

Abstract
Booklice in the genus Liposcelis are pests of stored grain products. They pose a considerable economic threat to global food security and safety. To date, the complete mitochondrial genome has only been determined for a single booklouse species Liposcelis bostrychophila. Unlike most bilateral animals, which have their 37 mt genes on one circular chromosome, ≈15 kb in size, the mt genome of L. bostrychophila has two circular chromosomes, 8 and 8.5 kb in size. Here, we report the mt genome of another booklouse, Liposcelis decolor. The mt genome of L. decolor has the typical mt chromosome of bilateral animals, 14,405 bp long with 37 genes (13 PCGs, 22 tRNAs and 2 rRNAs). However, the arrangement of these genes in L. decolor differs substantially from that observed in L. bostrychophila and other insects. With the exception of atp8-atp6, L. decolor differs from L. bostrychophila in the arrangement of all of the other 35 genes. The variation in the mt genome organization and mt gene arrangement between the two Liposcelis species is unprecedented for closely related animals in the same genus. Furthermore, our results indicate that the two-chromosome mt genome organization observed in L. bostrychophila likely evolved recently after L. bostrychophila and L. decolor split from their most recent common ancestor.
Introduction
Insect mitochondrial (mt) genomes usually consist of a single circular chromosome (13–20 kb) containing 13 protein-coding genes (PCGs), 22 transfer RNA genes (tRNAs) and two ribosomal RNA genes (rRNAs), which is typical of bilateral animals [1]. The mt genome usually contains one large non-coding element called the A+T-rich or control region, which contains the sites for genome replication and the initiation of gene transcription [1]. Due to several unique features, including conserved gene content, maternal inheritance, and rapid nucleotide evolution, mt genome sequences have been used to facilitate the understanding of animal evolution [2]. Currently, more than 300 insect mt genomes have been sequenced [3]. Various types of gene rearrangements have been observed in insect mt genomes. Rearrangements of tRNA genes are the most common whereas rearrangements of protein-coding and rRNA genes are less common [4]–[6]. Studies in recent years also revealed variations of mt genome organization in bilateral animals [1]. For instance, mt genomes that consist of multiple chromosomes have been reported in parasitic lice [7]–[9], booklice [3], rotifera [10] and nematodes [11]–[13]. Variations in mt genome organization may provide a novel perspective for understanding animal evolution [14]–[16], in addition to genome sequences [3], [17], RNA secondary structures [7], [18]–[20], and gene rearrangements [5], [18], [21], [22].
Several types of atypical mt genome organization have been reported in psocodean insects (superorder Psocodea) in recent years. Psocodea contains two orders of insects: Psocoptera (booklice and barklice) and Phthiraptera (chewing and sucking lice). The mt genomes of human lice, Pediculus humanus, P. capitis and Pthirus pubis, consist of 14 to 20 mini-chromosomes, each one is 1.8 to 4 kb in size and contains one to five genes [7], [9]. The chewing louse, Coloceras sp., has a typical mt chromosome with 37 genes and a circular mt DNA molecule that is approximately half the size of the typical mt chromosome [8]. The booklouse, Liposcelis bostrychophila, has a bipartite mt genome with two chromosomes: one chromosome is ≈8 kb in size and has 16 genes and the other is ≈8.5 kb in size and has 22 genes [3]. Extensive gene rearrangement has been found in the mt genomes of most of the 12 Psocodea species that have been completely or nearly completely sequenced to date, including the booklouse, L. bostrychophila [3].
During the last two decades, the booklice of the genus Liposcelis have emerged as serious pests of stored commodities worldwide [23], [24]. The genus Liposcelis has 126 known species worldwide and includes four groups (A, B, C, and D)[25], [26]. The booklouse. L. decolor, investigated in the current study, belongs to group C, while L. bostrychophila belongs to group D. Many previous studies have indicated that there is great variation among the four Liposcelis groups with respect to morphology, physiology, biochemistry and molecular biology [25], [27], [28]. In particular, analyses of ITS (internal transcribed spacers) sequences indicated that Liposcelis species of groups C and D have the highest nucleotide divergence among the four groups [25].
To understand whether the bipartite mt genome organization observed in L. bostrychophila occurred in other booklice of the genus Liposcelis, we sequenced the mt genome of L. decolor. We found that, unlike L. bostrychophila, L. decolor has the typical mt chromosome of bilateral animals. However, the arrangement of mt genes in L. decolor differs substantially from that in L. bostrychophila and other known insects. Our results showed, for the first time, a high level of variation in both mt genome organization and mt gene arrangement between closely related animal species in the same genus.
Materials and Methods
Ethics statement
No specific permits were required for the insects collected in this study. The sampling locations were not privately owned or protected in any way and the collection did not involve endangered or protected species.
