000325. [DOI] [PubMed] [Google Scholar]
- Rolls ET. The functions of the orbitofrontal cortex. Brain Cogn. 2004;55:11–29. doi: 10.1016/S0278-2626(03)00277-X. [DOI] [PubMed] [Google Scholar]
- Rothkirch M, Schmack K, Schlagenhauf F, Sterzer P. Implicit motivational value and salience are processed in distinct areas of orbitofrontal cortex. Neuroimage. 2012;62:1717–1725. doi: 10.1016/j.neuroimage.2012.06.016. [DOI] [PubMed] [Google Scholar]
- Sabatinelli D, Keil A, Frank DW, Lang PJ. Emotional perception: correspondence of early and late event-related potentials with cortical and subcortical functional MRI. Biol Psychol. 2013;92:513–519. doi: 10.1016/j.biopsycho.2012.04.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Safren SA, Heimberg RG, Brown EJ, Holle C. Quality of life in social phobia. Depress Anxiety. 1996/1997;4:126–133. doi: 10.1002/(SICI)1520-6394(1996)4:3<126::AID-DA5>3.0.CO;2-E. [DOI] [PubMed] [Google Scholar]
- Schienle A, Schäfer A, Hermann A, Rohrmann S, Vaitl D. Symptom provocation and reduction in patients suffering from spider phobia: an fMRI study on exposure therapy. Eur Arch Psychiatry Clin Neurosci. 2007;257:486–493. doi: 10.1007/s00406-007-0754-y. [DOI] [PubMed] [Google Scholar]
- Schneier FR, Heckelman LR, Garfinkel R, Campeas R, Fallon BA, Gitow A, Street L, Del Bene D, Leibowitz MR. Functional impairment in social phobia. J Clin Psychiatry. 1994;55:322–331. [PubMed] [Google Scholar]
- Schneier FR, Johnson J, Hornig CD, Liebowitz MR, Weissman MM. Social Phobia. Comorbidity and morbidity in an epidemiologic sample. Arch Gen Psychiatry. 1992;49:282–288. doi: 10.1001/archpsyc.1992.01820040034004. [DOI] [PubMed] [Google Scholar]
- Schoenbaum G, Roesch M. Orbitofrontal cortex, associative learning, and expectancies. Neuron. 2005;47:633–636. doi: 10.1016/j.neuron.2005.07.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shin LM, Liberzon I. The neurocircuitry of fear, stress, and anxiety disorders. Neuropsychopharmacology. 2010;35:169–191. doi: 10.1038/npp.2009.83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stein MB, Goldin PR, Sareen J, Zorrilla LT, Brown GG. Increased amygdala activation to angry and contemptuous faces in generalized social phobia. Arch Gen Psychiatry. 2002;59:1027–1034. doi: 10.1001/archpsyc.59.11.1027. [DOI] [PubMed] [Google Scholar]
- Straube T, Glauer M, Dilger S, Mentzel HJ, Miltner WHR. Effects of cognitive-behavioral therapy on brain activation in specific phobia. Neuroimage. 2006;29:125–135. doi: 10.1016/j.neuroimage.2005.07.007. [DOI] [PubMed] [Google Scholar]
- Straube T, Kolassa IT, Glauer M, Mentzel HJ, Miltner WHR. Effect of task conditions on brain responses to threatening faces in social phobics: an event-related functional magnetic resonance imaging study. Biol Psychiatry. 2004;56:921–930. doi: 10.1016/j.biopsych.2004.09.024. [DOI] [PubMed] [Google Scholar]
- Straube T, Mentzel HJ, Miltner WHR. Common and distinct brain activation to threat and safety signals in social phobia. Neuropsychobiology. 2005;52:163–168. doi: 10.1159/000087987. [DOI] [PubMed] [Google Scholar]
- Tillfors M, Furmark T, Marteinsdottir I, Fredrikson M. Cerebral blood flow during anticipation of public speaking in social phobia: a PET study. Biol Psychiatry. 2002;52:1113–1119. doi: 10.1016/s0006-3223(02)01396-3. [DOI] [PubMed] [Google Scholar]
- Tzourio-Mazoyer N, Landeau B, Papathanassiou D, Crivello F, Etard O, Delcroix N, Mazoyer B, Joliot M. Automated anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI single-subject brain. Neuroimage. 2002;15:273–289. doi: 10.1006/nimg.2001.0978. [DOI] [PubMed] [Google Scholar]
- Wells A, Matthews G. Attention and emotion: A clinical perspective. Hillsdale, NJ: Lawrence Erlbaum Associates; 1994. [Google Scholar]
- Whalen PJ. Fear, vigilance, and ambiguity: initial neuroimaging studies of the human amygdala. Curr Dir Psychological Sci. 1998;7:177–188. [Google Scholar]
- Whalen PJ, Shin LM, McInerney SC, Fischer H, Wright CI, Rauch SL. A functional MRI study of human amygdala responses to facial expressions of fear versus anger. Emotion. 2001;1:70–83. doi: 10.1037/1528-3542.1.1.70. [DOI] [PubMed] [Google Scholar]
- Williams JMG, Watts FN, MacLeod C, Mathews A. Cognitive psychology and emotional disorders. Chichester, England: Wiley; 1988. [Google Scholar]