

Abstract
Adenosine receptors (ARs) comprise a group of G protein-coupled receptors (GPCR) which mediate the physiological actions of adenosine. To date, four AR subtypes have been cloned and identified in different tissues. These receptors have distinct localization, signal transduction pathways and different means of regulation upon exposure to agonists. This review will describe the biochemical characteristics and signaling cascade associated with each receptor and provide insight into how these receptors are regulated in response to agonists. A key property of some of these receptors is their ability to serve as sensors of cellular oxidative stress, which is transmitted by transcription factors, such as nuclear factor (NF)-κB, to regulate the expression of ARs. Recent observations of oligomerization of these receptors into homo- and heterodimers will be discussed. In addition, the importance of these receptors in the regulation of normal and pathological processes such as sleep, the development of cancers and in protection against hearing loss will be examined.
Keywords: adenosine, adenosine receptors, desensitization, nuclear factor-κB, sleep, hearing loss, cancer
1. Introduction
Adenosine is produced primarily from the metabolism of adenosine triphosphate (ATP) and exerts pleiotropic functions throughout the body. In the central nervous system (CNS), adenosine plays important functions such as modulation of neurotransmitter release [1], synaptic plasticity [2] and neuroprotection in ischemic, hypoxic and oxidative stress events [3–5]. In addition, adenosine plays different roles in a large variety of tissues. In the cardiovascular system, adenosine produces either vasoconstriction or vasodilation of veins and arteries [6]. Adenosine regulates T cell proliferation and cytokine production [7]. The nucleoside also inhibits lipolysis and stimulates bronchoconstriction [8,9].
Adenosine is produced both intracellularly and extracellularly. Intracellular adenosine is produced from its immediate precursor, 5′-adenosine monophosphate (5′-AMP), by the action of the enzyme 5′-nucleotidase. Adenosine can then follow several metabolic/synthetic pathways. It can be metabolized to inosine and hypoxanthine by adenosine deaminase and to uric acid by xanthine oxidase. Adenosine can also be transported out of the cell to the extracellular space by specific bi-directional nucleoside transporters [10]. Inhibitors of these transporters, such as dipyridamole, increase the extracellular concentrations of adenosine and are useful clinically to treat certain cardiovascular complications. Adenosine can also be converted back to 5′-AMP by the action of the enzyme adenosine kinase and subsequently to ADP and ATP. The extracellular 5′-AMP is produced by degradation of ATP (by ecto-nucleotidase) and cyclic AMP (by ecto-cyclic AMP phosphodiesterase) [10,11].
Unlike vesicular release of neurotransmitters upon electrical impulse, the release of adenosine is mediated mainly by the action of its transporters. Adenosine transporters are classified into equilibrative nucleoside transporters (ENTs) and concentrative nucleoside transporters (CNTs) [12]. ENTs are passive bidirectional transporters which transport adenosine across the plasma membrane based on its concentration gradients. The human genome encodes four ENT isoforms (ENT1-4). Among them, the best studied are ENT1 and ENT2 which are broadly classified depending on their sensitivity for inhibition by nitrobenzylthioinosine (NBMPR) [13]. These transporters are widely expressed in the CNS, with ENT1 showing the highest expression [14]. The CNTs are active Na+-dependent transporters that transport adenosine against its concentration gradient [5,15]. CNTs have been described in brain macrophages, thymocytes, liver, lung, choroid plexus, kidneys and intestines of animals [16].
Adenosine initiates its biological effects via four receptor subtypes, namely the A1, A2A, A2B and A3ARs. The A1 and A2AAR possess high affinity for adenosine while the A2B and A3AR show relatively lower affinity for adenosine receptors [5]. These receptors belong to the superfamily of G protein-coupled receptors (GPCR). Members of this family have a very similar molecular architecture. They display a seven transmembrane α-helical structure, with an extracellular amino-terminus and an intracellular carboxy-terminus. The N-terminal domain has N-glycosylation sites which influences trafficking of the receptor to the plasma membrane [17,18]. The carboxy-terminus contains serine and threonine residues which serve as phosphorylation sites for protein kinases and enable receptor desensitization. Furthermore, the carboxy-terminus and the third intracellular loop enable coupling of the ARs to G proteins [17,18].
ARs have traditionally been classified based on their differential coupling to adenylyl cyclase to regulate cyclic AMP levels. The A1 and A3ARs are coupled to Gi/o proteins, while A2AAR and A2BAR are coupled to Gs/olf proteins [5]. Therefore, activation of the A2A and A2BARs increase cyclic AMP production, resulting in activation of protein kinase A (PKA) and phosphorylation of the cyclic AMP response element binding protein (CREB). In contrast, activation of the A1 and A3AR inhibits cyclic AMP production and decreases PKA activity and CREB phosphorylation [3,5,12]. In some cases, the A1AR increases phospholipase C (PLC) activity through a pertussis toxin-sensitive G protein. The A1AR can directly couple to and inhibit cardiac K+ channels and types Q, N and P voltage sensitive Ca2+ channels. In turn, the A3AR can regulate the activity of PLC via a pertussis toxin-sensitive G protein in rat basophilic leukemia cells [19,20] or by direct coupling to Gq protein [5,21]. Activation of the A1AR can also increase mitogen-activated protein kinase (MAPK) pathway in cells of the Chinese hamster ovary (CHO) [22,23] and COS-7 fibroblast-like cells by βγ subunit of Gi/o protein [24]. Activation of the A2AAR can also promote activation of protein kinase C (PKC) in a cyclic AMP-dependent and independent mechanisms [5,25]. In contrast, activation of the A2BAR can stimulate PKC activity by direct coupling to Gq proteins [5].
In the CNS, A1ARs are widely distributed on neurons in the cortex, hippocampus and cerebellum [5]. These receptors are also present on astrocytes [26], oligodendrocytes [27] and microglia [28]. In neurons, A1AR are highly localized to synaptic regions, where they modulate the release of neurotransmitters, such as glutamate, acetylcholine, serotonin and GABA [3]. A2AAR have a more restrictive localization in the striatum and olfactory bulb [5]. These receptors are present in neurons, microglia and oligodendrocytes and possibly astrocytes [29,30]. The presence of A2AAR has been described in dendritic spines and postsynaptic regions of the basal ganglia [31]. These receptors are highly localized to presynaptic regions (in hippocampus), where they modulate the release of neurotransmitters such as glutamate, acetylcholine, GABA and noradrenaline [32–35]. A2BAR are expressed at low levels on neuronal and glial cells, such as microglia and astrocytes [36,37]. Low levels of the A3AR are detectable in the hippocampus, cortex, cerebellum and striatum [5], with cellular localization in neurons, astrocytes and microgila [38–40].
2. Desensitization of Adenosine Receptors
Continued or repeated exposures to an agonist usually result in the receptor-mediated response to plateau and then diminish despite the continual presence of agonist. In the field of receptor kinetics, this process is called desensitization [41]. Desensitization of GPCRs is a complex phenomenon involving multiple and temporally distinct components. The mechanisms underlying rapid desensitization often involve receptor phosphorylation by a family of G protein-coupled receptor kinases (GRKs) resulting in their preferential binding to arrestins molecules. This promotes desensitization by uncoupling the receptor from G-protein, leading to impaired receptor function [42]. Following desensitization, many GPCRs internalize by an arrestin-dependent process via clathrin-coated pits which leads to the eventual intracellular dephosphorylation of the receptor, and its re-insertion into the cell membrane to produce a resensitized state [43]. More prolonged agonist activation generally leads to the transfer of internalized receptor to a lysosomal compartment with subsequent down-regulation [44]. The process of desensitization, internalization, resensitization, or down-regulation forms an important part in receptor regulation which eventually controls receptor-mediated signaling pathways.
It has been shown that activation of all four AR subtypes eventually leads to their desensitization by different mechanisms. Most of these observations are made following administration of non-hydrolyzable agonists to cell cultures and animals. However, it is yet unclear how significant a role receptor desensitization plays in regulation of ARs by adenosine in vivo under normal conditions. This process should become more important with the use of selective AR subtype agonists for the treatment of diseases. Studies have shown that upon addition of agonists, the A1AR is phosphorylated and internalized slowly, with a half-life of several hours. In contrast, A2AAR and A2BAR demonstrate a more rapid rate of down-regulation, usually lasting about an hour. The down-regulation and desensitization of the A3AR occurs within minutes [45]. Thus, it is important to understand their regulation in order to design drugs that can exploit or avoid the receptor-mediated signaling to treat diseases.
2.1. A1AR
Several studies have been carried out to understand the molecular mechanisms underlying A1AR desensitization. In an early study, Parson and Stiles [46] showed that the A1AR in rat adipocytes desensitized upon chronic administration of A1AR agonist, R-phenylisopropyladenosine (R-PIA), over a period of six days by Alzet minipumps (Alza Corporation, Vacaville, CA, USA). A decrease in A1AR levels and reduced inhibition of isoproterenol-stimulated adenylyl cyclase activity was detected following R-PIA treatment. These changes were associated with decreased levels of pertussis-sensitive G proteins, but an increase in (cholera toxin-labeled) Gs proteins in the plasma membranes. Furthermore, adipocyte membranes obtained from R-PIA-treated rats showed enhance isoproterenol and forskolin-stimulated adenylyl cyclase. These studies suggest the response of adipocytes to chronic activation of the A1AR (i.e., loss of inhibition of adenylyl cyclase) is to increase the stimulatory response (i.e., increase in adenylyl cyclase activity). Alternatively, desensitization of the A1AR could relieve a tonic inhibitory tone which is reflected by enhanced agonist stimulation of adenylyl cyclase. A subsequent study performed in cultured adipocytes confirmed the in vivo findings and further demonstrated that desensitization of the A1AR was associated with desensitization of insulin-dependent glucose transport [47]. This suggests a common link between desensitization of the A1AR and insulin receptor. A later study by Longabaugh et al. [48] confirmed the previous findings that desensitization of the A1AR to R-PIA was linked to reductions in A1AR and Gi proteins and an increase in Gsα proteins, but showed that the changes in Giα proteins were not linked to alternations in their mRNA levels.
Studies in a clonal cell line, ductus deferens smooth muscle (DDT1-MF2 cells), showed differential rates of desensitization of the A1AR and A2AAR, which were not associated with changes in their coupling to G proteins. Increased phosphorylation of the A1AR was observed following exposure of these cells to agonists [49]. However, the mechanism(s) underlying phosphorylation was not determined. This study demonstrates that homologous desensitization of the A1AR was associated with phosphorylation and uncoupling of the receptor from its Gi protein. Similarly, Nie et al. [50] demonstrated rapid translocation of GRK from cytosol to the plasma membrane and subsequent phosphorylation of the A1AR within one hour of R-PIA treatment to DDT1-MF2 cells. Furthermore, purified preparations of the A1AR that were phosphorylated with purified recombinant GRK-2 displayed enhanced affinity for arrestin over Gi/Go G-proteins. In contrast to these findings, short-term exposure of CHO cells, stably expressing human A1AR, to R-PIA resulted in neither desensitization nor phosphorylation by GRK [51]. The reason for these different findings is not clear at present, but could reflect differences between hamster and human A1AR used in these studies. An interesting finding in DDT1-MF2 cells was that ectoenzyme adenosine deaminase, was shown to play an important role in agonist-induced A1AR regulation by enhancing desensitization and internalization in DDT1-MF2 cells after chronic exposure to R-PIA. It was demonstrated that upon exposure to the agonist, adenosine deaminase and A1AR colocalize and internalize by the same endocytotic pathway [52].
In primary cultures of cerebellar granule cells, chronic treatment with A1AR agonist and antagonist reciprocally regulates A1AR [53]. Exposure to A1AR agonist, N6-cyclopentyladenosine, resulted in time- and concentration-dependent reduction in the density of the A1AR and G-protein coupling corresponding to impaired agonist-induced adenylyl cyclase inhibition.
Coelho et al. [54] reported that hypoxia decreases the density of A1AR in rat hippocampal slices. This desensitization could be mimicked by 2-chloroadenosine (CADO), and was prevented by adding the A1AR antagonist, 1,3-dipropyl-8-cyclopentylxanthine (DPCPX). These results suggest that hypoxia leads to an increase in extracellular adenosine levels, and a subsequent, quite rapid (<90 min) desensitization of A1AR.
Jajoo et al. [55] examined the role of β-arrestin1/extracellular signal-regulated kinase (ERK1/2) MAPK pathway in the regulation of A1AR desensitization and recovery in DDT1-MF2 cells. They reported that the exposure of A1AR agonist, R-PIA, for 24 h resulted in a decrease of A1AR membrane protein which was associated with an unexpected 11-fold increase in A1AR mRNA. This effect of R-PIA was dependent on β-arrestin1, as knockdown of β-arrestin1 by siRNA blocked R-PIA-mediated down-regulation of the A1AR. In addition, β-arrestin1 knockdown by siRNA suppressed R-PIA-dependent ERK1/2 and activator protein-1 (AP-1) activities and reduced the induction of A1AR mRNA. Interestingly, withdrawal of the agonist after a 24 h exposure resulted in rapid recovery of plasma membrane A1AR, which was dependent on the de novo protein synthesis and on the activity of ERK1/2 but independent of β-arrestin1 and NF-κB. These findings suggest that the β-arrestin1/ERK1/2 pathway, which contributes to the desensitization and down-regulation of A1AR membrane protein, is also able to prime the transcriptional machinery for rapid synthesis of the A1AR upon withdrawal of the agonist.