Sample collection, DNA extraction, PCR and sequencing
L. decolor individuals were collected at grain storage facilities in Binzhou, Shandong Province, China in 2010, and identified to species according to their morphological characteristics [28]–[30]. Subsequently, the ITS sequence [25] and sequences of partial rrnL and cox1 genes [31] were used to confirm the species identification. The ITS sequence obtained has been deposited in GenBank under accession number KF874610. An L. decolor colony was maintained in the lab on a diet of whole wheat flour, skim milk, and yeast powder (10∶1∶1) in an incubator at 27 ± 0.5°C, 75–80% relative humidity and a scotoperiod of 24 hours. Voucher specimens (#Ps-01-01-03) were deposited at the Insect Collection, Southwest University, Chongqing, China. Total genomic DNA was extracted using a Tissue/Cell gDNA Mini Kit (Watson Biotechnologies, Shanghai, China) and stored at −20°C. Parts of cox1, cox3, cob, rrnS, rrnL and nad5 genes were amplified by PCR with conserved insect primers (Table S1) [32]. Species-specific primers were then designed for long PCR (Table S1).
Each long PCR reaction was performed in a 25 μL volume, containing 1 μL each of forward primer (10 μM) and reverse primer (10 μM), 4 μL of dNTPs (each 2.5 mM), 1 μL of template DNA (∼300 ng/μL), 2.5 μL MgCl2 (25 mM), 2.5 μL of 10× LA PCR reaction buffer, 12.75 μL ddH2O and 0.25 μL LA Taq DNA polymerase (5 U/μL, Takara). All reactions were carried out using C1000™ thermal cyclers (Bio-RAD, Hercules, CA, USA) with the follow conditions: initial 2 min denaturation at 94°C, 37 cycles of 94°C for 20 sec, 58°C for 50 sec, 68°C for 5–10 min (depending on target size, 1 min/kb), followed by a final extension at 68°C for 15 min. Gel-purified amplification products < 6 kb in size were ligated into pGEM-T Easy vectors (Promega, Madison, WI, USA), and introduced into Escherichia coli (Trans5α, TransGen Biotech, Beijing, China), following by ampicillin selection, then sequenced with M13 primers. Longer PCR products (> 6 kb) were directly sequenced with both forward and reverse PCR primers and internal primers by primer walking. All amplification products were sequenced with an ABI 3730 automated DNA sequencer (Applied Biosystems, Foster city, CA, USA) at the Beijing Genomics Institute (BGI) in Beijing, China.
Sequence assembly, annotation and analysis
SeqMan (DNAStar) was used to assemble the two overlapping nucleotide sequences, which were further confirmed by manually inspection. The protein-coding and rRNA genes were identified using the program ORF Finder (http://www.ncbi.nlm.nih.gov/gorf/gorf.html) and BLAST searches against the GenBank database, respectively. Subsequently, all of these genes were further confirmed by alignment with homologous genes from those of other louse and booklouse species. The transfer RNA genes were identified by their cloverleaf secondary structure using ARWEN [33] with default parameters and tRNAscan-SE 1.21 [34] with Search Mode = “EufindtRNA-Cove”, Genetic Code = “Invertebrate Mito” and Cove score cutoff = 0.1. The stem-loop secondary structure of the putative control regions was folded using the Mfold Server [35] under the RNA folding option with default parameters. The base composition and codon usage were analyzed with BioEdit (http://bioedit.software.informer.com/) and DAMBE 5.3.9 [36]. Sequences of mt genomes of other lice were retrieved from GenBank and MitoZoa (Table S2) [37].
Phylogenetic analyses
Phylogenetic analyses were conducted with the 11 Psocodea mt genome sequences currently available in GenBank including the new booklouse sequence obtained in this study. The mt genome sequence of the fruit fly, Drosophila melanogaster, served as an outgroup. Sequences of atp8, nad4L, and tRNA genes were too short and too variable to be correctly aligned among the psocodean species; these genes were thus excluded from the phylogenetic analyses. The nad4 was also excluded as this gene has not been identified in the human pubic louse, P. pubis [7]. The amino acid sequences from each protein-coding gene and the nucleotide sequence of each rRNA gene were aligned with MAFFT v7 [38]. The nucleotide sequences of each protein-coding gene were aligned based on the corresponding amino acid alignments using PAL2NAL [39] to ensure the correct reading frame; the poorly aligned sites were removed with GUIDANCE[40] using the default setting. Then, positions with gap in more than half of the species were removed. Substitution saturations of the nucleotide sequences were examined using DAMBE 5.3.9 following Xia et al. [41]. Whole PCG sequences were chosen to enter the next step if Iss (index of substitution saturation) is significantly lower than Iss.c (critical value for symmetrical tree topology) (P < 0.05). All of the protein-coding and rRNA genes, except nad3 and nad6, passed this test. Consequently, the third codon positions of nad3 and nad6 were excluded from phylogenetic analyses. The best fit models for the alignment of nucleotide sequence and amino acid sequence were determined using the Akaike Information Criterion in jModelTest 0.1.1 [42] and ProtTest 3 [43], respectively. Specifically, the GTR+I+G model and MtREV+I+G model were chosen for the nucleotide sequence dataset and the amino acid sequence dataset, respectively. Phylogenetic trees were estimated via Bayesian inference (BI) method using MrBayes v3.12 [44]. Four independent Markov chains were simultaneously run for 2,000,000 generations with a heating scheme (temp = 0.2). Trees were sampled every 100 generations (sample-freq = 100) and the first 25% of the generations were discarded as burn-in and the remaining samples were used to compute the consensus tree. Stationarity was considered to be reached when the average standard deviation of split frequencies was below 0.01 [45].
