

ABSTRACT
In this Commentary, we discuss two sets of genetically encoded molecular tools that have significantly enhanced our ability to observe and manipulate complex biochemical processes in their native context and that have been essential in deepening our molecular understanding of how intracellular signaling networks function. In particular, genetically encoded biosensors are widely used to directly visualize signaling events in living cells, and we highlight several examples of basic biosensor designs that have enabled researchers to capture the spatial and temporal dynamics of numerous signaling molecules, including second messengers and signaling enzymes, with remarkable detail. Similarly, we discuss a number of genetically encoded biochemical perturbation techniques that are being used to manipulate the activity of various signaling molecules with far greater spatial and temporal selectivity than can be achieved using standard pharmacological or genetic techniques, focusing specifically on examples of chemically driven and light-inducible perturbation strategies. We then describe recent efforts to combine these diverse and powerful molecular tools into a unified platform that can be used to elucidate the molecular details of biological processes that may potentially extend well beyond the realm of signal transduction.
KEY WORDS: Biosensors, FRET, Live cell imaging, Signaling, Targeted perturbations
Introduction
Inter- and intracellular communication is the key for living systems to cope with an ever-changing environment. Effective cell communication utilizes highly regulated signal transduction networks that convert input signals into specific cellular responses. To achieve these specific responses, each network comprises numerous signaling molecules, such as second messengers, kinases, phosphatases, ligands, etc., whose dynamics are in turn subject to rigorous spatial and temporal control (Kholodenko et al., 2010). Various molecular mechanisms are put in place to maintain the tight spatiotemporal control of signal transduction, and these frequently involve the compartmentalization of signaling molecules. For example, scaffolding proteins play a major role in partitioning signaling enzymes into distinct macromolecular complexes and in directing these nanomachines to different subcellular locations (reviewed in Pan et al., 2012; Scott and Pawson, 2009). In particular, various components of the 3′,5′-cyclic adenosine monophosphate (cAMP)-dependent protein kinase (PKA)-signaling pathway are known to be assembled onto A-kinase anchoring proteins (AKAPs) that are targeted throughout the cell (Wong and Scott, 2004), and a number of different scaffold proteins are involved in regulating mitogen-activated protein kinase (MAPK)-signaling pathways (Brown and Sacks, 2009; Kolch, 2005). Anchored signaling enzymes can further impose spatial control over diffusible signaling molecules, giving rise to dynamic gradients and microdomains of signaling activity (Dehmelt and Bastiaens, 2010; Kholodenko et al., 2010).
Thus, in order to dissect the molecular mechanisms that are involved in signal transduction, it is imperative to study these dynamic processes in the native context of a living cell, tissue or organism, as this ensures that the entire signaling network, together with its regulatory mechanisms, remains intact. Classic methods of studying signal transduction, which usually involve cell fixation or lysis, provide only limited temporal and spatial resolution, and often fail to capture biologically relevant single-cell behaviors such as oscillations owing to their reliance on cell populations. Similarly, classic cell perturbation strategies often lack the spatial and temporal resolution that are needed to investigate the relationship between specific signaling molecules. Many genetically encoded probes have, therefore, been developed to help to directly visualize and capture dynamic signaling processes in living biological systems, as well as to directly manipulate signaling molecules with greater precision. Because they are genetically encoded, these tools can be introduced into living systems as DNA and undergo de novo protein synthesis, followed by subcellular targeting through the use of appended localization signals. Furthermore, when these two sets of genetically encoded molecular tools are combined, they provide a powerful platform for dissecting the roles of specific signaling molecules in guiding cellular behaviors.
In this Commentary, we focus on two complementary types of genetically encodable molecular probe that can be utilized to either monitor or perturb the behaviors of various signaling molecules. To begin with, we will present an overview of the main varieties of genetically encoded, fluorescent protein (FP)-based biosensors and illustrate how different biosensor designs can be used to monitor various signaling events. This is followed by specific examples of the two main classes of molecular tools – chemically inducible perturbation systems and light-inducible perturbation systems – that enable the targeted manipulation of discrete biochemical processes within signaling pathways in living cells. We then discuss recent examples from the literature in order to highlight specific instances in which the combined application of these complementary technologies helped to provide new insights into the regulation and function of intracellular signaling pathways. Finally, we offer some of our thoughts regarding the future prospects of these exciting techniques.
Genetically encoded biosensors to monitor signaling dynamics
Our understanding of how specificity in signaling is achieved has been greatly enhanced by our ability to study real-time signaling dynamics in living cells. Early on, optical detection of cellular signaling was often accomplished using chemical dyes. For instance, upon its introduction in the 1980s, the fluorescent dye and Ca2+ indicator Fura-2 helped revolutionize the study of Ca2+ signaling and remains one of the most popular fluorescent probes for live cell imaging. Over the past two decades, the advent of FP technology, in conjunction with advances in microscopy, has enabled the visualization of a much wider range of signaling events with high spatiotemporal resolution (Box 1). A plethora of FP-based biosensors has been generated to monitor a variety of signaling molecules, such as cyclic nucleotide messengers, ligands, phosphoinositides, kinases, phosphatases, GTPases and many others (reviewed in Balla, 2009; Newman et al., 2011; Tantama et al., 2012).
Box 1. Using fluorescence in order to probe cellular function.
Methods that can be used to chemically modify purified antibodies without altering their reactivity towards antigens were first demonstrated in the 1930s (discussed in Coons, 1971). At the time, Albert Coons was testing for streptococcal antigens in lesions of rheumatic fever and generated the first fluorescein-conjugated antibodies (Coons, 1971; Coons et al., 1942). The field of immunology was soon ablaze with immunofluorescence. Initially used to visualize disease-related antigens (Mellors, 1968) and for diagnostic purposes (Nelson et al., 1964), immunofluorescence later became a powerful tool for localizing native cellular components (see Bloom et al., 1973). Eventually, researchers began to directly label purified cellular proteins with fluorescent tags (Taylor and Wang, 1978; Taylor et al., 1984). These fluorescent analogs could be introduced into living cells, making it possible to continuously record the dynamics of labeled molecules, rather than rely on static snapshots of fixed cells.
Scientists also began synthesizing dyes whose fluorescent properties were sensitive to particular conditions in the cell, such as pH, membrane potential or ion concentrations (reviewed in Tsien, 1989). Like fluorescent analogs, fluorescent chemical indicators could be introduced into living cells to visualize dynamic processes. However, in contrast to the aforementioned techniques, in which fluorescence is simply a passive marker, fluorescence plays an active and integral role in the function of these indicators. For example, the Ca2+ indicator fura-2 emits green fluorescence (∼520 nm) that is stimulated by 380 nm light in the Ca2+-free state and by 340 nm light in the Ca2+-bound state; the ratio of emission intensity from the excitation at these two wavelengths serves to indicate Ca2+ concentration (Grynkiewicz et al., 1985).
When FP technology first emerged (Tsien, 1998; Zhang, 2009), FPs were also used as passive markers of protein localization. Given their genetic encodability, along with the relative ease of recombinant DNA technology, FP tagging soon replaced the use of fluorescent analogs (Whitaker, 2000). Meanwhile, improved spectrally distinct FPs (Day and Davidson, 2009; Sample et al., 2009; Shaner et al., 2005) enabled the construction of FRET-based biosensors, with changes in FRET actively reporting on biochemical events in cells. The first such biosensor, the Ca2+ probe Cameleon, featured CaM and the M13 peptide sandwiched between either GFP and BFP or CFP and YFP (Miyawaki et al., 1997). Reporters for kinase activity were soon developed (Ting et al., 2001; Zhang et al., 2001) releasing a flood of biosensors, whose modular design is adaptable to detecting a seemingly endless variety of biochemical processes (Mehta and Zhang, 2011; Newman et al., 2011).
All biosensors consist of a sensing element that is coupled to a reporting unit. The sensing element usually comprises endogenous proteins or de novo peptide sequences that target the biochemical activity of interest, whereas the reporting unit consists of one or more fluorescent proteins, wherein fluorescence serves as an indicator for the biochemical status of the sensing element. The simplest biosensor design consists of the fusion of a single FP to a protein or protein domain of interest. Because FPs typically do not encode any innate localization, the distribution of the fluorescent signal can serve as a proxy for the localization of the target protein. However, appropriate controls are always needed to eliminate potential artifacts of FP fusions, as some FPs may spontaneously accumulate in the nucleus (Seibel et al., 2007) or have a tendency to form aggregates in certain cell types. In addition, the size of an FP can sometimes sterically hinder the formation of signaling complexes. Smaller fluorescent tags could thus be used when size matters (Fessenden and Mahalingam, 2013; Jung et al., 2013; Zürn et al., 2010). Alternatively, an FP can be linked to a sensing domain that specifically binds to a molecule of interest that typically resides in a cellular membrane, such as the pleckstrin homology (PH) domain of Akt, which recognizes 3′ phosphoinositides in the plasma membrane (Gray et al., 1999) (Fig. 1A). The production or degradation of the target molecule results in the translocation of these probes to or from the membrane, which can be measured as a change in the fluorescence intensity at the compartment of interest.
Fig. 1.
