

Abstract
Objective
To determine to what extent oligoclonal band (OCB) specificities are clonally interrelated and to what degree they are associated with corresponding B-cell responses in the peripheral blood (PB) of multiple sclerosis (MS) patients.
Methods
Mass-spectrometric proteomic analysis of isoelectric focused (IEF) cerebrospinal fluid (CSF) immunoglobulin G (IgG) was used in combination with next-generation deep-immune repertoire sequencing of PB and CSF IgG heavy chain variable regions from MS patients.
Results
We find evidence for ongoing stimulation and maturation to antibody-expressing B cells to occur primarily inside the central nervous system (CNS) compartment. B cells participating in OCB production can also be identified in PB; these cells appear to migrate across the blood–brain barrier and may also undergo further antigen stimulation in the periphery. In individual patients, different bands comprising OCBs are clonally related.
Interpretation
Our data provide a high-resolution molecular analysis of OCBs and strongly support the concept that OCBs are not merely the terminal result of a targeted immune response in MS but represent a component of active B cell immunity that is dynamically supported on both sides of the blood-brain barrier.
The presence of soluble clonal immunoglobulin G (IgG), also referred to as oligoclonal bands (OCBs), in the cerebrospinal fluid (CSF) represents a central immunodiagnostic feature for multiple sclerosis (MS), detected in >95% of patients.1 OCBs result from intrathecal antigen-driven immune responses2,3 against as yet unknown target antigens. Typically visualized as discrete bands on isoelectric focusing (IEF) gel electrophoresis, OCBs have been associated with a more rapid conversion from clinically isolated syndrome to clinically definite MS,4–6 providing evidence that they may reflect more active central nervous system (CNS)-directed autoimmunity or otherwise contribute to tissue damage. Under treatment with natalizumab, a peripherally acting anti-VLA4 monoclonal antibody that blocks immune cell migration into the CNS and effectively reduces MS disease activity, CSF IgG levels can decrease and OCBs disappear.7,8 These changes are associated with a decrease in intrathecal lymphocytes,7,9 suggesting that an ongoing exchange of immune cells between the peripheral blood (PB) and the CNS is required to maintain intrathecal B-cell stimulation and OCBs.
Although oligoclonal CSF IgG has been shown to be a product of intrathecal plasma cells and B cells,3,10 essentially nothing is known about how B cells and/or plasma cells migrate to the CNS compartment and establish immunologically active sites in MS brain. It is likely that antigen-directed affinity maturation and terminal differentiation of B cells contributing to OCBs occur within perivascular infiltrates and meningeal lymphoidlike follicles in the CNS.11,12 Attempts to define the antigenic specificity of B-cell immunity in MS have yielded mixed and partially conflicting results. Whereas some studies demonstrated binding of intrathecally produced IgG and/or OCBs to myelin,13 myelin proteins,14,15 and Epstein–Barr virus antigens,16 others have reported an absence of reactivity to myelin antigens.17
Our own work suggests that IgG-expressing B cells and/or plasma cells participate in an ongoing exchange across the blood-brain barrier (BBB), a process operative during clinically active and also seemingly quiescent phases of the disease.18 It is not known whether B cells or plasma cells that express Ig corresponding to OCBs are also present in the PB, where they may contribute to CNS-targeted immune responses. Bystander activation in the periphery has been suggested as a mechanism by which proinflammatory B cells might support CNS-directed T-cell–mediated autoimmunity in MS19; however, there is currently no evidence in MS linking antigen-driven immunity in the periphery to targeted humoral immune responses in the CNS compartment.
We performed extensive mass-spectrometric proteomic analysis of CSF OCBs from 5 MS patients; previously described IgG heavy chain variable region (IgG-VH) sequence data sets obtained by deep-immune repertoire sequencing of PB and CSF were used for reference to match proteomic data with IgG-VH transcripts.18 Our findings suggest that ongoing antigenic stimulation and maturation of B cells to antibody-expressing plasma cells and plasmablasts occurs mostly inside the CNS compartment. More importantly, we show that B cells involved in OCB expression may also egress into the PB, where they may undergo further affinity maturation.
Patients and Methods
Patients and Samples
CSF samples from 5 patients who provided informed consent and who met diagnostic criteria for MS20 were available for our studies (Table 1; Supplementary Table S1 for additional clinical information). All patients displayed multiple OCBs in their CSF that were not detectable in their serum. Immediately after CSF collection via lumbar puncture (LP), CSF was centrifuged and supernatants were stored separately from cell pellets at −80°C; PB was obtained during the same patient visit. All 5 patients have been previously reported.18 IgG-VH transcriptomes were generated from CSF cells and PB mononuclear cells (PBMCs) using deep immune repertoire sequencing (DIRS). OCB IgG proteomes were analyzed by mass spectrometry using IgG isolated from CSF supernatants and separated by IEF. The study was approved by the University of California, San Francisco Committee on Human Research.
Table 1.
Patient Characteristics
| Patient ID | Age, yr | Sex | CSF, ml | WBC | OCB | VHref-CSF | VHref-PB |
|---|---|---|---|---|---|---|---|
| MS-1 | 29 | F | 8 | 1 | + | 294 | 135,346 |
| MS-2 | 37 | F | 10 | 1 | + | 419 | 60,609 |
| MS-3 | 39 | F | 10 | ND | + | 1,025 | 127,973 |
| MS-4 | 39 | F | 10 | 3 | + | 61 | 78,136 |
| MS-5 | 22 | M | 8 | 4 | + | 1,723 | 50,207 |
Shown are age, sex, CSF volume obtained for this study, CSF WBC, presence of OCBs as determined by a clinical laboratory (+ if ≥2 CSF-restricted IgG bands), and numbers of IgG-VH sequences per patient contained in our IgG-VH reference database. Refer to Supplementary Table S1 for additional clinical information. VHref-CSF represents IgG-VH sequences that are restricted to the CSF or belong to bicompartmental clusters of IgG-VH sequences; VHref-PB contains IgG-VH sequences from peripheral blood without a link to the CSF compartment (see Patients and Methods).