Chick embryo retina cultures of eight days treated for 48 h with CGS 21680 and N6-(2(3,5-dimethoxyphenyl)-2-(2-methylphenyl)ethyl)-adenosine (DPMA), selective A2AAR agonists, showed an increased expression of A1AR. This effect was blocked by the selective A2AAR antagonist, ZM 241385, and PKA inhibitor, H89 [56]. These in vitro findings were confirmed in an in vivo study where chick embryo retinas were treated for 48 h with CGS 21680. In this study, CGS 21680 was able to increase the expression of A1AR which was blocked by selective A2AAR antagonists, SCH 58261 and ZM 241385. Interestingly, under normal conditions, endogenous adenosine produced and released into the environment of the retina appears to control A2AAR-mediated expression of A1AR, since the treatment with ZM 241385 and SCH 58261 alone was capable of reducing the expression of A1AR [57]. These findings indicate that up-regulation of A1AR by long-term activation of A2AAR depends on classical AR signaling pathway.
A study done in rat hippocampus challenged with hypoxia showed internalization and desensitization of the A1AR, but without a reduction in the overall amount of these receptors [54]. Another study in C6 glioma cells subjected to hypoxia (2, 6 and 24 h) showed down-regulation of the A1AR and up-regulation of A2AAR receptor. This effect was shown to be dependent on the release of adenosine, since treatment with adenosine deaminase was able to block this effect. Peculiarly, the effect of the A2AAR expression induced by hypoxia was inhibited by A1AR antagonist, DPCPX, but not by the A2AAR antagonist, ZM 241385, indicating that the increased expression of the A2AAR receptor induced by hypoxia depends on the A1AR [58].
2.2. A2AAR
A2AAR desensitization is mediated by multiple, temporally distinct, agonist-dependent processes. Short-term agonist exposure induced a rapid desensitization of A2AAR-stimulated adenylyl cyclase activity which was associated with diminished receptor-Gs coupling, and agonist-stimulated phosphorylation of the A2AAR receptor protein. Longer agonist treatment, however, resulted in down-regulation in total receptor number and up-regulation of α-subunits of inhibitory G-protein [59]. The structural requirements necessary for the agonist-induced desensitization of A2AAR resides mainly in its carboxy-terminus. Palmer and Stiles [60] introduced various mutations in the 95 amino acid sequence from the carboxy-terminus of A2AAR which contains ten different phosphorylation sites and identified threonine-298 to be essential in agonist-induced receptor phosphorylation and short-term desensitization. However, it is not involved in long-term desensitization of A2AAR function. These findings confirm that short-term and long-term desensitization of A2AAR could be mediated by structurally distinct regions of the receptor protein and may involve different mechanisms.
GRKs play an important role in mediating agonist-induced phosphorylation and subsequent desensitization of GPCR action. Out of the different isoforms of GRKs, few of them have been shown to regulate A2AAR desensitization. NG108-15 mouse neuroblastoma/rat glioma cells over-expressing wild-type GRK-2 showed marked reduction in the adenylyl cyclase activity after acute A2AAR stimulation and enhanced sensitivity of A2AAR to desensitization. This phenomenon was found to be dependent on the levels of GRK-2. In cells expressing very high levels of GRK-2, low agonist concentration was sufficient to induce GRK-dependent desensitization [61]. This GRK-2-mediated A2AAR desensitization is reportedly inhibited by tumor necrosis factor (TNF)-α in human monocytoid THP-1 cells representing a novel cross-talk between TNF-α receptor and A2AAR. TNF-α treatment in THP-1 cells not only reduced the translocation of GRK-2 to the plasma membrane but also decreased GRK-2 association with the plasma membrane preventing A2AAR activity and enhancing receptor function [62]. Inhibitors of receptor internalization, such as hypertonic sucrose or concanavalin A did not affect agonist-stimulation of A2AAR or agonist-induced desensitization of receptor response in NG108-15 cells. However, incubation of these cells with sucrose or concanavalin A did affect the resensitization of A2AAR response following agonist removal [63].
2.3. A2BAR
Point mutation or deletion studies of rat A2BAR stably expressed in CHO cells revealed that a serine residue (Ser329), close to the carboxy-terminus, is critical for the rapid agonist-induced desensitization and internalization of the receptor [64]. A2BAR undergoes rapid agonist-induced desensitization and internalization in a GRK-2 and arrestin-dependent manner. Expression of a dominant negative mutant of GRK-2 in NG108-15 cells [65] or antisense-induced inhibition of non-visual arrestins (arrestin-2 and -3) in human embryonic kidney (HEK-293) cells [66] efficiently reduced the rate of agonist-induced desensitization and internalization of endogenous A2BAR. Recycling of A2BAR after agonist-induced endocytosis was also affected in cells with reduced arrestin levels. Interestingly, overexpression of arrestin-2 or arrestin-3 rescued A2BAR internalization and recycling of the receptor protein. Overexpression of arrestin-3, however, showed a significantly faster rate of recycling than arrestin-2, suggesting an isoform specific role of arrestin in regulating A2BAR trafficking [67]. In another study, A2BAR was shown to internalize in an arrestin- and dynamin-sensitive manner [68].
On the other hand, second messenger-dependent kinases, such as PKA and PKC, did not seem to be involved in agonist-mediated phosphorylation and subsequent desensitization of A2BAR [69]. Human astrocytoma cells which endogenously expressed A2BAR showed that TNFα, a pro-inflammatory cytokine, markedly reduced agonist-dependent receptor phosphorylation on threonine residues and attenuated agonist-mediated A2BAR desensitization [70]. TNFα-induced inhibition of A2BAR desensitization could result in prolonged A2BAR responsiveness and may contribute to the excessive astrocytic activation that occurs in neurodegenerative diseases.
In a recent study, it was proposed that A2AAR is involved in the surface expression of A2BAR in human embryonic kidney cell line, AD-293, transfected simultaneously with mouse A2AAR and A2BAR. It was found that newly synthesized A2BAR is retained in the endoplasmic reticulum (ER) and is eventually targeted for degradation by the proteosomes. Inhibition of proteosome activity was not sufficient to enhance maturation and surface expression of the receptor. Furthermore, it was shown that co-transfection of A2AAR with A2BAR enhanced surface expression of A2BAR through F(X)6LL motif in A2AAR carboxy-terminus. These findings support the notion that A2AAR and A2BAR form heterodimer complexes for trafficking and function [71].
2.4. A3AR
Agonist occupation of A3AR results in rapid desensitization of receptor function as a result of phosphorylation of the receptor protein by members of the family of GRKs [72,73]. To identify the amino acid residue important for A3AR phosphorylation, Palmer and Stiles [74] demonstrated that a triple mutant, Thr307/Ala307, Thr318/Ala318 and Thr319/Ala319 within the carboxy-terminus, showed marked reduction in agonist-stimulated phosphorylation and desensitization of rat A3AR. Individual mutations of each residue showed that Thr318 and Thr319 are the two major sites for phosphorylation where Thr318 appeared to be necessary to observe phosphorylation at Thr319, but not vice versa. Moreover, mutation of two palmitoylation sites within the carboxy-terminus, Cys302 and Cys305 of rat A3AR which controls the GRK phosphorylation sites, displayed a significant level of basal phosphorylation even in the absence of an agonist. This suggests an important regulatory role of these palmitoylation sites in receptor desensitization.
Receptor kinetics of A3AR was studied by Trincavelli et al. [75] in human astrocytoma cells. Short-term exposure to the agonist 2-chloro-N6-(3-iodobenzyl)-N-methyl-5′-carbamoyladenosine (Cl-IBMECA) caused rapid receptor desensitization followed by internalization within 15 and 30 min, respectively. Agonist removal resulted in recycling of the A3ARs to the cell surface within 120 min. Long-term exposure (1–24 h) resulted in marked receptor down-regulation and the restoration of receptor levels associated with recovery of receptor function was also slow (24 h). MAPK was shown to regulate agonist-induced phosphorylation of A3AR where its stimulation mediated activation of ERK1/2 within 5 min of agonist exposure in CHO cells stably expressing A3AR. Treatment with PD98059, a well-characterized MAPK kinase inhibitor, showed impaired receptor phosphorylation, desensitization and internalization by inhibiting GRK-2 translocation from cytosol to the plasma membrane. These data suggests that A3AR activation is regulated by ERK1/2 in a feedback mechanism which controls GRK-2 activity and prevents receptor phosphorylation [76]. These findings could explain the dual and opposite role of A3AR in neuroprotection and neurodegenerative diseases.
3. NF-κB Regulation of A1AR and A2AAR Expression
Previous studies from our laboratories indicated that the expression of the A1AR is positively regulated by oxidative stress. This observation was made from studying the action of cisplatin, a chemotherapeutic agent, on the levels of A1AR in the chinchilla cochlea [77]. It was shown that cisplatin, via an oxidative stress pathway, induces A1AR in the cochlea which could serve as a last-ditch effort to rescue the cells in the inner ear from apoptosis. More detailed studies in DDT1-MF2 cells showed that cisplatin-induced A1AR expression resulted from activation of NF-κB by reactive oxygen species (ROS) produced by cisplatin. More importantly, an NF-κB consensus sequence was located 623 base pairs upstream of the start site of the human promoter A construct [78]. Such a mechanism of A1AR induction could help to precondition the cell or tissue to subsequent episodes of oxidative stress. For example, noise exposure, which increases ROS in the chinchilla cochlea (via NADPH oxidases), increases A1AR in the inner ear [79] while osmotic diuresis increases A1AR in the kidneys by a similar mechanism [80]. A similar mechanism could explain the induction of the A1AR and A3AR in the gut in a rabbit model of ileitis [81] and in the brain after the induction of cerebral ischemia in rats [82].
We have previously shown that activation of NF-κB is also a key regulator of A2AAR expression. Treatment of PC12 cells with nerve growth factor (NGF) significantly reduced A2AAR gene expression by an approximately three 3-fold within three days [83]. This effect was associated with a rapid activation of NF-κB via the low affinity p75 NGF receptor and was mimicked by other activators of NF-κB (e.g., ceramide, H2O2) [84]. These results are consistent with findings of NF-κB consensus sites present in the A2AAR gene promoter [85]. However, other mechanisms could be invoked to explain NGF regulation of A2AAR. These include ERK and stress-activated protein kinase/c-Jun N-terminal kinase (JNK) [86].
To evaluate the significance of NF-κB in the regulation of the A1AR expression, we examined the expression of ARs in brain tissues from mice with deletion of the gene for the p50 subunit of NF-κB. This gene knockout (KO) renders these mice immune deficient, but they are viable and able to reproduce [87]. These mice showed reduced levels of A1AR in brain cortical plasma membranes, compared to their wild type counterparts. Interestingly, the levels of the G proteins alpha subunits (Gαi3) were also significantly reduced in the p50 KO mice, but the levels of Gαi1 were unchanged. The deficit in A1AR/Gi protein expression was associated with increased neuronal apoptosis [88]. These findings suggest that NF-κB tonically regulates neuronal A1AR expression and survival.
Based on the observation that the A2AAR is negatively regulated by NF-κB, we were interested in the expression of this receptor in p50 KO mice. We observed higher expression of the A2AAR in striatal tissues from p50 KO mice, compared to wild type mice. As anticipated, less A1AR mRNA and protein was detected in the striatal tissues in p50 KO mice as compared with F2 mice [89]. These studies suggest that absence of the NF-κB p50 subunit leads to dysregulation of ARs in the striatum, as observed for the A1AR in other brain regions. Overall, these studies suggest an essential role of the NF-κB p50 subunit in the regulation of A1 and A2AAR expression in the brain.
Alterations in A2AAR in striatum could alter locomotor activity, since this receptor exhibits inhibitory actions on dopamine D2 receptor (D2R). The wild type and p50 KO mice did not show any difference in basal locomotor activity, but the p50 KO mice showed a hypersensitivity to caffeine-induced locomotor activity evaluated during the dark phase (when mice are normally active) [90]. The p50 KO mice also demonstrated increased sensitivity to intraperitoneal injections of SCH 58261, an A2AAR antagonist [90] but not to the selective A1AR antagonist, DPCPX. These data suggest that the increase in A2AAR in p50 KO mice provides larger striatal target (A2AAR) for inhibition by caffeine, resulting in behavioral hypersensitivity.
4. Adenosine Receptor Oligomers
GPCRs are usually treated as individual units responsible for a specific signaling pathway. However, it is now known that these receptors can form complex structures with each other through physical links in its protein structure. When binding occurs between the same GPCRs, it forms a homodimer. On the other hand, when the link is between different receptors, it is a heterodimer. The term heteromer is currently defined as a macromolecular complex consisting of two or more functional receptors with different biochemical characteristics of individual receptors [91]. Moreover, the activation of a member of the heteromeric complex alters the binding properties of other receptor(s) present in the complex [91].
In this regard, a variety of papers in the literature indicate that ARs are capable of forming homodimers with each other and heterodimers with other GPCRs. One of the earliest evidence of the formation of homodimers of A1ARs has been demonstrated in intact tissue in the cortex of different species by immunoblotting and coimmunoprecipitation techniques [92]. Using immunogold electron microscopy, the authors confirmed the presence of homodimers. The presence of A1AR homodimers has also been demonstrated in hippocampal pyramidal neurons and Purkinje cells of the cerebellum [93]. Recently it was shown that HEK-293T cells transfected with the cDNA for the A1AR express receptor homodimers which promoted phosphorylation of ERK. The expression of A1AR homodimers in the cortex could explain the biphasic effects of the small and high doses of caffeine on motor activity [94]. Homodimers of A2AAR receptors have been demonstrated in HeLa and HEK-293T cells co-transfected with different constructs of A2AAR receptor. Such cells produce functional receptors expressed on the cell surface and the formation of the homodimer apparently was not dependent on the activity of the receptor, since treatment with the A2AAR agonist CGS 21680 did not alter the bioluminescence resonance energy transfer (BRET) signal detected. Therefore, these data suggest that A2AAR homodimers are constitutively expressed in native tissues and cultured cell lines [95]. However, it is yet unclear how these homodimeric A1ARs differ functionally from their monomeric counterparts.
