

Abstract
Synechocystis sp. PCC 6803 is a genetically tractable model organism for photosynthesis research. The genome of Synechocystis sp. PCC 6803 consists of a circular chromosome and seven plasmids. The importance of small regulatory RNAs (sRNAs) as mediators of a number of cellular processes in bacteria has begun to be recognized. However, little is known regarding sRNAs in Synechocystis sp. PCC 6803. To provide a comprehensive overview of sRNAs in this model organism, the sRNAs of Synechocystis sp. PCC 6803 were analyzed using deep sequencing, and 7,951,189 reads were obtained. High quality mapping reads (6,127,890) were mapped onto the genome and assembled into 16,192 transcribed regions (clusters) based on read overlap. A total number of 5211 putative sRNAs were revealed from the genome and the 4 megaplasmids, and 27 of these molecules, including four from plasmids, were confirmed by RT-PCR. In addition, possible target genes regulated by all of the putative sRNAs identified in this study were predicted by IntaRNA and analyzed for functional categorization and biological pathways, which provided evidence that sRNAs are indeed involved in many different metabolic pathways, including basic metabolic pathways, such as glycolysis/gluconeogenesis, the citrate cycle, fatty acid metabolism and adaptations to environmentally stress-induced changes. The information from this study provides a valuable reservoir for understanding the sRNA-mediated regulation of the complex physiology and metabolic processes of cyanobacteria.
Introduction
Cyanobacteria are among the oldest life forms on the earth, appearing in the fossil record as long as 3.5 billion years ago [1]. Synechocystis sp. PCC 6803 was the first photosynthetic organism for which the genome was completely sequenced [2]. This organism’s genomic information coupled with the rich biochemistry and physiological information available for Synechocystis sp. PCC 6803 has made this strain one of the most popular genetically tractable model organisms for physiological studies of photosynthesis and systems biology and an opportune biological model for biofuel-producing microalgae [3]–[5].
Non-coding RNAs are known to be key genetic regulators in organisms from all domains of life. Bacteria, as well as eukaryotes, possess a significant number of non-coding RNAs. In bacteria, these regulatory RNAs are generally referred to as small regulatory RNAs (sRNAs) because they typically range from 50 to 200 nt in length [6]. Over the past few years, considerable progress has been made in understanding the mechanisms and regulation of bacterial sRNAs [7]. A variety of RNA-based regulatory mechanisms have been proven to control a variety of processes, including stress responses, metabolic pathways, developmental processes and pathogenesis [8]–[12]. In recent years, deep sequencing has emerged as a new and powerful experimental method for transcriptome analysis [13], and sRNAs have been detected in a wide variety of microbial genomes, including those of Escherichia coli [14], Mycobacterium tuberculosis [15], Streptococcus pneumoniae [16], Bacillus subtilis [17] and Helicobacter pylori [18],Nodularia spumigena CCY9414[19],Anabaena sp. PCC 7120 [20].
As in eukaryotes, the importance of sRNAs as mediators of a number of cellular processes has begun to be recognized in bacteria [9], [21]–[26]. However, apart from E. coli, reports about sRNAs in other bacteria are very limited, and relatively little information on sRNAs in cyanobacteria is available. Since the first cyanobacterial antisense RNA (asRNA) was found in the filamentous strain Calothrix sp. PCC7601 [27], the repertoire of cyanobacterial sRNAs has been expanding. In 1997, Watanabe et al. identified a single sRNA, called 6Sa RNA, using an elaborate biochemical protocol in the freshwater cyanobacteria Synechococcus sp. PCC6301 [28], which later turned out to be the cyanobacterial homolog of the 6S RNA [29]. Another functional RNA studied early on in cyanobacteria was the RNA subunit of RNaseP from the unicellular cyanobacterium Synechocystis sp. PCC 6803 and the heterocyst-forming strains Anabaena sp. PCC 7120 and Calothrix sp. PCC 7601 [30]–[33]. However, the first genome-wide and systematic screen for ncRNAs in cyanobacteria was based on comparative genome analyses in four marine cyanobacteria, Prochlorococcus MED4, MIT 9313, SS120 and Synechococcus WH 8102, and followed by experimental validation, which identified seven different ncRNAs, called Yfr1-7 for cYanobacterial Functional RNA [34]. Among these ncRNAs, Yfr7 is homologous to the 6S RNA [35] and well-known for its global regulatory functions in bacteria [29], [36]. In Synechocystis sp. PCC 6803, the Yfr2 sRNA family consists of three members known as Yfr2a–c [37]. In addition, 12 new ncRNAs and 24 asRNAs have been found in Prochlorococcus sp. MED4 by using high-density microarrays and genome information [38]. Comparative genome analyses have also been used for the identification of sRNAs in four model cyanobacteria, including three unicellular model cyanobacteria, Synechocystis sp. PCC 6803, Synechococcus elongatus PCC6301 and Thermosynechococcus elongatus BP1, and the toxic Microcystis aeruginosa NIES843 [37]. Seventy-eight putative RNA elements were predicted as small RNA candidates in Synechocystis sp. PCC 6803, and experimental analyses validated two new sRNAs originating from the fabF-hoxH and apcC-prmA intergenic spacers and three highly expressed sRNAs belonging to the Yfr2 family of sRNAs [37].
A computational search and a tiling microarray covering approximately 40% of the chromosome by overlapping probes have been developed specifically to detect asRNAs in Synechocystis sp. PCC 6803. Sixty intergenic sRNAs and 73 asRNAs were detected. Among these RNAs, there were 54 new sRNAs and 72 new asRNAs [39]. Recently, a differential RNA-sequencing approach examined the global transcriptional profile of Synechocystis sp. PCC 6803, resulting in the identification of 314 sRNA candidates in intergenic regions [40]. In addition to intergenic sRNAs and asRNAs, bacteria also contain regulatory elements within the 5′-leader regions of several mRNAs [14]. In this study, we report novel sRNAs identified in Synechocystis sp. PCC 6803 using a deep sequencing analysis focusing on low-molecular-weight RNAs (≤ 200 nt). By applying this approach, 5211 novel transcribed sRNA regions were identified, and 589 were found expressed from plasmid regions. The expression of 11 intergenic sRNAs, nine antisense sRNAs and seven 5′-leader regions was confirmed by RT-PCR. Possible target genes regulated by all identified putative sRNAs were predicted by IntaRNA and further analyzed for functional categorizations for biological pathways. Overall, our results provide new insights towards understanding the complex regulatory network of sRNAs in Synechocystis sp. PCC 6803.