CSF = cerebrospinal fluid; F = female; Ig = immunoglobulin; IgG-VH = IgG heavy chain variable region; M = male; ND = not determined; OCB = oligoclonal band; PB = peripheral blood; WBC = white blood cell count.
DIRS
In brief, RNA from the CSF cell pellet or PBMC (1 × 106) was isolated and reverse transcribed using the SMARTer RACE cDNA Amplification Kit (Clontech, Mountain View, CA). cDNA was amplified using the SMARTer RACE universal 5′ primer mix and an IgG isotype–specific 3′ primer (5′-GGG AAG ACS GAT GGG CCC TTG GTG G-3′) to allow for unbiased, Ig heavy chain variable germline gene (IGHV)-independent amplification of IgG-VHs. IgG-VH transcript libraries were prepared using GS FLX Titanium kits (Lib-L chemistry, 454 sequencing; Roche, Basel, Switzerland). Polymerase chain reaction (PCR) amplification was performed at 95°C for 5 minutes; 95°C for 30 seconds, 65°C for 30 seconds, and 72°C for 1 minute for 33 cycles; and 72°C for 7 minutes. IgG-VH PCR products were purified using AMPure XP beads (Beckman Coulter Genomics, Waltham, MA) and quantified using a Quant-iT PicoGreen dsDNA kit (Invitrogen, Carlsbad, CA). Purified PCR products were diluted to 1 × 109 molecules/μl and subjected to emulsion PCR and unidirectional sequencing (from the IgG isotype–specific primer end) using the GS FLX Titanium Lib-L chemistry (Roche).
IgG-VH Repertoire Sequence Analysis
For all IgG-VH sequencing reads, IGHV and Ig heavy chain joining (IGHJ) germline gene usage, and IgG heavy chain CDR3 region (H-CDR3) were determined using VDJFasta software as previously described21; in addition to unique sequence identifiers resulting from DIRS, each sequence was tagged with a patient-specific identifier and the compartment of origin (CSF or PBMC). Reads not passing quality criteria (uniquely classified IGHV, IGHJ, and H-CDR3) were filtered from subsequent analysis. An average of 16,892 (standard deviation [SD] = ±10,248) CSF IgG-VHs and 318,130 (SD = ±114,984) PBMC IgG-VHs were generated from each sample (CSF IgG-VHs per patient: MS-1, 5,661; MS-2, 20,752; MS-3, 25,658; MS-4, 6,164; MS-5, 26,226; PBMC IgG-VHs per patient: MS-1, 507,411; MS-2, 235,515; MS-3, 243,301; MS-4, 347,616; MS-5, 256,809). Only sequences with contiguous reading frames from H-CDR1 to the joining region were used to generate our reference databases (see below). Clusters of clonally related IgG-VH sequences were identified using a distance metric approach as previously described (Levenshtein distance of 0 or 1 between their H-CDR3 amino acid sequences).22
Our study was designed (1) to identify proteomic IgG signatures in CSF (OCB peptides) that would allow us to determine whether OCBs are exclusively produced by CSF B cells or can also be linked to peripheral B-cell repertoires, and (2) to determine whether separate IgG bands (OCBs) on IEF electrophoresis might contain clonally related IgG. Availability of DIRS data permitted clustering of IgG-VHs into groups of clonally related sequences that were derived exclusively from CSF, exclusively from PB, or from both compartments. To match IgG-VH transcriptomic with proteomic data, 2 IgG-VH sequence databases containing sequences that resulted from DIRS of CSF and PBMCs of all 5 patients were generated:
VHref-CSF contained a pool of 3,522 IgG-VH sequences from all 5 patients reported here that were either derived from CSF-restricted IgG-VH sequence clusters, or found in PB and clonally related to CSF-derived IgG-VHs. In VHref-CSF 2,308 IgG-VHs were derived from CSF and 1,214 from PB. VHref-CSF permitted identification of OCB peptides that are clearly linked to the CSF compartment, either because they are directly found in the CSF IgG-VH transcriptome, or because they map to PB IgG-VH sequences that are clonally related to CSF IgG-VHs.
VHref-PB contained a pool of 452,270 PBMC IgG-VH transcripts from all 5 patients reported here. None of the IgG-VHs in VHref-PB had clonally related sequences among CSF IgG-VHs. VHref-PB permitted identification of OCB peptides mapping to IgG-VH sequences contained in PB-restricted clusters, that is, for which no clonally related IgG-VHs could be found in the CSF-associated IgG-VH transcriptome from the same patient.
Isolation of IgG from CSF, IEF, and Mass Spectrometry
Four to 6ml of cell-free CSF supernatant was incubated overnight with Protein G Sepharose 4 Fast Flow beads (GE Healthcare, Buckinghamshire, UK) in the presence of protease inhibitors and IgG eluted using glycine buffer at pH 2.8. The eluate was immediately neutralized using Tris buffer and processed for IEF using the ReadyPrep 2D Cleanup kit (BioRad, Hercules, CA) according to the manufacturer’s instructions. Eluates resuspended in rehydration buffer (BioRad) were loaded onto pH 3 to 10, 11cm immobilized pH gradient (IPG) strips (BioRad) and rehydrated for 16 hours. IEF on IPG strips was performed according to the manufacturer’s instructions. At the completion of the IEF run, the IPG strips were fixed in trichloroacetic acid (Sigma, St Louis, MO) and sulfosalicylic acid (Sigma) for 1 hour and stained with a Colloidal Blue Staining Kit (Invitrogen) for 1 hour. Discrete bands representing clonal IgG were excised from the IPG strips and processed for mass spectrometry. Established methods of tryptic in-gel digestion were used for processing of IPG strip slices containing OCBs (see Supplementary Methods for details).