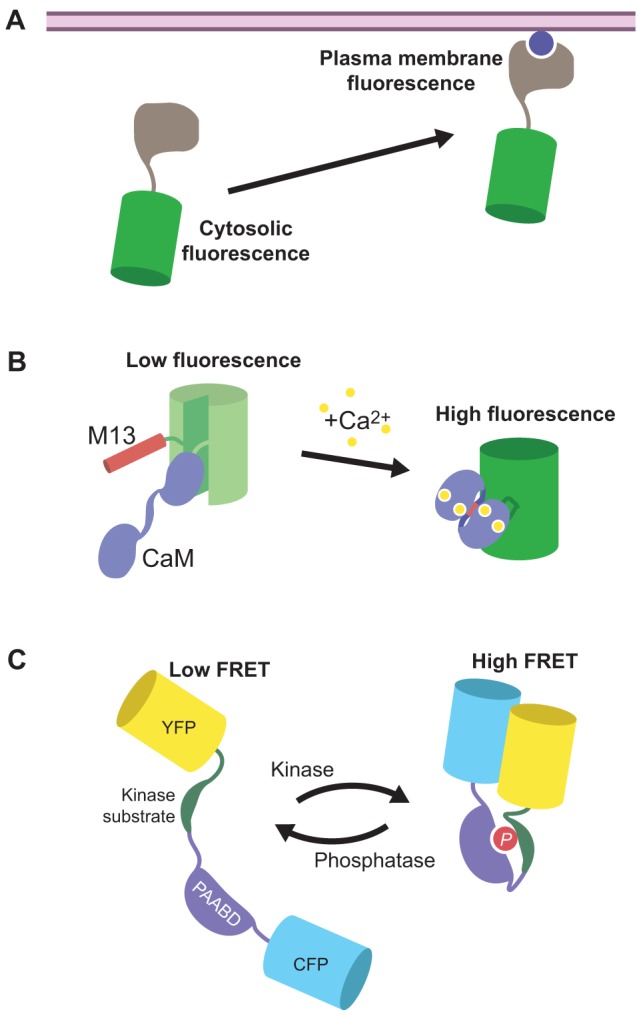
Different types of FP-based biosensor. (A) The fusion of an FP such as GFP (green) to a specific binding domain (gray) can be used to report on the production of certain signaling molecules (blue circle), for example through the redistribution of the probe – and thus fluorescence – from the cytosol (left) to the plasma membrane (right). (B) The Ca2+ sensor GCaMP consists of a molecular switch that contains calmodulin (CaM) and M13 inserted into a circularly permuted GFP (green), in which the native N- and C-termini of GFP are linked together, and new termini are generated from within the core β-barrel structure of GFP; the addition of Ca2+ causes CaM to bind to M13, which leads to increased GFP fluorescence. (C) FRET-based reporter for kinase activity. A kinase-specific substrate peptide and a phosphoamino-acid-binding domain (PAABD) are sandwiched between two FPs that can undergo FRET (e.g. CFP and YFP). Phosphorylation (P) of the substrate through the cognate kinase induces binding of the substrate peptide by the PAABD, resulting in a conformational rearrangement that produces a change in FRET.
Another class of single-FP biosensors exploits the photophysical properties of the chromophore itself in order to report on physiological events. In this case, the sensing unit comprises a molecular switch that is inserted within the fluorescent protein. A popular reporter of this type is derived from the fusion of green fluorescent protein (GFP), calmodulin (CaM) and M13 into GCaMP, an intensity-based Ca2+ indicator (Nakai et al., 2001) (Fig. 1B). GCaMP incorporates a circularly permuted variant of GFP, in which the native N- and C-termini of GFP are linked together and new termini are generated from within the core β-barrel structure of GFP. The Ca2+-binding protein CaM is fused to the new C-terminus, and the CaM-binding peptide M13 is fused to the new N-terminus, thereby serving as the molecular switch. Crystal structure studies of this reporter have shown that in the Ca2+-free state, the chromophore is exposed to the solvent and, thus, exists in its protonated, non-fluorescent state. Upon Ca2+ binding, the molecular switch undergoes a conformational change, partially blocking access of the solvent to the chromophore and resulting in increased fluorescence intensity from the non-protonated fluorescent species (Akerboom et al., 2009). Recently, the repertoire of intensiometric Ca2+ indicators has been expanded to include FPs of different hues ranging from blue to red (Akerboom et al., 2013; Zhao et al., 2011), thus providing great promise for multicolor imaging with other FP-based biosensors.
The largest class of FP biosensors features a generalized, modular design, wherein the molecular switch is sandwiched between a pair of FPs (Fig. 1C) that are capable of undergoing Förster resonance energy transfer (FRET) (for review, see Campbell, 2009; Ibraheem and Campbell, 2010; Newman et al., 2011; Zaccolo, 2004). FRET is a biophysical phenomenon that is characterized by the non-radiative transfer of excited-state energy from a donor fluorophore to an acceptor fluorophore through dipole-dipole coupling. Efficient energy transfer requires a sufficient amount of overlap between the donor emission spectrum and the acceptor absorption spectrum, as well as the spatial proximity (<10 nm) between the two fluorophores and their relative orientation with respect to each other. Thus, when the signaling activity of interest is turned on, the molecular switch undergoes a conformational change that alters the distance between and/or relative orientation of the two fluorophores, resulting in a change in FRET.
The modular nature of these biosensors facilitates the incorporation of diverse molecular switch designs, making this a broad and versatile class of reporters (Newman et al., 2011). For example, multiple FRET-based GTPase biosensors have been generated by tethering a GTPase of interest to a particular effector protein or domain and sandwiching this molecular switch between two FPs (Kiyokawa et al., 2011; Mochizuki et al., 2001; Nakamura et al., 2005). In the GDP-bound state, the GTPase is inactive and does not interact with the effector. However, when GDP is exchanged for GTP, the GTPase becomes activated and binds the effector, altering the conformation of the molecular switch and producing a FRET change. Similarly, kinase activity reporters (KARs) feature a molecular switch that is composed of a kinase-specific substrate tethered to a phosphoamino-acid-binding domain (PAABD), which is inserted between the two FPs (Fig. 1C) (Fuller et al., 2008; Gavet and Pines, 2010; Harvey et al., 2008; Violin et al., 2003; Zhang et al., 2001). Phosphorylation of the substrate by the cognate kinase induces binding between substrate and PAABD, thereby promoting a conformational rearrangement and a change in FRET.
An important consideration when using FP-based biosensors is the potential for these probes to perturb the processes they are used to monitor (Miyawaki, 2003), particularly when overexpressed reporters compete with endogenous targets for a limited supply of signaling molecules or when the sensing domain of a reporter contains an active enzyme. It is, therefore, essential to introduce as little of the biosensor as possible into the cells and to design controls to verify that the system is not adversely affected by the biosensor. Controls should also be performed to verify biosensor responses. For instance, controlling for correct interactions of the sensors with upstream regulators, their insertion into the correct activating compartments, and the proper recycling of sensors back into inactive compartments are all crucial to reading out physiologically relevant GTPase activation dynamics. Similarly, it is important to keep in mind that biochemical pathways are often regulated by the action of opposing processes. For example, the FRET response from a KAR can be reversed upon dephosphorylation of the substrate peptide through endogenous phosphatases. Thus, it should be noted that these biosensors do not strictly report kinase activity per se so much as they reflect how the balance between kinase and phosphatase activity – itself a crucial aspect of the native signaling environment – is influencing the action of a particular kinase. These dynamic changes in the steady-states of biochemical reactions are central to the processing of cellular information (Dehmelt and Bastiaens, 2010), and biosensors allow us to directly visualize these dynamics and tease apart the underlying molecular mechanisms.
Genetically encoded tools that can be used to perturb biochemical events in living cells
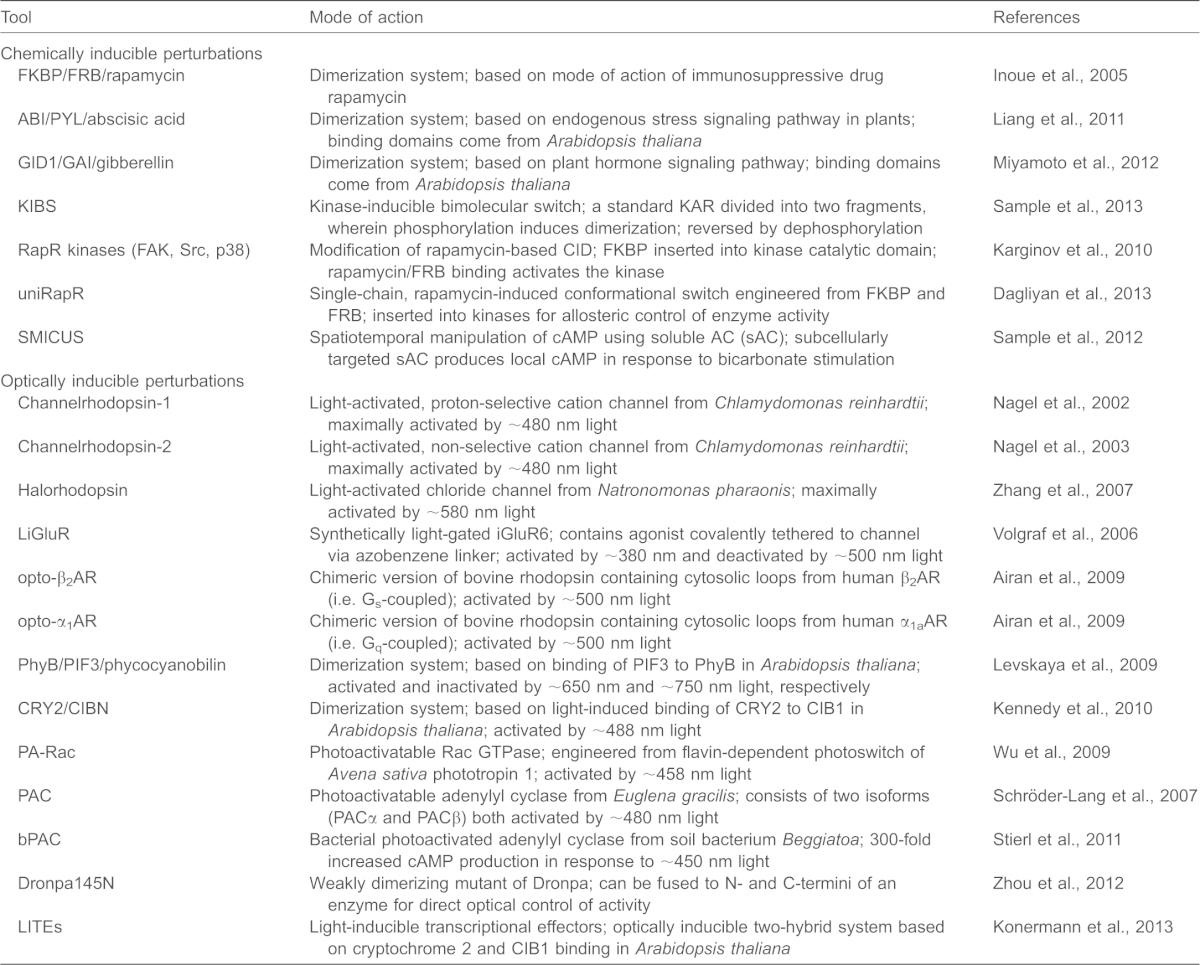
Dissecting causal relationships between specific signaling molecules within a dynamic and interconnected signaling network requires not only tools to monitor but also methods to perturb signaling events with spatial and temporal precision. Although pharmacological and genetic perturbations can be specific, only a limited degree of subcellular spatial control can be achieved when using these approaches. Below, we discuss examples of the two main strategies for targeted perturbation that are currently available, and that are based on either chemically or optically inducible switches. A list of currently available perturbation tools is provided in Table 1.