Materials and Methods
Sample preparation
Liquid cultures of wild-type Synechocystis sp. PCC 6803 were grown at 30°C in BG11 medium under continuous illumination with white light (30 μmol photons m−2 s−1). Cells were grown to an OD730 of 0.8 and harvested by centrifugation at 3000 g, then flashed frozen in liquid nitrogen and stored at –80°C until use.
RNA preparation, library construction, and sequencing
Total RNA from Synechocystis sp. PCC 6803 cultures grown in 100 ml liquid medium was isolated using the TRIzol reagent (INVITROGEN, Carlsbad, CA, USA) according to the manufacturer’s protocol. Genomic DNA was digested by treatment with DNase (TAKARA, DALIAN, CA, CHINA), and rRNAs were removed using the Ribo-Zero rRNA Removal (Gram-negative bacteria) kit (EPICENTRE, Madison, WI, USA). Preparation of small RNA libraries was performed using the TruSeq Small RNA Sample Prep kit (ILLUMINA, San Diego, CA, USA) following the manufacturer’s instructions. Briefly, 1 μg of total RNA was ligated with adapters at the 3′ and 5′ ends without any size fractionation. Adapter-ligated RNA was reverse-transcribed with SuperScript II Reverse Transcriptase (INVITROGEN, Carlsbad, CA, USA) and then PCR-amplified for 10 cycles. Libraries (transcript sizes ≤ 200 nt) were purified using 6% TBE PAGE gels after electrophoresis. Library quality and concentration were evaluated using the Agilent 2100 Bioanalyzer DNA 1000 assay. The obtained cDNAs were sequenced using an Illumina Genome Analyzer IIx. The deep sequencing data have been submitted to the NCBI Sequence Read Archive with the accession number SRR935472.
Analysis of the deep sequencing data and prediction of candidate sRNAs
After removing adaptor sequences from the raw reads obtained using Solexa sequencing technology and the low-quality sequences, the remaining sequencing reads were mapped onto the Synechocystis sp. PCC 6803 genome and its four megaplasmids using BWA [41]. A filtering procedure was implemented in PerlScript to extract the BWA output, and those reads that mapped onto any of the intergenic regions and regions that are complementary to known annotated genes were further analyzed. A cluster is defined as a genomic region that has mapped sequence reads with the same strand orientation contiguously overlapping with each other. Candidate sRNAs were extracted from all predicted transcribed regions (clusters) based on the following two criteria. First, the depth for a given transcribed region is more than 10. Second, the novel transcribed regions that were larger than 50 nt were extracted as candidate sRNAs.
RT-PCR validation
Total RNA isolated using the TRIzol reagent (INVITROGEN, Carlsbad, CA, USA) was treated with RNase-free DNase I (FERMENTAS, Life Sciences, Germany) and two micrograms of total RNA was used as a template to synthesize first-strand cDNA using a One Step SYBR PrimeScript RT-PCR kit (TAKARA) following the manufacturer’s instructions. The sRNA primers were designed using Primer5. The detailed information for these primers is available in Online Resource Table S7 in File S1. The cycling parameters were: 95°C for 3 min followed by 35 cycles of 95°C for 15 s, 57°C for 30 s and 72°C for 30 s.
Target prediction and the analysis of candidate sRNAs
To obtain the target information, we searched the target genes of all the putative sRNAs using the target prediction program IntaRNA taking the sRNAs as query and all annotated protein-coding genes of Synechocystis sp. PCC 6803 as the subject. GO annotations with the default parameters and GO enrichment with EASE scores of 0.05 were performed with the functional annotation tool DAVID [42], [43]. The KEGG pathway annotations were performed using the KEGG (Kyoto Encyclopedia of Genes and Genomes) pathway database (http://www.genome.jp/kegg/tool/map_pathway2.html) with the default parameters.
Results
A general strategy for the high-throughput identification of sRNAs
To comprehensively identify and characterize the sRNAs of wild-type Synechocystis sp. PCC 6803 using deep sequencing, a total of 7.95 million low-molecular-weight RNA (81 nt) reads were obtained from cultures harvested at exponential phase. From the raw sequences generated by high-throughput sequencing, the 3′- and 5′-adaptor sequences were trimmed off. Low-quality sequences were identified as those sequencing reads that averaged Phred scores smaller than 20 and sequencing reads shorter than 18 nucleotides. After discarding the low-quality reads, a total of 6.12 million reads could be successfully mapped onto the Synechocystis sp. PCC 6803 genome and its four megaplasmids. As ribosomal RNAs (rRNAs) account for the vast majority of total RNAs in a bacterial cell, but on account of the removal of rRNAs, these sequences represented only 26.9% of the total reads in our sample. As shown in Figure 1, 70.5% of the reads were mapped to annotated regions corresponding to rRNAs (38.1%), tRNAs (35.3%), and mRNAs (26.6%), whereas the remaining 29.5% were mapped to non-annotated regions corresponding to intergenic regions (79.5%), the cis-antisense strands of known genes (20.3%) and misc_rna (RNA of unknown function, 0.2%).
Figure 1. Pie charts classifying the deep sequencing reads.

A, The percentages of all the deep sequencing reads mapped to the previously annotated or non-annotated regions of the Synechocystis sp. PCC 6803 genome. B, The relative proportions of the deep sequencing reads (n = 4,320,855) mapped to the annotated regions. C, The relative percentages of the deep sequencing reads (n = 1,807,035) mapped to the nonannotated regions. IGR, intergenic region; Antisense, reads located on the cis-antisense strand of a known gene; misc_rna, RNA of unknown function.
Generally, sRNAs can be generated either as primary transcripts or as being processed from longer precursor transcripts [44]. In the current study, the latter case was ignored because it was impossible to discriminate the processed sRNAs from RNA degradation products. To predict novel transcribed regions, all overlapping reads of mapped non-annotated regions were assembled, and 16,192 novel transcribed regions (clusters) were generated. It should be noted that 43 of the 89 BSRD (Bacterial Small regulatory RNA Database) registered sRNAs [45] were detected in the current study, indicating the reliability of our method.
