

Abstract
We examined the effects of morpho-orthographic decomposition on complex word processing using a combination of masked priming and ERP recordings. The process of morpho-orthographic decomposition was primed by the prior presentation of complex non-words (formed by the combination a legal stem and legal affix, e.g. huntity, cornity, scanity) as prime stimuli. Targets were semantically transparent complex words (e.g. hunter), semantically opaque pseudocomplex words (e.g. corner), and simplex words (e.g. scandal) that contained the same stem as primes or a different stem (e.g., huntity-hunter vs. farmity-hunter). We found a large early (150–200 ms) priming effect for transparent complex words only, followed by widely distributed priming effects between 200 and 300 ms and more spatially focused N400 priming effects for all types of target. Furthermore, in the 150–200 ms time-window, the ERP waveforms generated by pseudocomplex words patterned with those of complex words, both of which generated less negative-going waveforms compared with simplex words. In the N400 time-window, on the other hand, complex words differed from both pseudocomplex and simplex words. The results provide further support for early morpho-orthographic segmentation processes that operate independently of semantic transparency, and suggest that the output of these processes only affects the subsequent processing of truly complex words.
Keywords: morphological processing, masked priming, event related potentials, visual word recognition, morpho-orthographic segmentation
Introduction
Morphology is the aspect of language concerned with the structure and formation of words, and a morpheme is often described as the smallest linguistic unit that serves a grammatical function (Aronoff & Fudeman, 2010). Recognizing a word involves determining what the word means on the basis of its form--its orthographic form in the case of reading, or phonological form in the case of listening. Words can be similar in meaning (synonyms) or form (homophones or homographs), but morphologically related words share both form and meaning. Morphologically related words therefore serve as an excellent model for examining the ways in which the representation of form and meaning interact during language comprehension. One standard paradigm used to investigate such issues is the priming paradigm, where a given target word is preceded by different types of prime stimuli, and the influence of these different primes on target word processing is evaluated. Morphologically related primes that share both semantic and orthographic features with targets (hunter-hunt) facilitate processing relative to unrelated primes (singer-hunt). Morphological priming is not simply the product of semantic and orthographic priming, since it can be distinguished from purely orthographic priming on the one hand, and from purely semantic priming on the other. That is, morphological priming produces stronger and longer lasting facilitation than does semantic priming, while orthographic priming tends to result in inhibition (e.g. Drews & Zwitserlood, 1995; Grainger, Colé, & Segui, 1991). Thus, morphological priming effects are typically interpreted as reflecting access to some form of explicit representation of morphological structure during the recognition of morphologically complex words.
One key manipulation in prior research involves the distinction between morphologically related words that are also semantically related (semantically transparent morphological relationship: e.g., farmer, farm), compared with morphologically related words that have no obvious semantic relation (semantically opaque morphological relationship: e.g., apartment, apart) and words that only share a superficial morphological relation (pseudo-morphological relationship: e.g., corner, corn). A clear influence of semantic transparency in driving morphological priming effects has been found in in studies using paradigms in which the primes are consciously available to participants, for example, delayed repetition priming (Marslen-Wilson & Zhou, 1999), cross-modal priming (Marslen-Wilson, Tyler, Waksler, & Older, 1994), and long stimulus onset asynchrony (SOA) priming (Rastle, Davis, Marslen-Wilson, & Tyler, 2000). However, results from many studies using the masked priming paradigm show statistically equivalent priming effects for semantically transparent primes (hunter-hunt) and semantically opaque or pseudocomplex primes (corner-corn) (e.g. Gold & Rastle, 2007; Lavric, Clapp, & Rastle, 2007; Longtin, Segui, & Halle, 2003; McCormick, Rastle, & Davis, 2008; Rastle, et al., 2000; Rastle, Davis, & New, 2004). In the masked priming paradigm, a prime is visually presented for a very brief period, around 30–50 milliseconds, and followed by clearly visible target stimuli. Prime visibility is further reduced by presenting a masking stimulus immediately before primes and, in certain studies, with a brief masking stimulus appearing between prime and target. The short prime duration, as well as the presence of masking stimuli, prevents the subject from consciously perceiving the prime, thereby eliminating any predictive strategies that subjects might use upon noticing the relations between primes and targets. Moreover, the close temporal proximity of the prime and target allow little time for the prime to be processed in isolation from the target, and thus any effects of the prime on responses to the target are presumed to reflect early automatic lexical processes (Forster & Davis, 1984). Therefore the emergence of priming for semantically opaque morphological primes in the masked priming paradigm has been interpreted as reflecting a morphological decomposition procedure that segments visually presented complex words into an orthographically defined stem and affix irrespective of semantic transparency.
Questions nonetheless remain about the point at which semantic information influences the processes involved in complex word recognition. Although many individual studies show statistically non-significant differences in the magnitude of facilitation for semantically transparent and semantically opaque prime–target targets, some studies have reported significant differences (Diependaele, Sandra, & Grainger, 2005; 2009; Feldman, O’Connor, & Del Prado Martín, 2009; Morris, Frank, Grainger, & Holcomb, 2007; for a review see Rastle & Davis, 2008). Moreover, (Crepaldi, Rastle, Coltheart, & Nickels, 2010) have recently shown that irregularly inflected words, which cannot be orthographically decomposed in any simple fashion, prime their base forms more than orthographically matched and semantically unrelated control words. Crepaldi et al. selected nouns and verbs with irregular past tense and plural forms (e.g. fall, mouse) as targets. Each target word was paired with three different primes: the irregular past tense or plural form of the target (e.g. fell-FALL), monomorphemic words that were morphologically unrelated but orthographically related to the targets (e.g. fill-FALL), and finally monomorphemic words that were unrelated either semantically or orthographically to the targets (e.g., hope-FALL). Participants performed a lexical decision task on the targets. Results showed that response times were faster for targets that were preceded by an irregular inflection (fell-FALL) than for those preceded by an orthographically related (fill-FALL) or completely unrelated prime (hope-FALL). Because ‘fell’ and ‘fall’ do not share any orthographically defined morphological units, the ‘fell-FALL’ priming effect cannot be attributed to morpho-orthographic overlap between prime and target, but rather must be attributed to their semantic overlap, providing further support for models of complex word recognition that include a level of morpho-semantic representation located “at some intermediate level between the morpho-orthographic segmentation stage and the semantic system” (Crepaldi, et al., 2010, p. 91).