Table 1. Genetically encoded tools that can be used to perturb biochemical events in living cells.
Chemically inducible perturbation tools
Many proteins are controlled by oligomerization. Exogenously triggered dimerization is a very effective strategy for precisely controlling the activation of discrete biochemical pathways (Crabtree and Schreiber, 1996), and chemically inducible dimerization (CID) has been used to control protein activity for many years (reviewed in DeRose et al., 2013). This approach involves generating chimeric proteins that are based on a pair of binding domains that specifically interact with one another in the presence of the immunosuppressive compound rapamycin. These systems are easily introduced into living cells because the protein components are genetically encoded and because rapamycin can easily permeate cellular membranes. CID is generally used to perturb local enzyme activity through a translocation-based approach, wherein the addition of rapamycin triggers the relocalization of an enzyme of interest from a substrate-poor region to a substrate-rich region (Fig. 2A). For example, constitutively active and dominant-negative forms of the GTPase Rac were inducibly translocated to the plasma membrane in order to rapidly and selectively perturb local Rac signaling (Inoue and Meyer, 2008; Inoue et al., 2005). Similarly, the induced translocation of a yeast inositol phosphatase, as well as phosphoinositide 3-kinase (PI3K), was used to perturb plasma membrane phosphoinositide signaling (Inoue and Meyer, 2008; Suh et al., 2006). In addition, the focal activation of caged rapamycin can also be used to achieve an even more spatially localized control of enzyme activity (Karginov et al., 2011; Umeda et al., 2011).
Fig. 2.
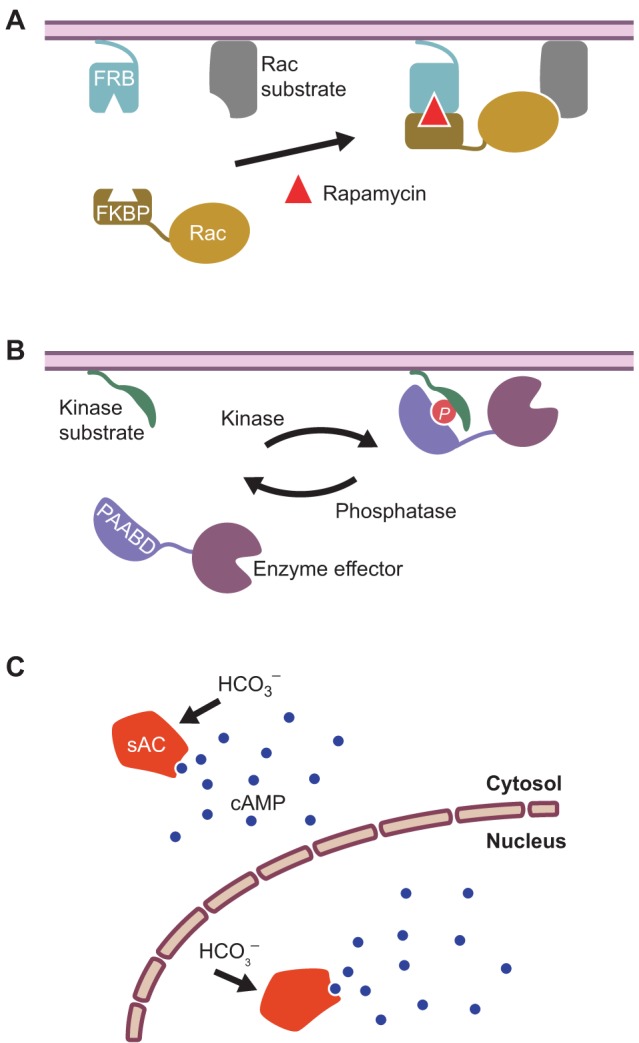
Examples of chemically inducible perturbation tools. (A) A chemically inducible dimerization system utilizing a pair of rapamycin-binding domains. One binding domain, derived from the FKBP-rapamycin binding (FRB) protein, is tethered to the plasma membrane and the other, derived from the FK506-binding protein (FKBP), is fused to an enzyme effector such as the GTPase Rac in the cytosol. Rapamycin treatment induces the translocation of the enzyme into the vicinity of its substrate, thereby promoting activity. (B) A kinase-inducible bimolecular switch. Here, a membrane-tethered, kinase-specific substrate peptide is shown, together with a phosphoamino-acid-binding domain (PAABD) that is coupled to an enzyme effector in the cytosol. Phosphorylation (P) of the substrate peptide reversibly recruits the PAABD–enzyme fusion protein to the membrane, where the enzyme is active. (C) Spatiotemporal manipulation of cAMP using soluble adenylyl cyclase (sAC). sAC is exclusively stimulated by bicarbonate (HCO3–) and can be expressed in different subcellular compartments to selectively produce cAMP in different regions of the cell, for instance in the cytosol or nucleus.
More recently, a number of alternative CID systems have also been developed that are orthogonal to rapamycin-based CID, in that they can be used in cells at the same time as rapamycin without producing unwanted cross-talk. For example, two CID systems have been constructed on the basis of hormone signaling in plants; one involves the dimerization of the proteins gibberellin insensitive dwarf 1 (GID1) and gibberellin insensitive (GAI) upon the addition of the hormone gibberellin (Miyamoto et al., 2012); the other utilizes the plant hormone abscisic acid (ABA) to induce dimerization between the PYR1-like (PYL) and ABA insensitive 1 (ABI1) proteins (Liang et al., 2011). In another variation, a kinase-inducible biomolecular switch (KIBS) was used to induce dimerization. Here, the molecular switch consists of two fragments – a kinase-specific substrate and a PAABD – that are induced to associate upon phosphorylation (Fig. 3B) (Sample et al., 2013). Because dephosphorylation can then trigger the dissociation of the switch, it may be possible to dynamically control local enzyme activity using this system.
Fig. 3.
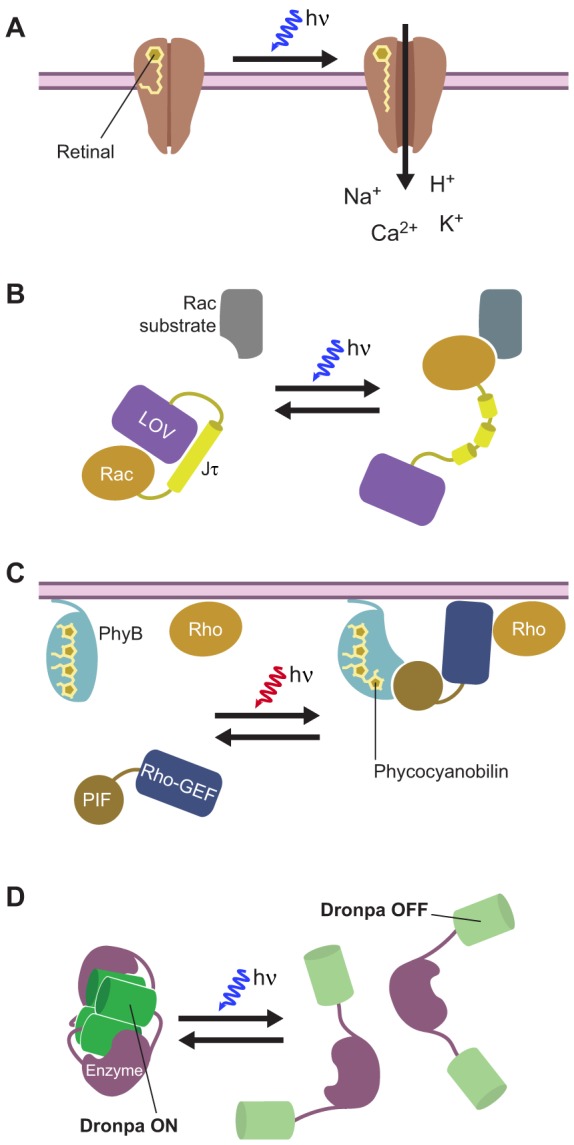
Examples of optically inducible perturbation tools. (A) The light-induced ion channel channel rhodopsin. The associated retinal moiety photoisomerizes upon illumination with blue light, which promotes channel opening and subsequent ion influx. (B) Photoactivatable Rac. A constitutively active Rac isoform is fused to the light–oxygen–voltage (LOV) domain and Jα helix from Avena sativa phototropin 1, which results in an intramolecular complex that inhibits Rac activity. Upon illumination with blue light, a reversible conformational change disrupts the Jα helix and induces Rac activity. (C) Light-inducible control of protein-protein interactions using phytochrome B (PhyB) and a phytochrome-interacting factor (PIF). Phycocyanobilin (PCB) is covalently bound to the PhyB protein, which is tethered to the plasma membrane. Upon illumination with red light, PCB undergoes a photoisomerization reaction that results in the binding of PhyB to the PIF. This can be used to control the translocation of a PIF-tethered effector enzyme from the cytosol to the plasma membrane. In this example, a guanine nucleotide exchange factor for a Rho-family GTPase (Rho-GEF) is tethered to PIF and used to activate a Rho-family GTPase (Rho) in the plasma membrane. (D) Optical control of enzyme activity with a photoswitchable FP. A weakly oligomerizing Dronpa mutant (Dronpa145N; green) is tethered to the N- and C-termini of an effector enzyme; in the fluorescent state (Dronpa ON), two of the fusion proteins associate to form a Dronpa145N tetramer, thereby blocking enzyme activity. Switching Dronpa145N into the non-fluorescent state (Dronpa OFF) by using blue light causes dissociation of the tetramer and releases the active enzyme.
