

Abstract
Flavonoids are a class of phenolic compounds commonly found in fruits, vegetables, grains, flowers, tea, and wine. They differ in their chemical structures and characteristics. Such compounds show various biological functions and have antioxidant, antimicrobial, anti-inflammatory, and antiapoptotic properties. The aim of this study was to evaluate the in vitro interactions of flavonoids with fluconazole against Candida tropicalis strains resistant to fluconazole, investigating the mechanism of synergism. Three combinations formed by the flavonoids (+)-catechin hydrated, hydrated quercetin, and (−)-epigallocatechin gallate at a fixed concentration with fluconazole were tested. Flavonoids alone had no antifungal activity within the concentration range tested, but when they were used as a cotreatment with fluconazole, there was significant synergistic activity. From this result, we set out to evaluate the possible mechanisms of cell death involved in this synergism. Isolated flavonoids did not induce morphological changes or changes in membrane integrity in the strains tested, but when they were used as a cotreatment with fluconazole, these changes were quite significant. When evaluating mitochondrial damage and the production of reactive oxygen species (ROS) only in the cotreatment, changes were observed. Flavonoids combined with fluconazole were shown to cause a significant increase in the rate of damage and the frequency of DNA damage in the tested strains. The cotreatment also induced an increase in the externalization of phosphatidylserine, an important marker of early apoptosis. It is concluded that flavonoids, when combined with fluconazole, show activity against strains of C. tropicalis resistant to fluconazole, promoting apoptosis by exposure of phosphatidylserine in the plasma membrane and morphological changes, mitochondrial depolarization, intracellular accumulation of ROS, condensation, and DNA fragmentation.
INTRODUCTION
Yeasts are the most common opportunistic agents in fungal infections of immunocompromised patients, and new fungal pathogens have emerged over the last decade (1, 2). Throughout the last 20 years, the most commonly isolated yeasts from systemic fungal infections have been species of Candida (3). The National Network of Health Security reports that Candida spp. are the third most common cause of bloodstream infections associated with intensive care units in the United States (4).
Among invasive fungal infections, Candida tropicalis has been reported in the literature to be a major non-albicans Candida species causing fungemia in patients with malignancies (5, 6). In Brazil, C. tropicalis is a common agent in hospitals (5). A survey performed in hospitals from northeast Brazil showed that, among cultures positive for Candida, C. tropicalis was the second most commonly isolated species. This scenario can be explained by the higher level of resistance of non-albicans Candida species (C. tropicalis, C. glabrata, C. parapsilosis, and C. krusei) to certain antifungal drugs in comparison to that of C. albicans (7–9).
The emergence of antimicrobial resistance and the limited efficacy of current antifungal agents have motivated the exploration for new drugs with relatively low toxicity that can reduce the chances of developing resistance (10–12).
Natural resources, such as plants, microorganisms, and invertebrates, provide sources for the discovery of potential bioactive molecules (12, 13). According to the World Health Organization (WHO), more than 80% of people use traditional medicines, mostly derived from plants and their by-products, to treat infectious diseases (12, 14).
Natural products from plants have recently attracted scientific interest for their antifungal properties (15, 16). Research in this field may lead to the development of drugs effective against pathogenic fungi (17, 18). Flavonoids (FLAV) are a group of natural substances with various phenolic structures garnering considerable scientific and therapeutic interest (Fig. 1). These compounds are widely distributed in nature and have diverse biological activities, such as antioxidant, antimicrobial, anti-inflammatory, and antiapoptotic properties, which have been observed for (+)-catechin hydrate (CATEQ), quercetin hydrate (QUERC), and (−)-epigallocatechin gallate (EPIG) (19–21).
FIG 1.

Chemical structures of the flavonoids (−)-epigallocatechin gallate (a), (+)-catechin hydrated (b), and hydrated quercetin (c).
The aim of the current study was to evaluate and compare the synergistic effects of catechin, quercetin, and epigallocatechin gallate with fluconazole in fluconazole-resistant strains of C. tropicalis via broth microdilution susceptibility tests, flow cytometry assays, and single-cell gel electrophoresis (alkaline comet assay) to investigate whether the synergism promotes yeast death through apoptosis.
MATERIALS AND METHODS
Isolates.
We used six fluconazole-resistant strains of C. tropicalis (22). The strains were inoculated onto Sabouraud dextrose agar (HiMedia, Mumbai, India) and incubated at 35°C for 24 h. They were then sown onto CHROMagar Candida (HiMedia, Mumbai, India) to assess their purity.
Antifungal susceptibility testing and drug interaction evaluation.
Broth microdilution (BMD) susceptibility testing was performed according to Clinical and Laboratory Standards Institute (CLSI) document M27-A3 (23) using RPMI broth (pH 7.0) buffered with 0.165 M morpholinepropanesulfonic acid (MOPS; Sigma Chemicals, St. Louis, MO) (23). Fluconazole (Merck Sharp & Dohme, São Paulo, Brazil) was dissolved with distilled water, and solutions of CATEQ, QUERC, and EPIG (Sigma Chemicals) were prepared in dimethyl sulfoxide (DMSO; Sigma Chemicals). All solutions were stored at −20°C until further use. Fluconazole was tested over the concentration range of 0.125 to 64 μg/ml, and the flavonoids CATEQ, QUERC, and EPIG were tested over the concentration range of 0.25 to 128 μg/ml. The 96-well culture plates were incubated at 35°C for 24 h, and the results were read visually, as recommended by the CLSI (2012) (24). The MIC was the concentration that inhibited 50% of fungal growth. The in vitro drug interactions were evaluated according to the MIC, and the strains were classified as susceptible (S), susceptible dose dependent (SDD), or resistant (R). The cutoff points for C. tropicalis susceptibility to fluconazole were as follows: S, MIC ≤ 2 μg/ml; SDD, MIC = 4 μg/ml; and R, MIC ≥ 8 μg/ml (24).
After determining the MIC50 of each drug by itself, the checkerboard technique was applied (22, 24, 25). Thus, the strains were exposed to various concentrations (0.125 to 64 μg/ml) of fluconazole in combination with flavonoids (0.25 to 128 μg/ml). The results were read visually, and the MIC50 values were determined at 24 h. Synergistic interactions were assessed by calculating the fractional inhibitory concentration index (FICI); FICI = ([FLC]/[CFS]) + ([FLAC]/CFlaS), where [FLC] and [FLAC] are the concentrations of fluconazole and flavonoids that showed activity when combined, respectively, and [CFS] and [CFlaS] are the concentrations of these drugs with activities in isolation, respectively. This interpretation was performed according to the FICI value, where a FICI value of <0.5 indicates synergism (SYN), 0.5 < FICI ≤ 4.0 indicates indifference (IND), and a FICI value of >4.0 indicates antagonism (ANT). The strains C. parapsilosis ATCC 22019 and C. krusei ATCC 6258 were used as controls (22, 24).
Cell treatments.
For flow cytometry and comet assay experiments, single strains of fluconazole-resistant C. tropicalis were exposed to increasing concentrations (0.25 to 128 μg/ml) of the tested flavonoids, each combined with various concentrations (0.125 to 64 μg/ml) of fluconazole, for 24 h. Also, fluconazole-susceptible strains were treated with fluconazole (64 μg/ml) for 24 h at 35°C (26–28). Amphotericin B (Sigma Chemicals) was used as a cell death control, as its toxic effects induce apoptotic cell death in yeasts, characterized by the occurrence of nuclear chromatin condensation and fragmentation and the accumulation of reactive oxygen species (ROS) (29, 30). All experiments were performed in triplicate in three independent experiments.