Results and Discussion
Mitochondrial genome of Liposcelis decolor
The mt genome of L. decolor has one typical circular chromosome, unlike the booklouse, L. bostrychophila (Figure 1). The size and the circular organization of the mt chromosome of L. decolor was confirmed by two overlapping PCR amplicons, 9.1 kb (d1-d2 from cox3 to rrnS) and 5.5 kb in size (d3-d4 from rrnS to cox3) respectively (Figure 1, Figure 2 and Table S1). The two amplicons overlapped by 33 bp in cox3 and 92 bp in rrnS.
Figure 1. Mitochondrial genome of Liposcelis decolor.

Transcriptional orientation is indicated with arrows. Protein-coding genes, ribosomal RNA genes and transfer RNA genes are shown in orange, blue and green respectively. tRNA genes for the two serine and two leucine tRNAs: S1 = AGN, S2 = UCN, L1 = CUN, and L2 = UUR. The non-coding regions larger than 60 bp are indicated in black. CR = putative control region. Arrows and purple curves indicate primers and PCR fragment, respectively. See Table S1 for sequence of PCR primers.
Figure 2. PCR amplification and verification of mitochondrial DNA of Liposcelis decolor.

Long-PCR amplification of two fragments to verify the single circular mt genome in L. decolor. Lane C1, negative control without the forward primer d1 or d3; lane C2, negative control without the reverse primer d2 or d4; lane C3, negative control without the DNA template. Lane M: 1 kb marker (Biomed). “d1-d2”, the product of PCR with primers d1 and d2, etc. Primer details are given in Table S1.
Sequencing and assembly of these two PCR amplicons revealed that the mt genome of L. decolor is 14,405 bp in length and encodes 37 genes that are typically found in metazoan mt genomes (Figure 1 and Table S3) (GenBank accession number: JX870621). All of the protein-coding genes (PCGs) initiate translation at an ATN codon, except for the TTG codon used in cox1. TAA and TAG serve as stop codons for all of the PCGs (Table S4). Eleven of the 13 PCGs had average length for Psocodean species, but nad4 is shorter and the nad4L is longer than in other Psocodean species (Figure S1). All 22 tRNA coding genes usually found in the mt genomes of metazoans are present in L. decolor (Figure 3); all have the conventional cloverleaf shaped secondary structure except trnS1, which lacks the D-arm, as in other insects. There are three non-coding regions longer than 60 bp in the mt genome of L. decolor. The longest non-coding region (118 bp) lies between cob and trnY, has an A+T content of 82.20%; three stem-loop secondary structures can be found in this region (Figure 4A). An 80 bp non-coding region lies between nad2 and nad4 with an 88.75% A+T content but has no stem-loop secondary structure. A 69 bp non-coding region is between trnG and trnL1 with a 92.75% A+T content and has two stem-loops (Figure 4B). The 118-bp region is considered to be most likely the putative control region (Figure 1) due to it is longer sequence in the mt genome and the presence of typical loop secondary structure.
Figure 3. Putative secondary structures of the 22 tRNA genes identified in the mitochondrial genome of Liposcelis decolor.

Bars indicate Watson-Crick base pairings, and dots between G and U pairs mark canonical base pairings in RNA.
Figure 4. Stem-loop secondary structures of two non-coding regions of Liposcelis decolor.