IgG-VH Proteome/Transcriptome Comparisons
Masses of tryptic OCB peptides as determined by tandem mass spectrometry (see Supplementary Methods) were compared to in silico trypsin-digested IgG-VHs contained in the reference databases (VHref-CSF, VHref-PB). Due to VHref-CSF/PB only containing variable region (IgG-VH) sequences, OCB peptides mapping to IgG constant regions were not included in our analyses. We applied a multistep approach to determine whether OCB peptides were (1) patient specific, (2) mapped to IgG-VH clusters that support a B-cell exchange between CSF and PB, and 3) map to IgG-VH clusters represented by separate IgG bands on IEF electrophoresis. First, we identified patient-specific OCB peptides that were found exclusively in the same patient’s CSF proteome and IgG-VH transcriptome represented by VHref-CSF. For example, a CSF OCB peptide derived from Patient MS-1 was considered patient specific if it mapped exclusively to Patient MS-1 IgG-VH transcripts in VHref-CSF; patient-specific OCB peptides from Patient MS-2 had to map to MS-2 IgG-VH transcripts, MS-3 OCB peptides had to map to MS-3 IgG-VH transcripts, et cetera, to be considered patient specific. Second, we analyzed all OCB peptides to determine whether they mapped uniquely to a single patient’s PBMC IgG-VHs in VHref-PB but not in VHref-CSF; only 1 peptide was found meeting these criteria and was also considered patient specific. Third, OCB peptides were assigned to clusters of clonally related IgG-VH sequences (see next paragraph). Only patient-specific peptides with full sequence confirmation by tandem mass spectrometry (see Supplementary Methods) were used for further analysis. All patient-specific OCB peptides were analyzed via IgBlast (http://www.ncbi.nlm.nih.gov/igblast/) to determine similarity with known IGHV germline gene sequences.
OCB-Associated IgG-VH Clusters and Lineages
To determine whether peptides mapped to IgG-VHs expressed by clonally related B cells exclusively found in the CSF, in PB, or in both compartments, OCB peptides were first aligned to IgG-VH sequences contained in our databases (VHref-CSF/VHref-PB), and all IgG-VH transcripts containing exact OCB peptide sequence matches were retrieved from our databases. Then, IgG-VH sequences with identical IGHV and IGHJ usage, and highly similar/identical H-CDR3 amino acid sequence were retrieved from our IgG-VH databases to generate groups (clusters) of clonally related IgG-VH sequences representative of CSF OCB-derived peptides.
To understand the relationship of OCB peptides with IgG-VH lineage development during affinity maturation, we generated lineage trees (for examples see Figs 3) of clustered IgG-VH sequences containing OCB peptide sequences. IgG-VH sequences with a contiguous reading frame spanning at least from the 5′ end of H-CDR1 to the 3′ end of H-CDR3 were aligned using ClustalW 2.1.23 Putative germline sequences including the V-D-J junction were determined using SoDA24 and used as tree roots for lineage tree calculations. IgTree software25 (kindly provided by Dr R. Mehr, Bar-Ilan University, Ramat-Gan, Israel) was used to place IgG-VH sequences along lineage trees. Lineage trees were displayed using the hierarchic (for clusters up to 50 nodes) or organic (for clusters >50 nodes) layout in Cytoscape (version 2.8.326); in Figures 3, IgG-VHs exclusively found in CSF are labeled blue, those in PB are red, and those present in both compartments are green. Putative germline sequences are black and represent the root of each lineage tree. Unknown intermediates calculated by IgTree are beige. The size of tree nodes is proportional to the number of identical sequences contained within each node. IgG-VH amino acid alignments were generated using the MAFFT algorithm27 within Jalview28 software.
Figure 3.

B-cell clusters participating in oligoclonal band (OCB) production undergo immune stimulation in the periphery. (A) Lineage tree (organic layout) and (B) alignment of representative immunoglobulin G (Ig) heavy chain variable region (IgG-VH) amino acid sequences from Patient MS-5 are shown. Germline node (black arrow), node comprised of cerebrospinal fluid (CSF) and peripheral blood (PB) mononuclear cell (PBMC) IgG-VH (green arrow), and PB-derived subcluster identified by OCB peptide searches (gray arrow) are indicated. In the alignment, matched OCB peptides are shaded in gray; IgG-VH is derived from the Ig heavy chain variable germline gene (IGHV)3–9 germline segment. Only representative IgG-VHs are shown. Organic layout was chosen over hierarchic layout because the latter generates very extensive horizontal images. For additional information, please refer to the legends of Figures 1 and 2.
Results
Oligoclonal Band Peptides Map to IgG-VH Transcripts from CSF and/or PB
As expected, IEF of purified patient CSF IgG revealed OCB in all 5 CSF samples (Fig 4). Tryptic digestion of OCBs, tandem mass spectrometry, and matching of peptide masses to VHref databases yielded a combined 385 OCB peptides from the 5 patients reported here (Supplementary Table S2). About one-third (n = 135) of all identified OCB peptides were determined as not patient specific and were excluded from further analysis. A total of 249 OCB peptides were patient specific in VHref-CSF, 79 of which were fully sequence-confirmed and used for further analyses (Table 2; Supplementary Tables S2, S3); 1 additional sequence-confirmed and entirely H-CDR3–derived OCB peptide from Patient MS-1’s CSF could be specifically mapped to that patient’s IgG-VH transcriptome in VHref-PB but not in VHref-CSF (see Supplementary Table S3, gray shading). Together, 80 patient-specific OCB peptides were identified and used for further analysis.
Figure 4.

Oligoclonal bands (OCBs) represent diverse and partially related immunoglobulin G (IgG) populations. Shown are isoelectric focused (IEF) cerebrospinal fluid and serum IgG from Patients MS-1 to MS-5. IEF gels were stained and processed for mass spectrometry as described in Patients and Methods; analyzed gel slices are labeled with letters. *Gel slices yielding patient-specific OCB bands. **Gel slices yielding peptides linking to peripheral blood mononuclear cell–only clusters or subclusters. IEF bands yielding IgG OCB peptides belonging to the same cluster are connected by gray brackets; dashed and solid lines were used for better visual separation.
Table 2.
