

Abstract
Marine spatial planning (MSP), whereby areas of the ocean are zoned for different uses, has great potential to reduce or eliminate conflicts between competing management goals, but only if strategically applied. The recent literature overwhelmingly agrees that including stakeholders in these planning processes is critical to success; but, given the countless alternative ways even simple spatial regulations can be configured, how likely is it that a stakeholder-driven process will generate plans that deliver on the promise of MSP? Here, we use a spatially explicit, dynamic bioeconomic model to show that stakeholder-generated plans are doomed to fail in the absence of strong scientific guidance. While strategically placed spatial regulations can improve outcomes remarkably, the vast majority of possible plans fail to achieve this potential. Surprisingly, existing scientific rules of thumb do little to improve outcomes. Here, we develop an alternative approach in which models are used to identify efficient plans, which are then modified by stakeholders. Even if stakeholders alter these initial proposals considerably, results hugely outperform plans guided by scientific rules of thumb. Our results underscore the importance of spatially explicit dynamic models for the management of marine resources and illustrate how such models can be harmoniously integrated into a stakeholder-driven MSP process.
Keywords: marine spatial planning, bioeconomics, ecological modelling, marine-protected areas, reserve design
1. Introduction
Marine spatial planning (MSP), in which areas of the ocean are zoned for different uses, is becoming increasingly important for balancing competing management objectives [1]. The initial design of a marine spatial plan is crucial for its long-term effectiveness. Different configurations of spatial restrictions can lead to vastly different outcomes [2], and plans are costly to adjust once implemented. Current recommendations for MSP emphasize the importance of stakeholder participation in the design phase [3–7], and several recent planning processes have relied heavily on stakeholder input [8–10]. The advantages of this approach are compelling [11]; local users, such as recreational and commercial fishers, boaters, divers, scientists and conservationists, have first-hand knowledge of the system, and public participation facilitates acceptance of and compliance with the plan. However, there are also drawbacks. Placing stakeholders at the centre of the design process demands that scientific guidance be provided in a form that is accessible to non-experts. Further, because any plan that restricts users’ activities will be contentious [12], scientific guidance may be ignored.
How likely is it that an effective spatial plan can be developed without scientific guidance? What forms of guidance most improve outcomes? Neither of these questions has been explored within a realistic context, and in particular, the likely costs of planning without scientific guidance have not been quantified. Here, we assess the likely outcome of MSP for a case study involving the placement of no-take marine-protected areas (MPAs). This example is based on the Marine Life Protection Act process undertaken in southern California between 2008 and 2010 [13]. In that process, a network of MPAs were established in near-shore waters, with varying degrees of protection, including no-take marine reserves. This study models a simplified version of that process by considering only no-take reserves and by evaluating reserve design at a coarser spatial scale. Planning, in this case study, has dual objectives: an economic goal (increasing fishery yields), and a conservation goal (protecting the biomass of harvested species). Because fishing inevitably leads to some decline in fish biomass, the optimal management strategy depends on the relative weight that is placed on these two goals [14].
To quantify the difficulty of finding plans which perform well for both goals, we first identified the set of best possible plans—the Pareto efficiency frontier—and compared these optimal outcomes with haphazardly generated plans. We then filtered the haphazard plans with various guidelines, or scientific ‘rules of thumb’, to determine whether existing forms of scientific guidance can bias plans towards better outcomes. Finally, we demonstrated a new method of combining scientific guidance with stakeholder-based design, in which model-identified optimal plans are altered by stakeholders.
We evaluated these alternative approaches to spatial planning using a spatially explicit, dynamic bioeconomic model to predict the outcomes of alternative plans. Such bioeconomic models are becoming more common for evaluating spatial plans, particularly in the context of fishery management [15–18]. Our model represents populations of eight species caught by a mix of recreational and small-scale commercial fishing: the model simulates fisheries in southern California, representing all shallow habitat between Point Conception and the Mexican border, including the offshore islands. It treats the region as 135 spatial subpopulations of each species, and includes life-history information, the spatial distribution of habitat, likely dispersal patterns during each species’s pelagic larval phase and predictions about how fishing effort will be distributed [2]. The model represents a significant advance on earlier models used to evaluate MPA placement, which have often relied on overly simplified spatial structure or have failed to incorporate the dynamic responses of fish and fishing to management. Because the performance of each MPA plan depends on management outside of the MPAs, we evaluated cases where fishing effort is moderate (effort for each species maximizes yield with no MPAs) and ones in which each species has been seriously overfished.