As mentioned above, ARs can form heterodimers with each other. Ciruela and coworkers [96] showed that co-transfection of the A1 and A2AAR receptor cDNAs in HEK-293T cells led to the formation of A1/A2AAR heterodimers on the cell surface. Interestingly, activation of the A2AAR by CGS 21680 in these cells reduced the affinity of the A1AR receptor to a selective radioligand. However, activation of A1AR by the A1AR agonist, R-PIA, did not produce a reciprocal change in the affinity of the A2AAR [96]. The ability of the A2AAR to reduce the affinity of the A1AR was confirmed in experiments that measured the levels of intracellular calcium. Incubation of the A1/A2AAR-transfected HEK-293T cells with R-PIA increased accumulation of intracellular calcium, which was drastically reduced when these cells were pretreated with CGS 21680 [96]. In vivo studies in rat striatum demonstrated co-localization of A1 and A2AARs in the extrasynaptic terminals of axons as well as in the presynaptic active zone of excitatory glutamatergic neurons, an indication that these receptors could be forming heterodimers. Using synaptosomes enriched in striatal glutamatergic nerve terminals, the investigators showed that the A1AR inhibited K+-evoked glutamate release was abolished by A2AAR activation [96]. In cultures of rat astrocytes, Christovão-Ferreira et al. [97] showed that A1 and A2AAR form heterodimers. In these cells, activation of the A1AR inhibited GABA uptake while activation of the A2AAR increased uptake. Surprisingly, the effect of the A1AR was blocked not only by the selective antagonist DPCPX, but also by the SCH 58261, a selective A2AAR antagonist. In turn, the effect of A2AAR on GABA uptake was blocked by SCH 58261 and DPCPX, indicating cross antagonism and/or a physical interaction between these receptors [97]. Both receptors were shown to be internalized together when exposed to A1 and A2AAR agonists separately. Interestingly, the differential responses of A1 and A2AAR agonists on GABA uptake involved activation of the Gi/0 and Gs protein, respectively [97]. Thus, the presence of A1–A2AAR heterodimers in the central nervous system could increase the complexity by which these two receptors regulate neuronal excitability. However, the advantage of A1–A2AAR heterodimers regulation of GABA uptake over the effects of the individual ARs is unclear.
Evidence from the literature suggests that an interaction exists between adenosine receptors and adrenergic receptors. For example, activation of the A1AR in myocardium attenuates β1-induced cyclic AMP formation and PKA enzymatic activity [98]. In cardiomyocytes, activation of the A1AR receptor by selective agonist CCPA translocates protein kinase C ɛ (PKCɛ) to the membrane. This effect was attenuated by isoproterenol, a non-selective β-adrenergic receptor agonist or forskolin, an activator of adenylyl cyclase [99]. These results are suggestive of an interaction between A1AR and the β-adrenergic receptors. HEK-293T cells co-transfected with A1AR, β1 and β2 adrenergic receptors show A1AR/β1AR or A1AR/β2AR heterodimers which enhance the phosphorylation of ERK induced by isoproterenol or CCPA, as compared to cells transfected with only one type of receptor. In addition, the inhibitory effect of A1AR on cyclic AMP production mediated by CCPA was abolished in cells expressing both heterodimers. Analysis of samples of human heart tissue showed the formation of these heterodimers [100]. Thus, the data suggest that A1AR and adrenergic receptors are able to form heterodimers with different properties compared to the generally accepted pharmacological and functional properties of individual receptors.
The principal source of production of adenosine into the extracellular environment occurs through metabolism of ATP [11]. Thus, the amount of ATP released into the extracellular medium directly determines the activity of ARs. However, ATP itself influences the activity of its receptors, which are subdivided into P2X, P2Y, P2U and P2Z subtypes. All members of the P2Y family of receptors are G protein-coupled. Studies indicate that the P2Y1 receptor is able to form heterodimers with the A1AR in vitro. These heterodimers were less effective in inhibiting forskolin-stimulated cyclic AMP formation than the native A1AR [101]. Imaging by BRET in HEK-293T cells confirms the formation of P2Y1/A1AR heterodimers [102]. Immunostaining of rat cortical neurons showed that both receptors co-localize in the cell body and dendritic regions. In rat cortex slices these receptors are shown to be present together in cortical, hippocampal, and cerebellar neurons, mainly in the soma and dendrites, indicating that these receptors could be form heterodimers. Moreover, immunoprecipitation analysis and electron microscopy of these specific brain regions have reported an interaction between the A1AR and P2Y1 receptors [93,103].
A2AARs are highly expressed in the striatum [5], a region that also has an abundance of cannabinoid (CB)-1 receptors [104]. CB1 receptors are coupled to Gi/o proteins and belong to a subtype of receptor for exogenous cannabinoids and endocannabinoids. The latter are represented by 2-arachidonoylglycerol (2AG) and N-arachidonoylethanolamine (anandamide), which differ from the majority of neurotransmitters because they are produced in the post-synaptic neuron and released by retrograde transport to the pre-synaptic neuron to perform its functions [104]. Immunofluorescence studies in coronal sections of rat striatum revealed a co-localization between A2AAR and CB1 receptors in striatal neurons. Subsequent immunoprecipitation experiments confirmed A2AAR-CB1 in striatal cells [105]. BRET assays in HEK-293 cells, co-transfected with A2AAR and CB1 receptors, revealed the formation of heterodimers on the cell surface [105,106]. In vitro studies in human neuroblastoma SH-SY5Y cells constitutively expressing A2AAR and CB1 receptors revealed a functional cross-talk between these receptors. When these cells were treated as selective CB1 receptor agonist, arachidonyl-2′-chloroethylamide (ACEA), the accumulation of cyclic AMP induced by forskolin was reduced. However, in the presence of the selective A2AAR antagonist, ZM 241385, the decrease in cyclic AMP by ACEA was reduced. These data suggest that effective coupling of the CB1 receptor to Gi protein requires prior or simultaneous activation of the A2AAR [105]. Moreover, CGS 21680 increase cyclic AMP production in SH-SY5Y cells, which is blocked by either ZM 241385 or ACEA. Thus, activation of A2AAR, leading to the production of cyclic AMP, required concurrent activation of the CB1 receptor [105]. Taken together, the data described above suggest that the A2AAR can heterodimerize with the CB1 receptor, which controls how each of these receptors responds to their specific agonists [107,108].
The best studied heterodimer of ARs are with dopamine receptors (DRs). Using double immunofluorescence assays, Hillion et al. [109] demonstrated a high degree of co-localization between A2AAR and D2R in the cell membranes of SH-SY5Y cells stably transfected with the human D2R and in cultures of striatal rat neurons. The existence of A2AAR-D2R heterodimeric complex was confirmed by co-immunoprecipitation assays. Interestingly, co-administration of A2AAR and D2R agonists to SH-SY5Y cells transfected with D2R cDNA resulted in co-aggregation, co-internalization and co-desensitization of the A2AAR and D2R [109]. A2AAR-D2R heterodimers were also revealed in HEK-293T cells co-transfected with the cDNAs encoding these receptors by fluorescence resonance energy transfer (FRET) and co-immunoprecipitation studies [110–112]. In HEK-293T cells co-transfected with A2AAR and D2R, Borroto-Escuela et al. [110] demonstrated that quinpirole (D2R agonist) induced internalization of the receptor heterodimer complex, which was facilitated by CGS 21680 (A2AAR agonist). Furthermore, internalization of the complex was shown to be dependent on β-arrestin2 and Akt [110]. Other data also show that the A2AAR-D2R complex can associate with the CB1 receptor [106] or the metabotropic glutamate receptor, mGluR5 [113] to form a trimeric complex. Studies have also shown formation of A1AR-D1R heterodimers. For example, co-transfection of mouse fibroblasts with A1AR and D1R cDNAs led to the formation of heterodimers. These heterodimers disappeared when these cells are pretreated with the SKF 38393, a D1R agonist. Using immunofluorescence labeling and confocal microscopy detection, co-localization or A1AR and D1R were observed in fibroblast cultures and cortical neurons [114]. Immunoprecipitation studies of tissues obtained from the rat nucleus accumbens demonstrated the presence of A1AR-D1R heterodimers in this brain region. Interestingly, the levels of heterodimers were significantly reduced in rats previously treated with cocaine, suggesting that activation of the D1R component of the heterodimer complex triggers its dissolution [115]. The presence of heterodimers between ARs and DRs in cells of the central nervous system would suggest some added functional roles of these complexes other than those mediated by activation of the individual receptors. In the striatum, one can envision that the AR-DR heterodimers could help to fine tune the intracellular signaling processes emanating from extracellular adenosine and dopamine or therapeutic agents directed to these receptors. As such, a better appreciation of AR-DR heterodimers could provide better insights to the treatment of diseases involving the striatum, such as Parkinson’s and Huntington’s disease [116–118].
5. Adenosine Receptors in the Control of Sleep
Adenosine has been shown to serve as a sleep promoting factor. The levels of this nucleoside in the basal forebrain increase during prolonged wakefulness and resolves during sleep [119]. Furthermore, individuals expressing an adenosine deaminase polymorphism, which increases adenosine, show deeper sleep and higher slow-wave activity (SWA) [120]. Infusion of adenosine in the basal forebrains increases sleep in rats [121] and cats [122]. Similarly, inhibition of adenosine intracellular uptake by inhibition of its equilibrative transporter produces electrophysiological and behavioral parameters similar to those produced by sleep deprivation [123]. Several pieces of evidence support a role of the A1AR (localized to the basal forebrain) in mediating the sleep promoting actions of adenosine. For example, systemic or cerebroventricular administration of A1AR agonists to rats led to increased sleep drive [124,125]. Administration of selective A1AR antagonist or A1AR antisense oligonucleotides (to reduce A1AR expression) increased wakefulness in rats. Interestingly, an increase in A1AR density in the basal forebrain was observed following sleep deprivation, which could contribute to the subsequent sleep rebound [124]. It is believed that the source of the excessive levels of adenosine during wakefulness derives from astrocytes and is released via soluble N-ethylmaleimide-sensitive factor attachment protein receptor (SNARE)-dependent gliotransmission [126]. Mice expressing an astrocyte-specific dominant negative SNARE protein show reduced SWA, compared with their wild-type counterparts [126]. This deficit could be produced in wild-type mice by intracerebroventricular infusion of cyclopentyltheophylline. In addition, these investigators observed that mice expressing the dominant negative SNARE protein had a lower level of memory deficit induced by sleep deprivation as compared to their wild-type littermates. This suggests a role of adenosine (produced during sleep deprivation) in mediating the memory deficits [126].
Recent studies using A1AR and A2AAR KO mice, clearly demonstrate a role of the A2AAR (not the A1AR) in mediating both the sleep suppressing and arousal actions of caffeine [127]. However, these investigators showed that the A1AR in the tuberomammillary nucleus mediates non-rapid eye movement (non-REM) sleep by inhibiting histamine release [128].
Since the A1 and A2AARs are induced by NF-κB in cell cultures [78,84], we examined the expression of these receptors in the cortex and striatal tissues of mice with a global deletion in the p50 subunit of NF-κB. As anticipated, these mice showed reduced expression of the A1AR in different regions of the brain and significant elevations in A2AAR in the striatum, suggesting that NF-κB activation controls the normal expression of these receptors. We also evaluated the sleep patterns of these mice to determine whether the modest decrease in A1AR would produce any changes. These mice exhibited more slow wave and REM sleep under normal conditions than their wild type counterparts (B6129PF2/J strain) [129]. This finding was surprising, based on the purported role of the A1AR in regulating sleep (see above) in rats. Accordingly, a lower level of A1AR expression should lead to reduced sleep duration. The p50 KO mice also demonstrate a more rapid recovery to normal sleep patterns following sleep deprivation. These data suggest dissociation between A1AR expression and normal sleep patterns in mice and in mediating the homeostatic drive following sleep deprivation.
The p50 KO mice also showed increased expression of the A2AAR in the striatum [89] and possibly in other brain regions. The increased striatal expression of A2AAR in the KO mice could account for their increased responsiveness to caffeine [89]. Since the A2AAR has been implicated in mediating the increased sleep propensity to adenosine and arousal actions of caffeine [127], it is tempting to speculate that the increase in cortical A2AAR could explain the increases in slow wave and REM sleep observed in the P50 KO mice, in addition to their rapid recovery following sleep deprivation (see Figure 1).
Figure 1.
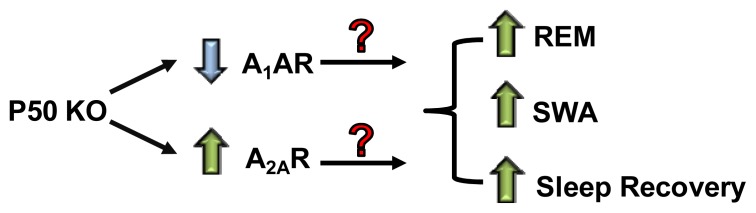
Proposed model of regulation of sleep by adenosine receptors in p50 KO mice. These mice express lower levels of A1AR in the cortex and striatum but higher expression of the A2AAR. P50 KO mice also demonstrate increased REM and SWA sleep and increased rate of sleep recovery following sleep deprivation. (?) indicates question as to which receptors to attribute the differences in sleep pattern to.
6. Adenosine Receptor and Protection against Hearing Loss
Early studies indicate that adenosine could modulate afferent neurotransmission in hair cell of the frog labyrinth system, a model system for studying hair cell [130]. These investigators showed that adenosine and adenosine uptake inhibitors reduced, while theophylline and adenosine deaminase increased, firing of hair cell afferents [130]. A subsequent study by Dulon et al. [131] demonstrated that application of adenosine to mammalian inner hair cells increased intracellular Ca2+ levels. Direct demonstration of A1AR in the rat cochlea, using several biochemical techniques, was later provided [132]. These receptors did not alter sound-evoked endocochlear potentials but activated antioxidant enzymes in the cochlea, thereby reducing the basal levels of malondialdehyde, a marker of lipid peroxidation and cell damage [77]. Activation of the A1AR contributes to the otoprotective actions of the adenosine analog, R-PIA, against cisplatin ototoxicity ([133,134] see Figure 2) and noise-induced hearing loss [135]. An interesting aspect of the A1AR in the cochlea is that it could be induced by oxidative derived from certain chemotherapeutic agents (such as cisplatin and aminoglycoside antibiotics) and noise. Induction of A1AR appears to be dependent on the activation of NF-κB by ROS which enhance transcriptional activity of the A1AR gene [78]. We propose that such feedback regulation could increase the cytoprotective activity of the A1AR in response to oxidative stress.
