

Abstract
Soy isoflavones are dietary components for which an association has been demonstrated with reduced risk of prostate cancer (PCa) in Asian populations. However, the exact mechanism by which these isoflavones may prevent the development or progression of PCa is not completely understood. There are a growing number of animal and in vitro studies that have attempted to elucidate these mechanisms. The predominant and most biologically active isoflavones in soy products, genistein, daidzein, equol, and glycetin, inhibit prostate carcinogenesis in some animal models. Cell-based studies show that soy isoflavones regulate genes that control cell cycle and apoptosis. In this review, we discuss the literature relevant to the molecular events that may account for the benefit of soy isoflavones in PCa prevention or treatment. These reports show that although soy isoflavone-induced growth arrest and apoptosis of PCa cells are plausible mechanisms, other chemo protective mechanisms are also worthy of consideration. These possible mechanisms include antioxidant defense, DNA repair, inhibition of angiogenesis and metastasis, potentiation of radio- and chemotherapeutic agents, and antagonism of estrogen- and androgen-mediated signaling pathways. Moreover, other cells in the cancer milieu, such as the fibroblastic stromal cells, endothelial cells, and immune cells, may be targeted by soy isoflavones, which may contribute to soy-mediated prostate cancer prevention. In this review, these mechanisms are discussed along with considerations about the doses and the preclinical models that have been used.
Keywords: Isoflavone, Genistein, Prostate Cancer, Chemoprevention
1. Introduction
Prostate cancer (PCa) is the most common malignancy and the second-leading cause of cancer death among men in the United States. It is estimated that 241,740 men will be diagnosed with PCa, and 28,170 men will die from PCa in 2012 [1]. Despite the high incidence, little is known regarding the etiology. Accepted risk factors are age, race, ethnicity, and geographical location. Although most of the localized PCa are surgically removable, there is still a subset of patients who present at late non-operable stages with little chance for an effective cure. Therefore, more efforts should be devoted for developing preventive strategies to reduce PCa prevalence and impact.
The incidence of PCa is much lower in Asian than that in Western populations [2]. However, Asian migrants to the United States have an increased incidence [3]. Therefore, environmental factors, especially the diet, have been presumed to play a key role in prostate carcinogenesis. Many Asian populations traditionally consume large amounts of soy food that is rich in isoflavones, such as genistein and daidzein. Mean serum or plasma concentrations of isoflavones in Japanese men are 10 to 100 times higher than those in European men [4]. Moreover, prostate fluid concentrations of daidzein and equol were much higher in Asian than in Western men [5]. Over the past decade, researchers have obtained evidence to suggest that soy isoflavones and their metabolites may be beneficial for the prevention or treatment of PCa. Some population-based studies have shown that soy isoflavone levels are associated with a reduced risk of PCa [6]. Animal and in vitro studies have yielded evidence in support of the notion that the use of isoflavones may be beneficial for PCa prevention and treatment. There are, however, findings from epidemiologic studies indicating a lack of associations between isoflavone or soy food intake and PCa risk. Thus, randomized clinical trials are additionally needed. There is an apparent shortage in clinical studies that examined the protective effects of soy isoflavones with PCa risk as an endpoint. There are many limitations for this kind of clinical studies such as the need for larger cohorts and longer period of intervention which is complicated by noncompliance and long duration of follow-up. Thus, most of the interventional studies used serum PSA levels as an end point and showed inconsistent results (Summarized in table 1). Some trials have shown that soy protein or isoflavone supplementation has no effects on serum PSA levels in healthy men [7–9], men at high risk of PCa [10,11], Men with localized PCa before treatment [12,13]or men with biochemical failure after a prior therapy [14–16]. However, other trials demonstrated the ability of soy isoflavones to reduce serum PSA or slow down PSA rising in subsets of men with biochemical failure after surgical intervention [17–19] or men with localized PCa and high PSA prior to therapy [20–22]. A small number of clinical studies addressed other protective effects of soy targeting endpoints other than PSA such as prostaglandins [23], VEGF [19], anti-apoptotic and proliferative markers [10] and AR [24].
Table 1.
Clinical Trials of Soy chemoprevention of PCa.