One complication of CID is that the enzyme that is to be translocated is present in the cytosol prior to dimer formation and may, therefore, exhibit ectopic activity. Other modifications of CID have thus been introduced to reduce this background activity, such as by initially sequestering the enzyme into an inactive pool elsewhere within the cell (DeRose et al., 2013). It has also proven possible to adapt CID for the direct allosteric regulation of enzymes. For example, the rapamycin-induced activation of the focal adhesion kinase (FAK), as well as the Src and p38 MAPKs, was recently achieved by inserting one of the aforementioned rapamycin-binding domains into the catalytic domain of each kinase (Karginov et al., 2010). Further engineering of this system resulted in the incorporation of both rapamycin-binding domains into a rationally designed unimolecular switch, dubbed uniRapR (Dagliyan et al., 2013). This modular conformational switch has also been successfully inserted into Src, enabling the direct and specific allosteric control of Src kinase activity both in vitro and in vivo (Dagliyan et al., 2013).
Whereas CID systems are generalizable and can be adapted for the control of several different enzymes and signaling pathways, other chemically induced perturbation systems are more narrowly applicable and are tailored to specific signaling pathways. For instance, we recently described a novel method for the spatiotemporal manipulation of cAMP using soluble adenylyl cyclase (SMICUS) that can be used to control the duration, magnitude and location of a cAMP signal (Sample et al., 2012). Classic techniques for manipulating cAMP concentrations usually produce large-scale variations in cAMP concentrations throughout the cell; however, our system utilizes a truncated version of soluble adenylyl cyclase (sAC) that retains the ability to catalytically produce cAMP from ATP and can be localized to different parts of the cell through tagging with endogenous targeting motifs. Furthermore, sAC is exclusively activated by CHO3− and not by direct activators of transmembrane adenylyl cyclase or ligands for G-protein-coupled receptors (GPCRs). Therefore, sAC serves as an orthogonal source to produce a local cAMP signal within a designated cellular space.
Light-inducible perturbation tools
In contrast to the more passive role light plays in monitoring signaling dynamics, it can also provide a powerful, non-invasive and, in most cases, orthogonal means for targeted perturbations. The high resolution with which light can be manipulated, both spatially and temporally, enables the investigation of rapid processes that either occur specifically in a subcellular compartment or in a subset of cells. Several techniques have been developed to harness light to control signaling processes (Gorostiza and Isacoff, 2008). A number of photoactivatable proteins, both naturally occurring and engineered, are available that provide optical control of channel activity, enzyme activity, and protein-protein interaction.
Naturally occurring, photoisomerizing chromophores can rapidly and reversibly switch the activation state of certain proteins. A classic example is the chromophore retinal, which binds the opsin proteins that are used in visual transduction in the eye. Certain recently discovered, naturally light-driven bacterial opsins, such as channelrhodopsin-2 (ChR2) (Nagel et al., 2003) and halorhodopsin (NpHR) (Zhang et al., 2007), represent new and useful members of the opsin family. Upon absorption of a photon, retinal isomerizes and triggers a set of structural changes that lead to channel opening, ion transport or interaction with downstream transducers (Fig. 3A) (Nagel et al., 2003; Nagel et al., 2005; Zhang et al., 2011). A modified ionotropic glutamate receptor (iGluR) has also been engineered that can be turned on and off by irradiation with different wavelengths of light, thereby functioning as both a light- and ligand-gated ion channel (Volgraf et al., 2006). Both of these approaches utilize light to drive ion channel activity. Similarly, the optogenetic control of biochemical pathways has been achieved by generating chimeras between vertebrate rhodopsin (vRh) and GPCRs (Airan et al., 2009; Kim et al., 2005). Here, the intracellular loops of vRh were replaced with those from the β2- adrenergic receptor to activate cAMP/PKA signaling through the heterotrimeric G protein Gs α subunit (Airan et al., 2009; Kim et al., 2005), or from the α1a- adrenergic receptor to activate phospholipase C (PLC) and Ca2+ signaling through the Gq α-subunit (Airan et al., 2009). These chimeric receptors were designated OptoXRs. In addition, the photoactivatable adenylyl cyclases, which are derived from light-sensitive adenylyl cyclases found in Euglena gracilis (Schröder-Lang et al., 2007) and certain soil bacteria (Beggiatoa spp.) (Stierl et al., 2011), can serve as alternatives to chemically activated sAC (see above).
An example of the rapid and reversible control of an enzyme using light is the fusion of the GTPase Rac1 to the photoreactive light-oxygen-voltage (LOV) domain and the C-terminal helical extension (Jα) from phototropin (Fig. 3B). In the dark, the flavin-binding LOV domain interacts with the Jα extension, occluding the effector-binding domain of Rac1. Illumination using light with a wavelength of 458 or 473 nm results in the formation of a covalent bond between a cysteine residue in the LOV domain and the associated flavin chromophore. This induces conformational changes that lead to the dissociation and unwinding of the Jα helix and, thus, opens up the Rac1 interaction domain for binding to effector molecules (Wu et al., 2009). This photoactivatable Rac variant has been utilized to study the role of Rac in cell migration in both Drosophila and Xenopus transgenic animals (Wang et al., 2010; Yoo et al., 2010).
Light can also be exploited to induce protein-protein interactions, which can be used to drive the translocation of proteins from one subcellular compartment to another. For instance, in plants, many light-sensitive processes are mediated by photoreceptive signaling proteins called phytochromes. These associate covalently with the chromophore phycocyanobilin (PCB), which photoisomerizes between two conformational states: Pr (red absorbing) and Pfr (far-red absorbing). Upon stimulation with red light, the phytochrome protein binds directly to its effector phytochrome interaction factor 3 (PIF3), and the Phy-PIF photoswitchable membrane recruitment system has been developed based on this interaction (Fig. 3C) (Levskaya et al., 2009). In this example, PhyB is tethered to the plasma membrane and a domain derived from the N-terminus of PIF6 is fused to a GEF that acts specifically on Rho-family GTPases. Irradiation with red light (650 nm) results in the isomerization of the chromophore to the Pfr state, thereby inducing translocation of the PIF-bound GEF to the plasma membrane and activation of the corresponding G-protein (Levskaya et al., 2009). In a similar concept, light-induced binding of the photolyase homology region (PHR) of cryptochrome 2 (CRY2) to the N-terminal portion of the transcription factor CIB1 has been exploited to control protein-protein interactions (Kennedy et al., 2010) and phosphoinositide metabolism (Idevall-Hagren et al., 2012).
Fluorescent proteins themselves can also be engineered into perturbation tools. Recently, it was shown that the reversibly photoswitchable GFP variant Dronpa can be used to regulate enzyme activity. Dronpa fluorescence is switched on by illumination with UV light (e.g. 405 nm) and switched off by illumination with blue light (e.g. 515 nm). Like most FPs, Dronpa is derived from a tetrameric parent molecule. The β strand 7 of Dronpa, which forms part of the dimer interface in the parent protein, becomes flexible in response to off-switching, and a weakly multimeric Dronpa mutant that contains a Lys-to-Asn substitution in β strand 7 (Dronpa145N) was shown to undergo reversible oligomerization in response to on- and off-switching (Zhou et al., 2012). As a proof of principle, the authors of this study then use this characteristic to enable the light-induced control of Cdc42 activity by fusing Dronpa145N to the N- and C-termini of the Cdc42 GEF intersectin. In the dark, the Dronpa subunits from a pair of chimeric proteins associate to form a Dronpa tetramer, as illustrated in Fig. 3D; this blocks its interaction with Cdc42 and produces ‘caged’ intersectin. Switching Dronpa off using blue light then dissociates the Dronpa tetramer, leading to the uncaging of intersectin and the subsequent activation of Cdc42 (Fig. 3D). The same effect can also be achieved when one of the Dronpa145N domains is substituted with wild-type Dronpa; in this case, the enzyme of interest is caged by an intramolecular Dronpa dimer, rather than by an intermolecular Dronpa tetramer (Zhou et al., 2012).
Genetically encoded biosensors and perturbation tools combine to provide new biological insights
As discussed above, genetically encoded molecular tools can facilitate both the visualization and perturbation of a variety of biochemical events within living cells. Combining perturbation tools with live cell imaging is increasingly used to delve into the organization of signaling networks and to uncover the molecular mechanisms that allow cells to integrate diverse inputs and achieve specific biological outputs. Below we highlight a few recent studies that specifically combined the use of genetically encoded biosensors with targeted perturbation tools to provide new insights into the spatiotemporal regulation of intracellular signaling and, thereby, illustrate the power of applying these strategies in tandem to the study of biological systems.