Figure 2.

Protection of rat cochlear outer hair cells from cisplatin-induced damage by A1AR agonist, R-PIA. Rats were pre-treated with R-PIA (10 μL of a 10 μM solution added to the round window). The remaining liquid was removed after 1 h and rats were administered cisplatin by intraperitoneal injections (16 mg/kg). Panels A–C are electron micrographs of the organ of Corti from rats treated with cisplatin and represent the hook, basal turn and the middle turn, respectively; Panels D–F shows the effect of pre-treatment with R-PIA for the respective regions; Panels G–I show the effect of R-PIA in presence of DPCPX, an A1AR antagonist, on cisplatin-induced damage. The inner hair cells are the three rows of cells with the “V” shaped stereociliary bundles. The inner hair cells are barely visible on the top of each micrograph. The data which was confirmed from different cochlear preparations support a protective role of the A1AR against cisplatin ototoxicity. (Reprinted with permission from [134], Copyright 2004 Elsevier). Scale bar = 5 μm.
The NADPH oxidase system appears to contribute significantly to cochlear ROS produced by noise trauma and cisplatin. Specifically, the NOX3 isoform of NADPH oxidase is localized primarily to the cochlea and mediates cisplatin ototoxicity [136]. As such, knockdown of NOX3 by trans-tympanic administration of NOX3 siRNA, protected against cisplatin ototoxicity [137]. ROS can increase DNA damage, and modify membrane lipids, cytosolic proteins and receptors on the cell surface [138]. Increased lipid peroxides appear to induce cochlear damage following noise exposure [139]. This explains the protective actions of antioxidants to treat hearing loss induced by noise or therapeutic agents [140]. However, the use of these agents to treat cisplatin-induced ototoxicity is limited by the potential interaction of the antioxidants and cisplatin to reduce its chemotherapeutic efficacy.
Recent studies have revealed another target of ROS, induction of inflammation in the cochlea. ROS can increase inflammatory processes in the cochlea by activating NF-κB [78,79]. NF-κB is a known mediator of inflammatory processes in the body [141]. More recently, we have shown that oxidative stress in the cochlea can contribute to the inflammatory process by activating signal transducer and activator of transcription 1 (STAT1) transcription factor [142]. In addition, STAT1 is able to couple the activation of transient receptor potential vanilloid receptor (TRPV)-1 in the cochlea to the induction of inflammation [143]. Accordingly, down-regulation of STAT1 ameliorates cisplatin-induced ototoxicity in the rats [142]. It is possible that the otoprotective actions of the A1AR against cisplatin ototoxicity involve inhibition of NF-κB and STAT1 transcription factors.
The cochlea expresses three subtypes of the ARs [144]. The A1ARs are expressed in inner hair cells, Deiters cells and spiral ganglion neurons. The A2AAR are present in the inner hair cells, Deiters cells, spiral ligament and spiral ganglion. However, the A3AR appear to have a more wide-spread expression. While it is clear that the activation of the A1AR confers otoprotection, the role of the A3AR is less clear. On the other hand, activation of the A2AAR appears to exacerbate cisplatin-induced ototoxicity [134]. As such, potentially useful AR drugs to treat ototoxicity could include agonists of the A1AR and antagonists of the A2AAR.
7. Adenosine Receptors in Cancer
The expression levels and function of A1AR in different cancer types is highly debatable. Panjehpour et al. [145] using semi-quantitative RT-PCR, reported that there was no significant difference between the expression levels of A1AR in non-neoplastic normal tissues and human breast tumor tissues. However, in another study, Mirza et al. [146] found high levels of A1AR in all breast tumor cell lines as well as in 15 of 24 human primary breast cancer tissues compared to normal mammary epithelial cells and matched normal breast tissues, respectively. In this study, knockdown of A1AR using siRNA attenuated MDA-MB-468 human breast tumor cell growth and proliferation by impairing the G1 checkpoint in the cell cycle. High expression of A1AR was also found in human colorectal adenocarcinoma [147], human leukemia Jurkat T cells [148] and human melanoma A375 cell lines [149]. Increased A1AR was also observed in the peritumoral regions of F98 glioblastoma cells [150]. The source of increased A1AR expression was found to be activated astrocytes and microglia which suppressed growth of glioblastoma upon A1AR activation. Despite its role in tumorigenesis, few studies have shown A1AR to have anti-tumor effects. In CW2 human colon cancer cells, A1AR activation increased apoptosis by activating caspases [151]. Anti-proliferative effects of A1AR were also reported in LoVo human metastatic colon cancer cells [152] and Sertoli-like TM4 cells [153].
A2AAR are found to be over-expressed in several cancer cell lines such as MCF-7 human breast cancer cells [154], Jurkat T-cell leukemia [148], A375 melanoma cells [149], U87MG human glioblastoma cells [155], HT29 and DLD-1 colon carcinoma cells [156], SH-SY5Y neuroblastoma cells [157], and non-small cell lung cancer cells [158] where it is shown to affect cell proliferation, apoptosis, angiogenesis and anti-tumor immunity. Activation of the A2AAR by CGS 21680 was shown to stimulate cell proliferation in MCF-7 breast cancer cells [154]. A2AAR also induced angiogenesis by promoting endothelial cell proliferation, migration and tube formation which is an important hallmark of cancer [159,160]. In another study, A2AAR was reported to protect the tumor by inhibiting anti-tumor T cells and A2AAR antagonist facilitated CD8+ T cell-mediated retardation of tumor growth [161]. These findings warrant the use of A2AAR antagonists to inhibit tumor growth and angiogenesis. In contrast, A2AAR activation in A375 melanoma cells reduced cell viability and cell clone formation [162], and activated caspases to induce apoptosis in Caco-2 human colon cancer cells [163].
The role of A2BAR in cancer can be broadly classified as pro-angiogenic. A2BAR activation in human retinal endothelial cells stimulated neovascularization through production of vascular endothelial growth factor (VEGF) [164]. A2BAR knockout mice, administered with Lewis lung carcinoma cells, demonstrated reduced tumor growth and longer survival times, compared to wild type control mice. It was suggested that tumor cells promote their growth by exploiting A2BAR-dependent regulation of VEGF in host immune cells [165]. In addition, knockdown of A2BAR by siRNA inhibited the release of pro-angiogenic growth factor, interleukin-8, in A375 human melanoma cells treated with chemotherapeutic agents like etoposide and doxorubicin [166]. The A2BAR promoter region contains a functional binding site for hypoxia-inducible factor (HIF), which leads to the induction of A2BAR under hypoxic conditions [167]. A2BAR is also over-expressed in colorectal carcinoma cells grown under hypoxic state, the inhibition of which significantly attenuated cell proliferation [168].
A3ARs are over-expressed in different types of cancer cells, such as PC-3MM human prostate cancer cells [169], colon and breast carcinoma tissues [170], A375 human melanoma cells [146], U87MG human glioblastoma cells [155,171], HL60 human promyelocytic leukemia cells [172], and hepatocellular carcinoma [173] compared to their normal counterparts, suggesting that these receptors could serve as potential molecular markers of these cancers. A role of the A3AR in mediating anti-tumor actions has been demonstrated in in vitro and in vivo models. For example, A3AR agonists inhibit the growth of melanoma [174], colon [175], leukemia [172] and prostate cancers [169,176] in animal models. A3AR agonist also inhibited liver metastasis in mice inoculated with human HCT-116 or murine CT-26 colon carcinoma cells [175]. The anti-tumor action of A3AR agonists could be explained by an increase in natural killer (NK) cell activity, which promotes killing of tumor cells [177]. In prostate cancer cells, Jajoo et al. [169] demonstrated that activation of the A3AR led to suppression of the high levels of ROS generated by these cells by inhibiting NADPH oxidases (Figure 3). Interestingly, activation of this receptor reduced ERK1/2 activity which normally controls the phoshorylation and activity of the p47phox subunit of this enzyme.
Figure 3.

Activation of A3AR suppresses ROS generation in AT6.1 prostate cancer cells. Cells were pretreated with the A3AR agonist, IB-MECA, the antagonist, MRS1523, or a combination of IB-MECA + MRS1523. ROS was determined by H2DCFDA fluorescence. IB-MECA significantly reduced fluorescence in these cells (A,B) which was reversed by MRS1523, indicating a role of the A3AR in this process. Similar effects were obtained with DPI and AEBSF, known inhibitors of NADPH oxidase (C,D). Experiments were replicated four times. Histograms represent the mean ± SEM. Asterisks (*) and (**) indicates statistically significant difference (p < 0.05) from vehicle and IB-MECA treatments, respectively. (Reprinted with permission from [169], Copyright 2009 Neoplasia Press, Inc.).
8. Conclusions
As described above, adenosine and ARs play a dynamic role in regulating normal cell physiology and also act as modulators in disease processes. A better understanding of the functions of these receptors, especially the newly identified receptor homomers and heteromers, could stimulate development of new therapies for the treatment of diseases.
Acknowledgments
The research described in this review, which was performed in the laboratory of the authors, was supported, in part, by NIH grants, R15 CA135494, R15DC011412, RO1-DC002396, and R01-CA166907, and from the Excellence in Academic Medicine Award from the SIU School of Medicine. Rafael Brito, who is a Ph.D. student, received a fellowship from Conselho Nacional de Desenvolvimento Cientifico e Tecnologico (CNPq) from Brazil (Processor# 2028752013-0) to work in Vickram Ramkumar’s laboratory.