Grainger and colleagues have proposed a dual-route model of morphological processing (Diependaele, et al., 2005; Diependaele, Morris, Serota, Bertand, & Grainger, in press; Morris, Porter, Grainger, & Holcomb, 2011), in which morphologically complex words activate, in parallel, units at two distinct representational levels: morpho-orthographic and morpho-semantic, the latter via whole-word form representations. Words that share a semantically opaque morphological or pseudo-morphological relationship (e.g. department-depart; corner-corn) prime each other only through shared representations at the morpho-orthographic level. In contrast, words that share a semantically transparent morphological relationship (e.g. hunter-hunt) will not only prime each other via morpho-orthographic representations, but will also be able to prime each other via the shared representations at the morpho-semantic level. This model therefore correctly predicted the pattern of results reported by Crepaldi et al. (2010). The present study provides a further test of this dual-route model, by examining the processing of semantically transparent derived words and semantically opaque pseudo-derived words as targets rather than prime stimuli. Following-up on our previous work we will combine the masked priming procedure with the recording of event-related potentials (ERPs), in order to provide a fine-grained temporal analysis of fast-acting automatic processes involved in the recognition of morphologically complex words
ERPs and masked morphological priming
Prior research using ERPs and masked priming has provided mixed evidence for an early influence of semantic transparency on masked morphological priming. In a study conducted in our lab, Morris et al. (2007) using the lexical decision task and a masked priming paradigm, examined ERPs and reaction times to targets primed by morphologically related words that were semantically transparent (e.g., hunter-hunt), morphologically related words that were semantically opaque (e.g., corner-corn) or orthographically, but not morphologically, related primes (e.g., scandal-scan). In this study both the N250 ERP component, thought to reflect the mapping of sublexical form representations onto whole-word form representations, and the N400 component, thought to reflect the mapping of whole-word form representations onto semantics, were found to be sensitive to semantic transparency (see Grainger & Holcomb, 2009, for a review of N250 and N400 priming effects). Semantically transparent primes generated the largest priming effects, orthographic primes the smallest effects, and semantically opaque primes intermediate effects. However, in an ERP masked priming study testing the same conditions as Morris et al. (2007), Lavric, Clapp and Rastle (2007) found no evidence for an influence of semantic transparency on priming effects on the N400 component. Priming effects were the same for targets preceded by semantically transparent and semantically opaque primes, and there was no priming effect for primes that shared an orthographic but no morphological relation. Finally, in a follow-up study (Morris, Grainger, & Holcomb, 2008), we found evidence for equivalent priming from semantically transparent (complex) and semantically opaque (pseudocomplex) primes between 200–250 ms post-target onset, with opaque priming effects disappearing between 250–300 ms, and priming effects emerging for all types of prime (transparent, opaque, orthographic) on the N400 component. Therefore, as with the evidence obtained from behavioral measures, we can only conclude from the ERP findings that there is evidence that pseudocomplex words (e.g., corner) undergo some form of morpho-orthographic decomposition. However, for the moment it is still not at all clear when semantic transparency begins to influence processing.
This mixed evidence in the ERP signal for an early phase of morpho-orthographic processing might be due to the fact that target stimuli in our prior studies, following the tradition of current research using masked priming to study morphological processing, were all monomorphemic. In these studies, morphological processes operating on complex prime stimuli are inferred as a result of the subsequent effects of such processes during the recognition of simplex targets (see Diependaele, Sandra, & Grainger, 2009; Giraudo & Grainger, 2001, for exceptions). Countering this tradition, however, in a recent study Lavric, Elchlepp, and Rastle (2012) directly compared processing of complex, pseudocomplex, and simplex words in an unprimed lexical decision task with ERP recordings. They found that the ERP waveforms patterned together for complex and pseudocomplex words in an early time-window (190–220 ms post-stimulus onset), both differing from the simplex word ERPs, followed by the complex and simplex words patterning together and differing from the pseudocomplex word ERPs in a slightly later time window (246–276 ms post-stimulus onset). This was taken as evidence for early morpho-orthographic decomposition operating on complex and pseudocomplex words, but not simplex words. In the present study we go one critical step further than in the Lavric et al. (2012) study, by priming the process of morpho-orthographic decomposition prior to the presentation of the different types of target word.
The present study
In the present study we examine responses to complex target words (i.e., words with a true morphological structure such as “hunter”), pseudocomplex target words (i.e., words with a superficial morphological structure such as “corner”), and simplex target words (i.e., words with no morphological structure such as “scandal”) preceded by complex nonword primes that can be related or unrelated to target words. This will allow us to directly examine ERP responses to morphological decomposition as it occurs, or not, during the processing of target words of varying levels of complexity. Rather than priming complex targets with primes that are the stems of the corresponding target, as in the Diependaele et al. (2009) and Giraudo and Grainger (2001) studies, here the complex targets were primed by complex nonword primes formed of a combination of the stem of the target and a suffix that did not generate an existing word (e.g., farmity-farmer). The rationale here is that these nonword primes will induce morpho-orthographic decomposition (Longtin & Meunier, 2005), and we should be able to trace the influence of this morpho-orthographic decomposition during the processing of the complex target in the absence of any morpho-semantic priming. More precisely, in the present study we examined the effects produced by the brief masked presentation of complex nonword primes (e.g., huntity, cornity, scanity) on the recognition of semantically transparent complex words (e.g. hunter), semantically opaque pseudocomplex words (e.g., corner), and simplex words (e.g., scandal). We hypothesize that for each of the three types of target, the complex nonword primes will undergo morpho-orthographic decomposition. We therefore ought to be able to observe the influence of such morpho-orthographic priming during the processing of target words of different levels of complexity.