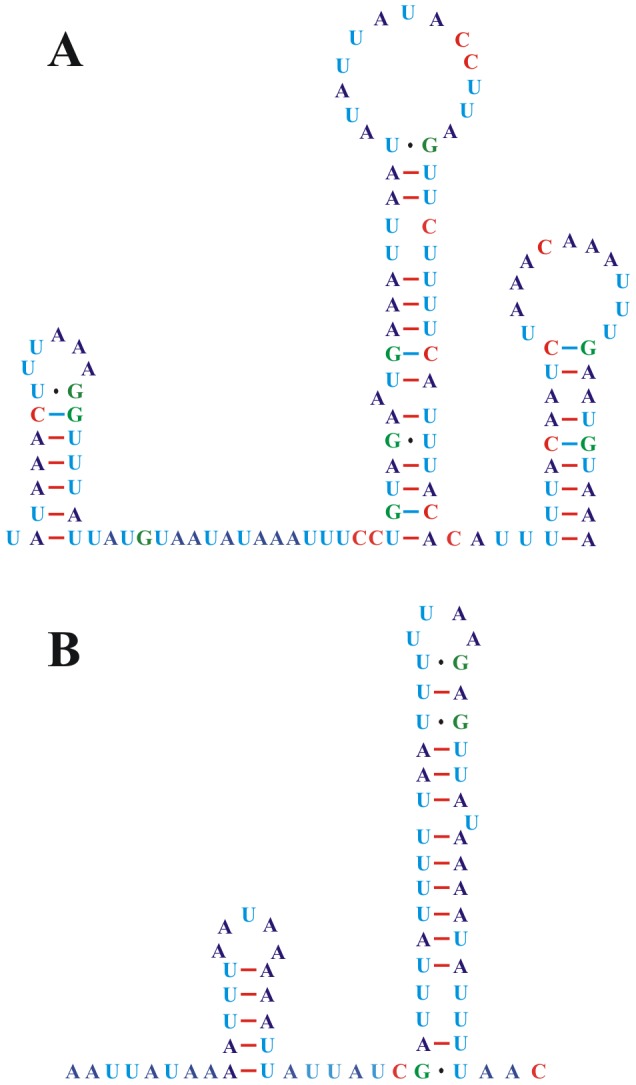
A, Stem-loop secondary structure of the 118 bp non-coding sequence; B, Stem-loop secondary structure of the 69 bp non-coding sequence.
The A+T content of the L. decolor mt genome is 75.23%. This is typical of psocodean insects but is higher than that of L. bostrychophila, 68.63% (Figure S2 and Table S5). The higher A+T content of L. decolor is present in all regions, both genes and non-coding regions (Table S5). The difference of A+T content between the two booklice is reflected further in the codon usage: the relative synonymous codon usages (RSCU) of the two booklice showed that L. decolor used more NNA and NNT codon than L. bostrychophila (Figure 5). The nucleotide composition of mt genome is usually conserved within a genus; however, it varies between L. decolor and L. bostrychophila. This variation may be related to mt genome fragmentation, because all of the psocodean fragmented mt genomes have a lower A+T content (Figure S2).
Figure 5. Relative synonymous codon usage (RSCU) for protein coding genes of Liposcelis decolor and L. bostrychophila.

Abbreviations of tRNA genes are according to the single letter according to the IPUC-IUB one-letter amino acid codes.
Mitochondrial gene rearrangement in L. decolor
The mt gene arrangement in L. decolor differs substantially from that of the hypothetical ancestor of insects and from that of the booklouse, L. bostrychophila (Figure 6). With the only exception of atp8-atp6, there is no gene boundary or gene block shared between L. decolor and L. bostrychophila, even though these two booklice belong to the same genus. Atp8-atp6, is a highly conserved ancestral gene boundary for animals and is assumed to be constrained by the function of a bicistronic atp8-atp6 transcript [46]. The mt gene arrangement is usually conserved within the same genus. For example, the two Pediculus species of human lice, both have extensively fragmented mt genomes but have the same mt gene arrangement [7]. Furthermore, Coloceras sp. and Campanulotes bidentatus, also have the same gene arrangement, except for a difference in the location of their trnQ. The extent of the variation in the mt gene arrangement between the two species of Liposcelis booklice is unprecedented for animals within the same genus, indicating that gene rearrangement occurred frequently after these two booklice diverged from each other. The genus Liposcelis is divided into four groups (A, B, C, D) phylogenetically [25], [26]; L. decolor and L. bostrychophila are in different groups, C and D respectively, although they often co-occur in a wide range of stored products in the same ecosystems. Previous studies have revealed that substantial variation exists among the Liposcelis groups at both morphological and molecular levels [25], . Whether or not the multipartite mt genome observed in L. bostrychophila occurred only in species of group D remains to be investigated.
Figure 6. Arrangement of mitochondrial genes in Liposcelis and the hypothetical ancestor of the arthropods.

Circular genomes have been arbitrarily linearized for ease of comparison. Gene names are the standard abbreviations used in the present study. tRNA genes are designated by the single letter according to the IPUC-IUB one-letter amino acid codes. Genes which are underlined are encoded on the opposite strand to the majority of genes in that particular genome. Black, gray, and white boxes represent putative control regions, pseudogenes, and transfer RNA genes, respectively. The boxes in 15 colors represent 13 protein coding genes and 2 ribosomal RNA genes. Shared gene-boundaries are labeled with square “brackets” above each genome.
Phylogenetic relationship of L. decolor to other species in the Psocodea
Bayesian inference was used to determine phylogenetic relationships among 11 species of Psocodea from the orders Psocoptera and Phthiraptera, with nucleotide and deducted amino acid sequences of mt genomes (Figure 7 and Figure S3). Although L. decolor and L. bostrychophila differ in both mt genome organization and mt gene arrangement, these two booklice are more closely related to each other than to other species in the Psocodea. The two Liposcelis species formed a sister clade to the parasitic lice (order Phthiraptera). This was also indicated by previous studies [3], [30], [47]. The close relationships between the two booklice, and between the booklice and the parasitic lice are strongly supported by our analyses. Our results indicate that the bipartite mt genome organization observed in L. bostrychophila likely evolved recently L. decolor and L. bostrychophila split from their most recent common ancestor. Furthermore, from the phylogenies of the Psocodea constructed in the present study and a number of previous studies [7], [9], it can be inferred that the bipartite mt genome organization in L. bostrychophila evolved independently from the much more fragmented mt genomes observed in the blood-sucking lice (suborder Anoplura).