The sequencing results were further analyzed for transcripts derived from intergenic regions and for reads complementary to known annotated genes. Transcript clusters were listed as putative candidate sRNAs only when they were represented by 10 or more reads at one cluster and were 50 nt longer. This procedure resulted in a total of 5211 candidate sRNAs, including the previously reported iron stress-repressed IsrR [9], PsbA3R [46], Yfr2a and Yfr2b [37].
Identification of sRNAs and validation by RT-PCR
Among the 5211 candidate sRNAs resulting from the deep sequencing analysis, 4622 were located on the chromosome, and 589 were located on the four megaplasmids: 110 on pSYSX, 80 on pSYSG, 200 on pSYSA and 199 on pSYSM. The putative sRNA candidates were placed into three different classes: class I compromises intergenic sRNAs, which are located within intergenic regions without overlapping adjacent genes; class II are referred to as antisense RNAs (asRNAs), located antisense to known genes; and class III comprises 5′-leader regions of mRNA transcripts, including riboswitches. To rule out the possibility of DNA contamination in total RNA sample, total RNA was digested with DNase I before reverse transcription and the results of 5S rRNA amplification showed that there was no DNA contamination after digestion in total RNA (Fig. 2)
Figure 2. PCR results of total RNA with DNase I digestion.
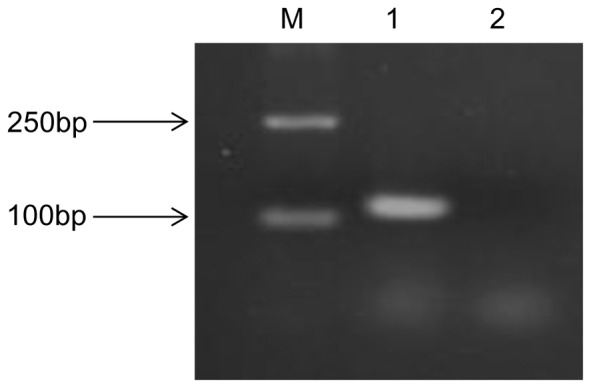
Electrophoresis of PCR amplicons of 5S rRNA using 2% agarose gels. M, Marker; 1, PCR amplicons of 5S rRNA in total RNA without digestion; 2, PCR amplicons of 5S rRNA in total RNA with DNase I digestion.
Class I, Intergenic small RNAs
An investigation that examined the global transcriptional profile of Synechocystis sp. PCC 6803 using RNA-seq resulted in the identification of 314 sRNA candidates in intergenic regions [40]. However, by using Solexa sequencing, we searched for cDNA clusters that occurred specifically within intergenic regions and discovered 423 clusters that represent possible intergenic sRNA candidates, including 335 on the chromosome and 88 on the four megaplasmids (Online Resource Table S1 in File S1). Twelve of the 56 putative top-scoring sRNAs identified by RNA-seq [40] are also presented in our results, and a total of 411 novel sRNA candidates are listed (Online Resource Table S1 in File S1). The expression of 9 out of the 13 intergenic candidate sRNAs could be detected by RT-PCR (Fig. 3), and the results are summarized in Table 1. The length of each main band on the SYPCC_igr_02, SYPCC_igr_07, SYPCC_igr_08 and SYPCC_igr_09 were almost as predicted (Fig. 3), whereas the other five candidate sRNAs tested exhibited more than one band and appeared to be different in size.
Figure 3. Validation of intergenic sRNA expression by RT-PCR.
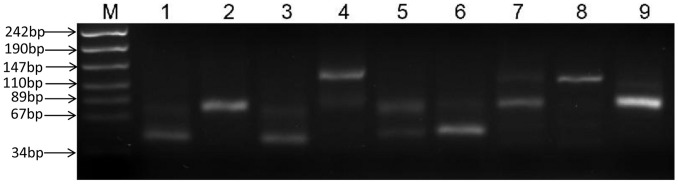
Electrophoresis of the PCR amplicons of the 9 selected intergenic sRNAs using 3% agarose gels. Lane M: Marker, Lane 1-9: SYPCC_Igr_01, SYPCC_Igr_02, SYPCC_Igr_03, SYPCC_Igr_04, SYPCC_Igr_05, SYPCC_Igr_06, SYPCC_Igr_07 SYPCC_Igr_08, and SYPCC_Igr_09.
Table 1. The 9 selected candidate intergenic sRNAs for RT-PCR validation.
| sRNA name | Location | Start (nt) | End (nt) | Strand |
| SYPCC_Igr_01 | pSYSA | 68081 | 68925 | + |
| SYPCC_Igr_02 | chromosome | 2530895 | 2530962 | + |
| SYPCC_Igr_03 | pSYSG | 15392 | 15481 | + |
| SYPCC_Igr_04 | chromosome | 496101 | 496321 | + |
| SYPCC_Igr_05 | chromsome | 1104709 | 1104929 | + |
| SYPCC_Igr_06 | chromosome | 3019919 | 3020058 | – |
| SYPCC_Igr_07 | pSYSM | 66503 | 66574 | + |
| SYPCC_Igr_08 | chromosome | 3039292 | 3039406 | + |
| SYPCC_Igr_09 | chromosome | 3305423 | 3305514 | + |
Class II, Antisense RNAs
It has been reported that, in the unicellular cyanobacterium Synechocystis sp. PCC 6803, asRNAs could account for up to 26% of all genes [39], [40] and that one gene may be predicted for several candidate sRNAs, such as slr1028 and slr0261 [40]. Overall, 4627 candidate asRNAs, including 15 of the asRNAs identified via tilling array and Northern blot analysis by Georg et al. [39], were suggested by our analysis, and the top-scoring (≥ 40 reads) putative asRNAs are provided in Online Resource Table S2 in File S1. Among these asRNAs, 116 originate from the four plasmids. It should be noted that the three previously reported known asRNAs in Synechocystis sp. PCC 6803, as1_flv4 [47], IsrR [9] and PsbA3R [46], which have been demonstrated to regulate the expression of the flv4-2 operon, iron stress-repressed mRNA isiA and the photosystem II D1 protein psbA3, respectively, were also detected in our analysis. The expression of 11 of the 14 candidate asRNAs was verified by RT-PCR (Fig. 4), and the results are summarized in Table 2.