Figure 1 describes how this hypothesized decomposition of prime stimuli might differentially affect processing of the three types of target tested in the present study. This figure is adapted from Morris et al. (2011) for the specific conditions tested in the present study. First of all, related primes will generate equivalent orthographic priming effects across all target types (arrow linking letters and lexical form representations in Figure 1). Second, related primes will activate embedded stems in the same way for each target type (arrow between letters and morpho-orthographic representations in Figure 1). According to our dual-route model, the earliest possible difference in priming effects across target type will arise at the level of morpho-orthographic representations and the mapping of these representations onto lexical form representations. If, as hypothesized in our model, morpho-orthographic stem representations only send activation to whole-word representations having a semantically transparent morphological relationship with the stem (arrow between morpho-orthographic stem representation and whole-word representation of complex words in Figure 1), then we should see an early difference in the priming effects to complex targets compared with both pseudocomplex and simplex targets. Furthermore, activation of the lexical form representation of the embedded stem should generate inhibitory effects on the processing of both pseudocomplex and simplex targets (lines with filled circles in Figure 1), and morpho-semantic representations should further facilitate processing of complex targets (arrows connecting lexical form and morpho-semantic representations in Figure 1) while having no influence on the processing of pseudocomplex and simplex targets.
Figure 1.
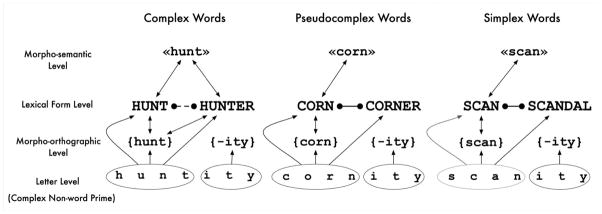
Processing of the different types of target word and corresponding related nonword primes tested in the present study (semantically transparent complex words – hunter, semantically opaque pseudocomplex words – corner, simplex words – scandal) according to our dual-route account of morphological processing. The figure shows how each combination of prime stimulus and target word (e.g., huntity-hunter) is processed at the letter, morpho-orthographic, lexical form, and morpho-semantic levels of representation. Excitatory connections are represented by arrows, and inhibitory connections by filled circles. The dashed inhibitory connection between “hunter” and “hunt” represents the modulation of lateral inhibition by semantic transparency.
Summing up the key predictions of the model presented in Figure 1 to be tested in the present study: 1) in the earliest priming effects (voltage differences between related and unrelated prime conditions) we expect to observe a pattern showing the strongest priming for complex targets compared with both pseudocomplex and simplex targets; 2) comparisons across the different types of target should reveal early morpho-orthographic decomposition for complex and pseudocomplex words, which should both differ from simplex words, whereas later differences should reveal effects of semantic transparency (Lavric, et al., 2012).
Results
Visual Inspection of ERPs
The ERPs for the complex, pseudocomplex and simplex prime conditions from 29 electrode sites are plotted against the ERPs for the corresponding unrelated prime conditions in Figures 2–4. Figure 5 shows the topography of ERP priming effects formed by subtracting ERPs to targets following related primes from ERPs to targets following unrelated primes, separately for the complex, pseudocomplex, and simplex target words. Figure 6 shows the scalp distribution of the effects of target type for the unrelated prime conditions only, in order to remove confounding effects of differences in priming effects across target type. This figure plots the difference in amplitude between ERPs to targets in the (a) complex and simplex conditions (b) pseudocomplex and simplex conditions and (c) pseudocomplex and complex conditions.
Figures 2–4.



ERP waveforms showing the responses to targets following related (solid line), and unrelated (dashed line) primes for complex words (Figure 2), pseudocomplex words (Figure 3) and simplex words (Figure 4). Negative voltages are plotted upward. Prime onset is marked by the vertical calibrating bar, and each tick mark on the x-axis represents 100 msec.
Figure 5.

The spatial distribution and time-course of priming effects for the complex (top), pseudocomplex (middle), and simplex (bottom) target words. These voltage maps were formed from difference waves calculated by subtracting voltages in the related prime condition from voltages in the unrelated prime condition.
Figure 6.

The spatial distribution of the effects of Target Type found in the 150–200 ms time-window (panel A) and the 350–500 ms time window (panel B). For each time-window effects of the three possible pairwise comparisons enabling the calculation of voltage differences are shown: Simplex vs. Complex, Simplex vs. Pseudocomplex, and Pseudocomplex vs. Complex. Only data from unrelated prime conditions were used in these calculations.
ERPs in the epoch from 0–800 ms after the onset of the target produced an initial small negative-going potential peaking between 50 and 100 ms immediately followed by a larger positivity peaking at around 200 ms. Following these early potentials, we observed a series of two negative deflections, the first peaking around 250 ms post-target (N250) and the second at around 400 ms post-target (N400). These two negativities were followed by a large positive deflection (the late positive component, or LPC).
The 150–200 ms time-window
Between 150 and 200 ms after target onset we found a significant Relatedness by Target Type by Anteriority interaction (F(8,176) = 5.9, p = .001) and a significant Relatedness by Target Type by Laterality interaction (F(4,88) = 3.1, p = .03). Follow-up analyses revealed a marginally significant interaction between Relatedness and Target Type at frontal pole sites (α = .01, F(2,44) = 4.23, p = .022), where responses to unrelated targets were significantly more negative than to related targets for complex items only (α = .017, F(1,22) = 7.45, p = 0.012). Given the spatial distribution and timing of this pattern it would appear to reflect an earlier onset of N250 priming for complex words.
Further follow-up analyses examined the effects of Target Type separately for the related and unrelated prime conditions.1 In the unrelated condition there was a significant Target Type by Anteriority interaction (F(8,176) = 3.09, p = .029). When examining each level of anteriority separately, we found an effect of Target Type at occipital sites (α = .01, F(2,46) = 5.54, p = .008). Pairwise comparisons on the occipital data (α = .017) showed a significant difference between Complex and Simplex targets (p = .002), a marginally significant difference between Pseudocomplex and Simplex targets (p = .037), and no significant difference between Complex and Pseudocomplex targets (p > .5). The pattern of effects of Target Type with unrelated primes is shown in panel A of Figure 6.
The 200–300 ms time-window
Between 200 and 300 ms after target onset, in the time-window typically associated with the N250, we found a significant main effect of Relatedness (F(1,22) = 9.43, p = .006), with unrelated primes generating more negative-going waveforms compared to related primes. In addition to the main effect of Relatedness we also found a significant Relatedness by Target Type by Anteriority interaction (F(8,176) = 3.16, p = .026).
