

Abstract
Melanin provides a crucial filter for solar UV radiation and its genetically determined variation influences both skin pigmentation and risk of cancer. Genetic evidence suggests that the acquisition of a highly stable melanocortin 1 receptor allele promoting black pigmentation arose around the time of savannah colonization by hominins at some 1–2 Ma. The adaptive significance of dark skin is generally believed to be protection from UV damage but the pathologies that might have had a deleterious impact on survival and/or reproductive fitness, though much debated, are uncertain. Here, I suggest that data on age-associated cancer incidence and lethality in albinos living at low latitudes in both Africa and Central America support the contention that skin cancer could have provided a potent selective force for the emergence of black skin in early hominins.
Keywords: skin cancer, melanin, UVB, albino
1. Introduction
It is increasingly being recognized that evolutionary considerations can shed light on our vulnerability to many common diseases [1,2]. Mismatches between the ancestral human environments that shaped our genetics and current lifestyles provide an evolutionary logic for some of the recognized risk factors for cancer [3–5]. A neglected aspect of these considerations is whether cancer could itself have provided selective pressure for evolutionary change in humans. At first glance, it would seem unlikely as most cancers in contemporary societies impact post-reproductively. There may, however, be one unique historical context in which lethal cancer did contribute to selection and this is with the ancestral origin of black skin in Africa. Skin pigmentation is perhaps the most striking polymorphic trait in humans, eliciting much debate as to its evolutionary origins and genetic basis [6–14]. Dark or black skin lowers the risk of ultraviolet radiation (UVR)-induced skin cancer by several orders of magnitude and, while this might be considered an incidental benefit, here I make a case for lethal skin cancer—in reproductive, young, early humans, as a potent selective force underlying the emergence of black skin as the ancestral pigmentation state.
2. Skin cancer and melanin
Skin cancers come in three unsavoury flavours—basal cell (BCC), squamous cell (SCC) carcinomas and malignant melanoma (MM)—and collectively they represent the commonest cancers of humans, with intense exposures (especially in early life) and chronic exposures being linked to MM and SCC/BCC, respectively [15,16]. The mutagenic impact of solar ultraviolet radiation (UVR), and particularly UVB (290–320 nm), is accepted as the predominant causal factor [17]. This is endorsed both by studies with human skin transplanted onto immune-deficient mice and UV-irradiated [18] and by mathematical modelling which finds a direct (linear regression) relation between the log of skin cancer incidence and total, annual UVR [19]. UVB is directly mutagenic, principally via the formation of unrepaired nucleotide transitions (CC → TT and C → T) at the site of dipyrimidine photoproducts in p53 and other oncogenes [20,21]. Light-skinned individuals are most at risk of all three skin cancer types and the lesions develop almost exclusively on sun-exposed parts of the body [22]. The intrinsic vulnerability of white skin to UVR-induced mutation is exemplified by the extraordinarily high frequency of covert p53 mutant clones in normal, exposed white skin—thousands per individual [23]—and the presence in most benign naevi of mutant V599E B-RAF, the hallmark mutation of melanoma [24]. Were it not for the efficacy of DNA repair of UV-induced DNA damage, those with white skin would all have cancer, and at a very young age, as evidenced by the impact of the inherited disorder of nucleotide excision DNA repair, xeroderma pigmentosum (XP) [25,26]. Black- or dark-skinned ethnic groups are substantially less at risk but when they do have a diagnosis of skin cancer, it is often on soles and palms—less pigmented regions of the body [27,28].
Melanin is the critical, biological effector of UVR damage limitation. Melanins (Greek, melas—black) are a family of aromatic polymers and are widespread in microbes, fungi, plants and animals, serving a wide variety of functions including coloration for sexual display, camouflage, decoy or defence signalling (cuttlefish ink), microbial virulence, innate immunity (in invertebrates), vision and neurological activity [29–31]. Exploiting the photo-protective capacity of melanin was however an early evolutionary innovation. The ability of melanin to absorb ultraviolet and gamma radiation aids survival of microbes and fungi that live in extreme environments, including high altitude and, rather dramatically, on the damaged walls of the now defunct Chernobyl reactor [32]. Additionally, melanized fungal cells can absorb and convert radiation or photon energy into chemical energy to support growth in a manner that parallels chlorophyll function in plants [33]. In the mammalian epidermis, melanin is synthesized from tyrosine, assembled in melanosomes and distributed via the melanocyte–keratinocyte functional unit (figure 1). There, it serves as a physical and chemical filter, absorbing UV(A,B) radiation photons, dissipating the energy as heat and providing a sink for free radicals produced by UVA. Melanin absorbs UVR maximally in the same range of the electro-magnetic radiation spectrum (280–320 nm) as does DNA [35].
Figure 1.

Diagram of melanin genetics, biosynthesis and skin disposition. (a) Simplified view of key molecules in melanin synthesis. Proteins (red) encoded by gene variants implicated in skin cancer via genome-wide association studies. (b) Maturation of melanosomes from round pheomelanosome (left) to mature, dark and ellipsoid eumelanosome (right). Length of melanosome, 0.4–1.0 µm. Electron micrographic images, adapted from reference [34]. Copyright © 1975 Wiley-Liss Inc. Used with permission from [34]. (c) Melanocyte–keratinocyte functional units in epidermis.
In black-skinned individuals, melanocytes synthesize brown/black eumelanin which is then packaged into peri-nuclear distributed, ellipsoid melanosomes of keratinocytes (figure 1). This appears to be a near optimal arrangement for UV filtration and DNA protection. In white skin, melanocytes synthesize a higher proportion of yellow and/or red pheomelanin and this is then assembled into clustered small, circular melanosomes in keratinocytes. The compound effect is minimal UV filtration. Moreover, pheomelanin is more photo-reactive than eumelanin and produces DNA-damaging free radicals when exposed to UV radiation, possibly enhancing carcinogenesis [29]. These inherited differences are determined by multiple genes encoding melanin synthesis and assembly in which the melanocortin receptor 1 (MC1R) plays a pivotal role (figure 1) [6,12,36]. Recent genome-wide associated studies (GWAS) reinforce these causal links by revealing that inherited allelic variants of genes directly involved in pigmentation (e.g. MC1R, tyrosinase gene, OCA-2) are associated with risk of skin cancer [37–39] (figure 1).
Ground levels of solar UVB radiation are influenced by many variables including sun spot flares, Earth's orbital position (relative to the sun), atmospheric ozone levels, clouds and moisture (or aridity), reflection (from snow and ice) and forestation cover [40–42]. The primary determinants of variable global levels and human exposure are however geographical latitude and altitude [41]. In equatorial regions, UVB levels are high all year round. At higher latitudes (north and south), exposure is less and varies with season [43]. The inherent risk of skin cancer is therefore determined by genetically programmed variation in skin pigmentation and geography [19,44]. Actual risk will be modulated by human behavioural variables, related to exposure. The pigmentation pattern of epidermis is, however, dynamic or adaptive and facultative increases in melanin—or tanning—in response to increased UVR exposure, can provide some protection. In this respect, red-haired individuals who have loss of function MC1R variants [35] and synthesize very little eumelanin are poor responders and therefore at increased risk of skin cancer [45].
UVB penetrates through the epidermis as far as the basement membrane interface with the dermis. This is sufficient depth and, depending upon melanization, sufficient energy to damage the DNA of keratinocyte stem/progenitors residing in the basal layer of the epidermis which are the likely cellular targets for BCC and SCC [46–48]. The degree of attenuation of UVR penetration, through the epidermis, by melanin is then critical in determining risk of skin cancer, providing a 500- to 1000-fold protection for black skin compared with white [19].
Globally, variation in skin pigmentation (measured by the proxy of reflectance) in indigenous populations correlates strongly with levels of UVR exposure quantified by satellite measurements, which in turn correlates strongly with latitude [41]. These data provide a persuasive case for the evolution of human skin coloration to be causally linked to UVR exposure [41,49]. This then begs the question of what particular UVR-associated pathology or pathologies would have provided the selective pressure for major evolutionary shifts in pigmentation. For any deleterious impact of UVR to have selective currency in human evolution, it would have to have impacted on survival and/or reproductive fitness.