Preparation of yeast cell suspensions.
Yeast cell suspensions were prepared from cultures in exponential growth phase. The cells were harvested, centrifuged (1,600 × g for 10 min at 4°C), washed twice with a 0.85% saline solution (1,200 × g for 5 min at 4°C), and then resuspended (∼106 cells/ml) in HEPES buffer from Sigma Chemicals supplemented with 2% glucose at pH 7.2 (22, 28).
Determination of cell density and membrane integrity.
The cell density and membrane integrity of the fungal strains were evaluated by the exclusion of 2 μg/ml propidium iodide (PI). Aliquots removed after 24 h of incubation with drugs were analyzed using flow cytometry. Ten thousand events were evaluated per experiment (n = 3), and cell debris was omitted from the analysis. Cell fluorescence was determined using flow cytometry in a Guava EasyCyte minisystem cytometer (Guava Technologies, Inc., Hayward, CA) with CytoSoft (version 4.1) software (22, 31, 32).
Detection of ROS in yeast.
For the detection of ROS produced over a 24-h culture period, cells were incubated with 20 μM 5-(and-6)-chloromethyl-2′,7′-dichlorodihydrofluorescein diacetate acetyl ester (CM-H2DCFDA) for 30 min in the dark at 35°C. Subsequently, the cells were harvested, washed, resuspended in phosphate-buffered saline (PBS), and immediately analyzed by flow cytometry (Guava EasyCyte minisystem; Guava Technologies, Inc., Hayward, CA). CM-H2DCFDA readily diffuses through the cell membrane, and it is hydrolyzed by intracellular esterases to form nonfluorescent dichlorofluorescein (DCFH), which is then rapidly oxidized to highly fluorescent 2′,7′-dichlorofluorescein (DCF) by any one of a broad range of intracellular oxidative stressors, aside from H2O2 (33). The fluorescence intensity of the DCF formed is proportional to the amount of ROS formed intracellularly (34).
Measurement of Δψm.
The mitochondrial transmembrane potential (Δψm) was determined by measurement of the retention of rhodamine-123 dye by fungal strains after 24 h of exposure. Cells were washed with PBS, incubated with rhodamine-123 (1 μg/ml) at 37°C for 30 min in the absence of light, and washed twice with PBS. Fluorescence was then measured using flow cytometry (Guava EasyCyte minisystem). Ten thousand events were evaluated per experiment (n = 3), and cell debris was omitted from the analysis (22, 35).
Yeast comet assay.
The alkaline comet assay was performed essentially as described by Miloshev et al. (2002) (36). Up to 200 μl of 0.5% agarose (normal melting point) was spread onto slides, and this supportive agarose layer was air dried before the application of cell suspensions onto the slides. Yeast cells were collected by centrifugation in an Eppendorf microcentrifuge for 5 min, washed with water, and resuspended in S buffer (1 M sorbitol, 25 mM KH2PO4, pH 6.5). Aliquots of approximately 5 × 104 cells/ml were mixed with 0.7% low-melting-point agarose containing 2 μg/ml Zymolyase 20T (Seikagaku Corp.), and the mixture was spread over the slides, covered with coverslips, and incubated for 20 min at 30°C to disintegrate the yeast cell walls and obtain spheroplasts. To minimize the activity of endogenous cellular enzymes, all further procedures were conducted in a cold room at 8 to 10°C. The coverslips were removed, and the slides were incubated in 30 mM NaOH, 1 M NaCl, 0.1% laurylsarcosine, 50 mM EDTA, pH 12.3, for 1 h to lyse the spheroplasts. The slides were rinsed three times for 20 min each time in 30 mM NaOH, 10 mM EDTA, pH 12.4, to unwind the DNA and then subjected to electrophoresis in the same buffer. The electrophoresis was carried out for 20 min at 0.5 V/cm and 24 mA. After electrophoresis, the slides were neutralized by submerging the slides in 10 mM Tris-HCl, pH 7.5, for 10 min, followed by consecutive 10-min incubations in 76% and 96% ethanol. Finally, the slides were air dried, stained with ethidium bromide (1 μg/ml), and visualized under a fluorescence microscope (31). All of the steps described above were conducted under dark conditions to prevent additional DNA damage. Images of 100 randomly selected cells (50 cells from each of the two replicate slides) were analyzed for each experimental group. The cells were scored visually and assigned to one of five classes, according to the tail length (from undamaged [score, 0] to maximally damaged [score, 4]), and a damage index value was calculated for each sample of cells. The damage index ranged from 0 (completely undamaged, 100 cells × 0) to 400 (maximum damage, 100 cells × 4) (37). The frequency of tailed cells, a DNA damage frequency indicator, was calculated on the basis of the number of cells with tails (indicating DNA strand breaks) versus the number of cells with no tails (22).
Annexin V staining.
Treated and untreated C. tropicalis cells were harvested by centrifugation and digested with 2 mg/ml Zymolyase 20T (Seikagaku Corp., Japan) in potassium phosphate buffer (PPB; 1 M sorbitol, pH 6.0) for 2 h at 30°C. Protoplasts of C. tropicalis were stained with fluorescein isothiocyanate (FITC)-labeled annexin V and PI using an FITC-annexin V apoptosis detection kit (Guava Nexin kit; Guava Technologies, Inc., Hayward, CA). Subsequently, cells were washed with PPB and incubated in annexin binding buffer containing 5 μl/ml FITC-annexin V and 5 μl of PI for 20 min. The cells were then analyzed by flow cytometry (Guava EasyCyte minisystem). For each experiment (n = 3), 10,000 events were evaluated, and cell debris was omitted from the analysis (38).
Leukocyte isolation and cultures.
Blood was collected from healthy, nonsmoker donors who had not taken any drugs for at least 15 days prior to sampling and placed in heparin, and leukocytes were isolated using density gradient centrifugation over Histopaque-1077. The protocols applied for isolation of leukocytes from whole blood were approved by the Ethics Committee in Research of the Federal University of Ceara (47/2013). Cells were washed and resuspended in RPMI 1640 medium supplemented with 20% fetal bovine serum, 2 mM glutamine, 100 U/ml penicillin, 100 μg/ml streptomycin (Invitrogen, Carlsbad, CA) at 37°C under a 5% CO2 atmosphere. Phytohemagglutinin (2%; Cultilab, Campinas, SP, Brazil) was added at the start of the cultures. After 24 h, cells were treated with the test compounds, as reported by Cavalcanti and coworkers (39).
Flavonoid toxicity to leukocytes.
The cytotoxicity of the tested compounds to leukocytes was evaluated by the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay (Sigma Chemicals, St. Louis, MO) (40). Briefly, cells were plated in 96-well plates (1.5 × 106 cells/ml). Compounds were dissolved in 1% DMSO (Sigma Chemical) at 0.19 to 100 μg/ml, and the resulting solutions were added to wells. After 72 h of exposure, the supernatant was replaced by fresh medium containing MTT (0.5 mg/ml). After 3 h, the MTT formazan product was dissolved in DMSO and the absorbance was measured at 595 nm (Beckman Coulter DTX-880 spectrometer) (39).
Statistical analysis.