2. Methods
The bioeconomic model used here represents seven fish species and one invertebrate species—black surfperch (Embiotoca jacksoni), California halibut (Paralichthys californicus), kelp bass (Paralabrax clathratus), kelp rockfish (Sebastes atrovirens), opaleye (Girella nigricans), ocean whitefish (Caulolatilus princeps), sheephead (Semicossyphus pulcher) and red sea urchin (Strongylocentrotus franciscanus)—chosen because they are targeted by commercial and/or recreational fishermen in southern California. The model divides the region into 135 patches and simulates an age-structured subpopulation of each species in each patch, incorporating data about the habitat distribution and life history of each species [2] (full details of the modelling are given in the electronic supplementary material). The subpopulations are connected by dispersal in the larval phase, with probabilities of each species dispersing from patch to patch generated by simulations of ocean currents [19,20]. We also simulated a fishing fleet for each species, and predicted how fishers will distribute their effort in response to alternative networks of no-take MPAs. The model predicts the equilibrium economic outcome (yield relative to maximum sustainable yield without MPAs) and conservation outcome (biomass as a fraction of unfished biomass), averaged over the eight species. Parameters follow those used in evaluations carried out as part of California's Marine Life Protection Act process [13] (parameter values can be found in the electronic supplementary material, table S1). Because the effectiveness of a given MPA network depends on management outside the MPAs, we compared alternative MPA networks within specific management contexts, including cases where the stocks outside of the MPAs were moderately fished and cases where stocks had been depleted by fishing.
Networks on the efficiency frontier were identified using genetic algorithms. Optimizations run from different starting conditions converged on the same frontier, indicating that networks on the frontier were at least locally optimal. We calculated expected outcomes and probability contours for all possible plans based on 100 000 randomly generated networks for which all combinations of closed and open patches were equally likely. For guideline-based networks, randomly generated networks of the stated sizes were filtered, so that only those meeting the guidelines were evaluated (see guidelines for spacing and habitat coverage, below). Optimally seeded networks were created by choosing a network on the efficiency frontier and randomly moving up to 25% of the component MPAs to previously unprotected locations. All simulation and optimization was performed in MATLAB (MathWorks, Natick, MA).
(a). Guidelines for spacing and habitat coverage
Various ‘rule-of-thumb’ guidelines have been proposed and applied [21] for designing networks of MPAs. Although such guidelines have been formulated in diverse ways, they generally have focused on the amount of area (or habitat) contained within individual MPAs and the spacing between adjacent MPAs. Here, we implement a flexible form of guidelines with two rules:
Rule 1: Every patch of habitat must have an MPA within R1 km.
Rule 2: MPAs only satisfy rule 1 if they contain at least R2 km2 of suitable habitat.
Note that on a linear coastline with continuous habitat, rule 1 is equivalent to specifying the maximum spacing between adjacent MPAs—a network with MPAs farther apart than 2 × R1 would fail to meet rule 1. However, guidelines specifying maximum spacing between MPAs do not work in cases where the distribution of habit is complex. If there are isolated patches of habitat (e.g. San Nicolas Island in southern California), no network of MPAs can avoid having a large gap. Our formulation has the advantage that rule 1 can be satisfied as long as each isolated patch has at least one associated MPA. Because of the presence of islands and complex coastlines, we used overwater distances between MPAs to evaluate rule 1. Similarly, if habitat is uniform, then rule 2 effectively specifies a minimum MPA size, but our formulation accounts better for the fact that habitat is distributed patchily in most seascapes. Together, these two rules can approximate a broad range real guidelines, including ones used in real planning processes.
We created a suite of related guidelines by using a wide range of possible values for the parameters R1 (30–120 km in 5 km increments) and R2 (1, 2 and 4 km2). Small values of R1 indicate stricter spacing rules and larger values of R2 indicate stricter minimum habitat requirements. For any pair of parameters, a candidate network is evaluated for all eight species, which have different habitat distributions. A network fails if it does not meet the habitat guidelines for any species. Depending on parameters R1 and R2, as well as the total size of the randomly generated network, a range of guideline selectivity was evaluated, from guidelines that accepted nearly every possible network (high R1, low R2) to guidelines that accepted only one in a million candidate networks (low R1, high R2). Guidelines accepting much less than 1 in a million possible networks were not evaluated, as they represent rules for which suitable networks are impossible or impractically difficult to find. The choice of guidelines had a substantial impact on the structure of the resulting network. For a given total MPA coverage, lower R1 resulted in networks with smaller maximum distances between patches of habitat and the nearest MPA, higher R2 resulted in higher mean total habitat protected, and there were substantial interactions between the two parameters.