In a series of follow-up analyses we examined the Target Type by Anteriority interaction (α = .017) for related and unrelated targets separately. We found that this interaction was not significant for either related or unrelated targets (all ps > .05). Inspection of Figure 5 suggests that the significant Relatedness by Target Type by Anteriority interaction likely reflects the slightly more anterior distribution of priming effects for complex targets plus the clearly posterior distribution of priming effects for simplex targets, even if such distributional differences are not significant when examined separately for each Target Type.
The 350–500 ms time-window
In this time-window we found a significant main effect of Target Type (F(2,44) = 6.41, p = .005). Follow-up analyses revealed that responses to transparent complex targets were less negative than to both pseudocomplex targets (α = .017, F(1,22) = 7.05, p = 0.014) and simplex targets (α = .017, F(1,22) = 17.04, p < 0.001), whereas responses to pseudocomplex and simplex targets did not differ (p = .6). In addition to the effects of Target Type, in this time window we also found significant interactions between Relatedness and Anteriority (F(4,88) = 4.61, p = .025) and Target Type and Anteriority (F(8,176) = 3.22, p = .027). Follow-up analyses (α = .01) revealed that the Relatedness by Anteriority interaction was driven by greater priming effects at posterior sites—a topographic distribution typical of the N400 (Central: F(1,22) = 4.7, p = .041; Parietal: F(1,22) = 10.16, p = .004; Occipital: F(1,22) = 5.7, p = .026). In contrast, the effects of Target Type on the N400 were greater at anterior than at posterior sites. Responses to complex targets were less negative than to those of both pseudocomplex targets (Frontal: F(1,22) = 7.22, p = 0.013; Central: F(1,22) = 11.32, p = .003; Parietal: F(1,22) = 7.72, p = 0.011) and simplex targets (Frontal Pole: F(1,22) = 8.72, p = .007; Frontal: F(1,22) = 14.14, p = .001; Central: F(1,22) = 31.48, p < .001; Parietal: F(1,22) = 6.47, p = .019).
Although we did not find a significant interaction between Target Type and Relatedness in this time window time-window, we examined the effects of Target Type for the unrelated prime condition only in order to remove the possibility that the more positive responses of complex targets were simply caused by priming from related primes (see Figure 6 panel B). We again found a significant main effect of Target Type (F(2,44) = 6.0, p = 0.007). The responses to transparent complex targets (α = .017) were less negative than to simplex targets (F(1,22) = 16.58, p = 0.001). The difference between complex and pseudocomplex targets was also marginally significant (F(1,22) = 5.02, p = 0.036), and responses to pseudocomplex and simplex targets did not differ (p = .35).
Discussion
Semantically transparent complex words (e.g., hunter), semantically opaque pseudocomplex words (e.g., corner), and simplex words (e.g., scandal) served as targets in a masked priming study where primes were formed of the embedded stem/pseudostem/word combined with a suffix to form an non-existing word (huntity, cornity, scanity). Effects of these related primes were measured relative to unrelated primes from the same stimulus category, using EEG recordings while participants monitored target words for animal names. We found priming effects for complex words, but not for pseudocomplex or simplex words in an early time window spanning 150–200ms post-target onset. Following that, all types of target showed widely distributed priming effects in the time window typically associated with the N250 ERP component, and following that, more spatially focused priming effects in the N400 time window. In all cases, related primes produced less negative-going waveforms than unrelated primes. Effects of target word complexity, independently of priming, were found in the 150–200 ms epoch as well as in the N400 epoch. In the early time window, complex target words in the unrelated prime condition patterned with pseudocomplex targets, which both generated less negative-going waveforms than simplex words. In the N400 epoch, on the other hand, pseudocomplex words patterned with simplex words which both generated more negative-going waveforms than complex words (see Figure 6 panel A).
The first important finding of the present study is the significant priming effect that was only seen for semantically transparent complex target words in the earliest time-window, between 150 and 200 ms post-target onset. The ERP waveforms generated by semantically transparent complex targets (e.g., hunter) were significantly more positive-going following a related nonword prime (huntity) compared with unrelated nonword primes (farmity) in frontal electrode sites. Given the timing and the spatial distribution of this effect, we think that it likely reflects an early onset of priming on the N250 component (see Grainger, Lopez, Eddy, Dufau, & Holcomb, 2012, for a similar early onset of N250 priming with an anterior distribution). Given than there were no significant priming effects for either semantically opaque pseudocomplex targets or simplex targets in this time-window, the early priming effect would appear to be driven by processes that are specific to semantically transparent complex words. Within the framework of our dual-route model presented in Figure 1, processing specific to semantically transparent complex words first arises during the mapping of sublexical morpho-orthographic representations onto the whole-word orthographic representations of morphologically complex words. That is, although pseudocomplex words might automatically undergo morpho-orthographic segmentation, the activation of the morpho-orthographic representation of the embedded stem (e.g., “corn” activated upon presentation of “corner”) would not directly influence activation of the whole-word orthographic representation of “corner”. In other words, although the process of morpho-orthographic segmentation might be blind to whether or not the stimulus is truly morphologically complex, the subsequent use of the segments extracted by this process is determined by whether or not the stimulus is truly complex. In line with this proposal is the fact that Diependaele et al. (2009) reported behavioral priming effects with stem primes and complex targets only when targets were transparent complex words.
Following the early priming effect for semantically transparent complex words, we found priming effects for all three types of target in the time window typically associated with the N250 ERP component. This general priming effect could simply be driven by the increased orthographic overlap between primes and targets in the related prime condition. N250 orthographic priming effects with nonword primes have been reported in several prior studies (e.g., Grainger, Kiyonaga, & Holcomb, 2006; Holcomb & Grainger, 2006; see Grainger & Holcomb, 2009, for a review). Orthographically related primes are thought to facilitate the mapping of sublexical form representations onto whole-word form representations, and the efficiency of this mapping is thought to be reflected in a modulation of N250 amplitude (Grainger & Holcomb, 2009). Our results suggest that when targets are not embedded in prime stimuli, as is the case in standard morphological priming (e.g., hunter-hunt; corner-corn; scandal-scan), then N250 priming effects are mainly determined by non-morphological orthographic overlap (i.e., number of letters shared by prime and target). We would nevertheless argue that the early priming effect (150 – 200 ms post-target onset) seen only for complex targets most likely reflects an earlier onset of N250 priming in this condition, rather than priming of a distinct ERP component. This early morphological priming would be driven by the combined influence of letter-level information and morpho-orthographic representations in determining the onset of N250 priming effects (see Figure 1).