The experiments measuring susceptibility, the in vitro profiles of synergism, and expression were repeated at least three times on different days. Arithmetic means and standard deviations were used to statistically analyze continuous variables (FICI). The geometric means were used to compare the MIC50 results statistically. The data obtained by flow cytometry were compared using one-way analysis of variance (ANOVA), followed by the Newman-Keuls test (P < 0.05). The data obtained by the MTT assay are presented as means ± standard errors of the means. Fifty percent inhibitory concentration (IC50) values and 95% confidence intervals (CIs) were obtained by nonlinear regression using the GraphPad program (Intuitive Software for Science, San Diego, CA).
RESULTS
Synergistic effect of flavonoids and fluconazole.
The fluconazole susceptibility profiles of the C. tropicalis strains were assessed using the microdilution technique previously described (23). Table 1 shows that there was no variation in the susceptibility of the different strains tested with fluconazole. All strains studied showed MIC50 values of 64 μg/ml. The synergism between flavonoids and fluconazole was determined using the checkerboard technique, in which the association of flavonoids with fluconazole showed a synergistic effect on fluconazole-resistant strains and exhibited FICIs ranging from 0.25 to 0.38 μg/ml (a synergistic effect is an FICI of <0.5).
TABLE 1.
Synergistic effect of fluconazole with CATEQ, QUERC, and EPIG against strains of Candida tropicalis resistant to fluconazole
| C. tropicalis straina | MIC50 (μg/ml)b |
FLC-FLAV interaction |
|||||||
|---|---|---|---|---|---|---|---|---|---|
| Standardc |
Combinationd |
FICI |
Interpretatione | ||||||
| FLC | CATEQ, QUERC, and EPIG | CATEQ + FLC 16 | QUERC + FLC 16 | EPIG + FLC 16e | CATEQ + FLC | FICI QUERC + FLC | FICI EPIG + FLC | ||
| 1 | 64 | >128 | 16 | 0.50 | 0.50 | 0.38 | 0.25 | 0.25 | SYN |
| 2 | 64 | >128 | <0.25 | <0.25 | <0.25 | 0.25 | 0.25 | 0.25 | SYN |
| 3 | 64 | >128 | <0.25 | <0.25 | 1.0 | 0.25 | 0.25 | 0.26 | SYN |
| 4 | 64 | >128 | 0.25 | 0.25 | 0.50 | 0.25 | 0.25 | 0.25 | SYN |
| 5 | 64 | >128 | 0.25 | 0.25 | <0.25 | 0.25 | 0.25 | 0.25 | SYN |
| 6 | 64 | >128 | 0.25 | <0.25 | <0.25 | 0.25 | 0.25 | 0.25 | SYN |
Yeasts isolated from biological samples.
FLC, fluconazole. The MIC50 value was defined as the lowest concentration that produced a 50% reduction in the growth of fungal cells after a 24-h incubation.
The procedure was performed according to the CLSI M27-A3 2008 protocol (23). Fluconazole concentrations ranged from 64 to 0.125 mg/liter, and flavonoid concentrations varied from 128 to 0.25 mg/liter.
The concentration of fluconazole was fixed at 16 mg/liter (FLC 16), and the flavonoid concentrations varied from 128 to 0.25 mg/liter. The synergistic effect of fluconazole and flavonoids was calculated on the basis of the FICI, which is equal to ([FLC]/[CFS]) + ([FLAC]/CFlaS), where [FLC] and [FLAC] are the concentrations of fluconazole and flavonoids that showed activity when combined, respectively, and [CFS] and [CFlaS] are the concentrations of these drugs with activities in isolation, respectively.
Interpretation was performed according to the value of FICI. An FICI of <0.5 is synergism (SYN), 0.5 < FICI ≤ 4.0 is indifference (IND), and an FICI of >4.0 is antagonism (ANT).
Cell treatments.
When the fluconazole-resistant strains were exposed to various concentrations (0.25 to 128 μg/ml) of flavonoids combined with various concentrations (0.25 to 16 μg/ml) of fluconazole for 24 h at 35°C, the best synergistic effect was achieved with 128 μg/ml of flavonoids combined with 16 μg/ml of fluconazole.
Loss of cell viability after cotreatment with flavonoids and fluconazole in C. tropicalis.
As shown in Fig. 2, the exposure of fluconazole-resistant strains to fluconazole did not cause a reduction in the number of viable cells compared to that for the control. However, cells treated with fluconazole in combination with flavonoids after 24 h of exposure showed a significant decrease (P < 0.05) in cell density: 18.47% ± 27.71% for catechin hydrate, 23.95% ± 3.32% for quercetin hydrate, and 8.33% ± 11.28% for epigallocatechin gallate compared to that for the control group.
FIG 2.
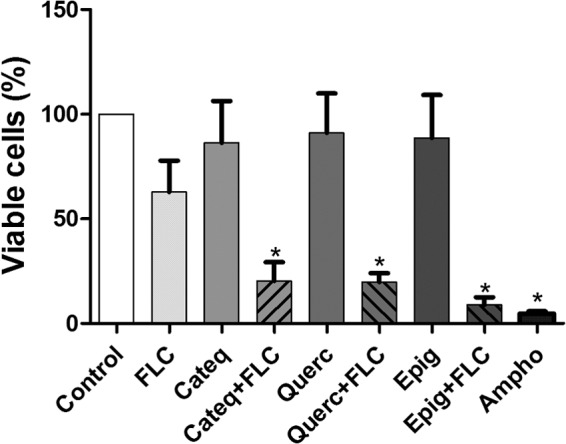
Effects of the different treatments on the viability of fluconazole (FLC)-resistant cells of C. tropicalis evaluated by flow cytometry after 24 h. Cells were treated with RPMI (negative control), amphotericin B (Ampho; 4 μg/ml; positive control), CATEQ, QUERC, and EPIG (128 μg/ml), fluconazole (64 μg/ml), and a fixed concentration of CATEQ, QUERC, and EPIG (128 μg/ml) with fluconazole (16 μg/ml). The data are presented as mean values ± SEMs from experiments performed in triplicate.
Changes in cell size/granularity by synergism of flavonoids and fluconazole.
Flow cytometry analysis (side scatter [SSC]-forward light scatter [FSC]) showed that resistant strains treated with fluconazole underwent cell shrinkage and nuclear condensation, as evidenced by the decrease in forward light scattering and a transient increase in side scattering, respectively. Interestingly, in all fluconazole-resistant C. tropicalis strains evaluated, changes in cell size/granularity were observed only after 24 h of exposure to fluconazole (16 μg/ml) in combination with flavonoids (Fig. 3).
FIG 3.

Analysis of changes in cell size/granularity (forward scatter-side scatter) in the presence of RPMI (negative control), amphotericin B (4 μg/ml; positive control), fluconazole (64 μg/ml), CATEQ, QUERC, and EPIG (128 μg/ml), and a fixed concentration of CATEQ, QUERC, and EPIG (128 μg/ml) with fluconazole (16 μg/ml) in isolated fluconazole-resistant C. tropicalis for a period of 24 h.
C. tropicalis plasma membrane damage.
Figure 4 shows the membrane integrity damage induced by the synergistic action of fluconazole plus (+)-catechin hydrate, quercetin hydrate, or (−)-epigallocatechin gallate in strains of C. tropicalis resistant to fluconazole. Treatment with fluconazole plus flavonoids (24 h of exposure) resulted in damage to the plasma membrane. Yeast cells treated with fluconazole in combination with the tested flavonoids for 24 h showed a significant increase (P < 0.05) in the population with membrane damage compared to the population with membrane damage in the control group (1.7% ± 0.58%): 39.43% ± 2.41% for catechin hydrate, 19.52% ± 1.27% for quercetin hydrate, and 13.61% ± 1.45% for epigallocatechin gallate.