3. Results and discussion
We found that a strategically designed marine spatial plan can substantially improve both economic and conservation outcomes. The best designs trace a smoothly downward-sloping efficiency frontier, suggesting that there is a natural trade-off between yield and biomass (figure 1, black curve). Under moderate fishing, an optimal MPA network can increase yields by 8% (while simultaneously increasing fish populations) or increase population biomass by 30% without reducing yields relative to the no-MPA outcome (figure 1a, star). The potential for MPAs to increase yields of species not being overfished contrasts with some previous results [22] and is possible because spatial heterogeneity in the system (particularly in ocean currents) means that adults in some locations are more valuable for population persistence than those in other locations. Spatial regulations that divert fishing away from these valued locations can increase the overall productivity of the fishery. Gains are more spectacular if species have been overfished (e.g. a more than doubling of both yield and biomass; figure 1b). In fact, strategically placed MPAs in an overfished system achieve yields and biomass similar to those available under moderate fishing (as seen by comparing the efficiency frontiers in figure 1a,b), emphasizing the broad potential of strategic MSP.
Figure 1.
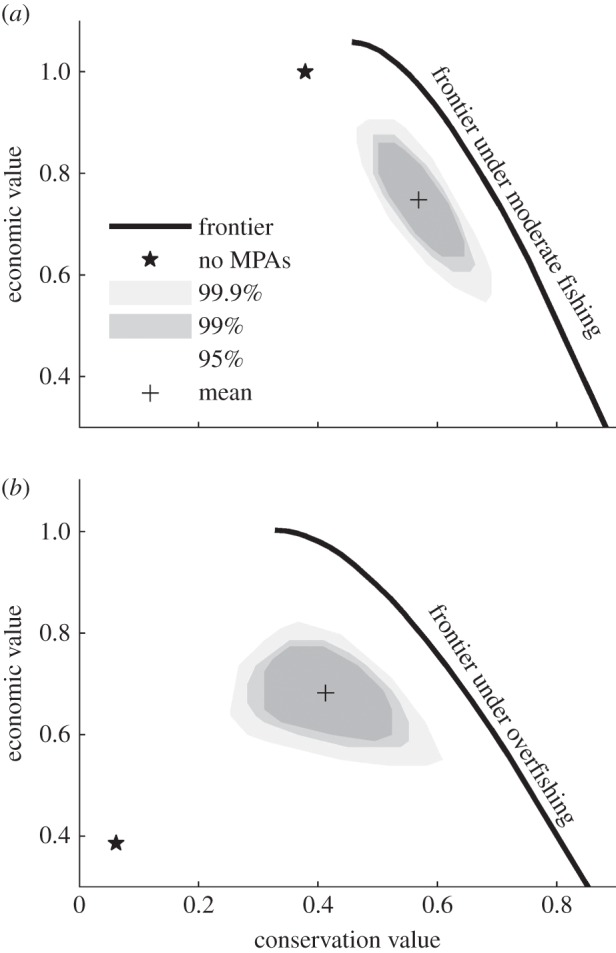
The effect of MPA network choice on conservation and economic outcomes under (a) moderate fishing and (b) overfishing. The star identifies the outcome with no MPAs. The black curve traces combinations of economic and conservation outcomes available with optimally placed MPAs. The areas contain 95%, 99% and 99.9% of all possible MPA networks, as indicated, and the cross indicates the expected outcome, averaged over all possibilities. Results are based on a model of near-shore fisheries in southern California.