In the N400 time-window, we again observed robust priming effects for all target types, which were albeit more spatially focused than the N250 priming effects. Related primes generated a reduced negativity compared with unrelated primes, and the size of this priming effect was comparable for the three types of target word. This can be interpreted as orthographic priming, generated by differences in prime-target orthographic overlap (which was the same across target types), affecting the build-up in activation of whole-word orthographic representations for target words, and the subsequent mapping between such whole-word representations and semantics (Holcomb & Grainger, 2006; Grainger & Holcomb, 2009).
However, independently of priming effects, the nature of the target item differentially affected ERPs in the present study. In the early 150–200 ms time window, we found that in the unrelated prime condition ERPs to complex and pseudocomplex targets patterned together, and both were significantly less negative-going than the ERPs to simplex targets (although only marginally so for the pseudocomplex targets). This result is in line with the pattern reported by Lavric et al. (2012) in an ERP study comparing the processing of these three types of target word without any priming manipulation. Like Lavric et al. (2012), we take this early pattern as reflecting automatic morpho-orthographic decomposition operating on complex and pseudocomplex target words alike. In line with these findings are the results from MEG studies (Lewis, Solomyak, & Marantz, 2011; Solomyak & Marantz, 2010) showing that activity originating in the left fusiform gyrus and peaking between 140 and 190 ms after the onset of a visually presented word (the M170 component) increases as a function of the decomposability of words. Morphological decomposability was measured as the ratio of the stem frequency to the whole word frequency, and the effect of decomposability was found for both transparent complex words and opaque pseudocomplex words.
In our N400 analysis, on the other hand, there was clear evidence that transparent complex targets generated reduced N400 amplitudes compared with both pseudocomplex and simplex targets (see Figure 6 panel B). One way to explain these effects of target type on later processing is in terms of an inhibitory influence of embedded words during the processing of target words containing the embedded item. According to the model presented in Figure 1, these inhibitory effects will be strongest for pseudocomplex and simplex targets, and should be smaller and possibly even absent for transparent complex targets (the dashed line connecting morphologically related whole-word representations in Figure 1). That is, related primes in the pseudocomplex (corner) and simplex (scandal) target conditions provide bottom-up support not only for the target word but also for the embedded word (corn, scan) that will compete with the target for identification, and this competition is thought to be reduced or even absent for complex targets. The fact that this specific pattern of effects of Target Type emerged in the N400 time-window is in line with the hypothesized role of lexical inhibition in driving these effects (see Massol, Grainger, Dufau, & Holocomb, 2010, for evidence for the time-course of lexical inhibition during priming). Alternatively, it could be the morpho-semantic representation that is shared by a complex word and its embedded stem that is the source of the reduced N400 amplitude for these target words compared with both pseudocomplex and simplex words. This shared morpho-semantic representation would be activated by the complex word target and its embedded stem, and send excitatory feedback to both of these representations. Future research should help clarify how mechanisms such as lateral inhibition and top-down feedback might underlie processing differences between complex, pseudocomplex, and simplex target words, and computational modeling will certainly play an important role here.
Summing up, the results of the present study are in line with the predictions of the model shown in Figure 1, according to which sublexical morpho-orthographic decomposition, once it has occurred, will only influence the subsequent processing of transparent complex words. First of all, in the line with the results of Lavric et al. (2012), we found evidence for early morpho-orthographic decomposition in the form of ERP differences between the different types of target in the 150–200 ms epoch. Here both complex and pseudocomplex words patterned together, and differed from simplex words. In the same early time-window, however, ERP priming effects were only evident for complex words. According to the model shown in Figure 1, this arises because the extraction of the stem from complex prime stimuli, which occurs for all target types, can only influence processing of related complex targets (e.g., huntity-hunter). Neither the stem “corn” extracted from “cornity”, nor the stem “scan” extracted from “scanity” can influence processing of related targets words (cornity-corner; scanity-scandal), given the hypothesized absence of connectivity between the relevant representations in the model (see Figure 1). Therefore, although we provide further evidence in favor of rapid morpho-orthographic decomposition that is blind to semantic information (operating on pseudocomplex words like “corner”) and even to lexical status (operating on pseudowords like “cornity”), we also show that subsequent processing can only capitalize on such decomposition when reading truly complex words.
Method
Participants
The participants for this study were 27 adults (16 women). The data from three participants were excluded from analysis, two because of excessive heartbeat artifact and one for excessive alpha activity. All participants were recruited from the Tufts University community and paid for their participation. The participants ranged in age from 18 to 22 years (mean 19.3 years). All were right-handed native English speakers with normal or corrected-to-normal vision, and none reported any linguistic or neurological impairment.
Stimuli
The stimuli for this study were 306 prime-target pairs in which the prime consisted of an existing English root combined with a bound suffix to form a morphologically complex nonword (e.g. ‘hunt’ + ‘-ity’ ⇨ ‘huntity’), and targets were either semantically transparent complex words containing the same root as the prime (e.g. huntity-HUNTER), semantically opaque pseudocomplex words containing the same root as the prime (cornity-CORNER), or simplex words that had the prime word embedded at initial position, but for which the letters following the embedded prime word did not form a common English suffix (scanity-SCANDAL). We constructed two lists each consisting of 306 prime-target pairs. In each list there were 102 semantically transparent complex targets, 102 semantically opaque pseudocomplex targets and 102 simplex targets. Primes were formed by dividing each target into either a stem and affix (in the case of the complex and pseudo-complex words, e.g hunter, corner) or into a possible stem and non-affix letter sequence (in the case of simplex words, e.g. scandal). Each stem was then combined with one of 23 possible affixes, taken from the CELEX English database (Baayen, Piepenbrock & Gulikers, 1995) to from a complex non-word prime. The list of affixes and their frequency of use in primes across the three conditions is given in Table 1.
Table 1.