FIG 4.

Effect of the different treatments of fluconazole for a period of 24 h on membrane integrity (determined by a PI exclusion test) in isolated fluconazole-resistant C. tropicalis. Cells were treated with RPMI (negative control), amphotericin B (4 μg/ml; positive control), fluconazole (64 μg/ml), CATEQ, QUERC, and EPIG (128 μg/ml) at a fixed concentration, and a fixed concentration of CATEQ, QUERC, and EPIG (128 μg/ml) with fluconazole (16 μg/ml) *, P < 0.05 compared to the control by ANOVA followed by the Newman-Keuls test.
Increased intracellular ROS generation induced by cotreatment with flavonoids and fluconazole in C. tropicalis.
In C. tropicalis resistant to fluconazole, increases (P < 0.05) in ROS levels were observed only in cultures coexposed to fluconazole (16 μg/ml) and the tested flavonoids (128 μg/ml) (Fig. 5).
FIG 5.
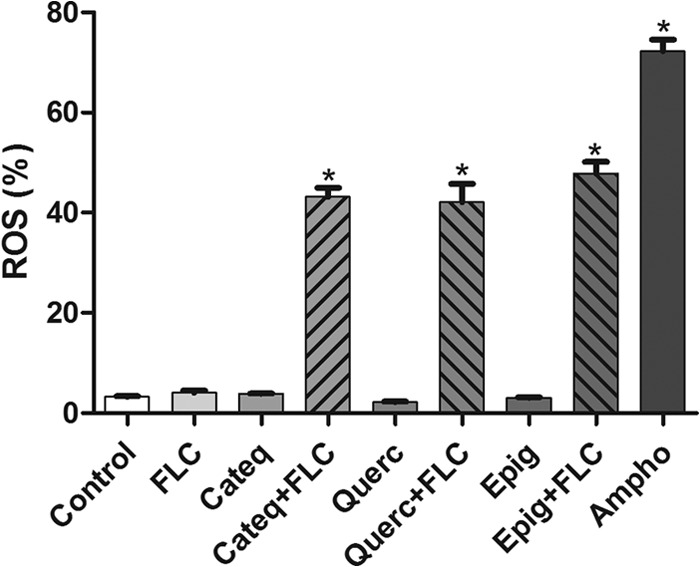
Percentages of DCF fluorescence-positive cells (indicating ROS production) in isolated fluconazole-resistant C. tropicalis after treatment with RPMI (negative control), amphotericin B (4 μg/ml; positive control), fluconazole (64 μg/ml); CATEQ, QUERC, and EPIG (128 μg/ml) at a fixed concentration, and a fixed concentration of CATEQ, QUERC, and EPIG (128 μg/ml) with fluconazole (16 μg/ml) *, P < 0.05 compared to the control by ANOVA followed by the Newman-Keuls test.
Phosphatidylserine externalization in C. tropicalis.
In Fig. 6, the population of cells in the lower right and upper right quadrants corresponds to early apoptotic cells (annexin V positive, 7-aminoactinomycin D [7AAD] negative) and late apoptotic cells (annexin V positive, 7AAD positive), respectively, with phosphatidylserine being externalized. After 24 h of exposure, the percentage of cells with externalized phosphatidylserine (the sums of early and late apoptotic stages) after a single treatment with the flavonoids (+)-catechin hydrate, quercetin hydrate, and (−)-epigallocatechin gallate and cultures treated only with fluconazole were very close to that for the negative-control cultures (0.14% ± 0.01%): 4.5% ± 1.2%, 4.5% ± 2.40%, 4% ± 2.42%, and 1.16% ± 1.2%, respectively. After 24 h of incubation, yeast cultures cotreated with fluconazole and the tested flavonoids, (+)-catechin hydrate, quercetin hydrate, and (−)-epigallocatechin gallate, showed a significant increase (P < 0.05) in the apoptotic cell percentages compared to the percentage for the control group (0.14% ± 0.01%): 36.5% ± 8.22%, 46.5% ± 18.8%, and 53% ± 10.9%, respectively. The association between fluconazole and the tested flavonoids clearly induced cell death at a level similar to that for amphotericin B, which was used as a positive control.
FIG 6.

Phosphatidylserine externalization, indicating early-stage apoptosis, shown by annexin V staining. The intensity of the fluorescence indicates the amount of exposed phosphatidylserine on cells treated with fluconazole (64 μg/ml), CATEQ, QUERC, and EPIG (128 μg/ml), amphotericin B (4 μg/ml; positive control), and a fixed concentration of CATEQ, QUERC, and EPIG (128 μg/ml) with fluconazole (16 μg/ml) for 24 h. *, P < 0.05 compared to the control by ANOVA followed by the Newman-Keuls test. PE, phycoerythrin.
Cotreatment with flavonoids and fluconazole induces mitochondrial dysfunction.
The strains of C. tropicalis cotreated with fluconazole (16 μg/ml) and flavonoids (128 μg/ml) showed mitochondrial dysfunction, characterized by reduction of the mitochondrial transmembrane potential (Δψm) after 24 h of exposure (Fig. 7).
FIG 7.
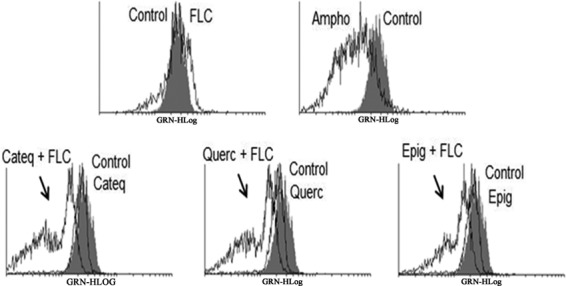
Histograms obtained by flow cytometry analysis of green fluorescence (GRN-XLog) of fluconazole-resistant C. tropicalis. The fluorescence of the cells shows the effects of different treatments on the mitochondrial transmembrane potential in strains exposed for 24 h to RPMI (negative control), amphotericin B (4 μg/ml; positive control), fluconazole (64 μg/ml), CATEQ, QUERC, and EPIG (128 μg/ml) at a fixed concentration, and a fixed concentration of CATEQ, QUERC, and EPIG (128 μg/ml) with fluconazole (16 μg/ml).
DNA damage induced by cotreatment with flavonoids and fluconazole or single treatments in C. tropicalis.
Figure 8 shows the DNA damage induced by the synergistic action of fluconazole plus (+)-catechin hydrate, quercetin hydrate, or (−)-epigallocatechin gallate on strains of C. tropicalis resistant to fluconazole. The individual analysis of single cells regarding the distribution of grades of DNA damage (Fig. 8A) showed that the flavonoids induced low levels of DNA damage (mainly grade 1). Moreover, corroborating that, cultures treated with single flavonoids alone did not suffer DNA damage, as evidenced by the means of the damage index and frequency (Fig. 8B and C). In contrast, C. tropicalis coexposure to fluconazole and the tested flavonoids for 24 h resulted in a significant increase (P < 0.05) in DNA strand break levels (Fig. 8A to C). Cells treated with fluconazole in combination with the flavonoid (+)-catechin hydrate, quercetin hydrate, or (−)-epigallocatechin gallate for 24 h exhibited damage index values (arbitrary units) of 64.38 ± 2.15, 74.62 ± 3.15, and 61.48 ± 2.56, respectively, and damage frequencies of 36.95% ± 3.17%, 31.83% ± 0.21%, and 29.42% ± 0.10%, respectively. Amphotericin B, used as a positive control, induced high levels of DNA strand breaks.