The precise spatial control of signaling pathways is often achieved through a balance between the synthesis and degradation of signaling molecules (e.g. cAMP) (Baillie, 2009; Cooper, 2003; Houslay, 2010; Stangherlin and Zaccolo, 2012), as well as through structural factors, such as cellular compartments and scaffold proteins (Dodge-Kafka et al., 2006), which establish local microdomains that contain varying concentrations of these molecules. To probe how local cAMP concentrations affect the specificity of cAMP–PKA signaling, we developed the aforementioned method for generating site-specific cAMP signals using soluble adenylyl cyclase (Sample et al., 2012). Using this method in conjunction with real-time imaging and mechanistic modeling, we were able to infer the existence of a pool of PKA holoenzyme that resides in the nucleus in HEK-293 cells. The existence of a resident pool of nuclear PKA had heretofore been a standing controversy in the field, with the traditional dogma holding that nuclear PKA activity only occurs through the slow translocation of activated PKA from the cytosol into the nucleus. However, by tuning cAMP signals in different subcellular compartments, such as the nucleus, cytoplasm or plasma membrane, and by monitoring PKA activity exclusively in the nucleus, we observed clear differences in the rate at which PKA activity arises in the nucleus. Specifically, when cAMP signals that are produced in or around the nucleus passed a critical threshold, we unequivocally demonstrated the rapid activation of a resident nuclear pool of PKA. However, cAMP that is produced at the plasma membrane activates a cytoplasmic pool of PKA, which then enters the nucleus by diffusion, as evidenced by the slow responses of the PKA activity reporter (Sample et al., 2012).
Targeted perturbations are also tremendously useful in unraveling the molecular events that are involved in complex processes such as cell migration. Cell migration depends on polarized morphogenesis, in which an initially stationary cell undergoes a symmetry-breaking event to form a migratory protrusion; and this typically involves the spontaneous self-organization of the intracellular machinery in response to stochastic variations in signaling molecules and interacting local and global feedback circuits that amplify these signals at the future site of polarization, while suppressing them elsewhere in the cell (Dehmelt and Bastiaens, 2010). For example, phosphoinositide signaling through PI3K has been shown to play an important role in regulating spontaneous membrane protrusions. To probe the feedback mechanisms that control PI3K signaling during this process, a recent study used a CID system to selectively induce plasma membrane PI3K activity in fibroblasts (Thevathasan et al., 2013). Using a genetically encoded biosensor that specifically detects 3′ phosphoinositides produced by PI3K, the authors initially observed a uniform biosensor response upon the induction of PI3K activity; however, this is rapidly followed by a spontaneous increase in PI3K activity in one part of the membrane, culminating in the formation of a robust cell protrusion. The authors then used this system to identify the GTPase HRas as a key component in the feedback process, as an HRas-targeted siRNA was shown to be able to abolish the local PI3K amplification (Thevathasan et al., 2013).
Similarly, another study sought to evaluate the effect of local Rac activation on Drosophila border cell migration (Wang et al., 2010). Border cells comprise a small group of six to eight cells that are derived from the follicular epithelium within each egg chamber of the Drosophila ovary. During stage 9 of oogenesis, these cells migrate en masse through the nurse cells to make contact with the developing oocyte, and it has long been known that Rac activity is required for this process. To investigate this further, transgenic flies were generated that express photoactivatable Rac (PA-RacQ61L) only in border cells. Light activation of PA-RacQ61L in a cell directly adjacent to the leading cell of the migrating cluster (characterized by a prominent, forward-directed protrusion) was shown to induce lateral movement away from the original path. This was followed by the subsequent activation of PA-RacQ61L in yet another cell, thereby further redirecting the cluster (Wang et al., 2010). Furthermore, when they expressed a FRET-based Rac GTPase biosensor in these cells, the authors observed an asymmetric pattern of Rac activity in endogenously migrating border cells, with the highest activity in elongating protrusions. Thus, through a combination of PA-Rac analogs and a FRET-based Rac biosensor, it was shown that local increases in Rac activity are sufficient to control the direction of a migrating cell cluster, indicating that asymmetric Rac GTPase activity is a key factor in direction sensing in vivo (Wang et al., 2010).
Photoactivatable signaling proteins are especially useful in unraveling the intricate signaling dynamics that characterize neuronal circuitry. For instance, light-induced ion channels such as channelrhodopsins can be used to generate action potentials within a selected neuron in order to map out specific synaptic connections (Petreanu et al., 2007). Furthermore, such optogenetic tools now often complement the use of genetically encoded Ca2+ indicators (GECIs), which are popular for measuring neuronal activity by monitoring Ca2+ transients that are evoked by action potentials and various synaptic inputs; and integrating these parallel approaches might dramatically alter the way neuronal signaling is studied. However, the activation spectra of different light-induced channels (e.g. channelrhodopsin, halorhodopsin, etc.) overlap significantly with the excitation spectra of commonly used GECIs (e.g. GCaMP, Yellow Cameleon, etc.), which makes it difficult to monitor Ca2+ without also activating the channel (Akerboom et al., 2013). Thus, numerous protein engineering efforts are underway to produce spectrally shifted variants of both channelrhodopsins (Prigge et al., 2012) and GECIs (Akerboom et al., 2013; Ohkura et al., 2012; Wu et al., 2013) in order to facilitate their simultaneous use. For example, a recently published study describes the construction of multiple color variants of GCaMP, including the red-shifted RCaMP (Akerboom et al., 2013). Using transgenic C. elegans that express RCaMP in muscle cells and ChR2 in either muscle or upstream motor neurons, the authors were able to successfully induce and monitor muscle contractions simultaneously in vivo.
Perspectives
The ability to precisely and dynamically alter and observe the state of various biological processes is becoming increasingly fundamental to furthering our understanding of how living systems function. With the aid of genetically encoded molecular tools, it has become possible to both perturb and visualize the dynamics of signaling molecules with spatial and temporal precision in living biological systems. By simultaneously applying these complementary techniques, researchers can more directly interrogate signaling networks and delineate causal relationships between the different molecular players. As discussed in this Commentary, these efforts have already yielded many new insights into the regulation and function of signaling pathways, although they are still largely in their infancy.
As with any emerging technology, there is always room for the development of new and innovative alternatives. In a recent study, for example, magnetic particles conjugated to either the GTPase Ran1 or the GEF Rcc1 were added to cytoplasmic extracts and then used to nucleate microtubule formation in response to an applied magnetic field (Hoffmann et al., 2013). Pushing this concept further, another study demonstrated the use of a magnetic tip to precisely control the location of a single magnetic particle that was conjugated to either Cdc42, Rac1 or a Rac1 GEF, within a living cell and, thus, spatially regulate actin polymerization (Etoc et al., 2013). Magnetic manipulation is an exciting approach that is complementary to existing perturbation tools and could offer remarkably precise spatiotemporal control over in vivo biochemistry. However, many challenges remain to be overcome, especially with regard to the development of genetically encoded magnetic particles, before this technique matures.
Although signaling molecules have been at the forefront in the application of these genetically encodable molecular probes, the potential scope for applying this approach to probe molecular and cellular processes goes beyond cell signaling. For instance, signaling is deeply integrated with other crucial cellular systems, in particular energy metabolism and gene regulation. Metabolic pathways, much like signaling pathways, are highly dynamic and perhaps even spatially compartmentalized (Deuschle et al., 2005), and researchers have already begun developing tools, including biosensors for metabolic markers, such as the ATP:ADP ratio (Berg et al., 2009) and the NADH–NAD+ redox state (Hung et al., 2011), that are aimed at dissecting the spatiotemporal organization of metabolic pathways. Furthermore, a recent study describing the generation of light-induced transcriptional effectors (LITEs) offers a genetically encoded system that not only enables the dynamic optical control of transcription but also of epigenetic regulation in living cells (Konermann et al., 2013).
Genetically encoded molecular tools are also shedding light on molecular processes within the context of intact animals (Akerboom et al., 2009; Dagliyan et al., 2013; Kittelmann et al., 2013; Lissandron et al., 2007; Tsuyama et al., 2013). However, outside of small or optically clear specimens, the ability to penetrate beyond superficial tissue layers to investigate processes in vital tissues remains a major barrier to whole-animal studies. This can be particularly problematic for chemical perturbation systems. In a recent study, for instance, the allosteric activation of a kinase in transgenic zebrafish embryos required the expression of the kinase in epidermal cells and the bathing of the embryos in a rapamycin solution (Dagliyan et al., 2013). Furthermore, both optogenetic systems and FP-based biosensors are also limited by their reliance on visible light, which penetrates poorly into biological tissues, whereas light with a wavelength between 650 and 900 nm is more suitable (Jöbsis, 1977; Weissleder and Ntziachristos, 2003). To this end, the development of several red-shifted FP variants (Miyawaki et al., 2012), together with similar efforts yielding red-shifted optogenetic perturbation tools (Prigge et al., 2012), may lead to the development of new probes that operate in this part of the spectrum. Similarly, technical advances, such as two-photon excitation (Helmchen and Denk, 2005) of both FPs (Drobizhev et al., 2011; Kawano et al., 2008) and optogenetic tools (Oron et al., 2012), to enable the use of standard probes with long wavelength illumination (Andrasfalvy et al., 2010; Breart et al., 2008; Kamioka et al., 2013) as well as the use of optical fibers for more direct application of light onto target areas (Jego et al., 2013; Sparta et al., 2011), are helping to overcome some of the constraints of using these tools intravitally. Thus, as these various strategies mature and become integrated (Dhakal et al., 2013), we can begin to glimpse the future uses of genetically encoded molecular tools to illuminate all aspects of biological function.
Supplementary Material
Footnotes
Competing interests
The authors declare no competing interests.
Funding
This work is supported by the National Institutes of Health [grant numbers R01 DK073368 and DP1 CA174423 to J.Z.]. Deposited in PMC for release after 12 months.