Figure 7. Phylogeny of the Psocodea inferred with mitochondrial genome sequences.

Numbers above the left branches show Bayesian posterior probability for the phylogenies from nucleotide sequences, the right from amino acid sequences. Only support above 50% is shown. The insects belong to Phthiraptera, Psocoptera, and Diptera are shown in gray, yellow and pink background respectively.
Supporting Information
Size of mitochondrial protein-coding genes and rRNA genes of the Psocodea. Lower horizontal bar, non-outlier smallest observation; lower edge of rectangle, 25 percentile; the central horizontal bar, median; upper edge of rectangle, 75 percentile; upper horizontal bar, non-outlier largest observation; small circle, outlier. Species are abbreviated as following: Ld, Liposcelis decolor; Lb, Liposcelis bostrychophila; Ls, Lepidopsocidae sp. RS-2001; Bm, Bothriometopus macrocnemis; Ac, Anaticola crassicornis.
(TIF)
A+T contents of the mitochondrial genomes of the Psocodea. The checked bars designate the fragmented genomes. The mt genomes sequenced incompletely are marked by asterisks. Species are abbreviated as following: Ld, Liposcelis decolor; Lb, Liposcelis bostrychophila; Ls, Lepidopsocidae sp. RS-2001; Bm, Bothriometopus macrocnemis; Cb, Campanulotes bidentatus; Cs: Coloceras sp. SLC-2011; Hm, Heterodoxus macropus; Ib, Ibidoecus bisignatus; Pc, Pediculus capitis; Ph, Pediculus humanus; Pp, Pthirus pubis; Ac, Anaticola crassicornis; Dm, Damalinia meyeri; Ps, Philopterus sp. SLC-2011; Qs, Quadraceps sp. SLC-2011; Bp, Brachionus plicatilis.
(TIF)
Alignments of mitochondrial gene sequences used for phylogenetic analyses. A. Nucleotide sequence alignment (gene order of this alignment: cox1-3, atp6, cob, nad1-3, nad5-6, rrnS and rrnL; sequences of nad3 and nad6 just with codon positions 1 and 2); B. Amino acid sequence alignment (gene order of this alignment: cox1-3, atp6, cob, nad1-3 and nad5-6). Species are abbreviated as following: Ld, Liposcelis decolor; Lb, Liposcelis bostrychophila; Ls, Lepidopsocidae sp. RS-2001; Bm, Bothriometopus macrocnemis; Cb, Campanulotes bidentatus; Cs: Coloceras sp. SLC-2011; Hm, Heterodoxus macropus; Ib, Ibidoecus bisignatus; Pc, Pediculus capitis; Ph, Pediculus humanus; Pp, Pthirus pubis; Dm, Drosophila melanogaster.
(DOC)
PCR primers used for the amplification of the mitochondrial genome of Liposcelis decolor .
(DOC)
GenBank accession numbers of all the insects mentioned in this study.
(DOC)
Summary of the mitochondrial genome of Liposcelis decolor . aGenes located in the different strand from that of cox1 are underlined. binc = intergenic nucleotides, indicating gap nucleotides (positive value) and overlapping nucleotides (negative value) of two adjacent genes. cAT-skew = (A−T)/(A+T), GC-skew = (G−C)/(G+C). dCR = control region (putative).
(DOC)
Start and stop codons of mitochondrial protein-coding genes of the Psocodea. Species are abbreviated as follows: Ld, Liposcelis decolor; Lb, Liposcelis bostrychophila; Ls, Lepidopsocidae sp. RS-2001; Bm, Bothriometopus macrocnemis; Cb, Campanulotes bidentatus; Cs, Coloceras sp. SLC-2011; Hm, Heterodoxus macropus; Ib, Ibidoecus bisignatus; Pc, Pediculus capitis; Ph, Pediculus humanus; Pp, Pthirus pubis; Ac, Anaticola crassicornis; Dm, Damalinia meyeri; Ps, Philopterus sp. SLC-2011.
(DOC)
Nucleotide compositions of the mitochondrial genomes of Liposcelis decolor and L. bostrychophila .
(DOC)
Funding Statement
This work was supported by the National Natural Sciences Foundation (31000860), the Program for Innovative Research Team in University (IRT0976), the Specialized Research Fund for the Doctoral Program of Higher Education(20100182120022), and the Fundamental Research Funds for the Central Universities (XDJK2013C051, XDJK2013A005). The funders had no role in study design, data collection and analysis, the decision to publish, or the preparation of the manuscript.