Conflicts of Interest
The authors declare no conflict of interest
References
- 1.Sebastiao A.M., Ribeiro J.A. Fine-tuning neuromodulation by adenosine. Trends Pharmacol. Sci. 2000;21:341–346. doi: 10.1016/s0165-6147(00)01517-0. [DOI] [PubMed] [Google Scholar]
- 2.de Mendoncca A., Ribeiro J.A. Adenosine and synaptic plasticity. Drug Dev. Res. 2001;52:283–290. [Google Scholar]
- 3.Cunha R.A. Adenosine as a neuromodulator and as a homeostatic regulator in the nervous system: Different roles, different sources and different receptors. Neurochem. Int. 2001;38:107–125. doi: 10.1016/s0197-0186(00)00034-6. [DOI] [PubMed] [Google Scholar]
- 4.Ferreira J.M., Paes-de-Carvalho R. Long-term activation of adenosine A(2a) receptors blocks glutamate excitotoxicity in cultures of avian retinal neurons. Brain Res. 2001;900:169–176. doi: 10.1016/s0006-8993(01)02279-x. [DOI] [PubMed] [Google Scholar]
- 5.Fredholm B.B., AP I.J., Jacobson K.A., Linden J., Muller C.E. International Union of Basic and Clinical Pharmacology. LXXXI. Nomenclature and classification of adenosine receptors—An update. Pharmacol. Rev. 2011;63:1–34. doi: 10.1124/pr.110.003285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Li J., Fenton R.A., Wheeler H.B., Powell C.C., Peyton B.D., Cutler B.S., Dobson J.G., Jr. Adenosine A2a receptors increase arterial endothelial cell nitric oxide. J. Surg. Res. 1998;80:357–364. doi: 10.1006/jsre.1998.5439. [DOI] [PubMed] [Google Scholar]
- 7.Hasko G., Pacher P. A2A receptors in inflammation and injury: Lessons learned from transgenic animals. J. Leukoc. Biol. 2008;83:447–455. doi: 10.1189/jlb.0607359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Bouma M.G., Stad R.K., van den Wildenberg F.A., Buurman W.A. Differential regulatory effects of adenosine on cytokine release by activated human monocytes. J. Immunol. 1994;153:4159–4168. [PubMed] [Google Scholar]
- 9.Van der Graaf P.H., Van Schaick E.A., Visser S.A., De Greef H.J., Ijzerman A.P., Danhof M. Mechanism-based pharmacokinetic-pharmacodynamic modeling of antilipolytic effects of adenosine A(1) receptor agonists in rats: Prediction of tissue-dependent efficacy in vivo. J. Pharmacol. Exp. Ther. 1999;290:702–709. [PubMed] [Google Scholar]
- 10.Latini S., Pedata F. Adenosine in the central nervous system: Release mechanisms and extracellular concentrations. J. Neurochem. 2001;79:463–484. doi: 10.1046/j.1471-4159.2001.00607.x. [DOI] [PubMed] [Google Scholar]
- 11.Zimmermann H. Extracellular metabolism of ATP and other nucleotides. Naunyn. Schmiedebergs Arch. Pharmacol. 2000;362:299–309. doi: 10.1007/s002100000309. [DOI] [PubMed] [Google Scholar]
- 12.Paes-De-Carvalho R. Adenosine as a signaling molecule in the retina: Biochemical and developmental aspects. An. Acad. Bras. Cienc. 2002;74:437–451. doi: 10.1590/s0001-37652002000300007. [DOI] [PubMed] [Google Scholar]
- 13.Jarvis S.M., Young J.D. Nucleoside transport in rat erythrocytes: Two components with differences in sensitivity to inhibition by nitrobenzylthioinosine and p-chloromercuriphenyl sulfonate. J Membr. Biol. 1986;93:1–10. doi: 10.1007/BF01871013. [DOI] [PubMed] [Google Scholar]
- 14.Anderson C.M., Xiong W., Geiger J.D., Young J.D., Cass C.E., Baldwin S.A., Parkinson F.E. Distribution of equilibrative, nitrobenzylthioinosine-sensitive nucleoside transporters (ENT1) in brain. J. Neurochem. 1999;73:867–873. doi: 10.1046/j.1471-4159.1999.0730867.x. [DOI] [PubMed] [Google Scholar]
- 15.Ritzel M.W., Yao S.Y., Huang M.Y., Elliott J.F., Cass C.E., Young J.D. Molecular cloning and functional expression of cDNAs encoding a human Na+-nucleoside cotransporter (hCNT1) Am. J. Physiol. 1997;272:C707–C714. doi: 10.1152/ajpcell.1997.272.2.C707. [DOI] [PubMed] [Google Scholar]
- 16.Griffith D.A., Jarvis S.M. Nucleoside and nucleobase transport systems of mammalian cells. Biochim. Biophys. Acta. 1996;1286:153–181. doi: 10.1016/s0304-4157(96)00008-1. [DOI] [PubMed] [Google Scholar]
- 17.Baldwin J.M. Structure and function of receptors coupled to G proteins. Curr. Opin. Cell Biol. 1994;6:180–190. doi: 10.1016/0955-0674(94)90134-1. [DOI] [PubMed] [Google Scholar]
- 18.Schoneberg T., Schulz A., Gudermann T. The structural basis of G-protein-coupled receptor function and dysfunction in human diseases. Rev. Physiol. Biochem. Pharmacol. 2002;144:143–227. [PubMed] [Google Scholar]
- 19.Ramkumar V., Stiles G.L., Beaven M.A., Ali H. The A3 adenosine receptor is the unique adenosine receptor which facilitates release of allergic mediators in mast cells. J. Biol. Chem. 1993;268:16887–16890. [PubMed] [Google Scholar]
- 20.Ali H., Muller C.E., Daly J.W., Beaven M.A. Methylxanthines block antigen-induced responses in RBL-2H3 cells independently of adenosine receptors or cyclic AMP: Evidence for inhibition of antigen binding to IgE. J. Pharmacol. Exp. Ther. 1991;258:954–962. [PubMed] [Google Scholar]
- 21.Jacobson K.A., Gao Z.G. Adenosine receptors as therapeutic targets. Nat. Rev. Drug Discov. 2006;5:247–264. doi: 10.1038/nrd1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Dickenson J.M., Blank J.L., Hill S.J. Human adenosine A1 receptor and P2Y2-purinoceptormediated activation of the mitogen-activated protein kinase cascade in transfected CHO cells. Br. J. Pharmacol. 1998;124:1491–1499. doi: 10.1038/sj.bjp.0701977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Schulte G., Fredholm B.B. Human adenosine A(1), A(2A), A(2B), and A(3) receptors expressed in Chinese hamster ovary cells all mediate the phosphorylation of extracellular-regulated kinase 1/2. Mol. Pharmacol. 2000;58:477–482. [PubMed] [Google Scholar]
- 24.Faure M., Voyno-Yasenetskaya T.A., Bourne H.R. cAMP and beta gamma subunits of heterotrimeric G proteins stimulate the mitogen-activated protein kinase pathway in COS-7 cells. J. Biol. Chem. 1994;269:7851–7854. [PubMed] [Google Scholar]
- 25.Socodato R., Brito R., Calaza K.C., Paes-de-Carvalho R. Developmental regulation of neuronal survival by adenosine in the in vitro and in vivo avian retina depends on a shift of signaling pathways leading to CREB phosphorylation or dephosphorylation. J. Neurochem. 2011;116:227–239. doi: 10.1111/j.1471-4159.2010.07096.x. [DOI] [PubMed] [Google Scholar]
- 26.Biber K., Klotz K.N., Berger M., Gebicke-Harter P.J., van Calker D. Adenosine A1 receptor-mediated activation of phospholipase C in cultured astrocytes depends on the level of receptor expression. J. Neurosci. 1997;17:4956–4964. doi: 10.1523/JNEUROSCI.17-13-04956.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Othman T., Yan H., Rivkees S.A. Oligodendrocytes express functional A1 adenosine receptors that stimulate cellular migration. Glia. 2003;44:166–172. doi: 10.1002/glia.10281. [DOI] [PubMed] [Google Scholar]
- 28.Gebicke-Haerter P.J., Christoffel F., Timmer J., Northoff H., Berger M., Van Calker D. Both adenosine A1- and A2-receptors are required to stimulate microglial proliferation. Neurochem. Int. 1996;29:37–42. [PubMed] [Google Scholar]
- 29.Li X.X., Nomura T., Aihara H., Nishizaki T. Adenosine enhances glial glutamate efflux via A2a adenosine receptors. Life Sci. 2001;68:1343–1350. doi: 10.1016/s0024-3205(00)01036-5. [DOI] [PubMed] [Google Scholar]
- 30.Melani A., Cipriani S., Vannucchi M.G., Nosi D., Donati C., Bruni P., Giovannini M.G., Pedata F. Selective adenosine A2a receptor antagonism reduces JNK activation in oligodendrocytes after cerebral ischaemia. Brain. 2009;132:1480–1495. doi: 10.1093/brain/awp076. [DOI] [PubMed] [Google Scholar]
- 31.Hettinger B.D., Lee A., Linden J., Rosin D.L. Ultrastructural localization of adenosine A2A receptors suggests multiple cellular sites for modulation of GABAergic neurons in rat striatum. J. Comp. Neurol. 2001;431:331–346. doi: 10.1002/1096-9861(20010312)431:3<331::aid-cne1074>3.0.co;2-w. [DOI] [PubMed] [Google Scholar]
- 32.Cunha R.A., Almeida T., Ribeiro J.A. Modification by arachidonic acid of extracellular adenosine metabolism and neuromodulatory action in the rat hippocampus. J. Biol. Chem. 2000;275:37572–37581. doi: 10.1074/jbc.M003011200. [DOI] [PubMed] [Google Scholar]
- 33.Lopes L.V., Cunha R.A., Kull B., Fredholm B.B., Ribeiro J.A. Adenosine A(2A) receptor facilitation of hippocampal synaptic transmission is dependent on tonic A(1) receptor inhibition. Neuroscience. 2002;112:319–329. doi: 10.1016/s0306-4522(02)00080-5. [DOI] [PubMed] [Google Scholar]
- 34.Rebola N., Oliveira C.R., Cunha R.A. Transducing system operated by adenosine A(2A) receptors to facilitate acetylcholine release in the rat hippocampus. Eur. J. Pharmacol. 2002;454:31–38. doi: 10.1016/s0014-2999(02)02475-5. [DOI] [PubMed] [Google Scholar]
- 35.Rebola N., Rodrigues R.J., Oliveira C.R., Cunha R.A. Different roles of adenosine A1, A2A and A3 receptors in controlling kainate-induced toxicity in cortical cultured neurons. Neurochem. Int. 2005;47:317–325. doi: 10.1016/j.neuint.2005.05.009. [DOI] [PubMed] [Google Scholar]
- 36.Allaman I., Lengacher S., Magistretti P.J., Pellerin L. A2B receptor activation promotes glycogen synthesis in astrocytes through modulation of gene expression. Am. J. Physiol. Cell Physiol. 2003;284:C696–C704. doi: 10.1152/ajpcell.00202.2002. [DOI] [PubMed] [Google Scholar]
- 37.Sebastiao A.M., Ribeiro J.A. Adenosine A2 receptor-mediated excitatory actions on the nervous system. Prog. Neurobiol. 1996;48:167–189. doi: 10.1016/0301-0082(95)00035-6. [DOI] [PubMed] [Google Scholar]
- 38.Brand A., Vissiennon Z., Eschke D., Nieber K. Adenosine A(1) and A(3) receptors mediate inhibition of synaptic transmission in rat cortical neurons. Neuropharmacology. 2001;40:85–95. doi: 10.1016/s0028-3908(00)00117-9. [DOI] [PubMed] [Google Scholar]
- 39.Hammarberg C., Schulte G., Fredholm B.B. Evidence for functional adenosine A3 receptors in microglia cells. J. Neurochem. 2003;86:1051–1054. doi: 10.1046/j.1471-4159.2003.01919.x. [DOI] [PubMed] [Google Scholar]
- 40.Wittendorp M.C., Boddeke H.W., Biber K. Adenosine A3 receptor-induced CCL2 synthesis in cultured mouse astrocytes. Glia. 2004;46:410–418. doi: 10.1002/glia.20016. [DOI] [PubMed] [Google Scholar]
- 41.Krupnick J.G., Benovic J.L. The role of receptor kinases and arrestins in G protein-coupled receptor regulation. Annu. Rev. Pharmacol. Toxicol. 1998;38:289–319. doi: 10.1146/annurev.pharmtox.38.1.289. [DOI] [PubMed] [Google Scholar]
- 42.Bunemann M., Hosey M.M. G-protein coupled receptor kinases as modulators of G-protein signalling. J. Physiol. 1999;517:5–23. doi: 10.1111/j.1469-7793.1999.0005z.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Goodman O.B., Jr., Krupnick J.G., Santini F., Gurevich V.V., Penn R.B., Gagnon A.W., Keen J.H., Benovic J.L. Beta-arrestin acts as a clathrin adaptor in endocytosis of the beta2-adrenergic receptor. Nature. 1996;383:447–450. doi: 10.1038/383447a0. [DOI] [PubMed] [Google Scholar]
- 44.Tsao P., von Zastrow M. Downregulation of G protein-coupled receptors. Curr. Opin. Neurobiol. 2000;10:365–369. doi: 10.1016/s0959-4388(00)00096-9. [DOI] [PubMed] [Google Scholar]
- 45.Klaasse E.C., Ijzerman A.P., de Grip W.J., Beukers M.W. Internalization and desensitization of adenosine receptors. Purinergic Signal. 2008;4:21–37. doi: 10.1007/s11302-007-9086-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Parsons W.J., Stiles G.L. Heterologous desensitization of the inhibitory A1 adenosine receptor-adenylate cyclase system in rat adipocytes. Regulation of both Ns and Ni. J. Biol. Chem. 1987;262:841–847. [PubMed] [Google Scholar]
- 47.Green A. Adenosine receptor down-regulation and insulin resistance following prolonged incubation of adipocytes with an A1 adenosine receptor agonist. J. Biol. Chem. 1987;262:15702–15707. [PubMed] [Google Scholar]
- 48.Longabaugh J.P., Didsbury J., Spiegel A., Stiles G.L. Modification of the rat adipocyte A1 adenosine receptor-adenylate cyclase system during chronic exposure to an A1 adenosine receptor agonist: Alterations in the quantity of GS alpha and Gi alpha are not associated with changes in their mRNAs. Mol. Pharmacol. 1989;36:681–688. [PubMed] [Google Scholar]
- 49.Ramkumar V., Olah M.E., Jacobson K.A., Stiles G.L. Distinct pathways of desensitization of A1- and A2-adenosine receptors in DDT1 MF-2 cells. Mol. Pharmacol. 1991;40:639–647. [PMC free article] [PubMed] [Google Scholar]