3. Why pale skin?
Light-coloured or poorly pigmented skin is generally believed to reflect an adaptive shift from black associated with migration into Europe from East Africa some 50 000–80 000 years ago [7,8,50,51]. Five genes involved in skin pigmentation have strong signatures of relatively recent positive selection in Europeans [52]. The selective pressures for this change are uncertain but one prominent explanation has been the advantage of poorly pigmented or pale skin for generating vitamin D and avoidance of rickets in cloudy northern latitudes [53,54] (table 1). This view has been challenged on the grounds that rickets would be a weak selector acting on Neolithic migrants: first, because there could have been adequate vitamin D provision from dietary (mainly fish) sources and, secondly, because rickets only came to prominence at the time of industrialization and urbanization, particularly in the nineteenth century, long after pale skin was adopted [7]. Vitamin D deficiency does have other deleterious impacts including on immune function [66]. An alternative hypothesis is that pale skin is less sensitive to cold and frostbite [67]. Whatever the evolutionary logic, the acquisition of pale skin has become a liability. But only so because pale-skinned Europeans have been subject to either voluntary or enforced migration to much sunnier climes (e.g. Queensland, Australia, and other subtropical zones) and, more recently, have availed themselves of youthful opportunities for intermittent high level sun exposure via inexpensive air travel and recreational holidays in the sun. In this context, skin cancer arises as the consequence of a mismatch between the ancestral environmental conditions that shaped our genetics and skin properties and our current behavioural and social activities [3]. This narrative is reasonably well established. What I address here is another and, in a sense, reciprocal evolutionary aspect of skin coloration and cancer risk.
Table 1.
Adaptive explanations for skin pigmentation as a fitness trait selected by UV radiation exposure levels.
|
light skin favoured at high latitudes (low UVR) |
references | |
|---|---|---|
| vitamin D provisiona | (rickets, osteoporosis)b | [53,54] |
| thermo-regulation | (frostbite) | [7,55] |
| pleiotypic effects of pigmentation genes on other non-skin functions | (?) | [56] |
|
dark skin favoured at low latitudes (high UVR)c |
references | |
| folate levels | (UVA folate photolysis)d | [43,50,57] |
| thermo-regulation | (sweat gland malfunction)e | [7,58] |
| immuno-regulation | (infectious disease) | [59]f, [60]g |
| effective foraging and hunting | (excessive sunburn and eye damage) | [6] |
| vitamin D levels | (vitamin D toxicity, calcification of the aorta, renal failure) | [54] |
| skin permeability | (xeric stress (dehydration), loss of barrier function and anti-microbial function of skin) | [61] |
| pleiotypic effects on other non-skin functions (e.g. eyesight) | (damage to vision) | [62] |
| combinatorial benefits | [63] | |
aFitness benefits.
bDeleterious phenotype.
cOther proffered selective advantages for black skin include camouflage, threat display or for heat absorption [64,65].
dLeading to neural tube defects in foetuses or to impaired spermatogenesis.
eRobins suggests that pale-skinned infants with a thinner epidermis might be particularly vulnerable to sunburn, hyperthermia and dehydration [7].
fWassermann's suggestion was that black skin was an indirect or pleiotypic consequence of adaptive changes to the adrenal gland that enhanced reticuloendothelial activity (and incidentally MSH) [32]. Other suggestions include protection from hypervitaminosis D and improved visual acuity in bright light [57].
gOmoto suggests that melanin could have provided protection from fungal infection and would therefore be a selective force in tropical, humid climates [60].
4. Ancestral environments and skin coloration
The question is whether, historically, skin cancer itself could have provided selective pressure for changes in allele frequency of genes favouring dark pigmentation, i.e. by selective survival of individuals bearing variants in MC1R and/or other genes that provided black pigmentation? Or—why were ancestral Homo sapiens black?
Early hominin evolution in East Africa at some 2–3 Ma was associated with a dramatic loss of the body hair development that is retained by our primate cousins [68,69]. Hair growth was retained on the head—the most UVR-exposed part of the body of a bipedal hominin. Some exotic explanations have been entertained for this dramatic phenotypic shift, including avoidance of fur parasites or of catching fire, a response to wearing clothes or an adaptation to an aquatic way of life [68–72]. But the most likely major adaptive advantage would have been for thermoregulation or facilitation of sweating and heat loss for physically active, hunter–gatherers in the savannah [69,73,74]. But what colour was the exposed skin of the first hairless hominins? Not black it would seem. The skin of our nearest primate relative, the chimpanzee, is, under the fur, essentially pale or white with melanocytes restricted to hair follicles [67]. The exposed and relatively hairless face and hands are also white in infant chimpanzees of three Pan subspecies (but black in Pan paniscus) and they become facultatively pigmented with age [75]. It has therefore been considered very likely, albeit not unambiguously so [76], that the first African hominins to discard hirsutism were also white- or pale-skinned [7,43,50].
Indigenous ethnic groups in East Africa and throughout sub-Saharan Africa all have black, highly melanized skin. So, when and why did hominin skin turn black? A clue comes from the genetics of skin pigmentation. African blacks have a strikingly invariant MC1R gene—compared with the same gene in Europeans where there is substantial allelic (non-synonymous base pair) and functional variation [51]. The MC1R gene is substantially divergent between the root haplotype for human diversity and the sequence in chimpanzees, suggesting low levels of constraint during the approximately 5 Myr of divergent evolution of the two species [51]. MC1R is critical for eumelanin production [37] (figure 1) and its coding region variability underpins adaptive colour changes in some animals [35,77]. The striking and unusual invariance of MC1R in indigenous Africans (maintained in African Americans), as well as low-frequency variation in Papua New Guinea, suggests that there has been very strong (purifying) selective pressure to retain minimal functional variation or loss in an allele that encodes very effective melanization [51] (see electronic supplementary material). But then if the ancestral hominin skin was initially pale or white, we should also conclude that at some stage there was positive advantage for the individual(s) who first expressed the founder MC1R allele. Rogers et al. [68] proposed that, around the time that humans first became hairless savannah-dwellers, the now common allele swept to fixation under the influence of potent environmental selective pressure. They calculate a minimum date for the time since that sweep occurred of approximately 1.2 Myr [68] which is not too discordant with the time estimate of 1.7 Ma when hominins (Homo ergaster or Homo erectus) were believed to have first inhabited the savannah [78]. The adoption of the more open savannah as a habitat may have resulted from climate change and consequential shifts in forestation and faunal species abundance [79]. Adaptation to the physical challenges of life in the savannah appears to have involved, or resulted in, co-option of a mosaic of fitness traits: larger brains, extensive exocrine sweat glands—and black skin [73].
What then might be the natural pressures that brought about selection in favour of black skin and that have maintained its stability in Africa? It is possible, as Charles Darwin [9] and, more recently, Jared Diamond [10,80] have argued, that the features of hair loss, black and white skin are not the products of natural selection at all but rather incidental variations endorsed by mate preference and sexual selection (box 1). Most, however, favour the view that black skin was acquired relatively early in human evolution as an adaptation to limit UV radiation damage [7,43,50,73,76]. An early proposal along these lines came in a paper delivered by Everard Home to The Royal Society in London in 1820: ‘On the black rete mucosum of the Negro, being a defence against the scorching effect of the sun's rays’ [83].
Box 1. Is it just sex?
‘Skin cancer and sunburn cause little debilitation and few deaths. As agents of natural selection they have an utterly trivial impact …’
(From The rise and fall of the third chimpanzee (Jared Diamond [10, p. 99]))
Is genetically determined skin colour, pale or pigmented, an adaptive trait—a product of UVR-based natural selection, or not? Jared Diamond [10] suggests not, based mainly on the inconsistency of pigmentation with latitude or geography—for example, the retention of black skin in Tasmanian aborigines (for ∼40 000 years) and the lack of acquisition of black coloration in indigenous migrants to the Andes some 10 000 years ago [10,80]. He favours, as does Aoki [81], prevalence of skin colouration dependent not on classical Darwinian natural or environmentally-based selection but on Darwinian sexual selection, i.e. mate preferences, as with the male peacock's exotic tail (or rather back) feathers' exuberance [9]. So, for white-skinned Europeans, the argument is that a population bottleneck resulted in only a very few surviving, or founder, migrants who just happened to be white and this coloration then resulted in a sexual preference which maintained the dominant phenotypic trait of pale skin.