OCB Peptides and Associated IgG-VH Clusters
| Patient ID | OCB Peptides | Associated IgG-VH Clusters | CSF-Restricted IgG-VH Clusters |
|---|---|---|---|
| MS-1 | 7 | 6 | 5 |
| MS-2 | 20 | 14 | 13 |
| MS-3 | 38 | 20 | 20 |
| MS-4 | 3 | 2 | 2 |
| MS-5 | 12 | 10 | 4 |
Shown are numbers of OCB peptides that could be uniquely assigned to a specific patient in our IgG-VH reference database and associated IgG-VH clusters. CSF-restricted IgG-VH clusters are those in which OCB peptides mapped to IgG-VH sequence clusters that were exclusively found in the CSF.
CSF = cerebrospinal fluid; IgG-VH = IgG heavy chain variable region; OCB = oligoclonal band.
General OCB Peptide Characteristics
The 80 patient-specific OCB peptides were derived from 18 IEF gel slices (see Fig 4). OCB peptide lengths ranged from 8 to 27 amino acids; peptide coverage per IgG-VH sequence ranged from 1 to 4; an overview of OCB peptides mapping to IgG-VH sequences is shown in Supplementary Figure S1. IGHV usage among all OCB peptides reported here and associated IgG-VHs is shown in Figure 5A; IGHV3–7 (17.7%), IGHV4–30-4/31 (11.8%), IGHV4–39 (17.7%), and IGHV4–59/61 (9.8%) were most frequently found; H-CDR3 length of peptide-associated IgG-VHs (see Fig 5B) ranged from 10 to 27 amino acids with an average H-CDR3 length of 18.1 ± 3.5 (SD) amino acids, as expected for a typical normal IgG population.
Figure 5.
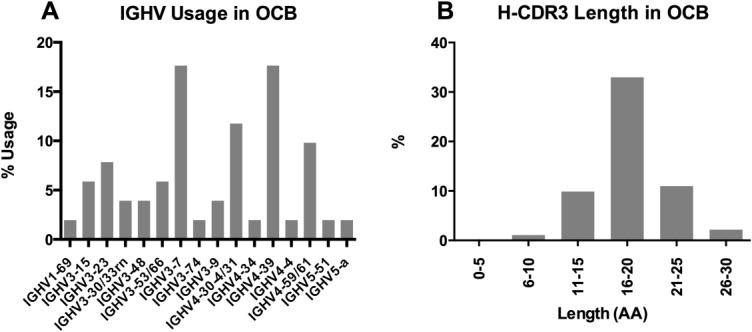
(A) Immunoglobulin G (IgG) heavy chain variable germline gene (IGHV) usage and (B) heavy chain CDR3 region (H-CDR3) length distribution of IgG heavy chain variable region clusters associated with oligoclonal band (OCB) peptides. AA = amino acids.
OCB Peptides Reflect Molecular Diversity of IgG-VHs
We were interested in understanding the molecular diversity associated with patient-specific OCB peptides reported here. For this purpose, we selected IgG-VHs that contained OCB peptide sequences from our VHref data sets and generated clusters of OCB-associated, clonally related IgG-VHs. The resulting clusters are reflective of mutational activity in the IgG-VHs, and clustered IgG-VHs can be used for phylogenetic lineage analysis. The 80 patient-specific OCB peptides could be assigned to 52 different clusters of clonally related IgG-VHs (see Table 2). Among the described peptides, 57 differed from IGHV germline sequences, indicating the presence of somatic mutations; 18 OCB peptides did not yield germline IGHV matches in IgBlast, generally because these sequences corresponded in large part to a non–germline-encoded H-CDR3 region and/or adjacent IGHJ sequence; 5 OCB peptides were identical to germline IGHV but mapped exclusively to IgG-VHs from Patient MS-2 or Patient MS-5 in VHref-CSF (see Supplementary Table S3). In general, OCB peptides that map largely or entirely to H-CDR3 sequences, or include mutations, are the most reliable tools to clearly assign peptides to a specific IgG-VH cluster. Germline gene-identical OCB peptides (n = 5; see Supplementary Table S3) were included in our analyses if they were patient specific, and within a patient also exclusively mapped to a single cluster of clonally related IgG-VHs.
In 1 patient (MS-3), we identified a number of highly similar OCB peptides incorporating amino acid mutations, demonstrating that progression of somatic hypermutation can be represented in OCBs (Fig 6). Numerous OCB peptides mapped to clusters of related IgG-VH sequences that were represented in >1 band on the IEF gels (see Fig 4, gray brackets; Supplementary Table S3), indicating the presence of related IgG in spatially separated OCB.
Figure 6.
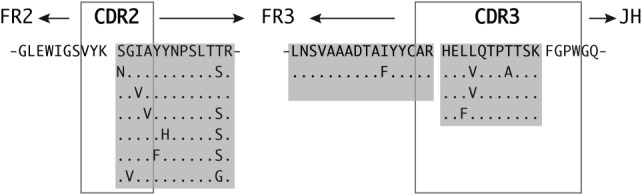
Oligoclonal band (OCB) peptides reveal signs of affinity maturation. Shown is an immunoglobulin G (IgG) heavy chain variable germline gene 4–39–derived, truncated IgG heavy chain variable region with OCB peptides identified by mass spectrometry in isoelectric focusing gel slices A, B, D, J, and K (see Fig 4; Supplementary Table S3) in Patient MS-3’s cerebrospinal fluid. Shaded in light gray are aligned amino acid sequences of identified OCB peptides reflective of ongoing affinity maturation in a B-cell cluster contributing to OCB production in Patient MS-3. Heavy chain CDR2 region (H-CDR2) and H-CDR3 are in gray frames. Dots indicate identity with the consensus sequence (top).