- 50.Nie Z., Mei Y., Ramkumar V. Short term desensitization of the A1 adenosine receptors in DDT1MF-2 cells. Mol. Pharmacol. 1997;52:456–464. doi: 10.1124/mol.52.3.456. [DOI] [PubMed] [Google Scholar]
- 51.Palmer T.M., Benovic J.L., Stiles G.L. Molecular basis for subtype-specific desensitization of inhibitory adenosine receptors. Analysis of a chimeric A1–A3 adenosine receptor. J. Biol. Chem. 1996;271:15272–15278. doi: 10.1074/jbc.271.25.15272. [DOI] [PubMed] [Google Scholar]
- 52.Saura C.A., Mallol J., Canela E.I., Lluis C., Franco R. Adenosine deaminase and A1 adenosine receptors internalize together following agonist-induced receptor desensitization. J. Biol. Chem. 1998;273:17610–17617. doi: 10.1074/jbc.273.28.17610. [DOI] [PubMed] [Google Scholar]
- 53.Hettinger-Smith B.D., Leid M., Murray T.F. Chronic exposure to adenosine receptor agonists and antagonists reciprocally regulates the A1 adenosine receptor-adenylyl cyclase system in cerebellar granule cells. J. Neurochem. 1996;67:1921–1930. doi: 10.1046/j.1471-4159.1996.67051921.x. [DOI] [PubMed] [Google Scholar]
- 54.Coelho J.E., Rebola N., Fragata I., Ribeiro J.A., de Mendonca A., Cunha R.A. Hypoxia-induced desensitization and internalization of adenosine A1 receptors in the rat hippocampus. Neuroscience. 2006;138:1195–1203. doi: 10.1016/j.neuroscience.2005.12.012. [DOI] [PubMed] [Google Scholar]
- 55.Jajoo S., Mukherjea D., Kumar S., Sheth S., Kaur T., Rybak L.P., Ramkumar V. Role of beta-arrestin1/ERK MAP kinase pathway in regulating adenosine A1 receptor desensitization and recovery. Am. J. Physiol. Cell Physiol. 2010;298:C56–C65. doi: 10.1152/ajpcell.00190.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Pereira M.R., Hang V.R., Vardiero E., de Mello F.G., Paes-de-Carvalho R. Modulation of A1 adenosine receptor expression by cell aggregation and long-term activation of A2a receptors in cultures of avian retinal cells: Involvement of the cyclic AMP/PKA pathway. J. Neurochem. 2010;113:661–673. doi: 10.1111/j.1471-4159.2010.06641.x. [DOI] [PubMed] [Google Scholar]
- 57.Brito R., Pereira M.R., Paes-de-Carvalho R., da Calaza K.C. Expression of A1 adenosine receptors in the developing avian retina: In vivo modulation by A(2A) receptors and endogenous adenosine. J. Neurochem. 2012;123:239–249. doi: 10.1111/j.1471-4159.2012.07909.x. [DOI] [PubMed] [Google Scholar]
- 58.Castillo C.A., Leon D., Ruiz M.A., Albasanz J.L., Martin M. Modulation of adenosine A1 and A2A receptors in C6 glioma cells during hypoxia: Involvement of endogenous adenosine. J. Neurochem. 2008;105:2315–2329. doi: 10.1111/j.1471-4159.2008.05314.x. [DOI] [PubMed] [Google Scholar]
- 59.Palmer T.M., Gettys T.W., Jacobson K.A., Stiles G.L. Desensitization of the canine A2a adenosine receptor: delineation of multiple processes. Mol. Pharmacol. 1994;45:1082–1094. [PMC free article] [PubMed] [Google Scholar]
- 60.Palmer T.M., Stiles G.L. Identification of an A2a adenosine receptor domain specifically responsible for mediating short-term desensitization. Biochemistry. 1997;36:832–838. doi: 10.1021/bi962290v. [DOI] [PubMed] [Google Scholar]
- 61.Mundell S.J., Luty J.S., Willets J., Benovic J.L., Kelly E. Enhanced expression of G protein-coupled receptor kinase 2 selectively increases the sensitivity of A2A adenosine receptors to agonist-induced desensitization. Br. J. Pharmacol. 1998;125:347–356. doi: 10.1038/sj.bjp.0702081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Khoa N.D., Postow M., Danielsson J., Cronstein B.N. Tumor necrosis factor-alpha prevents desensitization of Galphas-coupled receptors by regulating GRK2 association with the plasma membrane. Mol. Pharmacol. 2006;69:1311–1319. doi: 10.1124/mol.105.016857. [DOI] [PubMed] [Google Scholar]
- 63.Mundell S.J., Kelly E. The effect of inhibitors of receptor internalization on the desensitization and resensitization of three Gs-coupled receptor responses. Br. J. Pharmacol. 1998;125:1594–1600. doi: 10.1038/sj.bjp.0702234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Matharu A.L., Mundell S.J., Benovic J.L., Kelly E. Rapid agonist-induced desensitization and internalization of the A(2B) adenosine receptor is mediated by a serine residue close to the COOH terminus. J. Biol. Chem. 2001;276:30199–30207. doi: 10.1074/jbc.M010650200. [DOI] [PubMed] [Google Scholar]
- 65.Mundell S.J., Benovic J.L., Kelly E. A dominant negative mutant of the G protein-coupled receptor kinase 2 selectively attenuates adenosine A2 receptor desensitization. Mol. Pharmacol. 1997;51:991–998. doi: 10.1124/mol.51.6.991. [DOI] [PubMed] [Google Scholar]
- 66.Mundell S.J., Loudon R.P., Benovic J.L. Characterization of G protein-coupled receptor regulation in antisense mRNA-expressing cells with reduced arrestin levels. Biochemistry. 1999;38:8723–8732. doi: 10.1021/bi990361v. [DOI] [PubMed] [Google Scholar]
- 67.Mundell S.J., Matharu A.L., Kelly E., Benovic J.L. Arrestin isoforms dictate differential kinetics of A2B adenosine receptor trafficking. Biochemistry. 2000;39:12828–12836. doi: 10.1021/bi0010928. [DOI] [PubMed] [Google Scholar]
- 68.Claing A., Perry S.J., Achiriloaie M., Walker J.K., Albanesi J.P., Lefkowitz R.J., Premont R.T. Multiple endocytic pathways of G protein-coupled receptors delineated by GIT1 sensitivity. Proc. Natl. Acad. Sci. USA. 2000;97:1119–1124. doi: 10.1073/pnas.97.3.1119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Peters D.M., Gies E.K., Gelb C.R., Peterfreund R.A. Agonist-induced desensitization of A2B adenosine receptors. Biochem. Pharmacol. 1998;55:873–882. doi: 10.1016/s0006-2952(97)00560-1. [DOI] [PubMed] [Google Scholar]
- 70.Trincavelli M.L., Marroni M., Tuscano D., Ceruti S., Mazzola A., Mitro N., Abbracchio M.P., Martini C. Regulation of A2B adenosine receptor functioning by tumour necrosis factor a in human astroglial cells. J. Neurochem. 2004;91:1180–1190. doi: 10.1111/j.1471-4159.2004.02793.x. [DOI] [PubMed] [Google Scholar]
- 71.Moriyama K., Sitkovsky M.V. Adenosine A2A receptor is involved in cell surface expression of A2B receptor. J. Biol. Chem. 2010;285:39271–39288. doi: 10.1074/jbc.M109.098293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Ferguson G., Watterson K.R., Palmer T.M. Subtype-specific kinetics of inhibitory adenosine receptor internalization are determined by sensitivity to phosphorylation by G protein-coupled receptor kinases. Mol. Pharmacol. 2000;57:546–552. [PubMed] [Google Scholar]
- 73.Palmer T.M., Benovic J.L., Stiles G.L. Agonist-dependent phosphorylation and desensitization of the rat A3 adenosine receptor. Evidence for a G-protein-coupled receptor kinase-mediated mechanism. J. Biol. Chem. 1995;270:29607–29613. doi: 10.1074/jbc.270.49.29607. [DOI] [PubMed] [Google Scholar]
- 74.Palmer T.M., Stiles G.L. Identification of threonine residues controlling the agonist-dependent phosphorylation and desensitization of the rat A(3) adenosine receptor. Mol. Pharmacol. 2000;57:539–545. [PubMed] [Google Scholar]
- 75.Trincavelli M.L., Tuscano D., Marroni M., Falleni A., Gremigni V., Ceruti S., Abbracchio M.P., Jacobson K.A., Cattabeni F., Martini C. A3 adenosine receptors in human astrocytoma cells: Agonist-mediated desensitization, internalization, and down-regulation. Mol. Pharmacol. 2002;62:1373–1384. doi: 10.1124/mol.62.6.1373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Trincavelli M.L., Tuscano D., Marroni M., Klotz K.N., Lucacchini A., Martini C. Involvement of mitogen protein kinase cascade in agonist-mediated human A(3) adenosine receptor regulation. Biochim. Biophys. Acta. 2002;1591:55–62. doi: 10.1016/s0167-4889(02)00248-3. [DOI] [PubMed] [Google Scholar]
- 77.Ford M.S., Maggirwar S.B., Rybak L.P., Whitworth C., Ramkumar V. Expression and function of adenosine receptors in the chinchilla cochlea. Hear. Res. 1997;105:130–140. doi: 10.1016/s0378-5955(96)00204-3. [DOI] [PubMed] [Google Scholar]
- 78.Nie Z., Mei Y., Ford M., Rybak L., Marcuzzi A., Ren H., Stiles G.L., Ramkumar V. Oxidative stress increases A1 adenosine receptor expression by activating nuclear factor kappa B. Mol. Pharmacol. 1998;53:663–669. doi: 10.1124/mol.53.4.663. [DOI] [PubMed] [Google Scholar]
- 79.Ramkumar V., Whitworth C.A., Pingle S.C., Hughes L.F., Rybak L.P. Noise induces A1 adenosine receptor expression in the chinchilla cochlea. Hear. Res. 2004;188:47–56. doi: 10.1016/S0378-5955(03)00344-7. [DOI] [PubMed] [Google Scholar]
- 80.Pingle S.C., Mishra S., Marcuzzi A., Bhat S.G., Sekino Y., Rybak L.P., Ramkumar V. Osmotic diuretics induce adenosine A1 receptor expression and protect renal proximal tubular epithelial cells against cisplatin-mediated apoptosis. J. Biol. Chem. 2004;279:43157–43167. doi: 10.1074/jbc.M405666200. [DOI] [PubMed] [Google Scholar]
- 81.Sundaram U., Hassanain H., Suntres Z., Yu J.G., Cooke H.J., Guzman J., Christofi F.L. Rabbit chronic ileitis leads to up-regulation of adenosine A1/A3 gene products, oxidative stress, and immune modulation. Biochem. Pharmacol. 2003;65:1529–1538. doi: 10.1016/s0006-2952(03)00067-4. [DOI] [PubMed] [Google Scholar]
- 82.Lai D.M., Tu Y.K., Liu I.M., Cheng J.T. Increase of adenosine A1 receptor gene expression in cerebral ischemia of Wistar rats. Neurosci. Lett. 2005;387:59–61. doi: 10.1016/j.neulet.2005.07.013. [DOI] [PubMed] [Google Scholar]
- 83.Lee N.H., Weinstock K.G., Kirkness E.F., Earle-Hughes J.A., Fuldner R.A., Marmaros S., Glodek A., Gocayne J.D., Adams M.D., Kerlavage A.R., et al. Comparative expressed-sequence-tag analysis of differential gene expression profiles in PC-12 cells before and after nerve growth factor treatment. Proc. Natl. Acad. Sci. USA. 1995;92:8303–8307. doi: 10.1073/pnas.92.18.8303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Nie Z., Mei Y., Malek R.L., Marcuzzi A., Lee N.H., Ramkumar V. A role of p75 in NGF-mediated down-regulation of the A(2A) adenosine receptors in PC12 cells. Mol. Pharmacol. 1999;56:947–954. doi: 10.1124/mol.56.5.947. [DOI] [PubMed] [Google Scholar]
- 85.Chu Y.Y., Tu K.H., Lee Y.C., Kuo Z.J., Lai H.L., Chern Y. Characterization of the rat A2a adenosine receptor gene. DNA Cell Biol. 1996;15:329–337. doi: 10.1089/dna.1996.15.329. [DOI] [PubMed] [Google Scholar]
- 86.Malek R.L., Nie Z., Ramkumar V., Lee N.H. Adenosine A(2A) receptor mRNA regulation by nerve growth factor is TrkA-, Src-, and Ras-dependent via extracellular regulated kinase and stress-activated protein kinase/c-Jun NH(2)-terminal kinase. J. Biol. Chem. 1999;274:35499–35504. doi: 10.1074/jbc.274.50.35499. [DOI] [PubMed] [Google Scholar]
- 87.Sha W.C., Liou H.C., Tuomanen E.I., Baltimore D. Targeted disruption of the p50 subunit of NF-kappa B leads to multifocal defects in immune responses. Cell. 1995;80:321–330. doi: 10.1016/0092-8674(95)90415-8. [DOI] [PubMed] [Google Scholar]
- 88.Jhaveri K.A., Reichensperger J., Toth L.A., Sekino Y., Ramkumar V. Reduced basal and lipopolysaccharide-stimulated adenosine A1 receptor expression in the brain of nuclear factor-kappaB p50−/− mice. Neuroscience. 2007;146:415–426. doi: 10.1016/j.neuroscience.2006.12.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Xie X., Jhaveri K.A., Ding M., Hughes L.F., Toth L.A., Ramkumar V. Expression of striatal adenosine and dopamine receptors in mice deficient in the p50 subunit of NF-kappaB. Life Sci. 2007;81:1031–1041. doi: 10.1016/j.lfs.2007.07.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Xie X., Mhaskar Y., Arbogast L.A., Trammell R.A., Hughes L.F., Toth L.A. Adenosine receptor antagonists and behavioral activation in NF-kappaB p50 subunit knockout mice. Life Sci. 2009;85:226–234. doi: 10.1016/j.lfs.2009.05.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Ferre S., Baler R., Bouvier M., Caron M.G., Devi L.A., Durroux T., Fuxe K., George S.R., Javitch J.A., Lohse M.J., et al. Building a new conceptual framework for receptor heteromers. Nat. Chem. Biol. 2009;5:131–134. doi: 10.1038/nchembio0309-131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Ciruela F., Casado V., Mallol J., Canela E.I., Lluis C., Franco R. Immunological identification of A1 adenosine receptors in brain cortex. J. Neurosci. Res. 1995;42:818–828. doi: 10.1002/jnr.490420610. [DOI] [PubMed] [Google Scholar]