Figure 4. Validation of asRNA expression by RT-PCR.
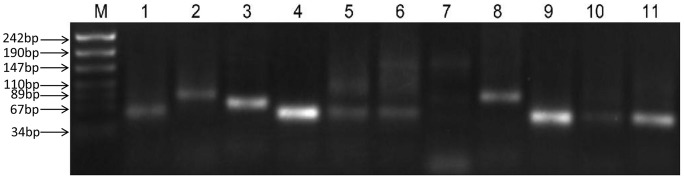
Electrophoresis of the PCR amplicons of 11 asRNAs using 3% agarose gels. Lane M: Marker, Lane 1–11: SYPCC_anti_01, SYPCC_anti_02, SYPCC_anti_03, SYPCC_anti_04, SYPCC_anti_05, SYPCC_anti_06, SYPCC_anti_07, SYPCC_anti_08, SYPCC_anti_09, SYPCC_anti_10, and SYPCC_anti_11.
Table 2. The 11 selected candidate asRNAs for RT-PCR validation.
| sRNA name | Location | Start (nt) | End (nt) | Strand |
| SYPCC_anti_01 | chromosome | 407802 | 407923 | + |
| SYPCC_anti_02 | chromosome | 1909216 | 1909443 | – |
| SYPCC_anti_03 | chromosome | 2724266 | 2724365 | – |
| SYPCC_anti_04 | chromosome | 279064 | 29161 | – |
| SYPCC_anti_05 | chromosome | 346136 | 346282 | + |
| SYPCC_anti_06 | chromosome | 2478723 | 2478824 | – |
| SYPCC_anti_07 | chromosome | 940674 | 940896 | + |
| SYPCC_anti_08 | chromosome | 1665302 | 1665520 | + |
| SYPCC_anti_09 | chromosome | 2219904 | 2220106 | – |
| SYPCC_anti_10 | chromosome | 1020080 | 1020302 | – |
| SYPCC_anti_11 | pSYSX | 91564 | 91738 | – |
Class III, 5’-leader region
In addition to intergenic sRNAs and asRNAs, bacteria contain regulatory elements within their 5′-leader regions [14], [48], [49]. Similar to sRNAs, these regulatory elements control transcription elongation, mRNA stability, and translation initiation in response to specific stimuli and metabolites [49]. In Synechocystis sp. PCC 6803, the distances between the identified and annotated genes’ transcriptional start sites and start codons for protein-coding genes range from 0 to 278 nt with a median distance of 42 nt [40]. Therefore, we define those sRNA candidates upstream of the 5′ ORF that are smaller than 300 nt as functional 5′-leader region sRNAs. A total of 161 5′-leader region candidates are shown in Online Resource Table S3 in File S1. Yfr2b, the 5′-untranslated leader of slr0199 [37], and the 6 other candidates reported by Mitschke et al. [40] were also detected in the current analysis. However, in contrast to the average length of 42 nt for the 5′-leader regions of the transcripts, the average length of predicted 5′-leader region candidates is ∼146 nt. This observation suggests that ‘long’ 5′-leader regions are likely to occur only when specialized functions are encrypted within them, and the most likely explanation for a long 5′-mRNA leader region is due to the inclusion of a signal-responsive regulatory RNA [17]. Seven out of 10 high read candidates selected for verification were detected by RT-PCR (Fig.5), and the results are summarized in Table 3.
Figure 5. Validation of 5′-leader region expression by RT-PCR.
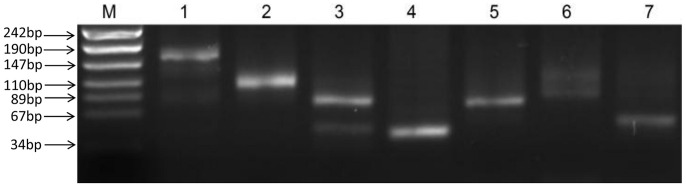
Electrophoresis of PCR amplicons of 7 5′-leader regions using 3% agarose gels. Lane M: Marker, Lane 1-7: SYPCC_Ir_01, SYPCC_Ir_02, SYPCC_Ir_03, SYPCC_Ir_04, SYPCC_Ir_05, SYPCC_Ir_06, and SYPCC_Ir_07.
Table 3. The 7 selected candidate 5′-leader regions for RT-PCR validation.
| sRNA name | Location | Start (nt) | End (nt) | Strand |
| SYPCC_Ir_01 | chromosome | 1832226 | 1832339 | + |
| SYPCC_Ir_02 | chromosome | 2730512 | 2730591 | + |
| SYPCC_Ir_03 | chromosome | 3433172 | 3433275 | + |
| SYPCC_Ir_04 | chromosome | 901005 | 901125 | + |
| SYPCC_Ir_05 | chromosome | 458917 | 459162 | + |
| SYPCC_Ir_06 | chromosome | 3168882 | 3169030 | + |
| SYPCC_Ir_07 | chromosome | 102301 | 102387 | – |
Functional categorization of candidate small RNA target genes
To better understand the role of sRNAs in different biological modules, sRNA- regulating target genes were predicted using IntaRNA [50]. Gene ontology (GO) assignment programs were utilized for the functional categorization of the predicted target genes, and the functional annotation tool DAVID [42], [43] was used to analysis the target GO annotation terms. The results are presented in Online Resource Table S4 in File S1.
Enriched GO terms were searched by GO enrichment analysis and defined by the functional significance of their target genes. The results are shown in Online Resource Table S5 in File S1. As shown in Figure 6A, the GO terms were distributed widely with regards to their respective biological processes, from transcription to RNA metabolite, to posttranslational modifications and signaling pathways. When classified as molecular functions, a total of 63% of the enriched GO terms are related to nucleoside and nucleotide binding, indicating extremely important roles for the sRNAs in the processes of replication and transcription (Fig. 6B). As for cellular components, the majority of the predicted targets are involved in different membrane systems (Fig. 6C).
Figure 6. GO analyses of predicted target sRNA genes.

GO analysis of target genes that are predicted to be involved in biological processes A, molecular functions B and their cellular components C. Classified target gene objects are depicted as percentages (in brackets) of the total number of gene objects with GO assignments.