Management gains are contingent on effective planning, however, and there is a substantial gap between optimal designs and the vast majority of possible options. On average, randomly designed MPA networks result in dramatically inferior outcomes for both economic and conservation goals (greater than 20% reduction in each metric under moderate fishing; figure 1a), and it is exceptionally unlikely that a randomly selected MPA network will approach the efficiency frontier (99.9% of options are more than 10% below optimal). If the system is overfished, most networks perform even less favourably (greater than 30% expected reduction in each metric; figure 1b). Given the huge number of options (greater than 1040 in this example) and their distribution of likely outcomes, expecting stakeholders to arrive at an efficient marine spatial plan without help is unrealistic. Even if a high-performing plan could be instantly recognized as such, there is a vanishingly small chance that one would be suggested in the first place. This quantitative assessment of the likelihood of achieving near-optimal outcomes is one of the first of its kind (although several studies have looked at the problem of spatial planning under uncertainty in more abstract contexts [23,24]), and it emphasizes the need for tools that can help stakeholders in developing effective marine spatial plans.
Within the context of MPA network design, considerable work has gone into deriving science-based rules of thumb to guide the design process [25]. While models have suggested that such guidelines can improve outcomes, the magnitude of this improvement has not been evaluated relative to optimally designed networks. The simplest kind of guidance specifies a fraction of the managed area to protect. If the area protected is constrained, then the expected outcomes follow a trade-off between economic and conservation goals if fishing effort is optimal or conservative, but trace a hump-shaped curve if the system is overfished (figure 2a). Outcomes on the left side of these hump-shaped curves are strictly inferior to those at the peak or right side, indicating that there is minimum desirable coverage of MPAs; the more severe the overfishing, the higher this minimum coverage (figure 2b). If other design guidance is not provided, however, then substantial MPA coverage is only likely to improve economic outcomes if the fishery is quite depleted; fish stocks must be reduced below 65% of the biomass that gives maximum sustainable yield before 10% or greater coverage of MPAs is demanded. Although guidelines for total MPA coverage improve results under overfished conditions, such guidance does not lead to MPA networks near the optimal outcomes. Options (99%) are more than 10% below the best outcomes for both metrics under moderate fishing and more than 40% below optimal under overfishing (figure 3, grey regions).
Figure 2.
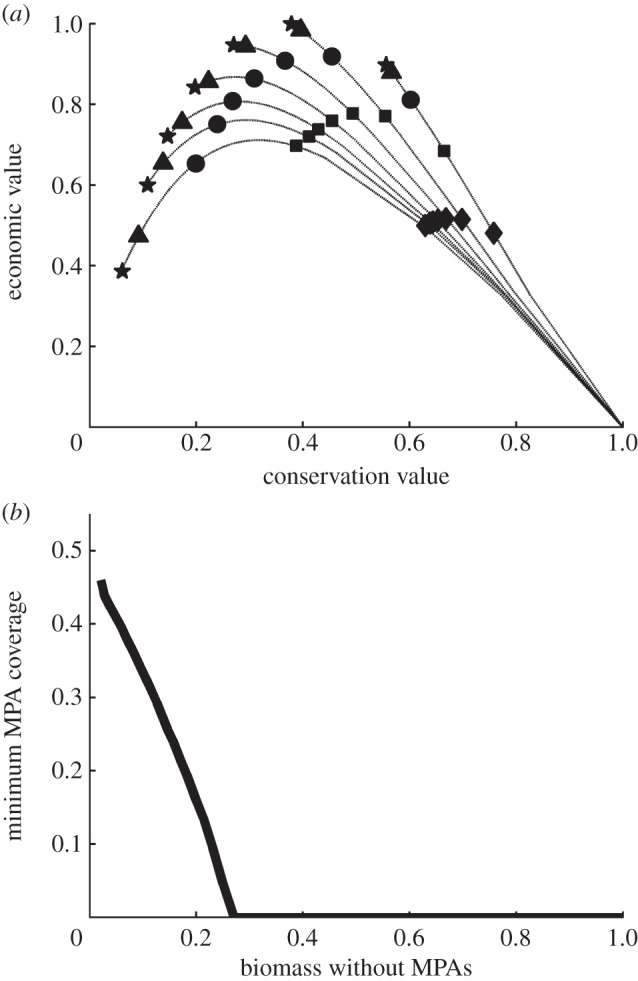
(a) Expected outcomes for various levels of MPA coverage in southern California (star = 0%, triangle = 8%, circle = 25%, square = 50%, diamond = 75%). Each curve represents expected values under a different level of fishing effort, ranging from four times (lower curve) to 0.5 times (upper curve) the effort that gives maximum sustainable yield with no MPAs. (b) The MPA coverage that maximizes expected economic value as a function of the equilibrium biomass without MPAs.
Figure 3.