FIG 8.
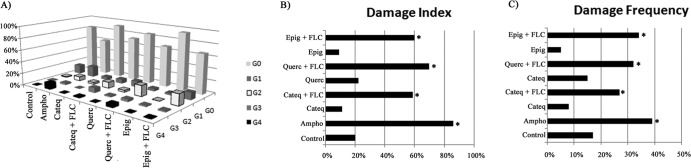
Effects of different treatments on the distribution of damage classes (grades [G] 0 to 4) of DNA caused by fluconazole after 24 h of exposure. The yeasts were exposed to RPMI (negative control), fluconazole (64 μg/ml), CATEQ, QUERC, and EPIG (128 μg/ml), amphotericin B (4 μg/ml; positive control), and a fixed concentration of CATEQ, QUERC, and EPIG (128 μg/ml) with fluconazole (16 μg/ml) for 24 h. *, P < 0.05 compared to the control by ANOVA followed by the Newman-Keuls test.
Cytotoxic activity of flavonoids in leukocytes.
Table 2 shows that the flavonoids quercetin and epigallocatechin gallate hydrate showed moderate cytotoxicity against human leukocytes, as analyzed by the MTT assay, compared with the results for the control group (P < 0.05). However, catechin hydrate showed no cytotoxicity when treated alone or in combination with fluconazole when cytotoxicity was compared to that for the control.
TABLE 2.
Cytotoxic activity of flavonoids on leukocytesa
| Compound | Leukocyte IC50 (μg/ml−1) |
|
|---|---|---|
| Without FLC | With FLC | |
| QUER | 17.45 (14.34–21.24) | 14.97 (13.06–17.17) |
| EPIG | 41.98 (38.57–51.68) | 40.42 (34.90–43.06) |
| CATEQ | >100 | >100 |
| FLC | >100 | |
Fluconazole (FLC) was used as a positive control. Data are presented as IC50 values and 95% CIs (in parentheses) from three independent experiments performed in triplicate.
DISCUSSION
Our findings suggest a potential synergistic effect of the combination of fluconazole with the tested flavonoids. Hirasawa and Takada (41) observed that various catechins present in green tea have significant antifungal activities when combined with antifungal drugs. Several recent studies have shown that epigallocatechin has antifungal activity against dermatophytes and yeasts as a single treatment or combined with other antifungal agents and can be applied as an alternative antifungal agent for fungal species resistant to traditional drugs (15, 19, 42). In our study, (−)-epigallocatechin gallate alone showed no antifungal activity, but when combined with fluconazole, it showed a synergistic effect against strains of C. tropicalis resistant to fluconazole. These findings corroborate those reported by Hirasawa and Takada (41), whose strains of C. albicans resistant to fluconazole demonstrated sensitivity to the combination of epigallocatechin gallate with fluconazole, suggesting that this combination might be useful in treating superinfections (43). Several antimicrobial effects have been observed for quercetin (44, 45). However, little is known about the antifungal activity of this flavonoid. The present data show that quercetin has no activity when used as a single treatment, but when combined with fluconazole, quercetin demonstrated a potent synergistic effect against strains of C. tropicalis resistant to fluconazole.
The molecular basis of the fluconazole resistance of the C. tropicalis strains investigated in the present study is currently unknown. However, on the basis of what is known for other characterized strains, we can speculate that it may involve the overexpression of efflux pumps encoded by either MDR or CDR genes or the acquisition of point mutations in the gene encoding ERG11 (46).
When cells were exposed to fluconazole combined with the flavonoids (+)-catechin hydrate, quercetin hydrate, and (−)-epigallocatechin gallate in the presence of a PI, a major portion of the cells became PI positive in comparison to the results for the control cultures. The increased PI uptake in the cells of fluconazole-resistant C. tropicalis treated with flavonoids and fluconazole demonstrates that these combinations can change the cell membrane structure, resulting in the loss of plasma membrane integrity in fungal cells and causing increased permeability. A study by Toyoshima et al. (47) described a similar mechanism of action of catechin against isolates of Trichophyton mentagrophytes through electron microscopy and suggested that catechin may act by lysing the cell membrane (41). This may explain the fact that cells treated with fluconazole combined with catechin showed greater damage to the membrane.
Fungal cells treated with the proposed synergistic combination showed increased levels of ROS, leading to the generation of hydroxyl radicals. Although ROS act as signal transducers, when accumulated in cells in the form of hydrogen peroxide, superoxide, and hydroxyl radicals, ROS are considered essential regulators of aging and have been reported to be a key element in the apoptosis of yeast (48–50). The synergistic interactions observed in this study indicate a probable prooxidant activity of flavonoids, which may be responsible for the induction of transcription factors associated with apoptosis and related to increased levels of proapoptotic proteins, mitochondrial injury, and ROS generation, as well as accelerated oxidative damage to DNA, proteins, and carbohydrates in vitro (51–53).
The flavonoids (+)-catechin hydrate, quercetin hydrate, and (−)-epigallocatechin gallate, when combined with fluconazole, promoted changes in the mitochondrial membrane potential. Tests verified mitochondrial dysfunction in the treated cells, suggesting that the synergistic combinations affect the mitochondrial respiratory function, preventing rhodamine-123 from accumulating in the mitochondria (35). Such a Δψm collapse can lead to transient pore openings in the mitochondrial membrane and the release of proapoptotic factors into the cytosol (50, 54). This fact can be explained by the ability of flavonoids to exhibit a prooxidant activity (52, 53) that favors increased intracellular levels of hydroxyl radicals that lead to mitochondrial membrane damage. Hwang et al. (55) showed that the flavonoid amentoflavone promotes mitochondrial dysfunction in C. albicans strains due to increased levels of ROS. Therefore, the flavonoids used synergistically with fluconazole in the present study seemed to indirectly promote the mitochondrial dysfunction as a result of increased levels of ROS. The increased intracellular ROS levels and mitochondrial dysfunction play an important role in apoptosis induction (48, 49, 56).
The treatment of C. tropicalis strains in this study with fluconazole plus flavonoids promoted DNA damage. However, the combination of fluconazole with quercetin hydrate showed greater damage to the DNA than the other combinations. This is clearly because quercetin, which exhibits two aromatic rings in its structure, can penetrate the phospholipid membranes (43, 57, 58) due to the hydrophobic nature of the molecule. However, with treatment with the combination of fluconazole and epigallocatechin gallate, a greater number of cells showed some type of DNA damage, regardless of the degree of injury. Condensation and extensive DNA fragmentation are features that often occur in the early stages of apoptosis, representing an irreversible step that leads to cell death (50, 59, 60).
The detection of apoptosis at an early stage can be determined using annexin V as a marker. In the presence of Ca2+, annexin binds with a high affinity to the phosphatidylserine present in the membranes of apoptotic cells (50). Our experimental evidence indicates that the combination of flavonoids with fluconazole induces apoptotic cell death in C. tropicalis, in which the generation and intracellular accumulation of reactive oxygen species seem to act as stimulators of early apoptosis signaling, in addition to directly damaging the mitochondria and the nuclear DNA. These data corroborate the results of Hwang et al. (50) and Cho and Lee (49), who found similar characteristics of cell death in yeasts treated with antimicrobial peptides.