To further evaluate the role of the identified possible target genes in biological/signaling pathways, the predicted target genes were superimposed onto the KEGG (Kyoto Encyclopedia of Genes and Genomes) pathway database (http://www.genome.jp/kegg/tool/map_pathway2.html). As shown in Online Resource Table S6 in File S1, the annotated genes were grouped into 89 known metabolic or signaling pathways, and many identified target genes are involved in pathways responsible for the control of key physiological processes. The pathways in which the target genes are predominantly enriched include “metabolic pathways” (62.8%), followed by “biosynthesis of secondary metabolites” (33%), “microbial metabolism in diverse environments” (14.3%), “two-component system” (8.8%), and “ABC transporters” (8.6%). Among those, 9 target genes (sll0427, sll0629, sll1317, slr1181, slr1655, slr1739, slr1834, smr0001 and ssr2831) were assigned to photosynthesis.
Small RNA targets involved in photosynthesis
The unicellular aquatic cyanobacterium Synechocystis sp. PCC 6803 has long been utilized as a model organism for photosynthesis studies. The regulation of photosynthesis genes in cyanobacteria has been studied in the past few years [9], [46]. According to the sRNA target data set contributed by this study, 9 genes were found to be involved in the photosynthetic pathway. As shown in Table 4, there are 4 genes involved in photosystem I and photosystem II, respectively. Among these genes, the D1 protein, encoded by the psbA gene, is believed to be the initial site of photoinhibition [51]. PsbT has been demonstrated to be involved in the efficient repair of photodamaged PSII under high light conditions [52], and there are also results indicating that PsaK and PsaL are essential for high light condition survival [53], [54]. Therefore, it is likely that the sRNAs in Synechocystis sp. PCC 6803 are involved in the regulation of the light energy distribution between PSI and PSII. Moreover, the PetA of the Cytb6f complex, which provides the electronic connection between PSII and PSI, was also identified. Taken together, sRNAs may be essential for the regulation of photosynthetic function, especially under high light conditions.
Table 4. The predicted target sRNA genes involved in photosynthesis.
| Gene ID | Description |
| sll0427 | psbO; photosystem II manganese-stabilizing polypeptide |
| sll0629 | psaK; photosystem I subunit X |
| sll1317 | petA; apocytochrome f |
| slr1181 | psbA1; photosystem II D1 protein |
| slr1655 | psaL; photosystem I reaction center protein subunit XI |
| slr1739 | photosystem II protein |
| slr1834 | psaA; photosystem I P700 chlorophyll a apoprotein A1 |
| smr0001 | psbT; photosystem II reaction center protein T |
| ssr2831 | psaE; photosystem I reaction center subunit IV |
Discussion
Although sRNAs have been increasingly recognized as major modulators of gene expression in bacteria [9], [21], [46], [55], only a limited number of studies have sought to identify the global set of expressed sRNAs in cyanobacteria [34], [37], [39], [40], [56]. Here, we describe the deep sequencing-based identification of sRNAs in the model cyanobacterium Synechocystis sp. PCC 6803. We utilized the strategy of analyzing millions of short reads from deep sequencing for the prediction of expressed sRNAs. Although the deep sequencing analyses used in this study focused on identifying low-molecular-weight RNAs (≤ 200 nt) by limiting the lengths of the RNA samples, assemblages of contiguously overlapping reads also overlap with longer regulatory RNAs.
During the past decade, several groups have extensively searched for sRNAs in Synechocystis sp. PCC 6803 using comparative genome analyses and experimental approaches [37], [39], [40]. Therefore, we examined some overlaps between the candidate sRNAs in this study and those identified in previous studies. In this study, 5211 candidate sRNAs were identified from exponentially growing cells, 27 of which were validated by RT-PCR. A total of only 33 overlapping candidate sRNAs were identified. The possible reasons are as follows. (1) It is quite possible that some of the bioinformatics-predicted sRNAs are not expressed at all. (2) This result is also possible because the majority of sRNAs are only expressed under stress conditions and are typically very tightly regulated [7] and may not have been expressed in our exponentially grown healthy sample pool. (3) Additionally, different bioinformatics-based approaches, such as [34] and ‘orphan’ transcriptional signals [40], only rely on transcriptional start sites, which may have resulted in our different results. Comparative bioinformatics is limited to the discovery of sRNAs that are conserved in closely related species, whereas experimental approaches, such as deep sequencing, should be able to comprehensively discover sRNAs in bacteria.
Plasmids containing sRNAs have been reported in various bacterial species [17], [57], [58]. Our results demonstrate that the plasmids in Synechocystis sp. PCC 6803 also express sRNAs, and 4 of those, including 3 intergenic RNAs (Table 1) and 1 asRNA (Table 2), were verified by RT-PCR, which suggests plasmids containing sRNAs could be a common feature of prokaryotic sRNAs. In fact, one of these candidate sRNAs, a cluster coded in pSYSA, has been identified in more than 13,000 reads, agreeing with previously reported results from Chlamydia trachomatis [57]. In addition, some candidate sRNAs were found in the antisense orientation to known genes, implying a role in the regulation of plasmid-coded genes. However, the roles of plasmid-encoded sRNAs are less clear and have been reported to be involved in the control of plasmid replication [59] and plasmid copy number [60], [61] and are perhaps related to the regulation of chromosomal gene expression [62].
sRNAs can be generated either as primary transcripts or through their processing from longer precursor transcripts [44]. In the current study, some of the identified sRNAs were observed for more than one transcript (Fig. 3), suggesting a working mechanism for the sRNAs in Synechocystis sp. PCC 6803 that may also involve the post-transcriptional processing of the primary transcripts. More recently, two asRNAs, Psb2R and Psb3R, located in 5′-untranslated regions of psbA2 and psbA3 were validated as two distinct signals with fragment lengths of 129 and approximately 220 nucleotides and 157 and 181 nucleotides, respectively, in Synechocystis sp. PCC 6803 [46].