- 93.Namba K., Suzuki T., Nakata H. Immunogold electron microscopic evidence of in situ formation of homo- and heteromeric purinergic adenosine A1 and P2Y2 receptors in rat brain. BMC Res. Notes. 2010;3:323. doi: 10.1186/1756-0500-3-323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Gracia E., Moreno E., Cortes A., Lluis C., Mallol J., McCormick P.J., Canela E.I., Casado V. Homodimerization of adenosine A(1) receptors in brain cortex explains the biphasic effects of caffeine. Neuropharmacology. 2013;71:56–69. doi: 10.1016/j.neuropharm.2013.03.005. [DOI] [PubMed] [Google Scholar]
- 95.Canals M., Burgueno J., Marcellino D., Cabello N., Canela E.I., Mallol J., Agnati L., Ferre S., Bouvier M., Fuxe K., et al. Homodimerization of adenosine A2A receptors: Qualitative and quantitative assessment by fluorescence and bioluminescence energy transfer. J. Neurochem. 2004;88:726–734. doi: 10.1046/j.1471-4159.2003.02200.x. [DOI] [PubMed] [Google Scholar]
- 96.Ciruela F., Casado V., Rodrigues R.J., Lujan R., Burgueno J., Canals M., Borycz J., Rebola N., Goldberg S.R., Mallol J., et al. Presynaptic control of striatal glutamatergic neurotransmission by adenosine A1-A2A receptor heteromers. J. Neurosci. 2006;26:2080–2087. doi: 10.1523/JNEUROSCI.3574-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Cristovao-Ferreira S., Navarro G., Brugarolas M., Perez-Capote K., Vaz S.H., Fattorini G., Conti F., Lluis C., Ribeiro J.A., McCormick P.J., et al. A1R-A2AR heteromers coupled to Gs and G i/0 proteins modulate GABA transport into astrocytes. Purinergic Signal. 2013;9:433–449. doi: 10.1007/s11302-013-9364-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Dobson J.G., Jr. Mechanism of adenosine inhibition of catecholamine-induced responses in heart. Circ. Res. 1983;52:151–160. doi: 10.1161/01.res.52.2.151. [DOI] [PubMed] [Google Scholar]
- 99.Komatsu S., Dobson J.G., Jr., Ikebe M., Shea L.G., Fenton R.A. Crosstalk between adenosine A1 and beta1-adrenergic receptors regulates translocation of PKCepsilon in isolated rat cardiomyocytes. J. Cell. Physiol. 2012;227:3201–3207. doi: 10.1002/jcp.24008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Chandrasekera P.C., Wan T.C., Gizewski E.T., Auchampach J.A., Lasley R.D. Adenosine A1 receptors heterodimerize with beta1- and beta2-adrenergic receptors creating novel receptor complexes with altered G protein coupling and signaling. Cell. Signal. 2013;25:736–742. doi: 10.1016/j.cellsig.2012.12.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Yoshioka K., Saitoh O., Nakata H. Heteromeric association creates a P2Y-like adenosine receptor. Proc. Natl. Acad. Sci. USA. 2001;98:7617–7622. doi: 10.1073/pnas.121587098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Yoshioka K., Saitoh O., Nakata H. Agonist-promoted heteromeric oligomerization between adenosine A(1) and P2Y(1) receptors in living cells. FEBS Lett. 2002;523:147–151. doi: 10.1016/s0014-5793(02)02965-4. [DOI] [PubMed] [Google Scholar]
- 103.Yoshioka K., Hosoda R., Kuroda Y., Nakata H. Hetero-oligomerization of adenosine A1 receptors with P2Y1 receptors in rat brains. FEBS Lett. 2002;531:299–303. doi: 10.1016/s0014-5793(02)03540-8. [DOI] [PubMed] [Google Scholar]
- 104.Fernandez-Ruiz J. The endocannabinoid system as a target for the treatment of motor dysfunction. Br. J. Pharmacol. 2009;156:1029–1040. doi: 10.1111/j.1476-5381.2008.00088.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Carriba P., Ortiz O., Patkar K., Justinova Z., Stroik J., Themann A., Muller C., Woods A.S., Hope B.T., Ciruela F., et al. Striatal adenosine A2A and cannabinoid CB1 receptors form functional heteromeric complexes that mediate the motor effects of cannabinoids. Neuropsychopharmacology. 2007;32:2249–2259. doi: 10.1038/sj.npp.1301375. [DOI] [PubMed] [Google Scholar]
- 106.Navarro G., Carriba P., Gandia J., Ciruela F., Casado V., Cortes A., Mallol J., Canela E.I., Lluis C., Franco R. Detection of heteromers formed by cannabinoid CB1, dopamine D2, and adenosine A2A G-protein-coupled receptors by combining bimolecular fluorescence complementation and bioluminescence energy transfer. Sci. World J. 2008;8:1088–1097. doi: 10.1100/tsw.2008.136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Martire A., Tebano M.T., Chiodi V., Ferreira S.G., Cunha R.A., Kofalvi A., Popoli P. Pre-synaptic adenosine A2A receptors control cannabinoid CB1 receptor-mediated inhibition of striatal glutamatergic neurotransmission. J. Neurochem. 2011;116:273–280. doi: 10.1111/j.1471-4159.2010.07101.x. [DOI] [PubMed] [Google Scholar]
- 108.Tebano M.T., Martire A., Chiodi V., Pepponi R., Ferrante A., Domenici M.R., Frank C., Chen J.F., Ledent C., Popoli P. Adenosine A2A receptors enable the synaptic effects of cannabinoid CB1 receptors in the rodent striatum. J. Neurochem. 2009;110:1921–1930. doi: 10.1111/j.1471-4159.2009.06282.x. [DOI] [PubMed] [Google Scholar]
- 109.Hillion J., Canals M., Torvinen M., Casado V., Scott R., Terasmaa A., Hansson A., Watson S., Olah M.E., Mallol J., et al. Coaggregation, cointernalization, and codesensitization of adenosine A2A receptors and dopamine D2 receptors. J. Biol. Chem. 2002;277:18091–18097. doi: 10.1074/jbc.M107731200. [DOI] [PubMed] [Google Scholar]
- 110.Borroto-Escuela D.O., Romero-Fernandez W., Tarakanov A.O., Ciruela F., Agnati L.F., Fuxe K. On the existence of a possible A2A-D2-beta-Arrestin2 complex: A2A agonist modulation of D2 agonist-induced beta-arrestin2 recruitment. J. Mol. Biol. 2011;406:687–699. doi: 10.1016/j.jmb.2011.01.022. [DOI] [PubMed] [Google Scholar]
- 111.Canals M., Marcellino D., Fanelli F., Ciruela F., de Benedetti P., Goldberg S.R., Neve K., Fuxe K., Agnati L.F., Woods A.S., et al. Adenosine A2A-dopamine D2 receptor-receptor heteromerization: Qualitative and quantitative assessment by fluorescence and bioluminescence energy transfer. J. Biol. Chem. 2003;278:46741–46749. doi: 10.1074/jbc.M306451200. [DOI] [PubMed] [Google Scholar]
- 112.Kamiya T., Saitoh O., Yoshioka K., Nakata H. Oligomerization of adenosine A2A and dopamine D2 receptors in living cells. Biochem. Biophys. Res. Commun. 2003;306:544–549. doi: 10.1016/s0006-291x(03)00991-4. [DOI] [PubMed] [Google Scholar]
- 113.Cabello N., Gandia J., Bertarelli D.C., Watanabe M., Lluis C., Franco R., Ferre S., Lujan R., Ciruela F. Metabotropic glutamate type 5, dopamine D2 and adenosine A2a receptors form higher-order oligomers in living cells. J. Neurochem. 2009;109:1497–1507. doi: 10.1111/j.1471-4159.2009.06078.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Gines S., Hillion J., Torvinen M., Le Crom S., Casado V., Canela E.I., Rondin S., Lew J.Y., Watson S., Zoli M., et al. Dopamine D1 and adenosine A1 receptors form functionally interacting heteromeric complexes. Proc. Natl. Acad. Sci. USA. 2000;97:8606–8611. doi: 10.1073/pnas.150241097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Toda S., Alguacil L.F., Kalivas P.W. Repeated cocaine administration changes the function and subcellular distribution of adenosine A1 receptor in the rat nucleus accumbens. J. Neurochem. 2003;87:1478–84. doi: 10.1046/j.1471-4159.2003.02121.x. [DOI] [PubMed] [Google Scholar]
- 116.Ciruela F., Ferre S., Casado V., Cortes A., Cunha R.A., Lluis C., Franco R. Heterodimeric adenosine receptors: A device to regulate neurotransmitter release. Cell. Mol. Life Sci. 2006;63:2427–2431. doi: 10.1007/s00018-006-6216-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Ferre S., Quiroz C., Orru M., Guitart X., Navarro G., Cortes A., Casado V., Canela E.I., Lluis C., Franco R. Adenosine A(2A) Receptors and A(2A) Receptor Heteromers as Key Players in Striatal Function. Front Neuroanat. 2011;5:36. doi: 10.3389/fnana.2011.00036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Franco R., Lluis C., Canela E.I., Mallol J., Agnati L., Casado V., Ciruela F., Ferre S., Fuxe K. Receptor-receptor interactions involving adenosine A1 or dopamine D1 receptors and accessory proteins. J. Neural. Transm. 2007;114:93–104. doi: 10.1007/s00702-006-0566-7. [DOI] [PubMed] [Google Scholar]
- 119.Porkka-Heiskanen T., Strecker R.E., Thakkar M., Bjorkum A.A., Greene R.W., McCarley R.W. Adenosine: A mediator of the sleep-inducing effects of prolonged wakefulness. Science. 1997;276:1265–1268. doi: 10.1126/science.276.5316.1265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Retey J.V., Adam M., Honegger E., Khatami R., Luhmann U.F., Jung H.H., Berger W., Landolt H.P. A functional genetic variation of adenosine deaminase affects the duration and intensity of deep sleep in humans. Proc. Natl. Acad. Sci. USA. 2005;102:15676–15681. doi: 10.1073/pnas.0505414102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Basheer R., Porkka-Heiskanen T., Stenberg D., McCarley R.W. Adenosine and behavioral state control: Adenosine increases c-Fos protein and AP1 binding in basal forebrain of rats. Brain Res. Mol. Brain Res. 1999;73:1–10. doi: 10.1016/s0169-328x(99)00219-3. [DOI] [PubMed] [Google Scholar]
- 122.Portas C.M., Thakkar M., Rainnie D.G., Greene R.W., McCarley R.W. Role of adenosine in behavioral state modulation: A microdialysis study in the freely moving cat. Neuroscience. 1997;79:225–235. doi: 10.1016/s0306-4522(96)00640-9. [DOI] [PubMed] [Google Scholar]
- 123.Porkka-Heiskanen T., Strecker R.E., McCarley R.W. Brain site-specificity of extracellular adenosine concentration changes during sleep deprivation and spontaneous sleep: An in vivo microdialysis study. Neuroscience. 2000;99:507–517. doi: 10.1016/s0306-4522(00)00220-7. [DOI] [PubMed] [Google Scholar]
- 124.Benington J.H., Kodali S.K., Heller H.C. Stimulation of A1 adenosine receptors mimics the electroencephalographic effects of sleep deprivation. Brain Res. 1995;692:79–85. doi: 10.1016/0006-8993(95)00590-m. [DOI] [PubMed] [Google Scholar]
- 125.Schwierin B., Borbely A.A., Tobler I. Effects of N6-cyclopentyladenosine and caffeine on sleep regulation in the rat. Eur. J. Pharmacol. 1996;300:163–171. doi: 10.1016/0014-2999(96)00021-0. [DOI] [PubMed] [Google Scholar]
- 126.Halassa M.M., Florian C., Fellin T., Munoz J.R., Lee S.Y., Abel T., Haydon P.G., Frank M.G. Astrocytic modulation of sleep homeostasis and cognitive consequences of sleep loss. Neuron. 2009;61:213–219. doi: 10.1016/j.neuron.2008.11.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Huang Z.L., Qu W.M., Eguchi N., Chen J.F., Schwarzschild M.A., Fredholm B.B., Urade Y., Hayaishi O. Adenosine A2A, but not A1, receptors mediate the arousal effect of caffeine. Nat. Neurosci. 2005;8:858–859. doi: 10.1038/nn1491. [DOI] [PubMed] [Google Scholar]
- 128.Oishi Y., Huang Z.L., Fredholm B.B., Urade Y., Hayaishi O. Adenosine in the tuberomammillary nucleus inhibits the histaminergic system via A1 receptors and promotes non-rapid eye movement sleep. Proc. Natl. Acad. Sci. USA. 2008;105:19992–19997. doi: 10.1073/pnas.0810926105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Jhaveri K.A., Ramkumar V., Trammell R.A., Toth L.A. Spontaneous, homeostatic, and inflammation-induced sleep in NF-kappaB p50 knockout mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006;291:R1516–R1526. doi: 10.1152/ajpregu.00262.2006. [DOI] [PubMed] [Google Scholar]
- 130.Bryant G.M., Barron S.E., Norris C.H., Guth P.S. Adenosine is a modulator of hair cell-afferent neurotransmission. Hear. Res. 1987;30:231–237. doi: 10.1016/0378-5955(87)90139-0. [DOI] [PubMed] [Google Scholar]
- 131.Dulon D., Mollard P., Aran J.M. Extracellular ATP elevates cytosolic Ca2+ in cochlear inner hair cells. Neuroreport. 1991;2:69–72. doi: 10.1097/00001756-199102000-00001. [DOI] [PubMed] [Google Scholar]
- 132.Ramkumar V., Ravi R., Wilson M.C., Gettys T.W., Whitworth C., Rybak L.P. Identification of A1 adenosine receptors in rat cochlea coupled to inhibition of adenylyl cyclase. Am. J. Physiol. 1994;267:C731–C737. doi: 10.1152/ajpcell.1994.267.3.C731. [DOI] [PubMed] [Google Scholar]
- 133.Ford M.S., Nie Z., Whitworth C., Rybak L.P., Ramkumar V. Up-regulation of adenosine receptors in the cochlea by cisplatin. Hear. Res. 1997;111:143–152. doi: 10.1016/s0378-5955(97)00103-2. [DOI] [PubMed] [Google Scholar]
- 134.Whitworth C.A., Ramkumar V., Jones B., Tsukasaki N., Rybak L.P. Protection against cisplatin ototoxicity by adenosine agonists. Biochem. Pharmacol. 2004;67:1801–1807. doi: 10.1016/j.bcp.2004.01.010. [DOI] [PubMed] [Google Scholar]
- 135.Wong A.C., Guo C.X., Gupta R., Housley G.D., Thorne P.R., Vlajkovic S.M. Post exposure administration of A(1) adenosine receptor agonists attenuates noise-induced hearing loss. Hear. Res. 2010;260:81–88. doi: 10.1016/j.heares.2009.12.004. [DOI] [PubMed] [Google Scholar]