OCB-Producing B-Cell Clusters Participate in Active Immunity on Both Sides of the BBB
To address the question whether OCBs are exclusively associated with intrathecal immune responses, or whether B cells in the periphery may also express the same or clonally related Ig genes, we analyzed the compartmental (ie, CNS vs periphery) contributions in OCB-associated IgG-VH clusters. We found the majority of patient-specific OCB peptides to be associated with IgG-VH transcripts observed exclusively in the CSF (see Table 2; Supplementary Table S3); this finding supports the well-accepted finding that significant activation of CNS-restricted B-cell responses, including affinity maturation of Ig genes and terminal differentiation to antibody-secreting B cells, occurs inside the CNS in MS patients.18,29–31
However, we also found a number of OCB peptides mapping to bicompartmental clusters composed of CSF and/or PB IgG-VH transcripts from Patients MS-1, MS-2, and MS-5 (see Figs 1, 2A–D, 3). In Patient MS-2, with clinically quiescent MS, we identified an OCB peptide from IEF band D (see Fig 4, MS-2) mapping to the H-CDR3 region (detected peptide: YFAWSAGK; corresponding H-CDR3: CVAVRYFAWSAGKLFDYW) of a bicompartmental IgG-VH cluster (see Fig 1) utilizing IGHV4–39 and IGHJ4 germline segments. Alignment and lineage analysis of IgG-VH sequences belonging to this cluster resulted in a lineage tree with 3 distinctive branches. The identified H-CDR3 OCB peptide mapped to a CSF sublineage; in the PB and remaining CSF sublineage, this sequence portion displayed a single amino acid replacement from V to A at amino acid position 6 of the described peptide.
Figure 1.

Bicompartmental B-cell lineage contributing to oligoclonal band (OCB) production in Patient MS-2. Shown are the (A) lineage tree (hierarchic layout) and (B) alignment of immunoglobulin G (IgG) heavy chain variable region (IgG-VH) amino acid sequences from cerebrospinal fluid (CSF) and peripheral blood (PB) mononuclear cells. Lineage trees are calculated using nucleotide sequences IgTree software and displayed using Cytoscape (see Patients and Methods). In the lineage tree, each round node represents at least 1 unique IgG-VH sequence ranging from at least the 5′ end of heavy chain CDR1 region (H-CDR1) to the 3′ end of H-CDR3; larger nodes represent up to hundreds of identical sequences. CSF-derived IgG-VHs are represented by blue nodes, PB-derived IgG-VHs are red. Numbers between nodes are numbers of nucleotide mutations; unlabeled connections between nodes represent single nucleotide mutations. Putative germline sequences were determined using SoDA24 and are labeled black; hypothetical intermediates calculated by IgTree are beige. Numbers on lines between nodes (edges) represent mutational steps (nucleotides) between nodes. Triangular nodes contain 2 or more singleton sequences in leaves. The OCB peptide (YFAWSAGK) identified from isoelectric focusing band D (see Fig 4) maps to the H-CDR3 and was only found in a CSF-restricted sublineage. Additional sublineages in PB and CSF suggest that B-cell affinity maturation and plasma cell maturation (OCB production) occur in parallel in both compartments. IGHV = Ig heavy chain variable germline gene.
Figure 2.

B-cells involved in oligoclonal band (OCB) production are present in the peripheral blood. Shown are (A–D) examples of bicompartmental IgG heavy chain variable region (IgG-VH) lineages (hierarchic layout) from Patient MS-5, and (E) a IgG-VH lineage from Patient MS-1 in which OCB peptides mapped to peripheral blood mononuclear cellimmunoglobulin G (IgG) sequences exclusively. Cerebrospinal fluid–derived IgG-VHs are represented by blue nodes, peripheral blood–derived IgG-VHs are red, and identical sequences found in both compartments are green. Numbers between nodes are numbers of nucleotide mutations; unlabeled connections between nodes represent single nucleotide mutations. Numbers in nodes are numbers of OCB peptides mapping to a specific node. Triangular nodes contain 2 or more singleton sequences in leaves. Ig heavy chain variable germline gene (IGHV) and Ig heavy chain joining (IGHJ) usage, matched peptide, and corresponding representative CDR3 are indicated per lineage. Amino acids differing from germline segments are underlined; in E, the detected peptide is entirely located in the non–germline-encoded heavy chain CDR3 region.
The largest number of OCB peptides mapping to bicompartmental IgG-VH clusters were found in Patient MS-5, with active MS and a gadolinium contrast-enhancing lesion on magnetic resonance imaging (MRI) 1 week prior to LP. In this patient, we identified 12 patient-specific OCB peptides, 8 of which mapped to bicompartmental IgG-VH clusters (see Supplementary Table S3; for examples, see Figs 2A–D and 3). Interestingly, 2 OCB peptides from Patient MS-5 mapped exclusively to PB IgG-VH sequences in a bicompartmental IgG-VH cluster (see Fig 3). In addition, we identified a single OCB peptide from Patient MS-1 (detected peptide: VMGFYEDSGYR; corresponding H-CDR3: CARVMGFYEDSGYRYFTGLNDYW; IGHV4-4, IGHJ4) that did not match any CSF-restricted or bicompartmental IgG-VH clusters; instead, this peptide mapped to an IgG-VH cluster that was exclusively comprised of PB-derived Ig transcripts found in VHref-PB (see Fig 2E).
Discussion
OCBs are produced by clonally expanded, terminally differentiated B cells within the CNS compartment; they are believed to mark a highly targeted immune response against a specific target antigen, or antigens that await unequivocal identification; OCBs are among the strongest indicators that an antigen-driven humoral immune process participates in MS pathology. Unfortunately, OCBs cannot be readily sequenced, and isolated proteomic analysis of OCBs is unreliable due to amino acid changes introduced in the Ig variable region during affinity maturation. Combining transcriptomic data of Ig genes involved in OCB expression with OCB proteomic analysis by mass spectrometry was previously applied to demonstrate that OCBs are produced by intrathecal plasma cells and B cells.2,3 Using next-generation DIRS, we recently demonstrated in MS and other neurological disorders that B-cell repertoires on both sides of the BBB are connected.18 The availability of soluble CSF IgG and comprehensive CSF and PB IgG-VH immune repertoires from 5 representative MS patients now enabled us to formally demonstrate that B cells participating in OCB production in the CSF can also be found in the periphery.