References
- 1. Boore JL (1999) Animal mitochondrial genomes. Nucleic Acids Research 27: 1767–1780. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Sheffield NC, Song H, Cameron L, Whiting MF (2008) A comparative analysis of mitochondrial genomes in Coleoptera (Arthropoda: Insecta) and genome descriptions of six new beetles. Molecular Biology and Evolution 25: 2499–2509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Wei DD, Shao R, Yuan ML, Dou W, Barker SC, et al. (2012) The multipartite mitochondrial genome of Liposcelis bostrychophila: insights into the evolution of mitochondrial genomes in bilateral animals. PLoS One 7: e33973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Satoh TP, Sato Y, Masuyama N, Miya M, Nishida M (2010) Transfer RNA gene arrangement and codon usage in vertebrate mitochondrial genomes: a new insight into gene order conservation. BMC Genomics 11: 479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Covacin C, Shao R, Cameron S, Barker SC (2006) Extraordinary number of gene rearrangements in the mitochondrial genomes of lice (Phthiraptera: Insecta). Insect Molecular Biology 15: 63–68. [DOI] [PubMed] [Google Scholar]
- 6. Shao R, Campbell NJH, Barker SC (2001) Numerous gene rearrangements in the mitochondrial genome of the wallaby louse, Heterodoxus macropus (Phthiraptera). Molecular Biology and Evolution 18: 858–865. [DOI] [PubMed] [Google Scholar]
- 7. Shao R, Zhu XQ, Barker SC, Herd K (2012) Evolution of extensively fragmented mitochondrial genomes in the lice of humans. Genome Biology and Evolution 4: 1088–1101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Cameron SL, Yoshizawa K, Mizukoshi A, Whiting MF, Johnson KP (2011) Mitochondrial genome deletions and minicircles are common in lice (Insecta: Phthiraptera). BMC Genomics 12: 394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Shao RF, Kirkness EF, Barker SC (2009) The single mitochondrial chromosome typical of animals has evolved into 18 minichromosomes in the human body louse, Pediculus humanus . Genome Research 19: 904–912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Suga K, Welch DBM, Tanaka Y, Sakakura Y, Hagiwarak A (2008) Two circular chromosomes of unequal copy number make up the mitochondrial genome of the rotifer Brachionus plicatilis . Molecular Biology and Evolution 25: 1129–1137. [DOI] [PubMed] [Google Scholar]
- 11. Gibson T, Blok VC, Phillips MS, Hong G, Kumarasinghe D, et al. (2007) The mitochondrial subgenomes of the nematode Globodera pallida are mosaics: evidence of recombination in an animal mitochondrial genome. Journal of Molecular Evolution 64: 463–471. [DOI] [PubMed] [Google Scholar]
- 12. Gibson T, Blok VC, Dowton M (2007) Sequence and characterization of six mitochondrial subgenomes from Globodera rostochiensis: multipartite structure is conserved among close nematode relatives. Journal of Molecular Evolution 65: 308–315. [DOI] [PubMed] [Google Scholar]
- 13. Watanabe KI, Bessho Y, Kawasaki M, Hori H (1999) Mitochondrial genes are found on minicircle DNA molecules in the mesozoan animal Dicyema . Journal of Molecular Biology 286: 645–650. [DOI] [PubMed] [Google Scholar]
- 14. Liu YG, Kurokawa T, Sekino M, Tanabe T, Watanabe K (2013) Complete mitochondrial DNA sequence of the ark shell Scapharca broughtonii: an ultra-large metazoan mitochondrial genome. Comparative Biochemistry and Physiology D-Genomics & Proteomics 8: 72–81. [DOI] [PubMed] [Google Scholar]
- 15. Liu GH, Wang Y, Song HQ, Li MW, Ai L, et al. (2013) Characterization of the complete mitochondrial genome of Spirocerca lupi: sequence, gene organization and phylogenetic implications. Parasites & Vectors 6: 45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Kayal E, Bentlage B, Collins AG, Kayal M, Pirro S, et al. (2012) Evolution of linear mitochondrial genomes in medusozoan cnidarians. Genome Biology and Evolution 4: 1–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Nardi F, Spinsanti G, Boore JL, Carapelli A, Dallai R, et al. (2003) Hexapod origins: monophyletic or paraphyletic? Science 299: 1887–1889. [DOI] [PubMed] [Google Scholar]
- 18. Li H, Liu H, Shi AM, Stys P, Zhou XG, et al. (2012) The complete mitochondrial genome and novel gene arrangement of the unique-headed bug Stenopirates sp. (Hemiptera: Enicocephalidae). PLoS One 7: e29419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Yuan ML, Wei DD, Wang BJ, Dou W, Wang JJ (2010) The complete mitochondrial genome of the citrus red mite Panonychus citri (Acari: Tetranychidae): high genome rearrangement and extremely truncated tRNAs. BMC Genomics 11: 597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Wei SJ, Shi M, Sharkey MJ, van Achterberg C, Chen XX (2010) Comparative mitogenomics of Braconidae (Insecta: Hymenoptera) and the phylogenetic utility of mitochondrial genomes with special reference to Holometabolous insects. BMC Genomics 11: 371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Yan D, Tang Y, Xue X, Wang M, Liu F, et al. (2012) The complete mitochondrial genome sequence of the western flower thrips Frankliniella occidentalis (Thysanoptera: Thripidae) contains triplicate putative control regions. Gene 506: 117–124. [DOI] [PubMed] [Google Scholar]