The synergistic effect of fluconazole with flavonoids promotes exposure of the phosphatidylserine in the plasma membrane, changes in cell size/granularity, mitochondrial membrane depolarization, intracellular ROS accumulation, and DNA fragmentation in fluconazole-resistant strains of C. tropicalis. Based on the characteristics of cell death observed, we hypothesize that the proposed synergism exerts its antifungal activity via increased intracellular ROS, resulting in apoptosis.
The use of (+)-catechin hydrate alone or in association with fluconazole did not cause any cytotoxic effects on cultured peripheral human leukocytes (IC50 > 100). Corroborating our findings, Babich et al. (61) reported that catechin exhibited a lower toxicity than other compounds derived from catechin against HSC-2 carcinoma cells and HGF-2 fibroblasts. Although additional tests, such as reproductive toxicity analysis and mutagenesis evaluation, must be performed, the present results show that (+)-catechin hydrate plus fluconazole is probably safe for use for treatment of acute infections in vivo. In summary, the results suggest that the flavonoid (+)-catechin hydrate has potential as an adjuvant agent in combination with antifungals for the treatment of candidemias, although a study with a higher number of strains would be required to establish this conclusion.
In conclusion, combinations of the flavonoids (+)-catechin hydrate, quercetin hydrate, or (−)-epigallocatechin gallate with fluconazole demonstrated antifungal activity against strains of fluconazole-resistant C. tropicalis in vitro. Despite changing the plasma and mitochondrial membrane integrity, the synergism also seemed to interact with the DNA, leading to death by apoptosis, possibly due to the intracellular accumulation of ROS. The flavonoid catechin hydrate showed no toxicity toward the leukocytes.
ACKNOWLEDGMENTS
This work was supported by grants and fellowships from the National Council of Technological and Scientific Development (CNPq), Coordination for the Improvement of Higher Level or Education Personnel (CAPES/Brazil), and the Foundation of Ceara Support for Scientific and Technology (FUNCAP/Ceara).
We declare that we have no conflicts of interest concerning this article.
Footnotes
Published ahead of print 23 December 2013
REFERENCES
- 1.Hitoto H, Pihet M, Weil B, Chabasse D, Jean-Philippe B, Rachieru-Sourisseau P. 2010. Acremonium strictum fungaemia in a paediatric immunocompromised patient: diagnosis and treatment difficulties. Mycopathologia 170:161–164. 10.1007/s11046-010-9306-5 [DOI] [PubMed] [Google Scholar]
- 2.Araujo MF, Vieira IJC, Braz-Filho R, Vieira-da-Motta O, Mathias L. 2009. Chemical constituents from Swartzia apetala Raddi var. glabra and evaluation of their antifungal activity against Candida spp. Braz. J. Pharmacogn. 19:366–369. 10.1590/S0102-695X2009000300005 [DOI] [Google Scholar]
- 3.Lyon GM, Karatela S, Sunay S, Adiri Y. 2010. Antifungal susceptibility testing of Candida isolates from the Candida surveillance study. J. Clin. Microbiol. 48:1270–1275. 10.1128/JCM.02363-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Diekema D, Arbefeville S, Boyken L, Kroeger J, Pfaller M. 2012. The changing epidemiology of healthcare-associated candidemia over three decades. Diagn. Microbiol. Infect. Dis. 73:45–48. 10.1016/j.diagmicrobio.2012.02.001 [DOI] [PubMed] [Google Scholar]
- 5.Nucci M, Colombo AL. 2007. Candidemia due to Candida tropicalis: clinical, epidemiologic, and microbiologic characteristics of 188 episodes occurring in tertiary care hospitals. Diagn. Microbiol. Infect. Dis. 58:77–82. 10.1016/j.diagmicrobio.2006.11.009 [DOI] [PubMed] [Google Scholar]
- 6.Kothavade RJ, Kura MM, Valand AG, Panthaki MH. 2010. C. tropicalis: its prevalence, pathogenicity and increasing resistance to fluconazole. J. Med. Microbiol. 59:873–880. 10.1099/jmm.0.013227-0 [DOI] [PubMed] [Google Scholar]
- 7.Tobudic S, Kratzer C, Presterl E. 2012. Azole-resistant Candida spp.—emerging pathogens? Mycoses 55:24–32. 10.1111/j.1439-0507.2011.02146.x [DOI] [Google Scholar]
- 8.Chi HW, Yang YS, Shang ST, Chen KH, Yehb KM, Changb FY, Lin JC. 2011. Candida albicans versus non-albicans bloodstream infections: the comparison of risk factors and outcome. J. Microbiol. Immunol. Infect. 44:369–375. 10.1016/j.jmii.2010.08.010 [DOI] [PubMed] [Google Scholar]
- 9.Gonzalez GM, Elizondo M, Ayala J. 2008. Trends in species distribution and susceptibility of bloodstream isolates of Candida collected in Monterrey, Mexico, to seven antifungal agents: results of a 3-year (2004 to 2007) surveillance study. J. Clin. Microbiol. 46:2902–2905. 10.1128/JCM.00937-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Simões M, Lemos M, Simões LC. 2012. Phytochemicals against drug-resistant microbes, p 185–205 In Patra AK. (ed), Dietary phytochemicals and microbes. Springer, Dordrecht, Netherlands. 10.1007/978-94-007-3926-0_6 [DOI] [Google Scholar]
- 11.Maurya IK, Pathak S, Sharma M, Sanwal H, Chaudhary P, Tupec S, Deshpande M, Singh Chauhan V, Prasada R. 2011. Antifungal activity of novel synthetic peptides by accumulation of reactive oxygen species (ROS) and disruption of cell wall against Candida albicans. Peptides 32:1732–1740. 10.1016/j.peptides.2011.06.003 [DOI] [PubMed] [Google Scholar]
- 12.Rajeshkumar R, Sundararaman M. 2011. Emergence of Candida spp. and exploration of natural bioactive molecules for anticandidal therapy—status quo. Mycoses 55:60–73. 10.1111/j.1439-0507.2011.02156.x [DOI] [PubMed] [Google Scholar]
- 13.Saini ML, Saini R, Roy S, Kumar A. 2008. Comparative pharmacognostical and antimicrobial studies of Acacia species (Mimosaceae). J. Med. Plants Res. 12:378–386 [Google Scholar]