While there has been a rapid increase in the identification of bacterial sRNAs in recent years, the identification of sRNA targets has proceeded relatively slowly. In E. coli, the number of validated sRNAs is approximately 80; however, only approximately 20 sRNAs have been assigned cellular functions [14], [63]. However, small RNA-mediated post-transcriptional regulation has emerged as an important regulatory mechanism for gene expression in various cellular processes [64]. One small RNA concurrently regulates several target mRNAs by binding to the corresponding mRNA via either perfect or imperfect sequence complementarity or by modulating protein activities, such as the Hfq-dependent GcvB sRNA, which has been shown to target more than 20 different mRNAs [65]. By targeting mRNA transcripts or affecting the activities of proteins at one time, sRNAs regulate a wide range of metabolic processes, such as quorum sensing [66], two-component signal transduction systems [65], photosynthesis [9], responses to the environment [67] and the regulation of outer membrane porins [68]. In this study, we analyzed all putative target genes in the KEGG pathway database, and the targets of the sRNAs were shown to be involved in 89 known metabolic or signaling pathways, suggesting that these sRNAs might play a significant role in regulating different biological processes and pathways such as photosynthesis, responses to diverse environments and the biosynthesis of secondary metabolites. Therefore, this work provides a solid platform for further experimental analyses of sRNAs and for the identification of the biological functions of these specific sRNAs.
To summarize, we have comprehensively analyzed sRNA expression under normal growth conditions using Synechocystis sp. PCC 6803 and predicted 5211 candidate sRNAs. We also identified several novel sRNAs that are highly expressed in plasmid regions, and four of these, including three intergenic sRNAs and one asRNA, were verified experimentally. Though the functions for the novel specific sRNAs obtained in this study remain unknown, the reliability of the targets for all the predicted sRNAs was further analyzed for the functional categorization of the biological pathways. The results indicate that these sRNAs are indeed involved in various metabolic pathways. This current study also provides more information about the regulatory network of sRNAs and will advance our understanding of sRNA functions in cyanobacteria.
Supporting Information
Contains the Files: Table S1. List of all predicted candidate intergenic small RNAs. Table S2. List of some putative top-scoring antisense RNAs(≥40 reads). Table S3. List of all predicted 5′ leader region candidates. Table S4. Classification of predicted targets of candidate small regulatory RNAs according to their biological process, molecular function and cellular component. Table S5. Complete list of enriched GO terms of target genes in Synechocystis sp. PCC 6803. Table S6. Complete list of KEGG pathways in the identified target genes of small regulatory RNAs. Table S7. List of the primer oligonucleotides used in this study.
(DOCX)
Funding Statement
This work was supported jointly by the National Program on Key Basic Research Project (2011CB200902) and the National Natural Science Foundation of China (31270094). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Schopf JW, Packer BM (1987) Early Archean (3.3-billion to 3.5-billion-year-old) microfossils from Warrawoona Group, Australia. Science 237: 70–73. [DOI] [PubMed] [Google Scholar]
- 2. Ikeuchi M (1996) Complete genome sequence of a cyanobacterium Synechocystis sp. PCC 6803, the oxygenic photosynthetic prokaryote. Tanpakushitsu Kakusan Koso 41: 2579–2583. [PubMed] [Google Scholar]
- 3. Atsumi S, Higashide W, Liao JC (2009) Direct photosynthetic recycling of carbon dioxide to isobutyraldehyde. Nature Biotechnology 27: 1177–1180. [DOI] [PubMed] [Google Scholar]
- 4. Ducat DC, Way JC, Silver PA (2011) Engineering cyanobacteria to generate high-value products. Trends In Biotechnology 29: 95–103. [DOI] [PubMed] [Google Scholar]
- 5. Quintana N, Van der Kooy F, Van de Rhee MD, Voshol GP, Verpoorte R (2011) Renewable energy from Cyanobacteria: energy production optimization by metabolic pathway engineering. Applied Microbiology And Biotechnology 91: 471–490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Huang HY, Chang HY, Chou CH, Tseng CP, Ho SY, et al. (2009) sRNAMap: genomic maps for small non-coding RNAs, their regulators and their targets in microbial genomes. Nucleic Acids Research 37: D150–154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Waters LS, Storz G (2009) Regulatory RNAs in bacteria. Cell 136: 615–628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Park SY, Cromie MJ, Lee EJ, Groisman EA (2010) A bacterial mRNA leader that employs different mechanisms to sense disparate intracellular signals. Cell 142: 737–748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Duhring U, Axmann IM, Hess WR, Wilde A (2006) An internal antisense RNA regulates expression of the photosynthesis gene isiA. Proceedings of the National Academy of Sciences of the United States of America 103: 7054–7058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Lee EJ, Groisman EA (2010) An antisense RNA that governs the expression kinetics of a multifunctional virulence gene. Molecular Microbiology 76: 1020–1033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Hernandez JA, Alonso I, Pellicer S, Luisa Peleato M, Cases R, et al. (2010) Mutants of Anabaena sp. PCC 7120 lacking alr1690 and alpha-furA antisense RNA show a pleiotropic phenotype and altered photosynthetic machinery. Journal Of Plant Physiology 167: 430–437. [DOI] [PubMed] [Google Scholar]
- 12. Mandin P, Gottesman S (2010) Integrating anaerobic/aerobic sensing and the general stress response through the ArcZ small RNA. Embo Journal 29: 3094–3107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Marguerat S, Bahler J (2010) RNA-seq: from technology to biology. Cellular And Molecular Life Sciences 67: 569–579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Raghavan R, Groisman EA, Ochman H (2011) Genome-wide detection of novel regulatory RNAs in E. coli. Genome Research 21: 1487–1497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Pellin D, Miotto P, Ambrosi A, Cirillo DM, Di Serio C (2012) A genome-wide identification analysis of small regulatory RNAs in Mycobacterium tuberculosis by RNA-Seq and conservation analysis. PLoS One 7: e32723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Acebo P, Martin-Galiano AJ, Navarro S, Zaballos A, Amblar M (2012) Identification of 88 regulatory small RNAs in the TIGR4 strain of the human pathogen Streptococcus pneumoniae. RNA 18: 530–546. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Irnov I, Sharma CM, Vogel J, Winkler WC (2010) Identification of regulatory RNAs in Bacillus subtilis. Nucleic Acids Research 38: 6637–6651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Sharma CM, Hoffmann S, Darfeuille F, Reignier J, Findeiss S, et al. (2010) The primary transcriptome of the major human pathogen Helicobacter pylori. Nature 464: 250–255. [DOI] [PubMed] [Google Scholar]