The list of affixes and the frequency with which each was used to construct primes for each of the three types of target.
| Frequency
|
|||
|---|---|---|---|
| Complex | Pseudocomplex | Simplex | |
| -al | 2 | 2 | 2 |
| -able | 4 | 4 | 4 |
| -age | 4 | 5 | 4 |
| -ance | 6 | 6 | 6 |
| -ant | 5 | 6 | 6 |
| -ard | 6 | 5 | 5 |
| -ary | 7 | 6 | 6 |
| -ate | 3 | 3 | 4 |
| -ence | 3 | 3 | 3 |
| -er | 4 | 3 | 4 |
| -ern | 3 | 3 | 3 |
| -ery | 4 | 4 | 4 |
| -ible | 2 | 3 | 3 |
| -ic | 5 | 5 | 5 |
| -ify | 4 | 4 | 4 |
| -ish | 4 | 5 | 4 |
| -ist | 3 | 3 | 2 |
| -ity | 5 | 5 | 5 |
| -ize | 6 | 7 | 6 |
| -ment | 6 | 6 | 6 |
| -ness | 6 | 5 | 6 |
| -or | 4 | 4 | 4 |
| -ous | 6 | 5 | 6 |
For each type of target, 51 pairs were related in that the stem of the complex nonword prime was the same as the stem of the target, and 51 were unrelated. Related primes in one list were used as unrelated primes in the other list. Therefore, across all participants, each target appeared with both a related and unrelated prime, and the same primes appeared in the related and unrelated conditions, but each participant saw each item in only one condition.
Although we held the targets constant across the related and unrelated conditions, it was obviously not possible to hold the targets constant across the different target type conditions, where the categories in question contain different lexical items. As a result, across target types, targets were not matched on all variables known to affect lexical processing. In particular, affix and non-affix endings varied in frequency. In addition, affix tokens were not equally represented across the morphologically complex and pseudocomplex targets.
We measured semantic transparency using Latent Semantic Analysis (LSA; Landauer & Dumais, 1997). A one-way ANOVA comparing LSA scores for the three types of prime/target pairs revealed a significant difference (F(2,285) = 107.5, p < .001. Post-hoc tests showed that the LSA scores of transparent pairs differed from those of both pseudocomplex (Mdiff = .26 (SE = .02), p < .001), and orthographic pairs (Mdiff = .26 (SE = .02), p < .001) but the scores of the latter two types of prime/target pairs did not differ (Mdiff = .002 (SE = .02), p = 1.0) Across the three conditions, targets were equated for length, cumulative bigram frequency (CBF), neighbourhood size (NS), log frequency (LF), stem length, stem cumulative bigram frequency, stem neighbourhood size, stem log frequency, and suffix length2 (see Table 2). Oneway ANOVAs comparing scores on each of these variables for the three types of prime/target pairs revealed no significant differences (all ps > .1)
Table 2.
Length and frequency characteristics of the related primes and targets. Frequencies (log10 scale) were obtained from the CELEX English lexical database (Baayen, Piepenbrock & Gullikers, 1995). Token frequencies were computed per million.
| Complex | Pseudocomplex | Simplex | ||||
|---|---|---|---|---|---|---|
|
| ||||||
| Mean | (SE) | Mean | (SE) | Mean | (SE) | |
| LSA | 0.32 | (0.02) | 0.06 | (0.01) | 0.06 | (0.01) |
| Target Length | 7.08 | (0.12) | 6.83 | (0.13) | 6.88 | (0.11) |
| Target CBF | 14310.48 | (492.03) | 14132.79 | (498.45) | 13779.02 | (570.82) |
| Target NS | 0.77 | (0.12) | 0.97 | (0.14) | 1.02 | (0.17) |
| Target LF | 0.69 | (0.05) | 0.81 | (0.07) | 0.74 | (0.06) |
| Nonword Prime | ||||||
| Length | 7.61 | (0.10) | 7.46 | (0.11) | 7.40 | (0.11) |
| Stem Length | 4.49 | (0.09) | 4.34 | (0.09) | 4.26 | (0.07) |
| Stem CBF | 6836.33 | (383.92) | 7115.82 | (356.47) | 7481.60 | (373.00) |
| Stem NS | 5.62 | (0.48) | 6.53 | (0.56) | 6.94 | (0.52) |
| Stem LF | 1.40 | (0.07) | 1.37 | (0.08) | 1.31 | (0.08) |
| Suffix Length | 2.59 | (0.07) | 2.49 | (0.08) | 2.62 | (0.09) |
As the task was semantic categorization with animal names as the target category, we added 124 filler items consisting of 62 animal names each in either the target or the prime position. When the animal names were in the target position, the primes were nonwords similar to those of the experimental items in that they consisted of part of the animal name acting as a stem followed by an affix (e.g., cougance-COUGAR). These primes could be either related or unrelated to the animal name. Animal names were also included in the prime position in order to monitor prime visibility. In these cases, targets resembled those of the experimental pairs in that they were either real complex words, pseudocomplex words or simplex words. Participants detected 89% of the animal targets but only 3% of the animal primes suggesting that the primes were effectively masked.
Procedure
Participants were seated in a comfortable chair is a darkened room at a distance of 59 inches from the computer monitor. Each testing session began with a short practice block, followed by the experimental block. Participants were told that they would see a list of words appear on the computer monitor and were instructed to monitor the list of words for animal name and press a button on a game controller when an animal name appeared. There were 31 such words in the prime position and 31 in the target position. Participants were told to read all other words passively (i.e., critical stimuli did not require an overt response). Visual stimuli were presented on a 19-in. monitor, with a diagonal viewable screen size of 18 inches, and a width of approximately 14.5 inches, with a screen resolution of 800 by 600 pixels set to a refresh rate of 100 Hz (which allows 10-ms resolution of stimulus control). Stimuli were displayed at high contrast as white letters (Verdana font) on a black background. Each letter was 40 pixels tall by 20 pixels wide. Targets which ranged from 5 to 11 characters measured 1.8 to 3.9 inches and subtended a visual angle of 1.7 to 3.8 degrees. Primes were presented in lower case letters for 50 ms preceded by a 500 ms random consonant mask. The mask shared no letters in common with the target or with the prime. The target was presented immediately after the prime in upper case letters for 300 ms followed by a 900 ms ISI.