In taking this view, Diamond sought endorsement from Charles Darwin's expressed view that: ‘none of the differences between the races of man are of any direct or special service to him’ [9]. Actually, Darwin's views about skin coloration were more nuanced, and he certainly tried to identify an adaptive rationale. He considered the possibility that dark skin was protective against tropical, infectious diseases but was unpersuaded by the evidence [9]. He also considered the deleterious effects of sun exposure might be selective but concluded: ‘Whether the saving of skin from being thus burnt is of sufficient importance to account for a dark tint have been gradually acquired by man through natural selection, I am unable to judge’. In the end he did opt for selection based on sex, as he did for the loss of body hair: ‘The view that seems to me most probable is that man, or rather primarily woman, become divested of hair for ornamental purposes’ [9]. Diamond is therefore in good company. But read the fine print and Diamond himself says something else [10]. Namely that black skin probably was the result of natural selection (via an uncertain mechanism) but that subsequent variation in skin pigmentation and sexual selection has made the global correlation of skin coloration and UV exposure inconsistent. It would appear very plausible that, as in evolution in general [82], sexual selection has acted on a prior background of adaptive or natural selection, the outcome being the global patterns of graded skin coloration we see today.
5. Adaptive significance of black skin
Table 1 lists candidate UVR-associated pathologies that have been suggested as adaptive rationales for dark-skin pigmentation. Many authors [10,22,29,50,58,63,84,85] have, in the past, and in a somewhat cursory fashion, considered skin cancer as a potential selective force but dismissed it on the grounds that when potentially lethal, in relatively young adults (MM), it is rare and when common (BCC, SCC) it is usually not fatal and occurs predominantly in later life. That argument relates to a longstanding view, expressed by JBS Haldane, Peter Medawar and others, that phenotypes associated with ageing lie outside the reach of natural selection. Notwithstanding the ‘grandmother’ effect that could operate post-reproductively via kin selection [6,86], the observation that BCC and SCC are mostly benign or post-reproductive in character is misleading. The observation is essentially correct in the context of contemporary white-skinned Caucasians inhabiting temperate United States [44] or subtropical zones (for example Queensland, Australia, at 10–20° latitude) [22]. In those settings, the population lives mainly indoors, wears hats and clothes, and has some awareness of risk and access to preventive measures and medical intervention. The ancestral circumstances and culture of the early hominins would have been very different. The relevant, though demanding, questions are these: what might have been the frequency and age of onset of skin cancer in early pale-skinned and naked ancestral humans in tropical (0–10° latitude) East Africa at some 1–2 Ma? And what impact might it be expected to have had on survival and reproductive fitness? We cannot know but there is a striking but hitherto neglected clue to what the answer could be. This comes from a natural experiment—skin cancer in present day albinos with similar high levels of UVR exposure.
6. Skin cancer in albinos
Albinism has long been recognized, an early account being provided by Pliny in the first century AD (quoted in Witkop et al. [87]). Individuals with albinism are visually obvious and exceptional and as such have been the object of mythology, bigotry and cruel exploitation [88–91]. The term albinism (Latin, alba—white) refers to a group of inheritable disorders of melanin pigmentation but with normal numbers and disposition of melanocytes in skin (and eyes and hair) [87,88,92]. The many Nigerian languages each have a name for albino including, in Efik, mbakara-obot-ikot—white man from the bush [93]. Albinism in black ethnic groups throughout sub-Saharan Africa is relatively common—some 1 : 5000 compared with 1 : 20 000 in Europe and USA [88,94]—with some groups, including the Ibo in Nigeria [95] and the Tonga in Zimbabwe [96] having high rates of approximately 1 in 1000 (figure 2). Over 100 years ago, Watkins-Pitchford [99] suggested that the albino should display the greatest risk of all to cancer. Animal modelling endorses his concerns. Albino mice subjected to a single dose of UV irradiation equivalent to sunburning develop very high rates of melanoma when exposed before six weeks of age. SCC occurred at high rates with a second dose in young mice [100] or in adults with chronic, suberythemal irradiation [101]. Control, wild-type mice were completely resistant to these exposures.
Figure 2.

Tanzanian African albino. Copyright Reuters.
The most frequent genetic basis for albinism in Africa is via homozygosity (often via culturally imbedded inbreeding) for an autosomal recessive variant of the gene OCA2 (oculocutaneous albinism type II), which encodes the P protein critically involved in pigmentation [8,102] (figure 1) and whose natural variants in Europeans are linked to skin cancer risk in GWAS [39,103]. Most African albinos have the same 2.7 kb deletion OCA2 mutation, indicative of a founder allele arising before the dispersion of ethnic subgroups throughout Africa 2000–3000 years ago [104]. The same mutation is commonly found in albinos in America of African or mixed African descent [105]. OCA2 type II, unlike type I, retains tyrosinase activity and therefore OCA2 albinos do synthesize pheomelanin (see figure 1) and have a variable capacity to develop pigmented patches or ephelides on exposed skin which may, as in normal white skin, provide some protection from UV damage and cancer [106,107]. Early studies showed that isolated skin of an albino was similar to that of white Europeans in UVB transmission capacity but very different from that of African black skin [108].
There are no population-based databases that provide for accurate age incidence rates of skin cancer in African albinos. However, multiple clinical reports testify to the fact that the prevalence of skin cancer in African albinos, though variable according to geographical region, is exceptionally high in low-latitude (5–10°) regions with high year-round UVB exposures, including Tanzania [97,109–111], Cameroon [112] and Nigeria [93,113,114]. In South Africa, skin cancer rates in albinos vary with latitude and altitude, being relatively high in Soweto and the Transvaal and lower in the Transkei [107,115]. The risk of developing skin cancer in Soweto albinos was estimated to be some 1000 times that of pigmented blacks [116]. Erythema and burns occur in infant albinos and focal skin lesions develop as early as 5 years of age [97]. By the age of 20 years, most albino individuals in low-latitude regions have multiple actinic keratoses (figure 3) [97], the precursor lesions for SCC [117,118]. Many of these regress spontaneously but most, if not all albinos, have overt skin cancer in their twenties or thirties [97,115,119,120], with occasional presentation even in childhood [97]. Most cases are SCC, which accords with sustained or chronic, high level UVB exposure [22] and the prevalence of actinic keratoses. The latter and their ulcerating malignant progeny develop mainly on the face and shoulders, the parts of the body most obviously exposed [97,109,119,121] (figure 3). Africans with albinism are usually poorly educated and as young adults have outdoor trades or are peasant farmers with increased sun exposure [97,98]. Therefore not only is SCC in African albinos very common or even ubiquitous but the age incidence curve is shifted markedly to the left compared with the same cancer type in white Caucasians in the USA and is not very different from the age incidence curve of SCC in white-skinned individuals with XP—a defect in DNA damage repair (figure 4) [122]. XP in African blacks is rare but is also associated with high rates of SCC at a young age [123]. In 512 albinos investigated in Nigeria [119], 50% had actinic keratoses by 14 years of age and 50% had biopsy-confirmed carcinoma by 26 years of age. No albino in this cohort was older than 40 and almost all died of skin cancer [87,88]. In another series (125 cases) of African albinos with skin cancer in Tanzania recorded in 1985, 50% of cases had a diagnosis by 20 years of age and 80% by 30 years [97]. Note also that age of onset is earlier than those data indicate as diagnosis usually occurs a few years after onset of symptoms [98]. Many patients present with advanced disease (figure 3b) [124], this being variously ascribed to rural residence, poverty, self-medication and resort to traditional medicines [110]. Melanoma in albinos is rare [110] but does occur and may be under-diagnosed as it is usually amelanotic [125,126]. BCC is also relatively common [127] and both this and SCC appear to be both multiple and more likely to be recurrent and malignant than their counterparts in Caucasians [97,119,127,128]. Distant metastases of SCC to liver or bone may occur [97] and many albino patients, if untreated, have advanced, invasive lesions involving cartilage, bone and muscle [97,109]. The time interval between development of overt SCC in African albinos and localized invasion or metastasis and patients’ death is around 4 years in the largest series recorded to date [97], which suggests that most albinos, if untreated, will die of advanced SCC in their twenties. Not only is diagnosis of skin cancer in albino Africans delayed in many cases by several years from onset but surveillance and surgical or radio-therapeutic intervention are not systematically managed or affordable. Patients are frequently lost to follow up [97,110]. Accurate mortality rates are not available but few albinos (less than 10%) in equatorial areas of Africa live beyond their third decade [93,97,121,129], and skin cancer is considered to be the major determinant of their limited lifespan [97,112,119,129]:
‘… in the day-time they care not to go abroad. …When moon-shiny night's come, they are all life and activity…’. They were but short-lived.’