- 14.Duraipandiyan V, Ayyanar M, Ignacimuthu S. 2006. Antimicrobial activity of some ethnomedicinal plants used by Paliyar tribe from Tamil Nadu, India. BMC Complement. Altern. Med. 6:35. 10.1186/1472-6882-6-35 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Park BJ, Park JC, Taguchi H, Fukushimae K, Hyonf SH, Takatori K. 2011. In vitro antifungal activity of epigallocatechin 3-O-gallate against clinical isolates of dermatophytes. Yonsei Med. J. 52:535–538. 10.3349/ymj.2011.52.3.535 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Cavaleiro C, Pinto E, Gonçalves MJ, Salgueiro L. 2006. Antifungal activity of Juniperus essential oils against dermatophyte, Aspergillus and Candida strains. J. Appl. Microbiol. 100:1333–1338. 10.1111/j.1365-2672.2006.02862.x [DOI] [PubMed] [Google Scholar]
- 17.Pyun MS, Shin S. 2006. Antifungal effects of the volatile oils from Allium plants against Trichophyton species and synergism of the oils with ketoconazole. Phytomedicine 13:394–400. 10.1016/j.phymed.2005.03.011 [DOI] [PubMed] [Google Scholar]
- 18.Mondello F, De Bernardis F, Girolamo A, Cassone A, Salvatore G. 2006. In vivo activity of terpinen-4-ol, the main bioactive component of Melaleuca alternifolia Cheel (tea tree) oil against azole-susceptible and -resistant human pathogenic Candida species. BMC Infect. Dis. 6:158. 10.1186/1471-2334-6-158 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Han Y. 2007. Synergic anticandidal effect of epigallocatechin-O-gallate combined with amphotericin B in a murine model of disseminated candidiasis and its anticandidal mechanism. Biol. Pharm. Bull. 30:1693–1696. 10.1248/bpb.30.1693 [DOI] [PubMed] [Google Scholar]
- 20.Liu CM, Zheng YL, Lu J, Lua J, Zhanga ZF, Fana SH, Wua DM, Mab JQ. 2010. Quercetin protects rat liver against lead-induced oxidative stress and apoptosis. Environ. Toxicol. Pharmacol. 29:158–166. 10.1016/j.etap.2009.12.006 [DOI] [PubMed] [Google Scholar]
- 21.Gordon CN, Wareham WD. 2010. Antimicrobial activity of the green tea polyphenol (−)-epigallocatechin-3-gallate (EGCG) against clinical isolates of Stenotrophomonas maltophilia. Int. J. Antimicrob. Agents 36:129–131. 10.1016/j.ijantimicag.2010.03.025 [DOI] [PubMed] [Google Scholar]
- 22.da Silva CR, de Andrade Neto JB, Sidrim JJC, Ângelo MRF, Magalhães HIF, Cavalcanti BC, Brilhante RSN, Macedo DS, Moraes MO, Lobo MDP, Grangeiro TB, Nobre Júnior HV. 2013. Synergistic effects of amiodarone and fluconazole on Candida tropicalis resistant to fluconazole. Antimicrob. Agents Chemother. 57:1691–1700. 10.1128/AAC.00966-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Clinical and Laboratory Standards Institute 2008. Reference method for broth dilution antifungal susceptibility testing of yeasts. Approved standard M27-A3, 3rd ed. Clinical and Laboratory Standards Institute, Wayne, PA [Google Scholar]
- 24.Clinical and Laboratory Standards Institute 2012. Reference method for broth dilution antifungal susceptibility testing of yeasts. Fourth informational supplement. M27-S4. Clinical and Laboratory Standards Institute, Wayne, PA [Google Scholar]
- 25.Endo EH. 2007. Synergistic effect of the crude extract and fractions of Punica granatum against Candida albicans and synergy with fluconazole. Dissertation. State University of Maringá, Maringá, Brazil [Google Scholar]
- 26.Rudensky B, Broide E, Berko N, Wiener-Well Y, Yinnon AMM, Raveh D. 2008. Direct fluconazole susceptibility testing of positive Candida blood cultures by flow cytometry. Mycoses 51:200–204. 10.1111/j.1439-0507.2007.01466.x [DOI] [PubMed] [Google Scholar]
- 27.Pina-Vaz C, Rodrigues AG, Costa-de-Oliveira S, Ricardo E, Mardh PA. 2005. Potent synergic effect between ibuprofen and azoles on Candida resulting from blockade of efflux pumps as determined by FUN-1 staining and flow cytometry. J. Antimicrob. Chemother. 56:678–685. 10.1093/jac/dki264 [DOI] [PubMed] [Google Scholar]
- 28.Pina-Vaz C, Rodrigues AG. 2010. Evaluation of antifungal susceptibility using flow cytometry molecular and cell biology methods for fungi. Methods Mol. Biol. 638:281–289. 10.1007/978-1-60761-611-5_21 [DOI] [PubMed] [Google Scholar]
- 29.Phillips AJ, Sudbery I, Ramsdale M. 2003. Apoptosis induced by environmental stresses and amphotericin B in Candida albicans. Proc. Natl. Acad. Sci. U. S. A. 100:14327–14332. 10.1073/pnas.2332326100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Almeida B, Silva A, Mesquita A, Sampaio-Marques B, Rodrigues F, Ludovico P. 2008. Drug-induced apoptosis in yeast. Biochim. Biophys. Acta 1783:1436–1448. 10.1016/j.bbamcr.2008.01.005 [DOI] [PubMed] [Google Scholar]
- 31.Pinkerton DM, Banwell MG, Garson MJ, Kumar N, de Moraes MO, Cavalcanti BC, Barros FWA, Pessoa C. 2010. Antimicrobial and cytotoxic activities of synthetically derived tambjamines C and E-J, BE-18591, and a related alkaloid from the marine bacterium Pseudoalteromonas tunicate. Chem. Biodivers. 7:1311–1324. 10.1002/cbdv.201000030 [DOI] [PubMed] [Google Scholar]
- 32.Joung YH, Kim HR, Lee MK, Park AJ. 2007. Fluconazole susceptibility testing of Candida species by flow cytometry. J. Infect. 54:504–508. 10.1016/j.jinf.2006.09.016 [DOI] [PubMed] [Google Scholar]
- 33.Hempel SL, Buettner GR, O'Malley YQ, Wessels DA, Flaherty DM. 1999. Dihydrofluorescein diacetate is superior for detecting intracellular oxidants: comparison with 2′,7′-dichlorodihydrofluorescein diacetate, 5(and 6)-carboxy-2′,7′-dichlorodihydrofluorescein diacetate, and dihydrorhodamine 123. Free Radic. Biol. Med. 27:146–159. 10.1016/S0891-5849(99)00061-1 [DOI] [PubMed] [Google Scholar]
- 34.LeBel CP, Ischiropoulos H, Bondy SC. 1992. Evaluation of the probe 2′,7′-dichlorofluorescein as an indicator of reactive oxygen species formation and oxidative stress. Chem. Res. Toxicol. 5:227–231. 10.1021/tx00026a012 [DOI] [PubMed] [Google Scholar]
- 35.Ludovico P, Sansonetty F, Côrte-Real M. 2001. Assessment of mitochondrial membrane potential in yeast cell populations by flow cytometry. Microbiology 147:3335–3343 [DOI] [PubMed] [Google Scholar]
- 36.Miloshev G, Mihaylov I, Anachkova B. 2002. Application of the single cell electrophoresis on yeast cells. Mutat. Res. 513:69–74. 10.1016/S1383-5718(01)00286-8 [DOI] [PubMed] [Google Scholar]