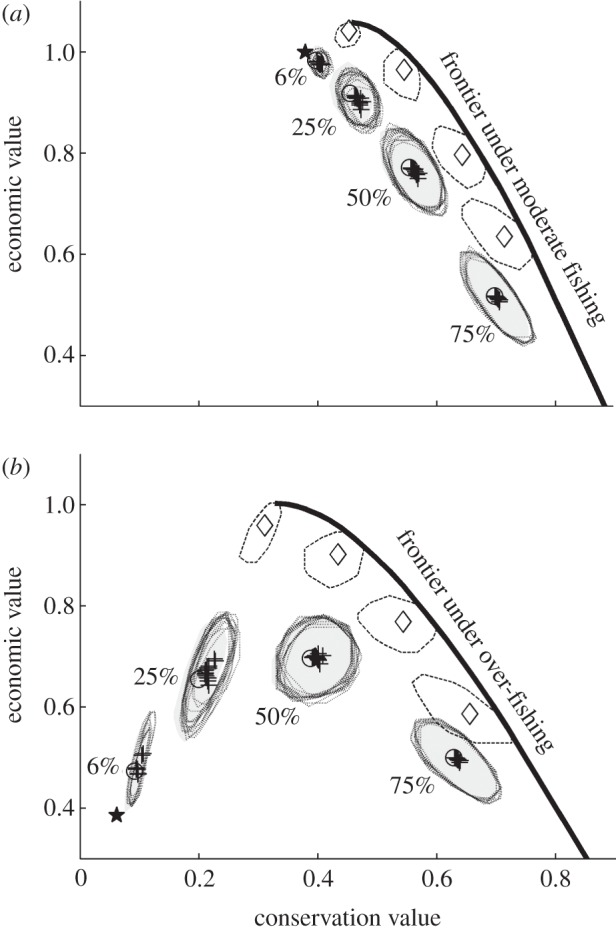
A comparison of likely outcomes of alternative methods for guiding MPA design under (a) moderate fishing and (b) overfishing. Expected outcomes with fixed total MPA coverage are marked with circles (shaded areas indicate 95% probability contours). Expected outcomes under various minimum spacing and habitat coverage guidelines are marked with crosses (95% contour indicated with dotted lines). Expected outcomes if MPA networks are guided by model optimized ‘seeded’ proposals are marked with diamonds (95% probability contours marked with dashed lines).
More sophisticated rules-of-thumb guidelines specify the size of MPAs and the maximum spacing between them. These rules seek to distil complex information about organisms’ movement into simple rules for design [25], although their effectiveness has recently been questioned [26]. We examined a range of guidelines with rules governing spacing of MPAs and the minimum amount of habitat protected in each (electronic supplementary material, Methods); rules which are similar to those used in California's Marine Life Protection Act process [21]. We found that networks meeting these guidelines are little better than random (figure 3, dotted shapes). The most dramatic effect of guidelines is to effectively exclude small MPA networks, implicitly setting a minimum coverage rule. Thus, guidelines can improve outcomes under overfishing, but do not outperform simple rules specifying total MPA coverage. Several recent and ongoing MPA network planning processes around the world incorporate similar guidelines [21,27,28], and we provide the first demonstration that this strategy is likely to fall short of its goals in nearly all cases.
Our results indicate that rule-of-thumb types of guidelines are ultimately an inefficient way to incorporate science into the design of marine spatial plans. The strength of spatially explicit models is their ability to rapidly find optimal designs from among an astronomical number of options. Here, we propose a novel method for incorporating models into a stakeholder-driven design process. Models can be used to identify efficient networks, which then serve as the starting points for stakeholder participation in the design process. Even if stakeholders substantially alter these initial networks, then the results are still near-optimal. Expected outcomes are within a few per cent of the efficiency frontier, and 95% of possible outcomes are better than the likely outcomes of guideline-based designs (figure 3, solid shapes). By using this approach to MPA network design, stakeholders gain maximum benefit from the predictive power of the models while retaining the freedom to choose among millions of possible networks, accounting for goals and details not captured by the models, and thus improving the final plan.