It is well accepted that OCB are produced by CSF B cells3,10; our finding that most OCB peptides map to CSF-derived IgG-VH sequences underscores the importance of such intrathecal antigen-driven immune responses. However, it is not known whether clusters of related B cells participating in OCB production are also present in the periphery; our studies did reveal such a connection in 3 of 5 patients. Two patients with clinically quiescent MS displayed single OCB-producing B-cell clusters that potentially participated in exchange across the BBB. The largest number of bicompartmental IgG-VH clusters was identified in Patient MS-5, an individual with active MS and a contrast-enhancing lesion on brain MRI 1 week prior to LP. This finding suggests enhanced migration of B cells across the BBB during active disease. Our data do not permit a clear statement regarding directionality of B-cell exchange across the BBB. However, B cells that participate in OCB production likely represent a well-established and temporally invariable intrathecal immune response.32 Hence, our findings in Patient MS-5 provide circumstantial evidence that during active MS, B cells may egress from the CNS to the periphery. Based on our findings that some CSF-associated B-cell clusters apparently undergo extensive somatic hypermutation (ie, affinity maturation) in the periphery, it is also possible that B cells that have already participated in intrathecal immune stimulation and OCB production can egress from the CNS and undergo antigen-directed restimulation in the periphery. Clonal B-cell reservoirs may be a source of replenishment for OCB-producing B cells; such reservoirs may be located within the CSF, in meningeal lymphoid folliclelike structures,11 intraparenchymal B-cell infiltrates, or the periphery.
Placing OCB peptides along lineage trees has helped to clarify the relationship between OCB production and B-cell affinity maturation. We found a number of instances in which OCB peptides could be clearly assigned to an IgG-VH cluster but within this cluster mapped to a distinct sublineage, an observation that pertains to both CSF-restricted and bicompartmental IgG-VH clusters. Thus, it is possible that clonal B-cell expansion and plasma cell differentiation necessary for OCB production are a function of dynamic B-cell immunity rather than its final result. Within permissive inflammatory environments, plasma cell maturation and affinity maturation likely occur in parallel, resulting in antibody populations derived from clonally related B cells that may contain heavy chain variable regions reflective of different stages of affinity maturation.
An important question awaiting elucidation is whether OCB contribute to tissue damage in MS. B-cell–depleting therapy with the anti-CD20–targeting monoclonal antibodies rituximab and ocrelizumab was recently found to rapidly and effectively suppress inflammatory disease activity in MS.33–35 Interestingly, in one report OCB were unchanged 24 weeks after rituximab therapy despite a reduction in intrathecal B-cell numbers,36 to some degree suggesting an inferior role of OCBs in MS immune pathology. Conversely, there is evidence that limiting migration of lymphocytes across the BBB by natalizumab (anti-VLA4 antibody) significantly reduces intrathecal antibody production.7,8 Our data presented here support the concept that disease-relevant B-cell immune responses are active on both sides of the BBB, a connection that could be effectively interrupted by natalizumab.
Antigens recognized by OCBs or intrathecal B cells remain unknown, despite extensive efforts by many groups, and despite growing evidence that these immune responses must be driven by specific immunogenic stimuli. We find that clonally related IgG-VH sequences are associated with OCB that are spatially separated on IEF gels. Thus, immune responses against target epitopes could be represented by numerous OCBs; in other words, the presence of numerous OCBs does not necessarily constitute an intrathecal immune response against multiple target epitopes. Our finding that B-cell clusters producing OCBs in the CNS also undergo somatic hypermutation in the periphery strongly suggests the presence of identical or highly similar antigens on both sides of the BBB. Recently, antibodies against the potassium channel protein KIR4.1 have been described in almost 50% or MS patients’ serum,37 and injection of purified KIR4.1 antibodies into the CSF compartment of mice was shown to induce CNS tissue damage. The initial trigger of KIR4.1-directed autoimmunity is unknown; however, molecular mimicry between peripheral and CNS self or foreign antigens could be an important concept supporting bicompartmental B-cell immunity in MS. The pathological relevance of immune responses against myelin oligodendrocyte glycoprotein (MOG) and myelin basic protein (MBP) has been controversially discussed in MS; mimicry between MOG and the milk protein butyrophilin has been demonstrated for both antibodies and T cells in MS.38,39 MBP-specific T-cell clones have also been found to react to viral peptides.40
We provide evidence for a deep connection between OCB and B-cell clusters in patients with MS, and we demonstrate for the first time that “OCBs” (ie, antibodies represented by OCBs) can theoretically be produced in the periphery. In single individuals, multiple different bands are comprised of closely related protein sequences corresponding to clonally expanded B cells present in the nervous system and also in the periphery. Remarkably, some B-cell clusters or subclusters corresponding to OCBs are observed only in the PB. Perhaps sampling at an earlier or different time or at a significantly greater sequencing depth would have identified the B cells corresponding to these OCBs in the CSF, or alternatively some OCBs could be the product of B cells present in the meninges or brain parenchyma but not in CSF. Whatever the explanation, the finding that some OCB specificities are connected only to PB B cells indicates that disease-relevant B cells circulate between the CNS and peripheral compartments. Equally important, this finding suggests that the periphery represents a site of persistence and also likely of activation of pathogenic B cells in MS. Lastly, our findings add support to the concept that molecular sequencing of B-cell receptors could be used as a diagnostic and monitoring tool for longitudinal tracking of disease activity in MS.18
Acknowledgments
These studies were supported by grants from Pfizer, the Nancy Davis Foundation, Small Ventures USA, the Brass Family Foundation, the National Multiple Sclerosis Society (RG-4868 to HCvB), the NIH National Institute of Neurologic Disorders and Stroke (K02NS072288 to HCvB; R01NS026799 to SLH). H.-C.v.B. was also supported by an endowment from the Rachleff Family Foundation. Mass spectrometry analysis was performed by the University of California, San Francisco Sandler-Moore Mass Spectrometry Core Facility, which acknowledges support from the Sandler Family Foundation, the Gordon and Betty Moore Foundation, and NIH/National Cancer Institute Cancer Center Support Grant P30 CA082103.