- 136.Mukherjea D., Whitworth C.A., Nandish S., Dunaway G.A., Rybak L.P., Ramkumar V. Expression of the kidney injury molecule 1 in the rat cochlea and induction by cisplatin. Neuroscience. 2006;139:733–740. doi: 10.1016/j.neuroscience.2005.12.044. [DOI] [PubMed] [Google Scholar]
- 137.Mukherjea D., Jajoo S., Kaur T., Sheehan K.E., Ramkumar V., Rybak L.P. Transtympanic administration of short interfering (si)RNA for the NOX3 isoform of NADPH oxidase protects against cisplatin-induced hearing loss in the rat. Antioxid. Redox Signal. 2010;13:589–598. doi: 10.1089/ars.2010.3110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Vlajkovic S.M., Housley G.D., Thorne P.R. Adenosine and the auditory system. Curr. Neuropharmacol. 2009;7:246–256. doi: 10.2174/157015909789152155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Ohinata Y., Miller J.M., Schacht J. Protection from noise-induced lipid peroxidation and hair cell loss in the cochlea. Brain Res. 2003;966:265–273. doi: 10.1016/s0006-8993(02)04205-1. [DOI] [PubMed] [Google Scholar]
- 140.Rybak L.P., Whitworth C.A., Mukherjea D., Ramkumar V. Mechanisms of cisplatin-induced ototoxicity and prevention. Hear. Res. 2007;226:157–167. doi: 10.1016/j.heares.2006.09.015. [DOI] [PubMed] [Google Scholar]
- 141.Barnes P.J., Karin M. Nuclear factor-kappaB: A pivotal transcription factor in chronic inflammatory diseases. N. Engl. J. Med. 1997;336:1066–1071. doi: 10.1056/NEJM199704103361506. [DOI] [PubMed] [Google Scholar]
- 142.Kaur T., Mukherjea D., Sheehan K., Jajoo S., Rybak L.P., Ramkumar V. Short interfering RNA against STAT1 attenuates cisplatin-induced ototoxicity in the rat by suppressing inflammation. Cell Death Dis. 2011;2:e180. doi: 10.1038/cddis.2011.63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Mukherjea D., Jajoo S., Sheehan K., Kaur T., Sheth S., Bunch J., Perro C., Rybak L.P., Ramkumar V. NOX3 NADPH oxidase couples transient receptor potential vanilloid 1 to signal transducer and activator of transcription 1-mediated inflammation and hearing loss. Antioxid. Redox Signal. 2011;14:999–1010. doi: 10.1089/ars.2010.3497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Vlajkovic S.M., Abi S., Wang C.J., Housley G.D., Thorne P.R. Differential distribution of adenosine receptors in rat cochlea. Cell Tissue Res. 2007;328:461–471. doi: 10.1007/s00441-006-0374-2. [DOI] [PubMed] [Google Scholar]
- 145.Panjehpour M., Hemati S., Forghani M.A. Expression of A1 and A3 adenosine receptors in human breast tumors. Tumori. 2012;98:137–141. doi: 10.1177/030089161209800119. [DOI] [PubMed] [Google Scholar]
- 146.Mirza A., Basso A., Black S., Malkowski M., Kwee L., Pachter J.A., Lachowicz J.E., Wang Y., Liu S. RNA interference targeting of A1 receptor-overexpressing breast carcinoma cells leads to diminished rates of cell proliferation and induction of apoptosis. Cancer Biol. Ther. 2005;4:1355–1360. doi: 10.4161/cbt.4.12.2196. [DOI] [PubMed] [Google Scholar]
- 147.Khoo H.E., Ho C.L., Chhatwal V.J., Chan S.T., Ngoi S.S., Moochhala S.M. Differential expression of adenosine A1 receptors in colorectal cancer and related mucosa. Cancer Lett. 1996;106:17–21. doi: 10.1016/0304-3835(96)04289-9. [DOI] [PubMed] [Google Scholar]
- 148.Gessi S., Varani K., Merighi S., Morelli A., Ferrari D., Leung E., Baraldi P.G., Spalluto G., Borea P.A. Pharmacological and biochemical characterization of A3 adenosine receptors in Jurkat T cells. Br. J. Pharmacol. 2001;134:116–126. doi: 10.1038/sj.bjp.0704254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Merighi S., Varani K., Gessi S., Cattabriga E., Iannotta V., Ulouglu C., Leung E., Borea P.A. Pharmacological and biochemical characterization of adenosine receptors in the human malignant melanoma A375 cell line. Br. J. Pharmacol. 2001;134:1215–1226. doi: 10.1038/sj.bjp.0704352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Synowitz M., Glass R., Farber K., Markovic D., Kronenberg G., Herrmann K., Schnermann J., Nolte C., van Rooijen N., Kiwit J., et al. A1 adenosine receptors in microglia control glioblastoma-host interaction. Cancer Res. 2006;66:8550–8557. doi: 10.1158/0008-5472.CAN-06-0365. [DOI] [PubMed] [Google Scholar]
- 151.Saito M., Yaguchi T., Yasuda Y., Nakano T., Nishizaki T. Adenosine suppresses CW2 human colonic cancer growth by inducing apoptosis via A(1) adenosine receptors. Cancer Lett. 2010;290:211–215. doi: 10.1016/j.canlet.2009.09.011. [DOI] [PubMed] [Google Scholar]
- 152.D’Ancona S., Ragazzi E., Fassina G., Mazzo M., Gusella M., Berti T. Effect of dipyridamole, 5′-(N-ethyl)-carboxamidoadenosine and 1,3-dipropyl-8-(2-amino-4-chlorophenyl)- xanthine on LOVO cell growth and morphology. Anticancer Res. 1994;14:93–97. [PubMed] [Google Scholar]
- 153.Shaban M., Smith R.A., Stone T.W. Purine suppression of proliferation of Sertoli-like TM4 cells in culture. Cell Prolif. 1995;28:673–682. doi: 10.1111/j.1365-2184.1995.tb00053.x. [DOI] [PubMed] [Google Scholar]
- 154.Etique N., Grillier-Vuissoz I., Lecomte J., Flament S. Crosstalk between adenosine receptor (A2A isoform) and ERalpha mediates ethanol action in MCF-7 breast cancer cells. Oncol. Rep. 2009;21:977–981. doi: 10.3892/or_00000311. [DOI] [PubMed] [Google Scholar]
- 155.Gessi S., Sacchetto V., Fogli E., Merighi S., Varani K., Baraldi P.G., Tabrizi M.A., Leung E., Maclennan S., Borea P.A. Modulation of metalloproteinase-9 in U87MG glioblastoma cells by A3 adenosine receptors. Biochem. Pharmacol. 2010;79:1483–1495. doi: 10.1016/j.bcp.2010.01.009. [DOI] [PubMed] [Google Scholar]
- 156.Gessi S., Merighi S., Varani K., Cattabriga E., Benini A., Mirandola P., Leung E., MacLennan S., Feo C., Baraldi S., et al. Adenosine receptors in colon carcinoma tissues and colon tumoral cell lines: Focus on the A(3) adenosine subtype. J. Cell Physiol. 2007;211:826–836. doi: 10.1002/jcp.20994. [DOI] [PubMed] [Google Scholar]
- 157.Peterfreund R.A., Gies E.K., Fink J.S. Protein kinase C regulates adenosine A2a receptor mRNA expression in SH-SY5Y cells. Eur. J. Pharmacol. 1997;336:71–80. doi: 10.1016/s0014-2999(97)01194-1. [DOI] [PubMed] [Google Scholar]
- 158.Mediavilla-Varela M., Luddy K., Noyes D., Khalil F.K., Neuger A.M., Soliman H., Antonia S. Antagonism of adenosine A2A receptor expressed by lung adenocarcinoma tumor cells and cancer associated fibroblasts inhibits their growth. Cancer Biol. Ther. 2013;14:860–868. doi: 10.4161/cbt.25643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Ahmad A., Ahmad S., Glover L., Miller S.M., Shannon J.M., Guo X., Franklin W.A., Bridges J.P., Schaack J.B., Colgan S.P., et al. Adenosine A2A receptor is a unique angiogenic target of HIF-2alpha in pulmonary endothelial cells. Proc. Natl. Acad. Sci. USA. 2009;106:10684–10689. doi: 10.1073/pnas.0901326106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Montesinos M.C., Desai A., Chen J.F., Yee H., Schwarzschild M.A., Fink J.S., Cronstein B.N. Adenosine promotes wound healing and mediates angiogenesis in response to tissue injury via occupancy of A(2A) receptors. Am. J. Pathol. 2002;160:2009–2018. doi: 10.1016/S0002-9440(10)61151-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Ohta A., Gorelik E., Prasad S.J., Ronchese F., Lukashev D., Wong M.K., Huang X., Caldwell S., Liu K., Smith P., et al. A2A adenosine receptor protects tumors from antitumor T cells. Proc. Natl. Acad. Sci. USA. 2006;103:13132–13137. doi: 10.1073/pnas.0605251103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Merighi S., Mirandola P., Milani D., Varani K., Gessi S., Klotz K.N., Leung E., Baraldi P.G., Borea P.A. Adenosine receptors as mediators of both cell proliferation and cell death of cultured human melanoma cells. J. Invest. Dermatol. 2002;119:923–933. doi: 10.1046/j.1523-1747.2002.00111.x. [DOI] [PubMed] [Google Scholar]
- 163.Yasuda Y., Saito M., Yamamura T., Yaguchi T., Nishizaki T. Extracellular adenosine induces apoptosis in Caco-2 human colonic cancer cells by activating caspase-9/-3 via A(2a) adenosine receptors. J. Gastroenterol. 2009;44:56–65. doi: 10.1007/s00535-008-2273-7. [DOI] [PubMed] [Google Scholar]
- 164.Grant M.B., Tarnuzzer R.W., Caballero S., Ozeck M.J., Davis M.I., Spoerri P.E., Feoktistov I., Biaggioni I., Shryock J.C., Belardinelli L. Adenosine receptor activation induces vascular endothelial growth factor in human retinal endothelial cells. Circ. Res. 1999;85:699–706. doi: 10.1161/01.res.85.8.699. [DOI] [PubMed] [Google Scholar]
- 165.Ryzhov S., Novitskiy S.V., Zaynagetdinov R., Goldstein A.E., Carbone D.P., Biaggioni I., Dikov M.M., Feoktistov I. Host A(2B) adenosine receptors promote carcinoma growth. Neoplasia. 2008;10:987–995. doi: 10.1593/neo.08478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Merighi S., Simioni C., Gessi S., Varani K., Mirandola P., Tabrizi M.A., Baraldi P.G., Borea P.A. A(2B) and A(3) adenosine receptors modulate vascular endothelial growth factor and interleukin-8 expression in human melanoma cells treated with etoposide and doxorubicin. Neoplasia. 2009;11:1064–1073. doi: 10.1593/neo.09768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Kong T., Westerman K.A., Faigle M., Eltzschig H.K., Colgan S.P. HIF-dependent induction of adenosine A2B receptor in hypoxia. FASEB J. 2006;20:2242–2250. doi: 10.1096/fj.06-6419com. [DOI] [PubMed] [Google Scholar]
- 168.Ma D.F., Kondo T., Nakazawa T., Niu D.F., Mochizuki K., Kawasaki T., Yamane T., Katoh R. Hypoxia-inducible adenosine A2B receptor modulates proliferation of colon carcinoma cells. Hum. Pathol. 2010;41:1550–1557. doi: 10.1016/j.humpath.2010.04.008. [DOI] [PubMed] [Google Scholar]
- 169.Jajoo S., Mukherjea D., Watabe K., Ramkumar V. Adenosine A(3) receptor suppresses prostate cancer metastasis by inhibiting NADPH oxidase activity. Neoplasia. 2009;11:1132–1145. doi: 10.1593/neo.09744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Madi L., Ochaion A., Rath-Wolfson L., Bar-Yehuda S., Erlanger A., Ohana G., Harish A., Merimski O., Barer F., Fishman P. The A3 adenosine receptor is highly expressed in tumor versus normal cells: Potential target for tumor growth inhibition. Clin. Cancer Res. 2004;10:4472–4479. doi: 10.1158/1078-0432.CCR-03-0651. [DOI] [PubMed] [Google Scholar]
- 171.Merighi S., Benini A., Mirandola P., Gessi S., Varani K., Leung E., Maclennan S., Borea P.A. Adenosine modulates vascular endothelial growth factor expression via hypoxia-inducible factor-1 in human glioblastoma cells. Biochem. Pharmacol. 2006;72:19–31. doi: 10.1016/j.bcp.2006.03.020. [DOI] [PubMed] [Google Scholar]
- 172.Kohno Y., Sei Y., Koshiba M., Kim H.O., Jacobson K.A. Induction of apoptosis in HL-60 human promyelocytic leukemia cells by adenosine A(3) receptor agonists. Biochem. Biophys. Res. Commun. 1996;219:904–910. doi: 10.1006/bbrc.1996.0331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Bar-Yehuda S., Stemmer S.M., Madi L., Castel D., Ochaion A., Cohen S., Barer F., Zabutti A., Perez-Liz G., Del Valle L., et al. The A3 adenosine receptor agonist CF102 induces apoptosis of hepatocellular carcinoma via de-regulation of the Wnt and NF-kappaB signal transduction pathways. Int. J. Oncol. 2008;33:287–295. [PubMed] [Google Scholar]
- 174.Madi L., Bar-Yehuda S., Barer F., Ardon E., Ochaion A., Fishman P. A3 adenosine receptor activation in melanoma cells: Association between receptor fate and tumor growth inhibition. J. Biol. Chem. 2003;278:42121–42130. doi: 10.1074/jbc.M301243200. [DOI] [PubMed] [Google Scholar]
- 175.Ohana G., Bar-Yehuda S., Arich A., Madi L., Dreznick Z., Rath-Wolfson L., Silberman D., Slosman G., Fishman P. Inhibition of primary colon carcinoma growth and liver metastasis by the A3 adenosine receptor agonist CF101. Br. J. Cancer. 2003;89:1552–1558. doi: 10.1038/sj.bjc.6601315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Fishman P., Bar-Yehuda S., Ardon E., Rath-Wolfson L., Barrer F., Ochaion A., Madi L. Targeting the A3 adenosine receptor for cancer therapy: Inhibition of prostate carcinoma cell growth by A3AR agonist. Anticancer Res. 2003;23:2077–2083. [PubMed] [Google Scholar]
- 177.Harish A., Hohana G., Fishman P., Arnon O., Bar-Yehuda S. A3 adenosine receptor agonist potentiates natural killer cell activity. Int. J. Oncol. 2003;23:1245–1249. [PubMed] [Google Scholar]