| Reference | Design | Participants | Soy or isoflavone intake in the intervention group | Duration of trial | Main outcomes | Results |
|---|---|---|---|---|---|---|
| Bosland et al, 2013 [14] | Randomized, double-blind trial | 177 men with localized prostate cancer after radical prostatectomy ((intervention (n=87) and placebo (n=90)) | Daily consumption of 20g soy protein isolate | 2 years | Serum PSA | Null |
| Lazarevic et al, 2011 [20] | Randomized, double-blind trial | 54 men with localized prostate cancer prior to prostatectomy ((intervention (n = 23) and placebo (n = 24)) | Daily consumption of 30 mg synthetic genistein | 3–6 weeks | Tissue and serum PSA | Genistein reduced serum PSA levels in patients with localized PCa |
| Napora et al, 2011 [15] | Randomized, double-blind trial | 33 men undergoing androgen ablation therapy for PCa ((intervention (n=17) and placebo (n=16)) | Daily consumption of 20 g of soy protein containing 160 mg of total isoflavones | 12 weeks | Metabolic and inflammatory parameters | Null |
| deVere White et al, 2010 [12] | Randomized, double-blind trial from 0–6 months and open-label study from 6–12 months | 53 men with localized prostate cancer ((intervention (n = 23) and placebo (n = 24)) | Daily consumption of 450 mg genistein, 300 mg daidzein, and other isoflavones | 6 months | Serum PSA | Null |
| Kwan et al, 2010 [17] | Phase II trial | 29 PCa patients with biochemical failure after radiotherapy | Daily consumption of 50–100 mg of isoflavones | 6 months | PSA doubling times | Soy induced significant prolongation of PSA doubling time in 41% of PCa patients |
| Swami et al, 2009 [23] | Randomized, double-blind trial | 25 men with localized prostate cancer ((intervention (n = 13) and placebo (n = 12)) | Daily consumption of 81.6 isoflavone aglycones | 2 weeks | Prostaglandin synthesis | Soy isoflavones significantly reduced prostate COX-2 mRNA and increased p21 mRNA in PCa patients |
| Pendleton et al, 2008 [18] | Open-labeled, Phase II, nonrandomized trial | 20 patients with biochemical failure after prior local therapy | Daily consumption of 47 mg of isoflavonoid | 12 months | Serum PSA | Soy isoflavones decreased the slope of PSA in PCa patients. |
| Grainger et al, 2008 [19] | Open-labeled randomized trial | 21 patients with biochemical failure after prior local therapy | Daily consumption of 40 g of soy protein contain 80 mg of isoflavones | 4 weeks | Serum PSA and VEGF | Soy protein prolonged PSA doubling time and reduced VEGF in PCa patients |
| Hamilton-Reeves et al, 2008 [10] | Randomized trial | 58 men at high risk of prostate cancer ((HGPIN (n=40) or atypical small acinar proliferation (n=13)) or low grade prostate cancer (n=5) | Daily consumption of 40 g protein of soy protein (107 mg isoflavones) | 6 months | Anti-apoptotic (BAX) and proliferative (PCNA) tumor markers and serum PSA | Soy isoflavones decreased anti-apoptotic tumor markers and reduced the incidence of developing Pca with no significant effect on serum PSA levels |
| Hamilton-Reeves et al, 2007 [24] | same as Hamilton-Reeves et al, 2008 | same as Hamilton-Reeves et al, 2008 | same as Hamilton-Reeves et al, 2008 | same as Hamilton-Reeves et al, 2008 | Tumor AR and ER-β | Soy isoflavones suppressed tumor AR expression with no significant effect on ER-β expression |
| Maskarinec et al, 2006 [7] | Randomized double-blind crossover clinical trial | 23 healthy men | Daily consumption of 35–38 mg soy isoflavones | 3 months | Serum PSA | Insignificant reduction in PSA in 14% |
| Dalais et al, 2004 [21] | Randomized, double-blind trial | 29 men diagnosed with localized prostate cancer prior to radical prostatectomy | Daily consumption of 117mg soy isoflavones | NA | Serum PSA | Soy isoflavones reduced total PSA in PCa patients |
| Kumar et al, 2004 [13] | Randomized, double-blind trial | 59 men with localized prostate cancer ((intervention (n = 29) and placebo (n = 30)) | Daily consumption of 60 mg soy isoflavones | 12 weeks | Serum PSA | Insignificant reduction in total and free PSA in the intervention arm relative to the placebo arm |
| deVere White et al, 2004 [16] | open-label pilot study | 52 patients with biochemical failure after prior therapy | Daily consumption of 450 mg genistein and 450 mg aglycone isoflavones | 6 months | Serum PSA | Null |
| Adams et al, 2004 [8] | Randomized, double-blind trial | 81 healthy men ((intervention (n = 34) and control (n = 47)) | Daily consumption of 83 mg soy isoflavones | 12 months | Serum PSA | Null |
| Jenkins et al, 2003 [9] | Randomized double-blind crossover clinical trial | 46 healthy middle-aged men | Daily consumption of 116 mg isoflavones | 3–4 weeks | Serum PSA | Null |
| Hussain et al, 2003 [22] | open-label pilot study | 39 patients with Pca under watchful waiting with rising PSA (group I) or had increasing serum PSA following local therapy (group II) or while receiving hormone therapy (group III). | Daily consumption of 200 mg soy isoflavones | 3–6 months | Serum PSA | Soy isoflavones decreased the rate of the rise of serum PSA |
| Urban et al, 2001 [11] | Randomized double-blind crossover clinical trial | 34 men with elevated PSA more than 4ng/ml | Daily consumption of 40 gm of isolated soy protein contain 42 mg genistein and 27 mg daidzein | 6 weeks | Serum PSA | Null |
The mechanistic basis for the protective effect of soy isoflavones on PCa is yet incompletely understood. In the present review, we summarize the existing knowledge and hypotheses on the molecular mechanisms by which soy isoflavones can influence the development of PCa.