- 22. Boore JL, Brown WM (1998) Big trees from little genomes: mitochondrial gene order as a phylogenetic tool. Current Opinion in Genetics & Development 8: 668–674. [DOI] [PubMed] [Google Scholar]
- 23. Athanassiou CG, Phillips TW, Aikins MJ, Hasan MM, Throne JE (2012) Effectiveness of sulfuryl fluoride for xontrol of sifferent life stages of stored-product psocids (Psocoptera). Journal of Economic Entomology 105: 282–287. [DOI] [PubMed] [Google Scholar]
- 24. Athanassiou CG, Arthur FH, Throne JE (2010) Efficacy of methoprene for control of five species of psocids (Psocoptera) on wheat, rice, and maize. Journal of Food Protection 73: 2244–2249. [DOI] [PubMed] [Google Scholar]
- 25. Wei DD, Yuan ML, Wang ZY, Wang D, Wang BJ, et al. (2011) Sequence analysis of the ribosomal internal transcribed spacers region in psocids (Psocoptera: Liposcelididae) for phylogenetic inference and species discrimination. Journal of Economic Entomology 104: 1720–1729. [DOI] [PubMed] [Google Scholar]
- 26. Lienhard C (1990) Revision of the western palaearctic species of Liposcelis Motschulsky (Psocoptera: Liposcelididae) Zoologische Jahrbücher, Abteilung für Systematik, Ökologie und Geographie der Tiere. 117: 117–174. [Google Scholar]
- 27. Cheng WX, Dou W, Chai YX, Wang JJ (2007) Comparison of biochemical and toxicological characterizations of glutathione S-transferases and superoxide dismutase between Liposcelis bostrychophila Badonnel and L. entomophila (Enderlein) (Psocoptera: Liposcelididae). Pesticide Biochemistry and Physiology 89: 151–157. [Google Scholar]
- 28.Li FS (2002) Psocoptera Of China. Beijing: Science Press pp: 77–79.
- 29. Hu F, Zhang GN, Wang JJ (2009) Antennal sensillae of five stored-product psocids pests (Psocoptera: Liposcelididae). Micron 40: 628–634. [DOI] [PubMed] [Google Scholar]
- 30. Lyal CHC (1985) Phylogeny and classification of the Psocodea, with particular reference to the lice (Psocodea: Phthiraptera). Systematic Entomology 10: 145–165. [Google Scholar]
- 31. Yang QQ, Zhao S, Kucerova Z, Stejskal V, Opit G, et al. (2013) Validation of the 16S rDNA and COI DNA barcoding technique for rapid molecular identification of stored product psocids (Insecta: Psocodea: Liposcelididae). Journal of Economic Entomology 106: 419–425. [DOI] [PubMed] [Google Scholar]
- 32. Simon C, Frati F, Beckenbach A, Crespi B, Liu H, et al. (1994) Evolution, weighting, and phylogenetic utility of mitochondrial gene-sequences and a compilation of conserved polymerase chain-reaction primers. Annals of the Entomological Society of America 87: 651–701. [Google Scholar]
- 33. Laslett D, Canback B (2008) ARWEN: a program to detect tRNA genes in metazoan mitochondrial nucleotide sequences. Bioinformatics 24: 172–175. [DOI] [PubMed] [Google Scholar]
- 34. Lowe TM, Eddy SR (1997) tRNAscan-SE: a program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Research 25: 955–964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Zuker M (2003) Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Research 31: 3406–3415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Xia X, Xie Z (2001) DAMBE: software package for data analysis in molecular biology and evolution. Journal of Heredity 92: 371–373. [DOI] [PubMed] [Google Scholar]
- 37. D'Onorio de Meo P, D'Antonio M, Griggio F, Lupi R, Borsani M, et al. (2012) MitoZoa 2.0: a database resource and search tools for comparative and evolutionary analyses of mitochondrial genomes in Metazoa. Nucleic Acids Research 40: D1168–1172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Katoh K, Misawa K, Kuma K, Miyata T (2002) MAFFT: a novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Research 30: 3059–3066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Suyama M, Torrents D, Bork P (2006) PAL2NAL: robust conversion of protein sequence alignments into the corresponding codon alignments. Nucleic Acids Research 34: W609–W612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Penn O, Privman E, Ashkenazy H, Landan G, Graur D, et al. (2010) GUIDANCE: a web server for assessing alignment confidence scores. Nucleic Acids Research 38: W23–W28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Xia X, Lemey P (2009) Assessing substitution saturation with DAMBE Phylogenetic Handbook: A Practical Approach to Phylogenetic Analysis and Hypothesis Testing, 2nd Edition: 615–630.