References
- Airan R. D., Thompson K. R., Fenno L. E., Bernstein H., Deisseroth K. (2009). Temporally precise in vivo control of intracellular signalling. Nature 458, 1025–1029 10.1038/nature07926 [DOI] [PubMed] [Google Scholar]
- Akerboom J., Rivera J. D. V., Guilbe M. M. R., Malavé E. C. A., Hernandez H. H., Tian L., Hires S. A., Marvin J. S., Looger L. L., Schreiter E. R. (2009). Crystal structures of the GCaMP calcium sensor reveal the mechanism of fluorescence signal change and aid rational design. J. Biol. Chem. 284, 6455–6464 10.1074/jbc.M807657200 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Akerboom J., Carreras Calderón N., Tian L., Wabnig S., Prigge M., Tolö J., Gordus A., Orger M. B., Severi K. E., Macklin J. J. et al. (2013). Genetically encoded calcium indicators for multi-color neural activity imaging and combination with optogenetics. Front Mol. Neurosci 6, 2 10.3389/fnmol.2013.00002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andrasfalvy B. K., Zemelman B. V., Tang J., Vaziri A. (2010). Two-photon single-cell optogenetic control of neuronal activity by sculpted light. Proc. Natl. Acad. Sci. USA 107, 11981–11986 10.1073/pnas.1006620107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baillie G. S. (2009). Compartmentalized signalling: spatial regulation of cAMP by the action of compartmentalized phosphodiesterases. FEBS J. 276, 1790–1799 10.1111/j.1742-4658.2009.06926.x [DOI] [PubMed] [Google Scholar]
- Balla T. (2009). Green light to illuminate signal transduction events. Trends Cell Biol. 19, 575–586 10.1016/j.tcb.2009.08.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berg J., Hung Y. P., Yellen G. (2009). A genetically encoded fluorescent reporter of ATP:ADP ratio. Nat. Methods 6, 161–166 10.1038/nmeth.1288 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bloom F. E., Wedner H. J., Parker C. W. (1973). The use of antibodies to study cell structure and metabolism. Pharmacol. Rev. 25, 343–358 [PubMed] [Google Scholar]
- Breart B., Lemaître F., Celli S., Bousso P. (2008). Two-photon imaging of intratumoral CD8+ T cell cytotoxic activity during adoptive T cell therapy in mice. J. Clin. Invest. 118, 1390–1397 10.1172/JCI34388 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown M. D., Sacks D. B. (2009). Protein scaffolds in MAP kinase signalling. Cell. Signal. 21, 462–469 10.1016/j.cellsig.2008.11.013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Campbell R. E. (2009). Fluorescent-protein-based biosensors: modulation of energy transfer as a design principle. Anal. Chem. 81, 5972–5979 10.1021/ac802613w [DOI] [PubMed] [Google Scholar]
- Coons A. H. (1971). The development of immunohistochemistry. Ann. N. Y. Acad. Sci. 177, 5–9 10.1111/j.1749-6632.1971.tb35025.x [DOI] [PubMed] [Google Scholar]
- Coons A. H., Creech H. J., Jones R. N., Berliner E. (1942). The demonstration of pneumococcal antigen in tissues by the use of fluorescent antibody. J. Immunol. 45, 159–170 [Google Scholar]
- Cooper D. M. F. (2003). Regulation and organization of adenylyl cyclases and cAMP. Biochem. J. 375, 517–529 10.1042/BJ20031061 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crabtree G. R., Schreiber S. L. (1996). Three-part inventions: intracellular signaling and induced proximity. Trends Biochem. Sci. 21, 418–422 10.1016/S0968-0004(96)20027-1 [DOI] [PubMed] [Google Scholar]
- Dagliyan O., Shirvanyants D., Karginov A. V., Ding F., Fee L., Chandrasekaran S. N., Freisinger C. M., Smolen G. A., Huttenlocher A., Hahn K. M. et al. (2013). Rational design of a ligand-controlled protein conformational switch. Proc. Natl. Acad. Sci. USA 110, 6800–6804 10.1073/pnas.1218319110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Day R. N., Davidson M. W. (2009). The fluorescent protein palette: tools for cellular imaging. Chem. Soc. Rev. 38, 2887–2921 10.1039/b901966a [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dehmelt L., Bastiaens P. I. H. (2010). Spatial organization of intracellular communication: insights from imaging. Nat. Rev. Mol. Cell Biol. 11, 440–452 10.1038/nrm2903 [DOI] [PubMed] [Google Scholar]
- DeRose R., Miyamoto T., Inoue T. (2013). Manipulating signaling at will: chemically-inducible dimerization (CID) techniques resolve problems in cell biology. Pflugers Arch. 465, 409–417 10.1007/s00424-012-1208-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deuschle K., Fehr M., Hilpert M., Lager I., Lalonde S., Looger L. L., Okumoto S., Persson J., Schmidt A., Frommer W. B. (2005). Genetically encoded sensors for metabolites. Cytometry A 64A, 3–9 10.1002/cyto.a.20119 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dhakal K., Gu L., Black B., Mohanty S. K. (2013). Fiber-optic two-photon optogenetic stimulation. Opt. Lett. 38, 1927–1929 10.1364/OL.38.001927 [DOI] [PubMed] [Google Scholar]
- Dodge-Kafka K. L., Langeberg L., Scott J. D. (2006). Compartmentation of cyclic nucleotide signaling in the heart: the role of A-kinase anchoring proteins. Circ. Res. 98, 993–1001 10.1161/01.RES.0000218273.91741.30 [DOI] [PubMed] [Google Scholar]
- Drobizhev M., Makarov N. S., Tillo S. E., Hughes T. E., Rebane A. (2011). Two-photon absorption properties of fluorescent proteins. Nat. Methods 8, 393–399 10.1038/nmeth.1596 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Etoc F., Lisse D., Bellaiche Y., Piehler J., Coppey M., Dahan M. (2013). Subcellular control of Rac-GTPase signalling by magnetogenetic manipulation inside living cells. Nat. Nanotechnol. 8, 193–198 10.1038/nnano.2013.23 [DOI] [PubMed] [Google Scholar]
- Fessenden J. D., Mahalingam M. (2013). Site-specific labeling of the type 1 ryanodine receptor using biarsenical fluorophores targeted to engineered tetracysteine motifs. PLoS ONE 8, e64686 10.1371/journal.pone.0064686 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fuller B. G., Lampson M. A., Foley E. A., Rosasco-Nitcher S., Le K. V., Tobelmann P., Brautigan D. L., Stukenberg P. T., Kapoor T. M. (2008). Midzone activation of aurora B in anaphase produces an intracellular phosphorylation gradient. Nature 453, 1132–1136 10.1038/nature06923 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gavet O., Pines J. (2010). Progressive activation of CyclinB1-Cdk1 coordinates entry to mitosis. Dev. Cell 18, 533–543 10.1016/j.devcel.2010.02.013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gorostiza P., Isacoff E. Y. (2008). Optical switches for remote and noninvasive control of cell signaling. Science 322, 395–399 10.1126/science.1166022 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gray A., Van Der Kaay J., Downes C. P. (1999). The pleckstrin homology domains of protein kinase B and GRP1 (general receptor for phosphoinositides-1) are sensitive and selective probes for the cellular detection of phosphatidylinositol 3,4-bisphosphate and/or phosphatidylinositol 3,4,5-trisphosphate in vivo. Biochem. J. 344, 929–936 10.1042/0264-6021:3440929 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grynkiewicz G., Poenie M., Tsien R. Y. (1985). A new generation of Ca2+ indicators with greatly improved fluorescence properties. J. Biol. Chem. 260, 3440–3450 [PubMed] [Google Scholar]
- Harvey C. D., Ehrhardt A. G., Cellurale C., Zhong H., Yasuda R., Davis R. J., Svoboda K. (2008). A genetically encoded fluorescent sensor of ERK activity. Proc. Natl. Acad. Sci. USA 105, 19264–19269 10.1073/pnas.0804598105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Helmchen F., Denk W. (2005). Deep tissue two-photon microscopy. Nat. Methods 2, 932–940 10.1038/nmeth818 [DOI] [PubMed] [Google Scholar]
- Hoffmann C., Mazari E., Lallet S., Le Borgne R., Marchi V., Gosse C., Gueroui Z. (2013). Spatiotemporal control of microtubule nucleation and assembly using magnetic nanoparticles. Nat. Nanotechnol. 8, 199–205 10.1038/nnano.2012.246 [DOI] [PubMed] [Google Scholar]
- Houslay M. D. (2010). Underpinning compartmentalised cAMP signalling through targeted cAMP breakdown. Trends Biochem. Sci. 35, 91–100 10.1016/j.tibs.2009.09.007 [DOI] [PubMed] [Google Scholar]
- Hung Y. P., Albeck J. G., Tantama M., Yellen G. (2011). Imaging cytosolic NADH-NAD(+) redox state with a genetically encoded fluorescent biosensor. Cell Metab. 14, 545–554 10.1016/j.cmet.2011.08.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ibraheem A., Campbell R. E. (2010). Designs and applications of fluorescent protein-based biosensors. Curr. Opin. Chem. Biol. 14, 30–36 10.1016/j.cbpa.2009.09.033 [DOI] [PubMed] [Google Scholar]