Although the case study presented here is simplified, it illustrates that even within a perfectly understood system, efficient management options are so rare that they are unlikely to be considered, much less adopted, without the use of model-based optimization. There have been important examples in which models have been used to generate spatial plans [29,30], and if combined with a stakeholder-driven process, the resulting plans will benefit from the models’ ability to find efficient designs from near-infinite options, from the stakeholders’ knowledge of local idiosyncrasies, and from the communities’ increased acceptance of the final plan. Continued development and validation of these models is a key step for achieving efficient planning. In cases where models are not available or sufficiently reliable, placing MPAs without guidance may still be appropriate (particularly under conditions of severe overfishing), but such a process should be undertaken with the awareness that the outcomes will fall far short of their potential, and without the illusion that rule-of-thumb-based scientific guidelines can help identify efficient plans.
Acknowledgements
We thank J. W. White and S. D. Gaines for helpful discussions, S. E. Lester for comments on this manuscript, S. Mitarai, J. R. Watson, C. Dong and J. C. McWilliams for sharing their dispersal estimates, E. Saarman for discussions about guidelines and two anonymous reviewers for comments.
Funding statement
This work was supported by the Waitt Foundation and NSF grant OCE-1155813.
References
- 1.White C, Halpern BS, Kappel CV. 2012. Ecosystem service tradeoff analysis reveals the value of marine spatial planning for multiple ocean uses. Proc. Natl Acad. Sci. USA 109, 4696–4701 (doi:10.1073/pnas.1114215109) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Rassweiler A, Costello C, Siegel DA. 2012. Marine protected areas and the value of spatially optimized fishery management. Proc. Natl Acad. Sci. USA 109, 11 884–11 889 (doi:10.1073/pnas.1116193109) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Gopnik M, Fieseler C, Cantral L, McClellan K, Pendleton L, Crowder L. 2012. Coming to the table: early stakeholder engagement in marine spatial planning. Mar. Policy 36, 1139–1149 (doi:10.1016/j.marpol.2012.02.012) [Google Scholar]
- 4.Pomeroy R, Douvere F. 2008. The engagement of stakeholders in the marine spatial planning process. Mar. Policy 32, 816–822 (doi:10.1016/j.marpol.2008.03.017) [Google Scholar]
- 5.UNEP WCMC. 2008. National and regional networks of marine protected areas: a review of progress. Cambridge, UK: UNEP World Conservation Monitoring Centre.
- 6.Gilliland PM, Laffoley D. 2008. Key elements and steps in the process of developing ecosystem-based marine spatial planning. Mar. Policy 32, 787–796 (doi:10.1016/j.marpol.2008.03.022) [Google Scholar]
- 7.Ehler C, Douvere F.2009. Marine spatial planning: a step-by-step approach towards ecosystem-based management. Paris, France: UNESCO. [DOI] [PubMed]
- 8.Flannery W, Cinnéide M. 2012. Deriving lessons relating to marine spatial planning from Canada's eastern Scotian shelf integrated management initiative. J. Environ. Policy Plan. 14, 37–41 (doi:10.1080/1523908X.2012.662384) [Google Scholar]
- 9.Arkema KK, Abramson SC, Dewsbury BM. 2006. Marine ecosystem-based management: from characterization to implementation. Front. Ecol. Environ. 4, 525–532 (doi:10.1890/1540-9295(2006)4[525:MEMFCT]2.0.CO;2) [Google Scholar]
- 10.Cowie-Haskell BD, Delaney JM. 2003. Integrating science into the design of the Tortugas ecological reserve. Mar. Technol. Soc. J. 37, 68–81 (doi:10.4031/002533203787537447) [Google Scholar]
- 11.Ferse SCA, Manes Costa M, Manes KS, Adhuri DS, Glaser M. 2010. Allies, not aliens: increasing the role of local communities in marine protected area implementation. Environ. Conserv. 37, 23–34 (doi:10.1017/S0376892910000172) [Google Scholar]
- 12.Agardy T, et al. 2003. Dangerous targets? Unresolved issues and ideological clashes around marine protected areas. Aquat. Conserv. Mar. Freshw. Ecosyst. 13, 353–367 (doi:10.1002/aqc.583) [Google Scholar]