- 19. Voss B, Bolhuis H, Fewer DP, Kopf M, Moke F, et al. (2013) Insights into the physiology and ecology of the brackish-water-adapted Cyanobacterium Nodularia spumigena CCY9414 based on a genome-transcriptome analysis. PLoS One 8: e60224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Flaherty BL, Van Nieuwerburgh F, Head SR, Golden JW (2011) Directional RNA deep sequencing sheds new light on the transcriptional response of Anabaena sp. strain PCC 7120 to combined-nitrogen deprivation. BMC Genomics 12: 332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Prevost K, Desnoyers G, Jacques JF, Lavoie F, Masse E (2011) Small RNA-induced mRNA degradation achieved through both translation block and activated cleavage. Genes & Development 25: 385–396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Thomason MK, Fontaine F, De Lay N, Storz G (2012) A small RNA that regulates motility and biofilm formation in response to changes in nutrient availability in Escherichia coli. Molecular Microbiology 84: 17–35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Wilms I, Voss B, Hess WR, Leichert LI, Narberhaus F (2011) Small RNA-mediated control of the Agrobacterium tumefaciens GABA binding protein. Molecular Microbiology 80: 492–506. [DOI] [PubMed] [Google Scholar]
- 24. Sonnleitner E, Gonzalez N, Sorger-Domenigg T, Heeb S, Richter AS, et al. (2011) The small RNA PhrS stimulates synthesis of the Pseudomonas aeruginosa quinolone signal. Molecular Microbiology 80: 868–885. [DOI] [PubMed] [Google Scholar]
- 25. Jorgensen MG, Nielsen JS, Boysen A, Franch T, Moller-Jensen J, et al. (2012) Small regulatory RNAs control the multi-cellular adhesive lifestyle of Escherichia coli. Molecular Microbiology 84: 36–50. [DOI] [PubMed] [Google Scholar]
- 26. Gogol EB, Rhodius VA, Papenfort K, Vogel J, Gross CA (2011) Small RNAs endow a transcriptional activator with essential repressor functions for single-tier control of a global stress regulon. Proceedings of the National Academy of Sciences of the United States of America 108: 12875–12880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Csiszar K, Houmard J, Damerval T, Tandeau de Marsac N (1987) Transcriptional analysis of the cyanobacterial gvpABC operon in differentiated cells: occurrence of an antisense RNA complementary to three overlapping transcripts. Gene 60: 29–37. [DOI] [PubMed] [Google Scholar]
- 28. Watanabe T, Sugiura M, Sugita M (1997) A novel small stable RNA, 6Sa RNA, from the cyanobacterium Synechococcus sp. strain PCC6301. Febs Letters 416: 302–306. [DOI] [PubMed] [Google Scholar]
- 29. Barrick JE, Sudarsan N, Weinberg Z, Ruzzo WL, Breaker RR (2005) 6S RNA is a widespread regulator of eubacterial RNA polymerase that resembles an open promoter. RNA 11: 774–784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Vioque A (1992) Analysis of the gene encoding the RNA subunit of ribonuclease P from cyanobacteria. Nucleic Acids Research 20: 6331–6337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Tous C, Vega-Palas MA, Vioque A (2001) Conditional expression of RNase P in the cyanobacterium Synechocystis sp. PCC6803 allows detection of precursor RNAs. Insight in the in vivo maturation pathway of transfer and other stable RNAs. Journal Of Biological Chemistry 276: 29059–29066. [DOI] [PubMed] [Google Scholar]
- 32. Hess WR, Fingerhut C, Schon A (1998) RNase P RNA from Prochlorococcus marinus: contribution of substrate domains to recognition by a cyanobacterial ribozyme. Febs Letters 431: 138–142. [DOI] [PubMed] [Google Scholar]
- 33. Schon A, Fingerhut C, Hess WR (2002) Conserved and variable domains within divergent rnase P RNA gene sequences of Prochlorococcus strains. International Journal Of Systematic And Evolutionary Microbiology 52: 1383–1389. [DOI] [PubMed] [Google Scholar]
- 34. Axmann IM, Kensche P, Vogel J, Kohl S, Herzel H, et al. (2005) Identification of cyanobacterial non-coding RNAs by comparative genome analysis. Genome Biology 6: R73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Axmann IM, Holtzendorff J, Voss B, Kensche P, Hess WR (2007) Two distinct types of 6S RNA in Prochlorococcus. Gene 406: 69–78. [DOI] [PubMed] [Google Scholar]
- 36. Wassarman KM (2007) 6S RNA: a regulator of transcription. Molecular Microbiology 65: 1425–1431. [DOI] [PubMed] [Google Scholar]
- 37. Voss B, Georg J, Schon V, Ude S, Hess WR (2009) Biocomputational prediction of non-coding RNAs in model cyanobacteria. BMC Genomics 10: 123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Steglich C, Futschik ME, Lindell D, Voss B, Chisholm SW, et al. (2008) The challenge of regulation in a minimal photoautotroph: non-coding RNAs in Prochlorococcus. PLoS Genetics 4: e1000173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Georg J, Voss B, Scholz I, Mitschke J, Wilde A, et al. (2009) Evidence for a major role of antisense RNAs in cyanobacterial gene regulation. Molecular Systems Biology 5: 305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Mitschke J, Georg J, Scholz I, Sharma CM, Dienst D, et al. (2011) An experimentally anchored map of transcriptional start sites in the model cyanobacterium Synechocystis sp. PCC6803. Proceedings of the National Academy of Sciences of the United States of America 108: 2124–2129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Li H, Durbin R (2009) Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 25: 1754–1760. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Huang da W, Sherman BT, Lempicki RA (2009) Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nature Protocols 4: 44–57. [DOI] [PubMed] [Google Scholar]
- 43. Dennis G Jr, Sherman BT, Hosack DA, Yang J, Gao W, et al. (2003) DAVID: Database for Annotation, Visualization, and Integrated Discovery. Genome Biology 4: P3. [PubMed] [Google Scholar]