- Idevall-Hagren O., Dickson E. J., Hille B., Toomre D. K., De Camilli P. (2012). Optogenetic control of phosphoinositide metabolism. Proc. Natl. Acad. Sci. USA 109, E2316–E2323 10.1073/pnas.1211305109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Inoue T., Meyer T. (2008). Synthetic activation of endogenous PI3K and Rac identifies an AND-gate switch for cell polarization and migration. PLoS ONE 3, e3068 10.1371/journal.pone.0003068 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Inoue T., Heo W. D., Grimley J. S., Wandless T. J., Meyer T. (2005). An inducible translocation strategy to rapidly activate and inhibit small GTPase signaling pathways. Nat. Methods 2, 415–418 10.1038/nmeth763 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jego S., Glasgow S. D., Herrera C. G., Ekstrand M., Reed S. J., Boyce R., Friedman J., Burdakov D., Adamantidis A. R. (2013). Optogenetic identification of a rapid eye movement sleep modulatory circuit in the hypothalamus. Nat. Neurosci. 16, 1637–1643 10.1038/nn.3522 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jöbsis F. F. (1977). Noninvasive, infrared monitoring of cerebral and myocardial oxygen sufficiency and circulatory parameters. Science 198, 1264–1267 10.1126/science.929199 [DOI] [PubMed] [Google Scholar]
- Jung D., Min K., Jung J., Jang W., Kwon Y. (2013). Chemical biology-based approaches on fluorescent labeling of proteins in live cells. Mol. Biosyst. 9, 862–872 10.1039/c2mb25422k [DOI] [PubMed] [Google Scholar]
- Kamioka Y., Sumiyama K., Mizuno R., Matsuda M. (2013). Live imaging of transgenic mice expressing FRET biosensors. Conf. Proc. IEEE Eng. Med. Biol. Soc. 2013, 125–128 [DOI] [PubMed] [Google Scholar]
- Karginov A. V., Ding F., Kota P., Dokholyan N. V., Hahn K. M. (2010). Engineered allosteric activation of kinases in living cells. Nat. Biotechnol. 28, 743–747 10.1038/nbt.1639 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karginov A. V., Zou Y., Shirvanyants D., Kota P., Dokholyan N. V., Young D. D., Hahn K. M., Deiters A. (2011). Light regulation of protein dimerization and kinase activity in living cells using photocaged rapamycin and engineered FKBP. J. Am. Chem. Soc. 133, 420–423 10.1021/ja109630v [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kawano H., Kogure T., Abe Y., Mizuno H., Miyawaki A. (2008). Two-photon dual-color imaging using fluorescent proteins. Nat. Methods 5, 373–374 10.1038/nmeth0508-373 [DOI] [PubMed] [Google Scholar]
- Kennedy M. J., Hughes R. M., Peteya L. A., Schwartz J. W., Ehlers M. D., Tucker C. L. (2010). Rapid blue-light-mediated induction of protein interactions in living cells. Nat. Methods 7, 973–975 10.1038/nmeth.1524 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kholodenko B. N., Hancock J. F., Kolch W. (2010). Signalling ballet in space and time. Nat. Rev. Mol. Cell Biol. 11, 414–426 10.1038/nrm2901 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim J-M., Hwa J., Garriga P., Reeves P. J., RajBhandary U. L., Khorana H. G. (2005). Light-driven activation of beta 2-adrenergic receptor signaling by a chimeric rhodopsin containing the beta 2-adrenergic receptor cytoplasmic loops. Biochemistry 44, 2284–2292 10.1021/bi048328i [DOI] [PubMed] [Google Scholar]
- Kittelmann M., Liewald J. F., Hegermann J., Schultheis C., Brauner M., Steuer Costa W., Wabnig S., Eimer S., Gottschalk A. (2013). In vivo synaptic recovery following optogenetic hyperstimulation. Proc. Natl. Acad. Sci. USA 110, E3007–E3016 10.1073/pnas.1305679110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kiyokawa E., Aoki K., Nakamura T., Matsuda M. (2011). Spatiotemporal regulation of small GTPases as revealed by probes based on the principle of Förster Resonance Energy Transfer (FRET): Implications for signaling and pharmacology. Annu. Rev. Pharmacol. Toxicol. 51, 337–358 10.1146/annurev-pharmtox-010510-100234 [DOI] [PubMed] [Google Scholar]
- Kolch W. (2005). Coordinating ERK/MAPK signalling through scaffolds and inhibitors. Nat. Rev. Mol. Cell Biol. 6, 827–837 10.1038/nrm1743 [DOI] [PubMed] [Google Scholar]
- Konermann S., Brigham M. D., Trevino A. E., Hsu P. D., Heidenreich M., Cong L., Platt R. J., Scott D. A., Church G. M., Zhang F. (2013). Optical control of mammalian endogenous transcription and epigenetic states. Nature 500, 472–476 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levskaya A., Weiner O. D., Lim W. A., Voigt C. A. (2009). Spatiotemporal control of cell signalling using a light-switchable protein interaction. Nature 461, 997–1001 10.1038/nature08446 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liang F-S., Ho W. Q., Crabtree G. R. (2011). Engineering the ABA plant stress pathway for regulation of induced proximity. Sci. Signal. 4, rs2 10.1126/scisignal.2001449 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lissandron V., Rossetto M. G., Erbguth K., Fiala A., Daga A., Zaccolo M. (2007). Transgenic fruit-flies expressing a FRET-based sensor for in vivo imaging of cAMP dynamics. Cell. Signal. 19, 2296–2303 10.1016/j.cellsig.2007.07.004 [DOI] [PubMed] [Google Scholar]
- Mehta S., Zhang J. (2011). Reporting from the field: genetically encoded fluorescent reporters uncover signaling dynamics in living biological systems. Annu. Rev. Biochem. 80, 375–401 10.1146/annurev-biochem-060409-093259 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mellors R. C. (1968). The application of labeled antibody technics in studying cell antigens. Cancer Res. 28, 1372–1381 [PubMed] [Google Scholar]
- Miyamoto T., DeRose R., Suarez A., Ueno T., Chen M., Sun T-P., Wolfgang M. J., Mukherjee C., Meyers D. J., Inoue T. (2012). Rapid and orthogonal logic gating with a gibberellin-induced dimerization system. Nat. Chem. Biol. 8, 465–470 10.1038/nchembio.922 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miyawaki A. (2003). Visualization of the spatial and temporal dynamics of intracellular signaling. Dev. Cell 4, 295–305 10.1016/S1534-5807(03)00060-1 [DOI] [PubMed] [Google Scholar]
- Miyawaki A., Llopis J., Heim R., McCaffery J. M., Adams J. A., Ikura M., Tsien R. Y. (1997). Fluorescent indicators for Ca2+ based on green fluorescent proteins and calmodulin. Nature 388, 882–887 10.1038/42264 [DOI] [PubMed] [Google Scholar]
- Miyawaki A., Shcherbakova D. M., Verkhusha V. V. (2012). Red fluorescent proteins: chromophore formation and cellular applications. Curr. Opin. Struct. Biol. 22, 679–688 10.1016/j.sbi.2012.09.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mochizuki N., Yamashita S., Kurokawa K., Ohba Y., Nagai T., Miyawaki A., Matsuda M. (2001). Spatio-temporal images of growth-factor-induced activation of Ras and Rap1. Nature 411, 1065–1068 10.1038/35082594 [DOI] [PubMed] [Google Scholar]
- Nagel G., Ollig D., Fuhrmann M., Kateriya S., Musti A. M., Bamberg E., Hegemann P. (2002). Channelrhodopsin-1: a light-gated proton channel in green algae. Science 296, 2395–2398 10.1126/science.1072068 [DOI] [PubMed] [Google Scholar]
- Nagel G., Szellas T., Huhn W., Kateriya S., Adeishvili N., Berthold P., Ollig D., Hegemann P., Bamberg E. (2003). Channelrhodopsin-2, a directly light-gated cation-selective membrane channel. Proc. Natl. Acad. Sci. USA 100, 13940–13945 10.1073/pnas.1936192100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagel G., Szellas T., Kateriya S., Adeishvili N., Hegemann P., Bamberg E. (2005). Channelrhodopsins: directly light-gated cation channels. Biochem. Soc. Trans. 33, 863–866 10.1042/BST0330863 [DOI] [PubMed] [Google Scholar]
- Nakai J., Ohkura M., Imoto K. (2001). A high signal-to-noise Ca(2+) probe composed of a single green fluorescent protein. Nat. Biotechnol. 19, 137–141 10.1038/84397 [DOI] [PubMed] [Google Scholar]
- Nakamura T., Aoki K., Matsuda M. (2005). Monitoring spatio-temporal regulation of Ras and Rho GTPase with GFP-based FRET probes. Methods 37, 146–153 10.1016/j.ymeth.2005.05.021 [DOI] [PubMed] [Google Scholar]
- Nelson J. D., Hempstead B., Tanaka R., Pauls F. P. (1964). Fluorescent Antibody Diagnosis of Infections. JAMA 188, 1121–1124 10.1001/jama.1964.03060390023006 [DOI] [PubMed] [Google Scholar]
- Newman R. H., Fosbrink M. D., Zhang J. (2011). Genetically encodable fluorescent biosensors for tracking signaling dynamics in living cells. Chem. Rev. 111, 3614–3666 10.1021/cr100002u [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohkura M., Sasaki T., Kobayashi C., Ikegaya Y., Nakai J. (2012). An improved genetically encoded red fluorescent Ca2+ indicator for detecting optically evoked action potentials. PLoS ONE 7, e39933 10.1371/journal.pone.0039933 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oron D., Papagiakoumou E., Anselmi F., Emiliani V. (2012). Two-photon optogenetics. Optogenetics: tools for controlling and monitoring neuronal activity (eds. Knöpfel, T. and Boyden, E. S.) 119–143Amsterdam: Elsevier; [DOI] [PubMed] [Google Scholar]
- Pan C. Q., Sudol M., Sheetz M., Low B. C. (2012). Modularity and functional plasticity of scaffold proteins as p(l)acemakers in cell signaling. Cell. Signal. 24, 2143–2165 10.1016/j.cellsig.2012.06.002 [DOI] [PubMed] [Google Scholar]
- Petreanu L., Huber D., Sobczyk A., Svoboda K. (2007). Channelrhodopsin-2-assisted circuit mapping of long-range callosal projections. Nat. Neurosci. 10, 663–668 10.1038/nn1891 [DOI] [PubMed] [Google Scholar]
- Prigge M., Schneider F., Tsunoda S. P., Shilyansky C., Wietek J., Deisseroth K., Hegemann P. (2012). Color-tuned channelrhodopsins for multiwavelength optogenetics. J. Biol. Chem. 287, 31804–31812 10.1074/jbc.M112.391185 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sample V., Newman R. H., Zhang J. (2009). The structure and function of fluorescent proteins. Chem. Soc. Rev. 38, 2852–2864 10.1039/b913033k [DOI] [PubMed] [Google Scholar]