- 42. Posada D (2008) jModelTest: phylogenetic model averaging. Molecular Biology and Evolution 25: 1253–1256. [DOI] [PubMed] [Google Scholar]
- 43. Darriba D, Taboada GL, Doallo R, Posada D (2011) ProtTest 3: fast selection of best-fit models of protein evolution. Bioinformatics 27: 1164–1165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Ronquist F, Huelsenbeck JP (2003) MrBayes 3: bayesian phylogenetic inference under mixed models. Bioinformatics 19: 1572–1574. [DOI] [PubMed] [Google Scholar]
- 45. Huelsenbeck JP, Ronquist F, Nielsen R, Bollback JP (2001) Bayesian inference of phylogeny and its impact on evolutionary biology. Science 294: 2310–2314. [DOI] [PubMed] [Google Scholar]
- 46. Simon M, Faye G (1984) Organization and processing of the mitochondrial oxi3/oli2 multigenic transcript in yeast. Mol Gen Genet 196: 266–274. [DOI] [PubMed] [Google Scholar]
- 47. Grimaldi D, Engel MS (2006) Fossil Liposcelididae and the lice ages (Insecta: Psocodea). Proc Biol Sci 273: 625–633. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Size of mitochondrial protein-coding genes and rRNA genes of the Psocodea. Lower horizontal bar, non-outlier smallest observation; lower edge of rectangle, 25 percentile; the central horizontal bar, median; upper edge of rectangle, 75 percentile; upper horizontal bar, non-outlier largest observation; small circle, outlier. Species are abbreviated as following: Ld, Liposcelis decolor; Lb, Liposcelis bostrychophila; Ls, Lepidopsocidae sp. RS-2001; Bm, Bothriometopus macrocnemis; Ac, Anaticola crassicornis.
(TIF)
A+T contents of the mitochondrial genomes of the Psocodea. The checked bars designate the fragmented genomes. The mt genomes sequenced incompletely are marked by asterisks. Species are abbreviated as following: Ld, Liposcelis decolor; Lb, Liposcelis bostrychophila; Ls, Lepidopsocidae sp. RS-2001; Bm, Bothriometopus macrocnemis; Cb, Campanulotes bidentatus; Cs: Coloceras sp. SLC-2011; Hm, Heterodoxus macropus; Ib, Ibidoecus bisignatus; Pc, Pediculus capitis; Ph, Pediculus humanus; Pp, Pthirus pubis; Ac, Anaticola crassicornis; Dm, Damalinia meyeri; Ps, Philopterus sp. SLC-2011; Qs, Quadraceps sp. SLC-2011; Bp, Brachionus plicatilis.
(TIF)
Alignments of mitochondrial gene sequences used for phylogenetic analyses. A. Nucleotide sequence alignment (gene order of this alignment: cox1-3, atp6, cob, nad1-3, nad5-6, rrnS and rrnL; sequences of nad3 and nad6 just with codon positions 1 and 2); B. Amino acid sequence alignment (gene order of this alignment: cox1-3, atp6, cob, nad1-3 and nad5-6). Species are abbreviated as following: Ld, Liposcelis decolor; Lb, Liposcelis bostrychophila; Ls, Lepidopsocidae sp. RS-2001; Bm, Bothriometopus macrocnemis; Cb, Campanulotes bidentatus; Cs: Coloceras sp. SLC-2011; Hm, Heterodoxus macropus; Ib, Ibidoecus bisignatus; Pc, Pediculus capitis; Ph, Pediculus humanus; Pp, Pthirus pubis; Dm, Drosophila melanogaster.
(DOC)
PCR primers used for the amplification of the mitochondrial genome of Liposcelis decolor .
(DOC)
GenBank accession numbers of all the insects mentioned in this study.
(DOC)
Summary of the mitochondrial genome of Liposcelis decolor . aGenes located in the different strand from that of cox1 are underlined. binc = intergenic nucleotides, indicating gap nucleotides (positive value) and overlapping nucleotides (negative value) of two adjacent genes. cAT-skew = (A−T)/(A+T), GC-skew = (G−C)/(G+C). dCR = control region (putative).
(DOC)
Start and stop codons of mitochondrial protein-coding genes of the Psocodea. Species are abbreviated as follows: Ld, Liposcelis decolor; Lb, Liposcelis bostrychophila; Ls, Lepidopsocidae sp. RS-2001; Bm, Bothriometopus macrocnemis; Cb, Campanulotes bidentatus; Cs, Coloceras sp. SLC-2011; Hm, Heterodoxus macropus; Ib, Ibidoecus bisignatus; Pc, Pediculus capitis; Ph, Pediculus humanus; Pp, Pthirus pubis; Ac, Anaticola crassicornis; Dm, Damalinia meyeri; Ps, Philopterus sp. SLC-2011.
(DOC)
Nucleotide compositions of the mitochondrial genomes of Liposcelis decolor and L. bostrychophila .
(DOC)