Figure 3.
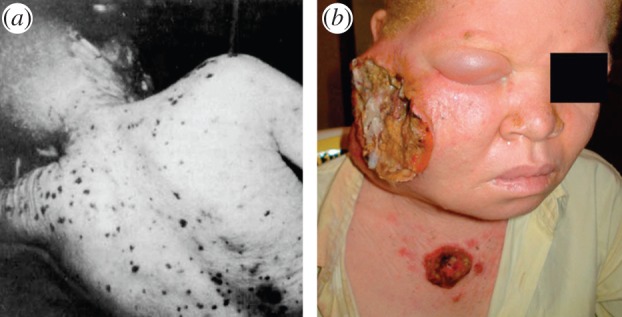
African albinos and skin cancer. (a) Multiple keratoses. Copyright © 1985 Wiley-Liss Inc. Used with permission from [97]. (b) Advanced squamous cell carcinoma on face of 28-year old albino female from Nigeria [98].
Figure 4.
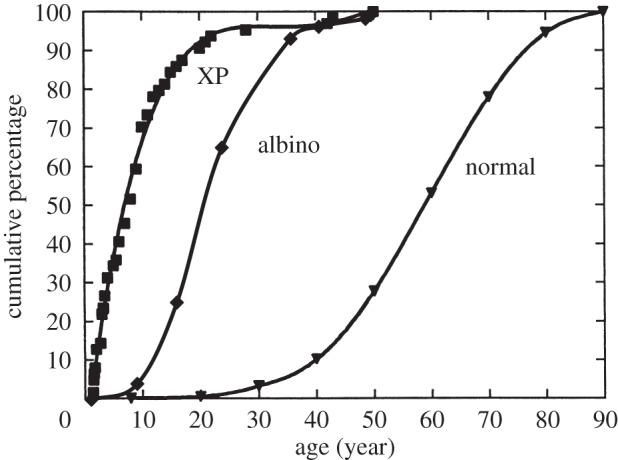
Age of onset of skin cancers in normal, XP and albino skin cancer patients. Adapted from Kraemer et al. [122] with permission from Copyright © 1997 National Academy of Sciences, USA. Albino data derived from a total of 189 cases in [97,110] (from two different regions of Tanzania). Age of onset, rather than diagnosis, is given as in original data [122]. To adjust for this, 2 years have been subtracted from the age of diagnosis of skin cancer in African albinos [97,110].
(From a 1699 account of albinism by Lional Wafer (quoted in Witkop et al. [87, p. 303]).)
7. Albinism and skin cancer outside of Africa
If the association between unpigmented skin, chronic UVB exposure and SCC is a consistent relationship then the pattern of UVR-induced SCC morbidity and mortality in African albinos should have parallels elsewhere in the world. Several indigenous or native American groups have exceptionally high rates of albinism [80]. The Kuna indigenous Americans of the San Blas Islands off the coast of Panama have one of the highest rates of OCA2 albinism, some 1 in 150 [130,131], reflecting their isolation and inbreeding [132]. The genetic basis is in a homozygous splice site single nucleotide variant in intron 17 of OCA2 [131]. As in tropical Africa, the Kuna albinos, who also live at low latitude (approx. 10°), suffer from extensive UVR-induced skin damage including multiple keratoses, solar elastoses and both SCC and BCC [130,132–134]. Virtually all albino Kuna have cancerous lesions by the beginning of their third decade; those with SCC usually die of metastases and/or local invasion of tissues and those with BCC from ulceration and infection, and without treatment these cancers are invariably fatal, and at a relatively young age (G. Gaudiano, personal communication, 2013) [133]. As in Africans with albinism, there are no other recorded common causes of premature death other than cancer. There are anecdotal reports of SCC in albinos in other regions of low latitude—Southern India [135], Papua New Guinea [136]—and high altitude—Nepal [137].
The conclusion that can be drawn is that white-skinned albinos with chronic exposure to high UVB levels are, particularly if untreated, more likely than not to die from skin cancer during their reproductively active years. This long-running natural experiment and its experimental parallel with UVR-exposed albino mice endorse the plausibility of the idea that black skin pigmentation could have arisen in early, pale-skinned hominins as a defence against lethal skin cancer.
Several other explanations have been proffered for the adaptive value of black skin in ancestral humans including, in particular, the UV-induced nutrient hydrolysis hypothesis [43] (see table 1 and the electronic supplementary material for references and discussion).
8. Concluding remarks
Extrapolation from the current risk of skin cancer in OCA2 albinos to that of early hominins in equatorial Africa is clearly speculative but if early humans were indeed pale-skinned, they would most probably have similarly suffered substantial affliction during reproductively active years from non-melanoma skin cancers. That skin cancer in African albinos might be germane to considerations of the adaptive significance of dark skin has been noted before [7,11,138,139], but never explored.
The age-related incidence and mortality from skin cancer, both historically and in contemporary albinos, have been modulated by many factors, including lifestyle, occupation and varying degrees of awareness, preventive measures and medical intervention [91]. In these cultural respects, the lethal impact of skin cancer would have been more severe in naked, pale-skinned and outdoor living hominins, dwelling in a habitat with the highest levels of year-round UVB radiation—in open and arid equatorial savannah. It is difficult to imagine a more potent prescription for cancer: maximum, sustained, whole-body carcinogenic exposure (UVB) coupled with minimal attenuation capacity (via melanin). Young hunter–gatherer males might have suffered the greatest UV exposure and risk of cancer. Death would have ensued at a young age from either metastases or localized invasion, ulceration, bleeding and infection. The detrimental impact on reproductive fitness would then have been severe, providing potent pressure for both the selective sweep of the highly stable African MC1R variant, promoting eumelanin synthesis and black skin and its subsequent stable maintenance for more than a million years. This critical gene clearly did diversify in sequence and function in the descendents of most of those migrants that left Africa to populate the rest of the world. In those, the selective pressures via UVR were both relaxed and different.
Acknowledgements
I am grateful to Dr Gioconda Gaudiano (Panama City) for information on skin cancer in the San Blas Kuna albinos.