- Sample V., DiPilato L. M., Yang J. H., Ni Q., Saucerman J. J., Zhang J. (2012). Regulation of nuclear PKA revealed by spatiotemporal manipulation of cyclic AMP. Nat. Chem. Biol. 8, 375–382 10.1038/nchembio.799 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sample V., Ni Q., Mehta S., Inoue T., Zhang J. (2013). Controlling enzymatic action in living cells with a kinase-inducible bimolecular switch. ACS Chem. Biol. 8, 116–121 10.1021/cb300393w [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schröder-Lang S., Schwärzel M., Seifert R., Strünker T., Kateriya S., Looser J., Watanabe M., Kaupp U. B., Hegemann P., Nagel G. (2007). Fast manipulation of cellular cAMP level by light in vivo. Nat. Methods 4, 39–42 10.1038/nmeth975 [DOI] [PubMed] [Google Scholar]
- Scott J. D., Pawson T. (2009). Cell signaling in space and time: where proteins come together and when they're apart. Science 326, 1220–1224 10.1126/science.1175668 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seibel N. M., Eljouni J., Nalaskowski M. M., Hampe W. (2007). Nuclear localization of enhanced green fluorescent protein homomultimers. Anal. Biochem. 368, 95–99 10.1016/j.ab.2007.05.025 [DOI] [PubMed] [Google Scholar]
- Shaner N. C., Steinbach P. A., Tsien R. Y. (2005). A guide to choosing fluorescent proteins. Nat. Methods 2, 905–909 10.1038/nmeth819 [DOI] [PubMed] [Google Scholar]
- Sparta D. R., Stamatakis A. M., Phillips J. L., Hovelsø N., van Zessen R., Stuber G. D. (2011). Construction of implantable optical fibers for long-term optogenetic manipulation of neural circuits. Nat. Protoc. 7, 12–23 10.1038/nprot.2011.413 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stangherlin A., Zaccolo M. (2012). Phosphodiesterases and subcellular compartmentalized cAMP signaling in the cardiovascular system. AJP: Heart and Circulatory Physiology 302, H379–H390 10.1152/ajpheart.00766.2011 [DOI] [PubMed] [Google Scholar]
- Stierl M., Stumpf P., Udwari D., Gueta R., Hagedorn R., Losi A., Gärtner W., Petereit L., Efetova M., Schwärzel M. et al. (2011). Light modulation of cellular cAMP by a small bacterial photoactivated adenylyl cyclase, bPAC, of the soil bacterium Beggiatoa. J. Biol. Chem. 286, 1181–1188 10.1074/jbc.M110.185496 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suh B-C., Inoue T., Meyer T., Hille B. (2006). Rapid chemically induced changes of PtdIns(4,5)P2 gate KCNQ ion channels. Science 314, 1454–1457 10.1126/science.1131163 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tantama M., Hung Y. P., Yellen G. (2012). Optogenetic reporters: Fluorescent protein-based genetically encoded indicators of signaling and metabolism in the brain. Prog. Brain Res. 196, 235–263 10.1016/B978-0-444-59426-6.00012-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taylor D. L., Wang Y. L. (1978). Molecular cytochemistry: incorporation of fluorescently labeled actin into living cells. Proc. Natl. Acad. Sci. USA 75, 857–861 10.1073/pnas.75.2.857 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taylor D., Amato P., Luby-Phelps K., McNeil P. (1984). Fluorescent analog cytochemistry. Trends Biochem. Sci. 9, 88–91 10.1016/0968-0004(84)90098-7 [DOI] [Google Scholar]
- Thevathasan J. V., Tan E., Zheng H., Lin Y. C., Li Y., Inoue T., Fivaz M. (2013). The small GTPase HRas shapes local PI3K signals through positive feedback and regulates persistent membrane extension in migrating fibroblasts. Mol. Biol. Cell 24, 2228–2237 10.1091/mbc.E12-12-0905 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ting A. Y., Kain K. H., Klemke R. L., Tsien R. Y. (2001). Genetically encoded fluorescent reporters of protein tyrosine kinase activities in living cells. Proc. Natl. Acad. Sci. USA 98, 15003–15008 10.1073/pnas.211564598 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsien R. Y. (1989). Fluorescent probes of cell signaling. Annu. Rev. Neurosci. 12, 227–253 10.1146/annurev.ne.12.030189.001303 [DOI] [PubMed] [Google Scholar]
- Tsien R. Y. (1998). The green fluorescent protein. Annu. Rev. Biochem. 67, 509–544 10.1146/annurev.biochem.67.1.509 [DOI] [PubMed] [Google Scholar]
- Tsuyama T., Kishikawa J., Han Y-W., Harada Y., Tsubouchi A., Noji H., Kakizuka A., Yokoyama K., Uemura T., Imamura H. (2013). In vivo fluorescent adenosine 5′-triphosphate (ATP) imaging of Drosophila melanogaster and Caenorhabditis elegans by using a genetically encoded fluorescent ATP biosensor optimized for low temperatures. Anal. Chem. 85, 7889–7896 10.1021/ac4015325 [DOI] [PubMed] [Google Scholar]
- Umeda N., Ueno T., Pohlmeyer C., Nagano T., Inoue T. (2011). A photocleavable rapamycin conjugate for spatiotemporal control of small GTPase activity. J. Am. Chem. Soc. 133, 12–14 10.1021/ja108258d [DOI] [PMC free article] [PubMed] [Google Scholar]
- Violin J. D., Zhang J., Tsien R. Y., Newton A. C. (2003). A genetically encoded fluorescent reporter reveals oscillatory phosphorylation by protein kinase C. J. Cell Biol. 161, 899–909 10.1083/jcb.200302125 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Volgraf M., Gorostiza P., Numano R., Kramer R. H., Isacoff E. Y., Trauner D. (2006). Allosteric control of an ionotropic glutamate receptor with an optical switch. Nat. Chem. Biol. 2, 47–52 10.1038/nchembio756 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang X., He L., Wu Y. I., Hahn K. M., Montell D. J. (2010). Light-mediated activation reveals a key role for Rac in collective guidance of cell movement in vivo. Nat. Cell Biol. 12, 591–597 10.1038/ncb2061 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weissleder R., Ntziachristos V. (2003). Shedding light onto live molecular targets. Nat. Med. 9, 123–128 10.1038/nm0103-123 [DOI] [PubMed] [Google Scholar]
- Whitaker M. (2000). Fluorescent tags of protein function in living cells. Bioessays 22, 180–187 [DOI] [PubMed] [Google Scholar]
- Wong W., Scott J. D. (2004). AKAP signalling complexes: focal points in space and time. Nat. Rev. Mol. Cell Biol. 5, 959–970 10.1038/nrm1527 [DOI] [PubMed] [Google Scholar]
- Wu Y. I., Frey D., Lungu O. I., Jaehrig A., Schlichting I., Kuhlman B., Hahn K. M. (2009). A genetically encoded photoactivatable Rac controls the motility of living cells. Nature 461, 104–108 10.1038/nature08241 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu J., Liu L., Matsuda T., Zhao Y., Rebane A., Drobizhev M., Chang Y-F., Araki S., Arai Y., March K. et al. (2013). Improved orange and red Ca2± indicators and photophysical considerations for optogenetic applications. ACS Chem. Neurosci. 4, 963–972 10.1021/cn400012b [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoo S. K., Deng Q., Cavnar P. J., Wu Y. I., Hahn K. M., Huttenlocher A. (2010). Differential regulation of protrusion and polarity by PI3K during neutrophil motility in live zebrafish. Dev. Cell 18, 226–236 10.1016/j.devcel.2009.11.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zaccolo M. (2004). Use of chimeric fluorescent proteins and fluorescence resonance energy transfer to monitor cellular responses. Circ. Res. 94, 866–873 10.1161/01.RES.0000123825.83803.CD [DOI] [PubMed] [Google Scholar]
- Zhang J. (2009). The colorful journey of green fluorescent protein. ACS Chem. Biol. 4, 85–88 10.1021/cb900027r [DOI] [PubMed] [Google Scholar]
- Zhang J., Ma Y., Taylor S. S., Tsien R. Y. (2001). Genetically encoded reporters of protein kinase A activity reveal impact of substrate tethering. Proc. Natl. Acad. Sci. USA 98, 14997–15002 10.1073/pnas.211566798 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang F., Wang L-P., Brauner M., Liewald J. F., Kay K., Watzke N., Wood P. G., Bamberg E., Nagel G., Gottschalk A. et al. (2007). Multimodal fast optical interrogation of neural circuitry. Nature 446, 633–639 10.1038/nature05744 [DOI] [PubMed] [Google Scholar]
- Zhang F., Vierock J., Yizhar O., Fenno L. E., Tsunoda S., Kianianmomeni A., Prigge M., Berndt A., Cushman J., Polle J. et al. (2011). The microbial opsin family of optogenetic tools. Cell 147, 1446–1457 10.1016/j.cell.2011.12.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao Y., Araki S., Wu J., Teramoto T., Chang Y. F., Nakano M., Abdelfattah A. S., Fujiwara M., Ishihara T., Nagai T. et al. (2011). An expanded palette of genetically encoded Ca2+ indicators. Science 333, 1888–1891 10.1126/science.1208592 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou X. X., Chung H. K., Lam A. J., Lin M. Z. (2012). Optical control of protein activity by fluorescent protein domains. Science 338, 810–814 10.1126/science.1226854 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zürn A., Klenk C., Zabel U., Reiner S., Lohse M. J., Hoffmann C. (2010). Site-specific, orthogonal labeling of proteins in intact cells with two small biarsenical fluorophores. Bioconjug. Chem. 21, 853–859 10.1021/bc900394j [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.