- 13.White JW, et al. 2012. A comparison of approaches used for economic analysis in marine protected area network planning in California. Ocean Coast. Manag. 74, 77–89 (doi:10.1016/j.ocecoaman.2012.06.006) [Google Scholar]
- 14.Lester SE, Costello C, Halpern BS, Gaines SD, White C, Barth JA. 2013. Evaluating tradeoffs among ecosystem services to inform marine spatial planning. Mar. Policy 38, 80–89 (doi:10.1016/j.marpol.2012.05.022) [Google Scholar]
- 15.Smith M. 2003. Economic impacts of marine reserves: the importance of spatial behavior. J. Environ. Econ. Manage. 46, 183–206 (doi:10.1016/S0095-0696(03)00024-X) [Google Scholar]
- 16.Kahui V, Alexander WRJ. 2007. A bioeconomic analysis of marine reserves for paua (abalone) management at Stewart Island, New Zealand. Environ. Resour. Econ. 40, 339–367 (doi:10.1007/s10640-007-9157-9) [Google Scholar]
- 17.Conrad J, Smith M. 2012. Nonspatial and spatial models in bioeconomics. Nat. Resour. Model. 25, 52–92 (doi:10.1111/j.1939-7445.2011.00102.x) [Google Scholar]
- 18.Valderrama D, Anderson J. 2007. Improving utilization of the Atlantic sea scallop resource: an analysis of rotational management of fishing grounds. Land Econ. 83, 86–103 [Google Scholar]
- 19.Watson J, Mitarai S, Siegel D, Caselle J, Dong C, McWilliams J. 2010. Realized and potential larval connectivity in the southern California Bight. Mar. Ecol. Prog. Ser. 401, 31–48 (doi:10.3354/meps08376) [Google Scholar]
- 20.Mitarai S, Siegel DA, Watson JR, Dong C, McWilliams JC. 2009. Quantifying connectivity in the coastal ocean with application to the southern California Bight. J. Geophys. Res. 114, C10026 (doi:10.1029/2008JC005166) [Google Scholar]
- 21.Saarman E, Gleason M, Ugoretz J, Airamé S, Carr M, Fox E, Frimodig A, Mason T, Vasques J. 2013. The role of science in supporting marine protected area network planning and design in California. Ocean Coast. Manag. 74, 45–56 (doi:10.1016/j.ocecoaman.2012.08.021) [Google Scholar]
- 22.Hilborn R, Micheli F, De Leo GA. 2006. Integrating marine protected areas with catch regulation. Can. J. Fish. Aquat. Sci. 63, 642–649 (doi:10.1139/f05-243) [Google Scholar]
- 23.Holland D, Herrera G. 2010. Benefits and risks of increased spatial resolution in the management of fishery metapopulations under uncertainty. Nat. Resour. Model. 23, 494–520 (doi:10.1111/j.1939-7445.2010.00071.x) [Google Scholar]
- 24.Smith MD, Sanchirico JN, Wilen JE. 2009. The economics of spatial-dynamic processes: applications to renewable resources. J. Environ. Econ. Manage. 57, 104–121 (doi:10.1016/j.jeem.2008.08.001) [Google Scholar]
- 25.Shanks AL, Grantham BA, Carr MH, Applications SE, The S, Reserves M. 2003. Propagule dispersal distance and the size and spacing of marine reserves. Ecol. Appl. 13, S159–S169 (doi:10.1890/1051-0761(2003)013[0159:PDDATS]2.0.CO;2) [Google Scholar]
- 26.Moffitt EA, White JW, Botsford LW. 2011. The utility and limitations of size and spacing guidelines for designing marine protected area (MPA) networks. Biol. Conserv. 144, 306–318 (doi:10.1016/j.biocon.2010.09.008) [Google Scholar]
- 27.Fernandes L, et al. 2012. Biophysical principles for designing resilient networks of marine protected areas to integrate fisheries, biodiversity and climate change objectives in the coral triangle. Jakarta, Indonesia: Coral Triangle Support Partnership. [Google Scholar]
- 28.Ardron J. 2008. Three initial OSPAR tests of ecological coherence: heuristics in a data-limited situation. ICES J. Mar. Sci. 65, 1527–1533 (doi:10.1093/icesjms/fsn111) [Google Scholar]
- 29.Sala E, Aburto-Oropeza O, Paredes G, Parra I, Barrera JC, Dayton PK. 2002. A general model for designing networks of marine reserves. Science 298, 1991–1993 (doi:10.1126/science.1075284) [DOI] [PubMed] [Google Scholar]
- 30.Fernandes L, et al. 2005. Establishing representative no-take areas in the great barrier reef: large-scale implementation of theory on marine protected areas. Conserv. Biol. 19, 1733–1744 (doi:10.1111/j.1523-1739.2005.00302.x) [Google Scholar]