References
- 1.Nesse RM, Stearns SC. 2008. The great opportunity: evolutionary applications to medicine and public health. Evol. Appl. 1, 28–48 (doi:10.1111/j.1752-4571.2007.00006.x) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Stearns SC. 2012. Evolutionary medicine: its scope, interest and potential. Proc. Biol. Sci. 279, 4305–4321 (doi:10.1098/rspb.2012.1326) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Greaves M. 2000. Cancer: the evolutionary legacy. Oxford, UK: Oxford University Press [Google Scholar]
- 4.Greaves M. 2007. Darwinian medicine: a case for cancer. Nat. Rev. Cancer 7, 213–221 (doi:10.1038/nrc2071) [DOI] [PubMed] [Google Scholar]
- 5.Eaton SB, et al. 1994. Women's reproductive cancers in evolutionary context. Q. Rev. Biol. 69, 353–367 (doi:10.1086/418650) [DOI] [PubMed] [Google Scholar]
- 6.Westerhof W. 2007. Evolutionary, biologic and social aspects of skin color. Dermatol. Clin. 25, 293–302 (doi:10.1016/j.det.2007.05.001) [DOI] [PubMed] [Google Scholar]
- 7.Robins AH. 1991. Biological perspectives on human pigmentation. Cambridge, UK: Cambridge University Press [Google Scholar]
- 8.Jobling MA, Hurles ME, Tyler-Smith C. 2004. Human evolutionary genetics. New York, NY: Garland Science [Google Scholar]
- 9.Darwin C. 1871,2004. The descent of man. London, UK: Penguin Books [Google Scholar]
- 10.Diamond J. 1991. The rise and fall of the third chimpanzee. London, UK: Vintage Press [Google Scholar]
- 11.Jurmain K, Kilgore L, Trevathan W, Ciochon RL. 2013. Introduction to physical anthropology. Stamford, CT: Wadsworth Cengage Learning [Google Scholar]
- 12.Rees JL. 2004. The genetics of sun sensitivity in humans. Am. J. Hum. Genet. 75, 739–751 (doi:10.1086/425285) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Bodmer WF, Cavalli-Sforza LL. 1976. Genetics, evolution and man. San Francisco, CA: W H Freeman [Google Scholar]
- 14.Cavalli-Sforza LL, Menozzi P, Piazza A. 1996. The history and geography of human genes. Princeton, NJ: Princeton University Press [Google Scholar]
- 15.Miller DL, Weinstock MA. 1994. Nonmelanoma skin cancer in the United States: incidence. J. Am. Acad. Dermatol. 30, 774–778 (doi:10.1016/S0190-9622(08)81509-5) [DOI] [PubMed] [Google Scholar]
- 16.Houghton AN, Polsky D. 2002. Focus on melanoma. Cancer Cell 2, 275–278 (doi:10.1016/S1535-6108(02)00161-7) [DOI] [PubMed] [Google Scholar]
- 17.IARC 1992. International agency for research on cancer, solar and ultraviolet radiation, vol. 55 Lyon, France: International Agency for Research on Cancer [Google Scholar]
- 18.Nomura T, et al. 1997. Induction of cancer, actinic keratosis, and specific p53 mutations by UVB light in human skin maintained in severe combined immunodeficient mice. Cancer Res. 57, 2081–2084 [PubMed] [Google Scholar]
- 19.Kollias N, Sayre RM, Zeise L, Chedekel MR. 1991. Photoprotection by melanin. J. Photochem. Photobiol. B Biol. 9, 135–160 (doi:10.1016/1011-1344(91)80147-A) [DOI] [PubMed] [Google Scholar]
- 20.Nakazawa H, English D, Randell PL, Nakazawa K, Martel N, Armstrong BK, Yamasaki H. 1994. UV and skin cancer: Specific p53 gene mutation in normal skin as a biologically relevant exposure measurement. Proc. Natl Acad. Sci. USA 91, 360–364 (doi:10.1073/pnas.91.1.360) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Pfeifer GP, You Y-H, Besaratinia A. 2005. Mutations induced by ultraviolet light. Mutat. Res. 571, 19–31 (doi:10.1016/j.mrfmmm.2004.06.057) [DOI] [PubMed] [Google Scholar]
- 22.Armstrong BK, Kricker A. 2001. The epidemiology of UV induced skin cancer. J. Photochem. Photobiol. B Biol. 63, 8–18 (doi:10.1016/S1011-1344(01)00198-1) [DOI] [PubMed] [Google Scholar]
- 23.Ståhl PL, Stranneheim H, Asplund A, Berglund L, Pontén F, Lundeberg J. 2011. Sun-induced nonsynonymous p53 mutations are extensively accumulated and tolerated in normal appearing human skin. J. Invest. Dermatol. 131, 504–508 (doi:10.1038/jid.2010.302) [DOI] [PubMed] [Google Scholar]
- 24.Poynter JN, Elder JT, Fullen DR, Nair RP, Soengas MS, Johnson TM, Redman B, Thomas NE, Gruber SB. 2006. BRAF and NRAS mutations in melanoma and melanocytic nevi. Melanoma Res. 16, 267–273 (doi:10.1097/01.cmr.0000222600.73179.f3) [DOI] [PubMed] [Google Scholar]
- 25.Friedberg EC, Henning KA. 1993. The conundrum of xeroderma pigmentosum: a rare disease with frequent complexities. Mutat. Res. 289, 47–53 (doi:10.1016/0027-5107(93)90129-4) [DOI] [PubMed] [Google Scholar]
- 26.Hanawalt PC, Sarasin A. 1986. Cancer-prone hereditary diseases with DNA processing abnormalities. Trends Genet. 2, 124–129 (doi:10.1016/0168-9525(86)90201-5) [Google Scholar]
- 27.Halder RM, Bang KM. 1988. Skin cancer in blacks in the United States. Dermatol. Clin. 6, 397–405 [PubMed] [Google Scholar]
- 28.Gohara MA. 2008. Skin cancer in skins of color. J. Drugs Dermatol. 7, 441–446 [PubMed] [Google Scholar]
- 29.Hill HZ. 1992. The function of melanin or six blind people examine an elephant. Bioessays 14, 49–56 (doi:10.1002/bies.950140111) [DOI] [PubMed] [Google Scholar]
- 30.Nosanchuk JD, Casadevall A. 2003. The contribution of melanin to microbial pathogenesis. Cell Microbiol. 5, 203–223 (doi:10.1046/j.1462-5814.2003.00268.x) [DOI] [PubMed] [Google Scholar]
- 31.Goding CR. 2007. Melanocytes: the new black. Int. J. Biochem. Cell Biol. 39, 275–279 (doi:10.1016/j.biocel.2006.10.003) [DOI] [PubMed] [Google Scholar]
- 32.Wember VV, Zhdanova NN. 2001. Peculiarities of linear growth of the melanin-containing fungi Cladosporium sphaerospermum Penz. and Alternaria alternata (Fr.) Keissler. Mikrobiol. Z. 63, 3–12 [PubMed] [Google Scholar]
- 33.Dadachova E, Bryan RA, Huang X, Moadel T, Schweitzer AD, Aisen P, Nosanchuk JD, Casadevall A. 2007. Ionizing radiation changes the electronic properties of melanin and enhances the growth of melanized fungi. PLoS ONE 2, e457 (doi:10.1371/journal.pone.0000457) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Quevedo WCJ, Fitzpatrick TB, Pathak MA, Jimbow K. 1975. Role of light in human skin color variation. Am. J. Phys. Anthropol. 43, 393–408 (doi:10.1002/ajpa.1330430321) [DOI] [PubMed] [Google Scholar]
- 35.Rees JL. 2003. Genetics of hair and skin color. Annu. Rev. Genet. 37, 67–90 (doi:10.1146/annurev.genet.37.110801.143233) [DOI] [PubMed] [Google Scholar]
- 36.Sturm RA. 2002. Skin colour and skin cancer: MC1R, the genetic link. Melanoma Res. 12, 405–416 (doi:10.1097/00008390-200209000-00001) [DOI] [PubMed] [Google Scholar]
- 37.Sturm RA, Teasdale RD, Box NF. 2001. Human pigmentation genes: identification, structure and consequences of polymorphic variation. Gene 277, 49–62 (doi:10.1016/S0378-1119(01)00694-1) [DOI] [PubMed] [Google Scholar]