- 44. Vogel J, Bartels V, Tang TH, Churakov G, Slagter-Jager JG, et al. (2003) RNomics in Escherichia coli detects new sRNA species and indicates parallel transcriptional output in bacteria. Nucleic Acids Research 31: 6435–6443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Li L, Huang D, Cheung MK, Nong W, Huang Q, et al. (2013) BSRD: a repository for bacterial small regulatory RNA. Nucleic Acids Research 41: D233–238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Sakurai I, Stazic D, Eisenhut M, Vuorio E, Steglich C, et al. (2012) Positive regulation of psbA gene expression by cis-encoded antisense RNAs in Synechocystis sp. PCC 6803. Plant Physiology 160: 1000–1010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Eisenhut M, Georg J, Klahn S, Sakurai I, Mustila H, et al. (2012) The antisense RNA As1_flv4 in the Cyanobacterium Synechocystis sp. PCC 6803 prevents premature expression of the flv4-2 operon upon shift in inorganic carbon supply. Journal Of Biological Chemistry 287: 33153–33162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Tucker BJ, Breaker RR (2005) Riboswitches as versatile gene control elements. Current Opinion In Structural Biology 15: 342–348. [DOI] [PubMed] [Google Scholar]
- 49. Serganov A, Nudler E (2013) A decade of riboswitches. Cell 152: 17–24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Busch A, Richter AS, Backofen R (2008) IntaRNA: efficient prediction of bacterial sRNA targets incorporating target site accessibility and seed regions. Bioinformatics 24: 2849–2856. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Kyle DJ, Ohad I, Arntzen CJ (1984) Membrane protein damage and repair: Selective loss of a quinone-protein function in chloroplast membranes. Proceedings of the National Academy of Sciences of the United States of America 81: 4070–4074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Iwai M, Katoh H, Katayama M, Ikeuchi M (2004) PSII-Tc protein plays an important role in dimerization of photosystem II. Plant And Cell Physiology 45: 1809–1816. [DOI] [PubMed] [Google Scholar]
- 53. Fujimori T, Hihara Y, Sonoike K (2005) PsaK2 subunit in photosystem I is involved in state transition under high light condition in the cyanobacterium Synechocystis sp. PCC 6803. Journal Of Biological Chemistry 280: 22191–22197. [DOI] [PubMed] [Google Scholar]
- 54. Wang Q, Jantaro S, Lu B, Majeed W, Bailey M, et al. (2008) The High Light-Inducible Polypeptides Stabilize Trimeric Photosystem I Complex under High Light Conditions in Synechocystis PCC 6803. Plant Physiology 147: 1239–1250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Eisenhut M, Georg J, Klahn S, Sakurai I, Mustila H, et al. (2012) The antisense RNA As1_flv4 in the Cyanobacterium Synechocystis sp. PCC 6803 prevents premature expression of the flv4-2 operon upon shift in inorganic carbon supply. J Biol Chem 287: 33153–33162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Voss B, Gierga G, Axmann IM, Hess WR (2007) A motif-based search in bacterial genomes identifies the ortholog of the small RNA Yfr1 in all lineages of cyanobacteria. BMC Genomics 8: 375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Albrecht M, Sharma CM, Reinhardt R, Vogel J, Rudel T (2010) Deep sequencing-based discovery of the Chlamydia trachomatis transcriptome. Nucleic Acids Research 38: 868–877. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Schluter JP, Reinkensmeier J, Daschkey S, Evguenieva-Hackenberg E, Janssen S, et al. (2010) A genome-wide survey of sRNAs in the symbiotic nitrogen-fixing alpha-proteobacterium Sinorhizobium meliloti. BMC Genomics 11: 245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Weaver KE (2007) Emerging plasmid-encoded antisense RNA regulated systems. Current Opinion In Microbiology 10: 110–116. [DOI] [PubMed] [Google Scholar]
- 60. Kurpiel PM, Hanson ND (2012) Point mutations in the inc antisense RNA gene are associated with increased plasmid copy number, expression of blaCMY-2 and resistance to piperacillin/tazobactam in Escherichia coli. Journal Of Antimicrobial Chemotherapy 67: 339–345. [DOI] [PubMed] [Google Scholar]
- 61. Okibe N, Suzuki N, Inui M, Yukawa H (2010) Antisense-RNA-mediated plasmid copy number control in pCG1-family plasmids, pCGR2 and pCG1, in Corynebacterium glutamicum. Microbiology 156: 3609–3623. [DOI] [PubMed] [Google Scholar]
- 62. Carlson JH, Whitmire WM, Crane DD, Wicke L, Virtaneva K, et al. (2008) The Chlamydia trachomatis plasmid is a transcriptional regulator of chromosomal genes and a virulence factor. Infection And Immunity 76: 2273–2283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Vogel J, Wagner EG (2007) Target identification of small noncoding RNAs in bacteria. Current Opinion In Microbiology 10: 262–270. [DOI] [PubMed] [Google Scholar]
- 64. Storz G, Vogel J, Wassarman KM (2011) Regulation by small RNAs in bacteria: expanding frontiers. Molecular Cell 43: 880–891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Coornaert A, Chiaruttini C, Springer M, Guillier M (2013) Post-transcriptional control of the Escherichia coli PhoQ-PhoP two-component system by multiple sRNAs involves a novel pairing region of GcvB. PLoS Genetics 9: e1003156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Lenz DH, Mok KC, Lilley BN, Kulkarni RV, Wingreen NS, et al. (2004) The small RNA chaperone Hfq and multiple small RNAs control quorum sensing in Vibrio harveyi and Vibrio cholerae. Cell 118: 69–82. [DOI] [PubMed] [Google Scholar]
- 67. Wassarman KM (2002) Small RNAs in bacteria: diverse regulators of gene expression in response to environmental changes. Cell 109: 141–144. [DOI] [PubMed] [Google Scholar]
- 68. Johansen J, Eriksen M, Kallipolitis B, Valentin-Hansen P (2008) Down-regulation of outer membrane proteins by noncoding RNAs: unraveling the cAMP-CRP- and sigmaE-dependent CyaR-ompX regulatory case. Journal Of Molecular Biology 383: 1–9. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Contains the Files: Table S1. List of all predicted candidate intergenic small RNAs. Table S2. List of some putative top-scoring antisense RNAs(≥40 reads). Table S3. List of all predicted 5′ leader region candidates. Table S4. Classification of predicted targets of candidate small regulatory RNAs according to their biological process, molecular function and cellular component. Table S5. Complete list of enriched GO terms of target genes in Synechocystis sp. PCC 6803. Table S6. Complete list of KEGG pathways in the identified target genes of small regulatory RNAs. Table S7. List of the primer oligonucleotides used in this study.
(DOCX)