Recording Procedure
The electroencephalogram (EEG) was recorded from 29 active tin electrodes held in place on the scalp by an elastic cap (Electrode-Cap International). In addition to the 29 scalp sites, additional electrodes were attached to below the left eye (to monitor for vertical eye movement/blinks), to the right of the right eye (to monitor for horizontal eye movements), over the left mastoid bone (reference) and over the right mastoid bone (recorded actively to monitor for differential mastoid activity). All EEG electrode impedances were maintained below 5 kΩ (impedance for eye electrodes was less than 10 kΩ). The EEG was amplified by an SA Bioamplifier with a bandpass of 0.01 and 40 Hz and the EEG was continuously sampled at a rate of 200 Hz throughout the experiment.
Data Analysis
ERPs time locked to the onset of target words in each category were formed offline from trials free of excessive artifact or response error. Trials characterized by EOG artifact in excess of 70 mV were rejected, resulting in 7.9% of trials being discarded. We calculated the mean voltage in each of four time-windows centered at 175, 250, and 425 ms (i.e. 150–200 ms, 200–300 ms, and 350–500 ms after target onset), relative to a 100 ms baseline (50 ms pre-stimulus and 50 ms post-stimulus). These time epochs were chosen to minimize temporal overlap between measured components and because they correspond roughly to peaks in the waveforms elicited by the stimuli that were identified by visual inspection, as well as to the time windows associated with a series of electrophysiological effects (N250, N400) that have been hypothesized to correspond to the component processes involved in word recognition (Grainger & Holcomb, 2009). The early 150–200 ms window corresponds to a pre-N250 period that might capture an earlier onset of N250 priming effects in certain conditions. The 50 ms pre and 50 post stimulus baseline was chosen in order to minimize the effect of waveforms elicited by the prime stimulus given the close temporal proximity of the prime and target in this paradigm.
We analysed the mean amplitude scores by selecting 15 representative sites distributed across the scalp (FP1, FPz, FP2, F3, Fz F4, C3, Cz, C4, P3, Pz, P4, O1, Oz, O2). This method of analyzing ERP data has been widely used in previous studies (Chauncey, Holcomb, & Grainger, 2008; Midgley, Holcomb, & Grainger, 2009) and provides a thorough analysis of the entire head while reducing the number of sites included in the analysis and hence the complexity of the interpretation. Our model included four within-subjects factors Relatedness, Target Type, Anteriority and Laterality. The Relatedness factor contrasted mean ERP amplitude scores for the related and unrelated word pairs, while the Target Type factor contrasted mean ERP amplitude scores for complex, pseudocomplex and simplex targets. To analyze the scalp distribution of the ERP effects, we included two factors, Anteriority and Laterality. The Anteriority factor represented the anterior-posterior distribution of effects and included five levels contrasting electrode locations from the back to the front of the head, while the Laterality factor represented the left-right distribution and included three levels contrasting electrode locations at left hemisphere, midline and right hemisphere locations.
List was included as a dummy variable. As there were no significant main effects of List (all ps > 0.1) we only report results concerning the main effects of the experimentally manipulated factors Relatedness and Target Type, and the interaction of these factors with the topographic factors Anteriority and Hemisphere. The alpha level was set at .05. The Greenhouse-Geisser correction was applied when evaluating effects with more than one degree of freedom in the numerator. In these cases, we report the uncorrected degrees of freedom and the corrected p-value. For interactions involving the experimental variables Relatedness and Target Type, we conducted follow-up simple effects tests. As there is no consensual standard for the level of familywise error to use when testing simple effects, we chose to adjust the alpha level by dividing by the number of tests performed (Bonferroni correction).
Highlights.
Evidence for early morphological decomposition of complex and pseudocomplex words.
Decomposition operates on any string (word or nonword) composed of a stem plus affix.
Decomposition only affects the subsequent processing of truly complex words.
A model of morpho-orthographic decomposition accounts for the data.
ERPs provide a sensitive means to track the effects of morpho-orthographic decomposition.
Acknowledgments
This research was supported by NIH Grant HD25889 to P. J. Holcomb., by ERC grant #230313 to J. Grainger
Footnotes
Although we generally avoid direct comparisons of ERPs generated by different items, since this runs the risk of confounding condition and item effects which can both influence early ERPs, here this comparison is motivated by the hypotheses under test and it also provides a comparison point with the Lavric et al. (2012) study. To this end we very carefully matched the different target types on a number of variables (see Table 1).
For simplex targets this was defined as the number of characters in the target that were not also contained in the embedded stem.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Aronoff M, Fudeman K. What is Morphology. 2. Wiley-Blackwell; 2010. [Google Scholar]
- Baayen RH, Piepenbrock R, Gulikers L. The CELEX lexical database (CD-ROM) Philadelphia: PA: Linguistic Data Consortium, University of Pennsylvania; 1995. [Google Scholar]
- Chauncey K, Holcomb P, Grainger J. Effects of stimulus font and size on masked repetition priming: An event-related potentials (ERP) investigation. Language and Cognitive Processes. 2008;23(1):183–200. doi: 10.1080/01690960701579839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crepaldi D, Rastle K, Coltheart M, Nickels L. ‘Fell’ primes ‘fall’, but does ‘bell’ prime ‘ball’? Masked priming with irregularly inflected primes. Journal of Memory and Language. 2010;63(1):83–99. [Google Scholar]
- Diependaele K, Morris J, Serota RM, Bertrand D, Grainger J. Breaking boundaries: Letter transpositions and morphological processing. Language and Cognitive Processes. 2013 in press. [Google Scholar]
- Diependaele K, Sandra D, Grainger J. Masked cross-modal morphological priming: Unravelling morpho-orthographic and morpho-semantic influences in early word recognition. Language and Cognitive Processes. 2005;20(1):75–114. [Google Scholar]