- 38.Pharoah PDP. 2008. Shedding light on skin cancer. Nat. Genet. 40, 817–818 (doi:10.1038/ng0708-817) [DOI] [PubMed] [Google Scholar]
- 39.Scherer D, Kumar R. 2010. Genetics of pigmentation in skin cancer: a review. Mutat. Res. 705, 141–153 (doi:10.1016/j.mrrev.2010.06.002) [DOI] [PubMed] [Google Scholar]
- 40.Frederick JE, Snell HE, Haywood EK. 1989. Solar ultraviolet radiation at the Earth's surface. Photochem. Photobiol. 50, 443–450 (doi:10.1111/j.1751-1097.1989.tb05548.x) [Google Scholar]
- 41.Chaplin G. 2004. Geographic distribution of environmental factors influencing human skin coloration. Am. J. Phys. Anthropol. 125, 292–302 (doi:10.1002/ajpa.10263) [DOI] [PubMed] [Google Scholar]
- 42.Madronich S, McKenzie RL, Björn LO, Caldwell MM. 1998. Changes in biologically active ultraviolet radiation reaching the Earth's surface. J. Photochem. Photobiol. B Biol. 46, 5–19 (doi:10.1016/S1011-1344(98)00182-1) [DOI] [PubMed] [Google Scholar]
- 43.Jablonski NG, Chaplin G. 2010. Human skin pigmentation as an adaptation to UV radiation. Proc. Natl Acad. Sci. USA 107(Suppl. 2), 8962–8968 (doi:10.1073/pnas.0914628107) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Scotto J, Kopf AW, Urbach F. 1974. Non-melanoma skin cancer among caucasians in four areas of the United States. Cancer 34, 1333–1338 (doi:10.1002/1097-0142(197410)34:4<1333::AID-CNCR2820340447>3.0.CO;2-A) [DOI] [PubMed] [Google Scholar]
- 45.English DR, Armstrong BK, Kricker A, Winter MG, Heenan PJ, Randell PL. 1998. Demographic characteristics, pigmentary and cutaneous risk factors for squamous cell carcinoma of the skin: a case-control study. Int. J. Cancer 76, 628–634 (doi:10.1002/(SICI)1097-0215(19980529)76:5<628::AID-IJC3>3.0.CO;2-S) [DOI] [PubMed] [Google Scholar]
- 46.Youssef KK, Van Keymeulen A, Lapouge G, Beck B, Michaux C, Achouri Y, Sotiropoulou PA, Blanpain C. 2010. Identification of the cell lineage at the origin of basal cell carcinoma. Nat. Cell Biol. 12, 299–305 [DOI] [PubMed] [Google Scholar]
- 47.Jensen UB, Lowell S, Watt FM. 1999. The spatial relationship between stem cells and their progeny in the basal layer of human epidermis: a new view based on whole mount labelling and lineage analysis. Development 126, 2409–2418 [DOI] [PubMed] [Google Scholar]
- 48.Clayton E, Doupé DP, Klein AM, Winton DJ, Simons BD, Jones PH. 2007. A single type of progenitor cell maintains normal epidermis. Nature 446, 185–189 (doi:10.1038/nature05574) [DOI] [PubMed] [Google Scholar]
- 49.Jablonski NG. 2006. Skin: a natural history. Berkeley, CA: Univesity of California Press [Google Scholar]
- 50.Jablonski NG, Chaplin G. 2000. The evolution of human skin coloration. J. Hum. Evol. 39, 57–106 (doi:10.1006/jhev.2000.0403) [DOI] [PubMed] [Google Scholar]
- 51.Harding RM, et al. 2000. Evidence for variable selective pressures at MC1R. Am. J. Hum. Genet. 66, 1351–1361 (doi:10.1086/302863) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Voight BF, Kudaravalli S, Wen X, Pritchard JK. 2006. A map of recent positive selection in the human genome. PLoS Biol. 4, e72 (doi:10.1371/journal.pbio.0040072) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Murray FG. 1934. Pigmentation, sunlight and nutritional disease. Am. Anthropol. 36, 438–445 (doi:10.1525/aa.1934.36.3.02a00100) [Google Scholar]
- 54.Loomis WF. 1967. Skin-pigment regulation of vitamin-D biosynthesis in man. Science 157, 501–506 (doi:10.1126/science.157.3788.501) [DOI] [PubMed] [Google Scholar]
- 55.Post PW, Daniels FJ, Binford RT. 1975. Cold injury and the evolution of ‘white’ skin. Hum. Biol. 47, 65–80 [PubMed] [Google Scholar]
- 56.Deol MS. 1975. Racial differences in pigmentation and natural selection. Ann. Hum. Genet. Lond. 38, 501–503 (doi:10.1111/j.1469-1809.1975.tb00640.x) [DOI] [PubMed] [Google Scholar]
- 57.Branda RF, Eaton JW. 1978. Skin color and nutrient photolysis: an evolutionary hypothesis. Science 201, 625–626 (doi:10.1126/science.675247) [DOI] [PubMed] [Google Scholar]
- 58.Roberts DF. 1977. Human pigmentation: its geographical and racial distribution and biological significance. J. Soc. Cosmet. Chem. 28, 329–342 [Google Scholar]
- 59.Wassermann HP, Heyl T. 1968. Quantitative data on skin pigmentation in South African races. S. Afr. Med. J. 42, 98–101 [PubMed] [Google Scholar]
- 60.Omoto K. 1984. The Negritos: genetic origins and micro-evolution. Acta Anthropogenet. 8, 137–147 [PubMed] [Google Scholar]
- 61.Elias PM, Menon G, Wetzel BK, Williams JW. 2010. Barrier requirements as the evolutionary ‘driver' of epidermal pigmentation in humans. Am. J. Hum. Biol. 22, 526–537 (doi:10.1002/ajhb.21043) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Daniels FJ, Post PW, Johnson BE. 1972. Theories of the role of pigment in the evolution of human races. In Pigmentation: its genesis and control (ed. Riley V.), pp. 13–22 New York, NY: Appleton-Century-Crofts [Google Scholar]
- 63.Blum HF. 1961. Does the melanin pigment of human skin have adaptive value? An essay in human ecology and the evolution of race. Q. Rev. Biol. 36, 50–63 (doi:10.1086/403275) [DOI] [PubMed] [Google Scholar]
- 64.Morison WL. 1985. What is the function of melanin? Arch. Dermatol. 121, 1160–1163 (doi:10.1001/archderm.1985.01660090074017) [PubMed] [Google Scholar]
- 65.Cowles RB. 1959. Some ecological factors bearing on the origin and evolution of pigment in the human skin. Am. Nat. 93, 283–293 (doi:10.1086/282085) [Google Scholar]
- 66.O'Brien MA, Jackson MW. 2012. Vitamin D and the immune system: beyond rickets. Vet. J. 194, 27–33 (doi:10.1016/j.tvjl.2012.05.022) [DOI] [PubMed] [Google Scholar]
- 67.Post PW, Szabó G, Keeling ME. 1975. A quantitative and morphological study of the pigmentary system of the chimpanzee with the light and electron microscope. Am. J. Phys. Anthropol. 43, 435–444 (doi:10.1002/ajpa.1330430325) [DOI] [PubMed] [Google Scholar]
- 68.Rogers AR, Iltis D, Wooding S. 2004. Genetic variation at the MCIR locus and the time since loss of human body hair. Curr. Anthropol. 45, 105–108 (doi:10.1086/381006) [Google Scholar]
- 69.Morris D. 1967. The naked ape. London, UK: Johnathan Cape [Google Scholar]
- 70.Pagel M, Bodmer W. 2003. A naked ape would have fewer parasites. Biol. Lett. 270, S117–S119 (doi:10.1098/rsbl.2003.0041) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Kushlan JA. 1985. The vestiary hypothesis of human hair reduction. J. Hum. Evol. 14, 29–32 (doi:10.1016/S0047-2484(85)80092-0) [Google Scholar]
- 72.Hamilton WJ. 1973. Life‘s colour code. New York, NY: McGraw-Hill [Google Scholar]