- 37.Collins AR. 2004. The comet assay for DNA damage and repair: principles, applications, and limitations. Mol. Biotechnol. 26:249–261. 10.1385/MB:26:3:249 [DOI] [PubMed] [Google Scholar]
- 38.Cavalcanti BC, Costa PM, Carvalho AA, Rodrigues FAR, Amorim RCN, Silva ECC, Pohlit AM, Costa-Lotufo LV, Moraes MO, Pessoa C. 2012. Involvement of intrinsic mitochondrial pathway in neosergeolide-induced apoptosis of human HL-60 leukemia cells: the role of mitochondrial permeability transition pore and DNA damage. Pharm. Biol. 50:980–993. 10.3109/13880209.2012.654921 [DOI] [PubMed] [Google Scholar]
- 39.Cavalcanti BC, Bezerra DP, Magalhães HI, Moraes MO, Lima MA, Silveira ER, Câmara CA, Rao VS, Pessoa C, Costa-Lotufo LV. 2009. Kauren-19-oic acid induces DNA damage followed by apoptosis in human leukemia cells. J. Appl. Toxicol. 29:560–568. 10.1002/jat.1439 [DOI] [PubMed] [Google Scholar]
- 40.Mosmann T. 1983. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J. Immunol. Methods 65:55–63. 10.1016/0022-1759(83)90303-4 [DOI] [PubMed] [Google Scholar]
- 41.Hirasawa M, Takada K. 2004. Multiple effects of green tea catechin on the antifungal activity of antimycotics against Candida albicans. J. Antimicrob. Chemother. 53:225–229. 10.1093/jac/dkh046 [DOI] [PubMed] [Google Scholar]
- 42.Park BJ, Taguchi H, Kamei K, Matsuzawa T, Hyon SH, Park JC. 2006. Antifungal susceptibility of epigallocatechin 3-O-gallate (ECGg) on clinical isolates of pathogenic yeast. Biochem. Biophys. Res. Commun. 347:401–405. 10.1016/j.bbrc.2006.06.037 [DOI] [PubMed] [Google Scholar]
- 43.Daglia M. 2012. Polyphenols as antimicrobial agents. Curr. Opin. Biotechnol. 23:174–181. 10.1016/j.copbio.2011.08.007 [DOI] [PubMed] [Google Scholar]
- 44.Ramadan MF, Asker MM. 2009. Antimicrobial and antiviral impact of novel quercetin-enriched lecithin. J. Food Biochem. 33:557–571. 10.1111/j.1745-4514.2009.00237.x [DOI] [Google Scholar]
- 45.Rodriguez-Vaquero MJ, Alberto MR, Manc-de-Nadra MC. 2007. Antibacterial effect of phenolic compounds from different wines. Food Control 18:93–101. 10.1016/j.foodcont.2005.08.010 [DOI] [Google Scholar]
- 46.Pfaller MA. 2012. Antifungal drug resistance: mechanisms, epidemiology, and consequences for treatment. Am. J. Med. 125:S3–S13. 10.1016/j.amjmed.2011.11.001 [DOI] [PubMed] [Google Scholar]
- 47.Toyoshima Y, Okubo S, Toda M. 1994. Effect of catechin on the ultrastructure of Trichophyton mentagrophytes. Kansenshogaku Zasshi 68:295–303 (In Japanese) [DOI] [PubMed] [Google Scholar]
- 48.Simon HU, Haj-Yehia A, Levi-Schaffer F. 2000. Role of reactive oxygen species (ROS) in apoptosis induction. Apoptosis 5:415–418. 10.1023/A:1009616228304 [DOI] [PubMed] [Google Scholar]
- 49.Cho J, Lee DG. 2011. The antimicrobial peptide arenicin-1 promotes generation of reactive oxygen species and induction of apoptosis. Biochim. Biophys. Acta 1810:1246–1250. 10.1016/j.bbagen.2011.08.011 [DOI] [PubMed] [Google Scholar]
- 50.Hwang B, Hwang JS, Lee J, Kim JK, Kim SR, Kim Y, Lee DG. 2011. Induction of yeast apoptosis by an antimicrobial peptide, papiliocin. Biochem. Biophys. Res. Commun. 408:89–93. 10.1016/j.bbrc.2011.03.125 [DOI] [PubMed] [Google Scholar]
- 51.Lee MH, Han DW, Hyon SH, Park JC. 2011. Apoptosis of human fibrosarcoma HT-1080 cells by epigallocatechin-3-O-gallate via induction of p53 and caspases as well as suppression of Bcl-2 and phosphorylated nuclear factor-κB. Apoptosis 16:75–85. 10.1007/s10495-010-0548-y [DOI] [PubMed] [Google Scholar]
- 52.Yin ST, Tang ML, Deng HM, Xing TR. 2009. Epigallocatechin-3-gallate induced primary cultures of rat hippocampal neurons death linked to calcium overload and oxidative stress. Naunyn Schmiedebergs Arch. Pharmacol. 379:551–564. 10.1007/s00210-009-0401-4 [DOI] [PubMed] [Google Scholar]
- 53.Suh KS, Chon S, Oh S, Kim SW, Kim JW, Kim YS, Woo JT. 2010. Prooxidative effects of green tea polyphenol (−)-epigallocatechin-3-gallate on the HIT-T15 pancreatic beta cell line. Cell Biol. Toxicol. 26:189–199. 10.1007/s10565-009-9137-7 [DOI] [PubMed] [Google Scholar]
- 54.Barroso G, Taylor S, Morshedi M, Manzur F, Gaviño F, Oehninger S. 2006. Mitochondrial membrane potential integrity and plasma membrane translocation of phosphatidylserine as early apoptotic markers: a comparison of two different sperm subpopulations. Fertil. Steril. 85:149–154. 10.1016/j.fertnstert.2005.06.046 [DOI] [PubMed] [Google Scholar]
- 55.Hwang I, Lee J, Jin HG, Woo ER, Lee DG. 2012. Amentoflavone stimulates mitochondrial dysfunction and induces apoptotic cell death in Candida albicans. Mycopathologia 173:207–218. 10.1007/s11046-011-9503-x [DOI] [PubMed] [Google Scholar]
- 56.Heiskanen KM, Bhat MB, Wang HW, Ma J, Nieminen AL. 1999. Mitochondrial depolarization accompanies cytochrome c release during apoptosis in PC6 cells. J. Biol. Chem. 274:5654–5658. 10.1074/jbc.274.9.5654 [DOI] [PubMed] [Google Scholar]
- 57.Van-Dijk C, Driessen AJ, Recourt K. 2000. The uncoupling efficiency and affinity of flavonoids for vesicles. Biochem. Pharmacol. 60:1593–1600. 10.1016/S0006-2952(00)00488-3 [DOI] [PubMed] [Google Scholar]
- 58.Alvesalo J, Vuorela H, Tammela P, Leinonen M, Saikku P, Vuorela P. 2006. Inhibitory effect of dietary phenolic compounds on Chlamydia pneumoniae in cell cultures. Biochem. Pharmacol. 71:735–741. 10.1016/j.bcp.2005.12.006 [DOI] [PubMed] [Google Scholar]
- 59.Ribeiro GF, Corte-Real M, Johansson B. 2006. Characterization of DNA damage in yeast apoptosis induced by hydrogen peroxide, acetic acid, and hyperosmotic shock. Mol. Biol. Cell 17:4584–4591. 10.1091/mbc.E06-05-0475 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Salvador VAG. 2009. Evaluation of apoptosis and necrosis in Saccharomyces cerevisiae during wine fermentations. Dissertation. University of Lisbon Técnca, Lisbon, Portugal [Google Scholar]
- 61.Babich H, Krupka ME, Nissim HA, Zuckerbraun HL. 2005. Differential in vitro cytotoxicity of (−)-epicatechin gallate (ECG) to cancer and normal cells from the human oral cavity. Toxicol. In Vitro 19:231–242. 10.1016/j.tiv.2004.09.001 [DOI] [PubMed] [Google Scholar]