- Diependaele K, Sandra D, Grainger J. Semantic transparency and masked morphological priming: The case of prefixed words. Memory & Cognition. 2009;37(6):895–908. doi: 10.3758/MC.37.6.895. [DOI] [PubMed] [Google Scholar]
- Drews E, Zwitserlood P. Morphological and orthographic similarity in visual word recognition. Journal of Experimental Psychology: Human Perception & Performance. 1995;21(5):1098–1116. doi: 10.1037//0096-1523.21.5.1098. [DOI] [PubMed] [Google Scholar]
- Feldman LB, O’Connor PA, Del Prado Martín FM. Early morphological processing is morpho-semantic and not simply morpho-orthographic: A violation of form-then-meaning accounts of word recognition. Psychonomic Bulletin & Review. 2009;16(4):684–691. doi: 10.3758/PBR.16.4.684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Forster KI, Davis C. Repetition priming and frequency attenuation in lexical access. Journal of Experimental Psychology: Learning, Memory, and Cognition. 1984;10(4):680–698. [Google Scholar]
- Giraudo H, Grainger J. Priming complex words: Evidence for supralexical representation of morphology. Psychonomic Bulletin & Review. 2001;8(1):127–131. doi: 10.3758/bf03196148. [DOI] [PubMed] [Google Scholar]
- Gold BT, Rastle K. Neural correlates of morphological decomposition during visual word recognition. Journal of Cognitive Neuroscience. 2007;19(12):1983–1993. doi: 10.1162/jocn.2007.19.12.1983. [DOI] [PubMed] [Google Scholar]
- Grainger J, Holcomb PJ. Watching the word go by: On the time-course of component processes in visual word recognition. Language and Linguistics Compass. 2009;3(1):128–156. doi: 10.1111/j.1749-818X.2008.00121.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grainger J, Colé P, Segui J. Masked morphological priming in visual word recognition. Journal of Memory & Language. 1991;30(3):370–384. [Google Scholar]
- Grainger J, Kiyonaga K, Holcomb PJ. The time course of orthographic and phonological code activation. Psychological Science. 2006;17(12):1021–1026. doi: 10.1111/j.1467-9280.2006.01821.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grainger J, Lopez D, Eddy M, Dufau S, Holcomb PJ. How word frequency modulates masked repetition priming: An ERP investigation. Psychophysiology. 2012;49:604–616. doi: 10.1111/j.1469-8986.2011.01337.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holcomb &, Grainger J. On the time-course of visual word recognition: An ERP investigation using masked repetition priming. Journal of Cognitive Neuroscience. 2006;18(10):1631–1643. doi: 10.1162/jocn.2006.18.10.1631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Landauer TK, Dumais ST. A solution to Plato’s problem: The latent semantic analysis theory of acquisition, induction, and representation of knowledge. Psychological Review. 1997;104(2):211–240. [Google Scholar]
- Lavric A, Clapp A, Rastle K. ERP Evidence of morphological analysis from orthography: A masked priming study. Journal of Cognitive Neuroscience. 2007;19(5):866–877. doi: 10.1162/jocn.2007.19.5.866. [DOI] [PubMed] [Google Scholar]
- Lavric A, Elchlepp H, Rastle K. Tracking hierarchical processing in morphological decomposition with brain potentials. Journal of Experimental Psychology: Human Perception and Performance. 2012;38(4):811–816. doi: 10.1037/a0028960. [DOI] [PubMed] [Google Scholar]
- Lewis G, Solomyak O, Marantz A. The neural basis of obligatory decomposition of suffixed words. Brain and Language. 2011;118(3):118–127. doi: 10.1016/j.bandl.2011.04.004. [DOI] [PubMed] [Google Scholar]
- Longtin CM, Meunier F. Morphological decomposition in early visual word processing. Journal of Memory and Language. 2005;53(1):26–41. [Google Scholar]
- Longtin CM, Segui J, Halle PA. Morphological priming without morphological relationship. Language & Cognitive Processes. 2003;18(3):313–334. [Google Scholar]
- Marslen-Wilson W, Zhou X. Abstractness, allomorphy, and lexical architecture. Language & Cognitive Processes. 1999;14(4):321–352. [Google Scholar]
- Marslen-Wilson W, Tyler LK, Waksler R, Older L. Morphology and meaning in the English mental lexicon. Psychological Review. 1994;101(1):3–33. [Google Scholar]
- Massol S, Grainger J, Dufau S, Holcomb PJ. Masked priming from orthographic neighbors: An ERP investigation. Journal of Experimental Psychology: Human Perception and Performance. 2010;36:162–174. doi: 10.1037/a0017614. [DOI] [PubMed] [Google Scholar]
- McCormick SF, Rastle K, Davis MH. Is there a ‘fete’ in ‘fetish’? Effects of orthographic opacity on morpho-orthographic segmentation in visual word recognition. Journal of Memory and Language. 2008;58(2):307–326. [Google Scholar]
- Midgley KJ, Holcomb PJ, Grainger J. Language effects in second language learners and proficient bilinguals investigated with event-related potentials. Journal of Neurolinguistics. 2009;22(3):281–300. doi: 10.1016/j.jneuroling.2008.08.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morris J, Frank T, Grainger J, Holcom PJ. Semantic transparency and masked morphological priming: An ERP investigation. Psychophysiology. 2007;44(4):506–521. doi: 10.1111/j.1469-8986.2007.00538.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morris J, Grainger J, Holcomb PJ. An electrophysiological investigation of early effects of masked morphological priming. Language and Cognitive Processes. 2008;23(7–8):1021–1056. doi: 10.1080/01690960802299386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morris J, Porter JH, Grainger J, Holcomb PJ. Effects of lexical status and morphological complexity in masked priming: An ERP study. Language and Cognitive Processes. 2011;26(4–6):558–599. doi: 10.1080/01690965.2010.495482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rastle K, Davis MH. Morphological decomposition based on the analysis of orthography. Language and Cognitive Processes. 2008;23(7–8):942–971. [Google Scholar]
- Rastle K, Davis MH, New B. The broth in my brother’s brothel: Morpho-orthographic segmentation in visual word recognition. Psychonomic Bulletin & Review. 2004;11(6):1090–1098. doi: 10.3758/bf03196742. [DOI] [PubMed] [Google Scholar]
- Rastle K, Davis MH, Marslen-Wilson WD, Tyler LK. Morphological and semantic effects in visual word recognition: A time-course study. Language & Cognitive Processes. 2000;15(4–5):507–537. [Google Scholar]
- Solomyak O, Marantz A. Evidence for early morphological decomposition in visual word recognition. Journal of Cognitive Neuroscience. 2010;22(9):2042–2057. doi: 10.1162/jocn.2009.21296. [DOI] [PubMed] [Google Scholar]