We thank the individuals who agreed to serve as subjects for this study; R. Mehr for providing IgTree; R. Niles for help with mass spectrometry experiments and data analyses; and T. Kuo, M. Sirota, S. Pitts, S. Potluri, and D. Shelton for helpful discussions and suggestions.
We dedicate this work to the memory of our friend and colleague David R. Cox, MD, PhD, whose contributions to the concepts and technologies reported here enabled these studies; we will always be inspired by David’s lifelong dedication to biomedical research as a means to prevent suffering and improve human health.
Authorship
J.B. and L.A. contributed equally.
Potential Conflicts of Interest
S.L.H.: board membership, BioMarin. H.-C.v.B.: consultancy, Novartis; grants/grants pending, Roche; grants/grants pending, Pfizer.
Supporting Information
Additional Supporting Information may be found in the online version of this article.
References
- Walsh MJ, Tourtellotte WW, Roman J, Dreyer W. Immunoglobulin G, A, and M–clonal restriction in multiple sclerosis cerebrospinal fluid and serum—analysis by two-dimensional electrophoresis. Clin Immunol Immunopathol. 1985;35:313–327. doi: 10.1016/0090-1229(85)90092-3. [DOI] [PubMed] [Google Scholar]
- Obermeier B, Lovato L, Mentele R, et al. Related B cell clones that populate the CSF and CNS of patients with multiple sclerosis produce CSF immunoglobulin. J Neuroimmunol. 2011;233:245–248. doi: 10.1016/j.jneuroim.2011.01.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Obermeier B, Mentele R, Malotka J, et al. Matching of oligoclonal immunoglobulin transcriptomes and proteomes of cerebrospinal fluid in multiple sclerosis. Nat Med. 2008;14:688–693. doi: 10.1038/nm1714. [DOI] [PubMed] [Google Scholar]
- Avasarala JR, Cross AH, Trotter JL. Oligoclonal band number as a marker for prognosis in multiple sclerosis. Arch Neurol. 2001;58:2044–2045. doi: 10.1001/archneur.58.12.2044. [DOI] [PubMed] [Google Scholar]
- Joseph FG, Hirst CL, Pickersgill TP, et al. CSF oligoclonal band status informs prognosis in multiple sclerosis: a case control study of 100 patients. J Neurol Neurosurg Psychiatry. 2009;80:292–296. doi: 10.1136/jnnp.2008.150896. [DOI] [PubMed] [Google Scholar]
- Tintore M, Rovira A, Rio J, et al. Do oligoclonal bands add information to MRI in first attacks of multiple sclerosis? Neurology. 2008;70:1079–1083. doi: 10.1212/01.wnl.0000280576.73609.c6. [DOI] [PubMed] [Google Scholar]
- Villar LM, Garcia-Sanchez MI, Costa-Frossard L, et al. Immunological markers of optimal response to natalizumab in multiple sclerosis. Arch Neurol. 2012;69:191–197. doi: 10.1001/archneurol.2011.971. [DOI] [PubMed] [Google Scholar]
- Harrer A, Tumani H, Niendorf S, et al. Cerebrospinal fluid parameters of B cell-related activity in patients with active disease during natalizumab therapy. Mult Scler. 2013;19:1209–1212. doi: 10.1177/1352458512463483. [DOI] [PubMed] [Google Scholar]
- Stuve O, Marra CM, Jerome KR, et al. Immune surveillance in multiple sclerosis patients treated with natalizumab. Ann Neurol. 2006;59:743–747. doi: 10.1002/ana.20858. [DOI] [PubMed] [Google Scholar]
- von Büdingen HC, Gulati M, Kuenzle S, et al. Clonally expanded plasma cells in the cerebrospinal fluid of patients with central nervous system autoimmune demyelination produce “oligoclonal bands.”. J Neuroimmunol. 2010;218:134–139. doi: 10.1016/j.jneuroim.2009.10.005. [DOI] [PubMed] [Google Scholar]
- Serafini B, Rosicarelli B, Magliozzi R, et al. Detection of ectopic B-cell follicles with germinal centers in the meninges of patients with secondary progressive multiple sclerosis. Brain Pathol. 2004;14:164–174. doi: 10.1111/j.1750-3639.2004.tb00049.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prineas JW. Multiple sclerosis: presence of lymphatic capillaries and lymphoid tissue in the brain and spinal cord. Science. 1979;203:1123–1125. doi: 10.1126/science.424741. [DOI] [PubMed] [Google Scholar]
- von Büdingen HC, Harrer MD, Kuenzle S, et al. Clonally expanded plasma cells in the cerebrospinal fluid of MS patients produce myelin-specific antibodies. Eur J Immunol. 2008;38:2014–2023. doi: 10.1002/eji.200737784. [DOI] [PubMed] [Google Scholar]
- Cruz M, Olsson T, Ernerudh J, et al. Immunoblot detection of oligoclonal anti-myelin basic protein IgG antibodies in cerebrospinal fluid in multiple sclerosis. Neurology. 1987;37:1515–1519. doi: 10.1212/wnl.37.9.1515. [DOI] [PubMed] [Google Scholar]
- O’Connor KC, Appel H, Bregoli L, et al. Antibodies from inflamed central nervous system tissue recognize myelin oligodendrocyte glycoprotein. J Immunol. 2005;175:1974–1982. doi: 10.4049/jimmunol.175.3.1974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cepok S, Zhou D, Srivastava R, et al. Identification of Epstein-Barr virus proteins as putative targets of the immune response in multiple sclerosis. J Clin Invest. 2005;115:1352–1360. doi: 10.1172/JCI23661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Owens GP, Bennett JL, Lassmann H, et al. Antibodies produced by clonally expanded plasma cells in multiple sclerosis cerebrospinal fluid. Ann Neurol. 2009;65:639–649. doi: 10.1002/ana.21641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- von Büdingen HC, Kuo TC, Sirota M, et al. B cell exchange across the blood-brain barrier in multiple sclerosis. J Clin Invest. 2012;122:4533–4543. doi: 10.1172/JCI63842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bar-Or A, Fawaz L, Fan B, et al. Abnormal B-cell cytokine responses a trigger of T-cell-mediated disease in MS? Ann Neurol. 2010;67:452–461. doi: 10.1002/ana.21939. [DOI] [PubMed] [Google Scholar]