- 73.Zihlman A, Cohn BA. 1988. The adaptive response of human skin to the savanna. Hum. Evol. 3, 397–409 (doi:10.1007/BF02447222) [Google Scholar]
- 74.Montagu A. 1964. Natural selection and man's relative hairlessness. J. Am. Med. Assoc. 187, 356–357 (doi:10.1001/jama.1964.03060180042012) [DOI] [PubMed] [Google Scholar]
- 75.Napier JR, Napier PH. 1967. A handbook of living primates, pp. 239–240 New York, NY: Academic Press [Google Scholar]
- 76.Quevedo WCJ, Fitzpatrick TB, Jimbow K. 1985. Human skin color: origin, variation and significance. J. Hum. Evol. 14, 43–56 (doi:10.1016/S0047-2484(85)80094-4) [Google Scholar]
- 77.Hoekstra HE, Hirschmann RJ, Bundey RA, Insel PA, Crossland JP. 2006. A single amino acid mutation contributes to adaptive beach mouse color pattern. Science 313, 101–104 (doi:10.1126/science.1126121) [DOI] [PubMed] [Google Scholar]
- 78.Klein RG. 1999. The human career: human biological and cultural origins, 2nd edn. Chicago, IL: University of Chicago Press [Google Scholar]
- 79.Bobe R, Behrensmeyer AK, Chapman RE. 2002. Faunal change, environmental variability and late Pliocene hominin evolution. J. Hum. Evol. 42, 475–497 (doi:10.1006/jhev.2001.0535) [DOI] [PubMed] [Google Scholar]
- 80.Diamond J. 1988. Survival of the sexiest. Discover 9, 74–81 [Google Scholar]
- 81.Aoki K. 2002. Sexual selection as a cause of human skin colour variation: Darwin's hypothesis revisited. Ann. Ham. Biol. 29, 589–608 (doi:10.1080/0301446021000019144) [DOI] [PubMed] [Google Scholar]
- 82.Fisher RA. 1930. The genetical theory of natural selection. Oxford, UK: Clarendon Press [Google Scholar]
- 83.Home E. 1821. On the black rete mucosum of the Negro, being a defence against the scorching effect of the sun's rays. Phil. Trans. R. Soc. Lond. 111, 1–6 (doi:10.1098/rstl.1821.0001) [Google Scholar]
- 84.Elias PM, Menon G, Wetzel BK, Williams JW. 2010. Barrier requirements as the evolutionary ‘driver’ of epidermal pigmentation in humans. Am. J. Hum. Biol. 22, 526–537 (doi:10.1002/ajhb.21043) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Rees JL, Flanagan N. 1999. Pigmentation, melanocortins and red hair. Q. J. Med. 92, 125–131 (doi:10.1093/qjmed/92.3.125) [DOI] [PubMed] [Google Scholar]
- 86.Hawkes K. 2010. How grandmother effects plus individual variation in frailty shape fertility and mortality: guidance from human-chimpanzee comparisons. Proc. Natl Acad. Sci. USA 107(Suppl. 2), 8977–8984 (doi:10.1073/pnas.0914627107) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Witkop CJ, Jr, Quevedo WCJ, Fitzpatrick TB. 1983. Albinism and other disorders of pigment metabolism. In The metabolic basis of inherited disease (eds Stanbury JB, Wyngaarden JB, Fredrickson DS, Goldstein JL, Brown MS.), pp. 301–346, 5th edn New York, NY: McGraw-Hill Book Company [Google Scholar]
- 88.King RA. 1998. Genetic hypomelanoses: disorders characterized by generalized hypomelanoses. In The pigmentary system: physiology and pathophysiology (eds Nordlund JJ, Boissy RE, Hearing VJ, King RA, Ortonne J-P.), pp. 553–575 New York, NY: Oxford University Press [Google Scholar]
- 89.Cruz-Inigo AE, Ladizinski B, Sethi A. 2011. Albinism in Africa: stigma, slaughter and awareness campaigns. Dermatol. Clin. 29, 79–87 (doi:10.1016/j.det.2010.08.015) [DOI] [PubMed] [Google Scholar]
- 90.Jeambrun P. 1998. Oculocutaneous albinism: clinical, historical and anthropological aspects. Arch. Pediatr. 5, 896–907 (doi:10.1016/S0929-693X(98)80136-X) [DOI] [PubMed] [Google Scholar]
- 91.Woolf CM. 2005. Albinism (OCA2) in Amerindians. Yrbk Phys. Anthropol. 48, 118–140 (doi:10.1002/ajpa.20357) [DOI] [PubMed] [Google Scholar]
- 92.Witkop CJ., Jr 1971. Albinism. In Advances in human genetics 2 (eds Harris H, Hirschhorn K.), pp. 61–142 New York, NY: Plenum Press; [PubMed] [Google Scholar]
- 93.Okoro AN. 1975. Albinism in Nigeria: a clinical and social study. Br. J. Dermatol. 92, 485–492 (doi:10.1111/j.1365-2133.1975.tb03116.x) [PubMed] [Google Scholar]
- 94.Hong ES, Zeeb H, Repacholi MH. 2006. Albinism in Africa as a public health issue. BMC Public Health 6, 212 (doi:10.1186/1471-2458-6-212) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Cervenka J, Witkop CJ, Jr, Okoro AN, King RA. 1979. Chromosome breaks and sister chromatid exchanges in albinos in Nigeria. Clin. Genet. 15, 17–21 [DOI] [PubMed] [Google Scholar]
- 96.Lund PM, Puri N, Durham-Pierre D, King RA, Brilliant MH. 1997. Oculocutaneous albinism in an isolated Tonga community in Zimbabwe. J. Med. Genet. 34, 733–735 (doi:10.1136/jmg.34.9.733) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Luande J, Henschke CI, Mohammed N. 1985. The Tanzanian human albino skin. Cancer 55, 1823–1828 (doi:10.1002/1097-0142(19850415)55:8<1823::AID-CNCR2820550830>3.0.CO;2-X) [DOI] [PubMed] [Google Scholar]
- 98.Opara KO, Jiburum BC. 2010. Skin cancers in albinos in a teaching hospital in eastern Nigeria: presentation and challenges of care. World J. Surg. Oncol. 8, 73 (doi:10.1186/1477-7819-8-73) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Watkins-Pitchford W. 1910. Pigmentation and cancer. Tvl. Med. J. 6, 23–25 [Google Scholar]
- 100.Noonan FP, Recio JA, Takayama H, Duray P, Anver MR, Rush WL, De Fabo EC, Merlino G. 2001. Neonatal sunburn and melanoma in mice. Nature 413, 271–272 (doi:10.1038/35095108) [DOI] [PubMed] [Google Scholar]
- 101.Noonan FP, Otsuka T, Bang S, Anver MR, Merlino G. 2000. Accelerated ultraviolet radiation-induced carcinogenesis in hepatocyte growth factor/scatter factor transgenic mice. Cancer Res. 60, 3738–3743 [PubMed] [Google Scholar]
- 102.Kedda MA, Stevens G, Manga P, Viljoen C, Jenkins T, Ramsay M. 1994. The tyrosinase-positive oculocutaneous albinism gene shows locus homogeneity on chromosome 15q11-q13 and evidence of multiple mutations in southern African negroids. Am. J. Hum. Genet. 54, 1078–1084 [PMC free article] [PubMed] [Google Scholar]
- 103.Nan H, Kraft P, Hunter DJ, Han J. 2009. Genetic variants in pigmentation genes, pigmentary phenotypes, and risk of skin cancer in Caucasians. Int. J. Cancer 125, 909–917 (doi:10.1002/ijc.24327) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Stevens G, Ramsay M, Jenkins T. 1997. Oculocutaneous albinism (OCA2) in sub-Saharan Africa: distribution of the common 2.7-kb P gene deletion mutation. Hum. Genet. 99, 523–527 (doi:10.1007/s004390050400) [DOI] [PubMed] [Google Scholar]
- 105.Durham-Pierre D, Gardner JM, Nakatsu Y, King RA, Francke U, Ching A, Aquaron R, del Marmol V, Brilliant MH. 1994. African origin of an intragenic deletion of the human P gene in tyrosinase positive oculocutaneous albinism. Nat. Genet. 7, 176–179 (doi:10.1038/ng0694-176) [DOI] [PubMed] [Google Scholar]