- Polman CH, Reingold SC, Banwell B, et al. Diagnostic criteria for multiple sclerosis: 2010 revisions to the McDonald criteria. Ann Neurol. 2011;69:292–302. doi: 10.1002/ana.22366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glanville J, Zhai W, Berka J, et al. Precise determination of the diversity of a combinatorial antibody library gives insight into the human immunoglobulin repertoire. Proc Natl Acad Sci U S A. 2009;106:20216–20221. doi: 10.1073/pnas.0909775106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levenshtein VI. Binary codes capable of correcting deletions, insertions, and reversals. Soviet Physics Dokl. 1965;10:707–710. [Google Scholar]
- Larkin MA, Blackshields G, Brown NP, et al. Clustal W and Clustal X version 2.0. Bioinformatics. 2007;23:2947–2948. doi: 10.1093/bioinformatics/btm404. [DOI] [PubMed] [Google Scholar]
- Volpe JM, Cowell LG, Kepler TB. SoDA: implementation of a 3D alignment algorithm for inference of antigen receptor recombinations. Bioinformatics. 2006;22:438–444. doi: 10.1093/bioinformatics/btk004. [DOI] [PubMed] [Google Scholar]
- Barak M, Zuckerman NS, Edelman H, et al. IgTree: creating immunoglobulin variable region gene lineage trees. J Immunol Methods. 2008;338:67–74. doi: 10.1016/j.jim.2008.06.006. [DOI] [PubMed] [Google Scholar]
- Smoot ME, Ono K, Ruscheinski J, et al. Cytoscape 2.8: new features for data integration and network visualization. Bioinformatics. 2011;27:431–432. doi: 10.1093/bioinformatics/btq675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Katoh K, Toh H. Recent developments in the MAFFT multiple sequence alignment program. Brief Bioinform. 2008;9:286–298. doi: 10.1093/bib/bbn013. [DOI] [PubMed] [Google Scholar]
- Waterhouse AM, Procter JB, Martin DM, et al. Jalview Version 2—a multiple sequence alignment editor and analysis workbench. Bioinformatics. 2009;25:1189–1191. doi: 10.1093/bioinformatics/btp033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qin Y, Duquette P, Zhang Y, et al. Clonal expansion and somatic hypermutation of V(H) genes of B cells from cerebrospinal fluid in multiple sclerosis. J Clin Invest. 1998;102:1045–1050. doi: 10.1172/JCI3568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Owens GP, Ritchie AM, Burgoon MP, et al. Single-cell repertoire analysis demonstrates that clonal expansion is a prominent feature of the B cell response in multiple sclerosis cerebrospinal fluid. J Immunol. 2003;171:2725–2733. doi: 10.4049/jimmunol.171.5.2725. [DOI] [PubMed] [Google Scholar]
- Baranzini SE, Jeong MC, Butunoi C, et al. B cell repertoire diversity and clonal expansion in multiple sclerosis brain lesions. J Immunol. 1999;163:5133–5144. [PubMed] [Google Scholar]
- Walsh MJ, Tourtellotte WW. Temporal invariance and clonal uniformity of brain and cerebrospinal IgG, IgA, and IgM in multiple sclerosis. J Exp Med. 1986;163:41–53. doi: 10.1084/jem.163.1.41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bar-Or A, Calabresi PA, Arnold D, et al. Rituximab in relapsing-remitting multiple sclerosis: a 72-week, open-label, phase I trial. Ann Neurol. 2008;63:395–400. doi: 10.1002/ana.21363. [DOI] [PubMed] [Google Scholar]
- Hauser SL, Waubant E, Arnold DL, et al. B-cell depletion with rituximab in relapsing-remitting multiple sclerosis. N Engl J Med. 2008;358:676–688. doi: 10.1056/NEJMoa0706383. [DOI] [PubMed] [Google Scholar]
- Kappos L, Li D, Calabresi PA, et al. Ocrelizumab in relapsing-remitting multiple sclerosis: a phase 2, randomised, placebo-controlled, multicentre trial. Lancet. 2011;378:1779–1787. doi: 10.1016/S0140-6736(11)61649-8. [DOI] [PubMed] [Google Scholar]
- Cross AH, Stark JL, Lauber J, et al. Rituximab reduces B cells and T cells in cerebrospinal fluid of multiple sclerosis patients. J Neuroimmunol. 2006;180:63–70. doi: 10.1016/j.jneuroim.2006.06.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Srivastava R, Aslam M, Kalluri SR, et al. Potassium channel KIR4.1 as an immune target in multiple sclerosis. N Engl J Med. 2012;367:115–123. doi: 10.1056/NEJMoa1110740. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stefferl A, Schubart A, Storch M, et al. Butyrophilin, a milk protein, modulates the encephalitogenic T cell response to myelin oligodendrocyte glycoprotein in experimental autoimmune encephalomyelitis. J Immunol. 2000;165:2859–2865. doi: 10.4049/jimmunol.165.5.2859. [DOI] [PubMed] [Google Scholar]
- Guggenmos J, Schubart AS, Ogg S, et al. Antibody cross-reactivity between myelin oligodendrocyte glycoprotein and the milk protein butyrophilin in multiple sclerosis. J Immunol. 2004;172:661–668. doi: 10.4049/jimmunol.172.1.661. [DOI] [PubMed] [Google Scholar]
- Wucherpfennig KW, Strominger JL. Molecular mimicry in T cell-mediated autoimmunity: viral peptides activate human T cell clones specific for myelin basic protein. Cell. 1995;80:695–705. doi: 10.1016/0092-8674(95)90348-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.