- 106.Bothwell JE. 1997. Pigmented skin lesions in tyrosinase-positive oculocutaneous albinos: a study in black South Africans. Int. J. Dermatol. 36, 831–836 (doi:10.1046/j.1365-4362.1997.00259.x) [DOI] [PubMed] [Google Scholar]
- 107.Kromberg JGR, Castle D, Zwane EM, Jenkins T. 1989. Albinism and skin cancer in southern Africa. Clin. Genet. 36, 43–52 (doi:10.1111/j.1399-0004.1989.tb03365.x) [DOI] [PubMed] [Google Scholar]
- 108.Thomson ML. 1955. Relative efficiency of pigment and horny layer thickness in protecting the skin of Europeans and Africans against solar ultraviolet radiation. J. Physiol. 127, 236–246 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Alexander GA, Henschke UK. 1981. Advanced skin cancer in Tanzanian albinos: preliminary observations. J. Natl Med. Assoc. 73, 1047–1054 [PMC free article] [PubMed] [Google Scholar]
- 110.Mabula JB, et al. 2012. Skin cancers among albinos at a university teaching hospital in northwestern Tanzania: a retrospective review of 64 cases. BMC Dermatol. 12, 5 (doi:10.1186/1471-5945-12-5) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Lookingbill DP, Lookingbill GL, Leppard B. 1995. Actinic damage and skin cancer in albinos in northern Tanzania: findings in 164 patients enrolled in an outreach skin care program. J. Am. Acad. Dermatol. 32, 653–658 (doi:10.1016/0190-9622(95)90352-6) [DOI] [PubMed] [Google Scholar]
- 112.Aquaron R. 1990. Oculocutaneous albinism in Cameroon. Ophthal. Paediatr. Genet. 11, 255–263 (doi:10.3109/13816819009015711) [DOI] [PubMed] [Google Scholar]
- 113.Asuquo ME, Ngim O, Ebughe G, Bassey EE. 2009. Skin cancers amongst four Nigerian albinos. Int. J. Dermatol. 48, 636–638 (doi:10.1111/j.1365-4632.2009.04017.x) [DOI] [PubMed] [Google Scholar]
- 114.Datubo-Brown DD. 1991. Primary malignant skin tumour in Nigerians. J. Natl Med. Assoc. 82, 345–348 [PMC free article] [PubMed] [Google Scholar]
- 115.Shapiro MP, Keen P, Cohen L, Murray JF. 1953. Skin cancer in the South African Bantu. Br. J. Cancer VII, 45–57 (doi:10.1038/bjc.1953.5) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Higginson J, Oettlé AG. 1960. Cancer incidence in the Bantu and ‘Cape Colored’ races of South Africa: report of a cancer survey in the Transvaal (1953–1955). J. Natl Cancer Inst. 24, 589–671 [DOI] [PubMed] [Google Scholar]
- 117.Ratushny V, Gober MD, Hick R, Ridky TW, Seykora JT. 2012. From keratinocyte to cancer: the pathogenesis and modeling of cutaneous squamous cell carcinoma. J. Clin. Invest. 122, 464–472 (doi:10.1172/JCI57415) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Alam M, Ratner D. 2001. Cutaneous squamous-cell carcinoma. N. Engl. J. Med. 344, 975–983 (doi:10.1056/NEJM200103293441306) [DOI] [PubMed] [Google Scholar]
- 119.King RA, Creel D, Cervenka J, Okoro AN, Witkop CJ. 1980. Albinism in Nigeria with delineation of new recessive oculocutaneous type. Clin. Genet. 17, 259–270 (doi:10.1111/j.1399-0004.1980.tb00145.x) [DOI] [PubMed] [Google Scholar]
- 120.Cervenka J, Witkop CJJ, Okoro AN, King RA. 1979. Chromosome breaks and sister chromatid exchanges in albinos in Nigeria. Clin. Genet. 15, 17–21 (doi:10.1111/j.1399-0004.1979.tb02024.x) [DOI] [PubMed] [Google Scholar]
- 121.Yakubu A, Mabogunje OA. 1993. Skin cancer in African albinos. Acta Oncol. 32, 621–622 (doi:10.3109/02841869309092440) [DOI] [PubMed] [Google Scholar]
- 122.Kraemer KH. 1997. Sunlight and skin cancer: another link revealed. Proc. Natl Acad. Sci. USA 94, 11–14 (doi:10.1073/pnas.94.1.11) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Chidzonga MM, Mahomva L, Makunike-Mutasa R, Masanganise R. 2009. Xeroderma pigmentosum: a retrospective case series in Zimbabwe. J. Oral Maxillofac. Surg. 67, 22–31 (doi:10.1016/j.joms.2007.09.026) [DOI] [PubMed] [Google Scholar]
- 124.Ofodile A, Oluwasanmi JO. 1979. Management of skin tumors in albinos. Niger. Med. J. 9, 589–591 [PubMed] [Google Scholar]
- 125.Perry PK, Silverberg NB. 2001. Cutaneous malignancy in albinism. Cutis 67, 427–430 [PubMed] [Google Scholar]
- 126.Schulze KE, Rapini RP, Duvic M. 1989. Malignant melanoma in oculocutaneous albinism. Arch. Dermatol. 125, 1583–1586 (doi:10.1001/archderm.125.11.1583b) [DOI] [PubMed] [Google Scholar]
- 127.Oluwasanmi JO, Olufemi Williams A, Alli AF. 1969. Superficial cancer in Nigeria. Br. J. Cancer 23, 714–728 (doi:10.1038/bjc.1969.88) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Itayemi SO, Abioye AA, Ogan O, Daramola JO, Oluwasanmi JO. 1979. Aggressive basal cell carcinoma in Nigerians. Br. J. Dermatol. 101, 465–468 (doi:10.1111/j.1365-2133.1979.tb00028.x) [DOI] [PubMed] [Google Scholar]
- 129.Schrire T. 1958. Aetiology of facial cancer: a speculative and deductive survey. S. Afr. Med. J. 11, 997–1002 [PubMed] [Google Scholar]
- 130.Keeler CE. 1963. Albinism, xeroderma pigmentosum, and skin cancer. Natl Cancer Inst. Monogr. 10, 349–359 [Google Scholar]
- 131.Carrasco A, Forbes EM, Jeambrun P, Brilliant MH. 2009. A splice site mutation is the cause of the high prevalence of oculocutaneous albinism type 2 in the Kuna population. Pigment Cell Melanoma Res. 22, 645–647 (doi:10.1111/j.1755-148X.2009.00575.x) [DOI] [PubMed] [Google Scholar]
- 132.Keeler C. 1970. Cuna moon-child albinism, 1950–1970. J. Hered. 61, 273–278 [DOI] [PubMed] [Google Scholar]
- 133.Keeler C. 1964. The Cuna moon-child syndrome. Dermatol. Trop. 35, 1–11 [DOI] [PubMed] [Google Scholar]
- 134.McFadden AW. 1961. Skin disease in the Cuna Indians. Arch. Dermatol. 84, 175–185 (doi:10.1001/archderm.1961.01580180129020) [Google Scholar]
- 135.Ramalingam VS, Sinnakirouchenan R, Thappa DM. 2009. Malignant transformation of actinic keratoses to squamous cell carcinoma in an albino. Indian J. Dermatol. 54, 46–48 (doi:10.4103/0019-5154.48986) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Hornabrook RW, McDonald WI, Carroll RL. 1980. Congenital nystagmus among the Red-skins of the Highlands of Papua New Guinea. Br. J. Ophthalmol. 64, 375–380 (doi:10.1136/bjo.64.5.375) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Guy PJ. 1990. De-pigmenting skin conditions, cancer, and surgery in Nepal. J. R. Army Med. Corps 136, 122–123 [PubMed] [Google Scholar]
- 138.Parra EJ. 2007. Human pigmentation variation: evolution, genetic basis, and implications for public health. Yrbk Phys. Anthropol. 50, 85–105 (doi:10.1002/ajpa.20727) [DOI] [PubMed] [Google Scholar]
- 139.Fitzpatrick TB. 1964. Introductory lecture. In Recent progress in photobiology (ed. Bowen EJ.), pp. 365–373 Oxford, UK: Blackwell Scientific Publications [Google Scholar]