1.1 Brief review of soy isoflavones metabolism and their cellular actions
Isoflavones are diphenolic compounds present in plants like soybeans, red clover, and kudzu root. Soybeans are the most common dietary source for isoflavones. The predominant isoflavones in soybean are genistein, daidzein, and, in lower concentration, glycitein. Bioavailability of isoflavones depends on the way they are extracted, processed, and cooked. Soybeans contain 2–5 mg of isoflavones per gram of protein. Isoflavones in soy foods exist in the form of glucosides, conjugated with sugars. After ingestion, soybean isoflavones undergo hydrolysis by intestinal β-glucosidases, which releases the main bioactive aglycones, daidzein, genistein and glycitein. These aglycones may be absorbed or further metabolized in the gut to several specific metabolites, including equol. The extent of this metabolism varies considerably among individuals. It is affected by the composition and activity of intestinal flora and the degree of carbohydrate-richness of the diet, which control intestinal fermentation. Increased fermentation results in an extensive biotransformation of isoflavones, with a significant increase in production of equol from daidzein [25].
Genistein is the predominant and most biologically active isoflavone in the soy diet, and it can be taken up by cells without prior metabolic activation to exert its effects [26]. The plasma half-life of genistein and daidzein is approximately 7.9 hours in adults; the maximum concentration is reached 6–8 hours after intake. Plasma concentrations of 50–800 ng/ml have been found for genistein, daidzein and equol in adults who consume modest amounts of soy foods [27]. Prostate tissue levels of soy isoflavones exceed serum levels by as much as 4–6 fold in people daily consuming soy [28]. Despite the elevated serum levels that can be achieved by soy isoflavone consumption, Phase I clinical trials showed minimal toxicity in subjects treated with a purified soy unconjugated isoflavone mixture at doses as high as 16mg/kg body weight [29]. Data about soy isoflavone pharmacokinetics and pharmacodynamics put them in the top of the list of chemopreventive agents that worthy to be examined thoroughly for their protective effects against PCa.
1.2. Molecular mechanisms of soy isoflavone signaling
The soy isoflavones genistein, daidzein, and glycitein have a structural similarity to 17β-estradiol, which explains their estrogenic activity and identifies them as phytoestrogens (fig. 1). Because of this structural and functional similarity to estradiol, soy isoflavones can bind to estrogen receptors alpha (ER-α) and beta (ER-β). The affinity of genistein to ER-β is about 20–30 times higher than to ER-α and is comparable to the affinity of 17β-estradiol [30,31]. Soy isoflavones also induce the transcriptional activity of ERβ more than ERα [32]. The combination of genistein, daidzein, and glycitein increases the selectivity for ERβ binding [30,33]. The distribution of ERα and ERβ varies among different types of tissues. For instance, tissue of female reproductive system, uterus and breast, are abundant in ERα, whereas bone and prostate tissue have greater amounts of ERβ. This divergence in tissue distribution of ERα and ERβ receptors leads to the dissimilar effects of isoflavones among different tissues. In addition to their estrogenic activities, there is a growing body of evidence showing that isoflavones can modulate the expression of some genes that control cell survival [34], cell cycle [35], and apoptosis [36]. They also have antioxidant properties [37], inhibit tyrosine kinase activity [38], suppress NF-κB, and regulate Akt, MAPK [39,40], and other signaling pathways.
Figure 1.
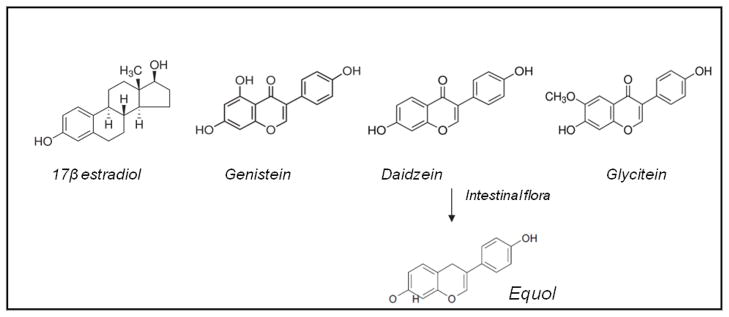
Molecular structures of 17-β-estradiol and isoflavones genistein, daidzein, equol and glycitein.
1.3. Effects of soy isoflavones on cell proliferation
Inhibition of PCa epithelial cell growth is one of the most recognized protective roles of soy isoflavones in PCa. We demonstrated in our recently published data that genistein inhibits the growth of LAPC-4 and PC-3 PCa cells in a dose-dependent manner [41]. Although our data showed this growth inhibitory effect was mediated mainly by the anti-androgenic activity of genistein especially at the low physiological doses, other contributory mechanisms have been proposed in literature. This includes, but not limited to, the ability of soy isoflavones to modify the expression of some central genes for cell survival [34], cell cycle [35], and apoptosis [39]. Soy isoflavones also reduce androgen receptor (AR) and prostate-specific antigen (PSA) levels, inhibit mammalian target of rapamycin (mTOR) activity, and induce growth arrest in several PCa cell lines. We will discuss the proposed mechanisms of the growth inhibitory effects of soy isoflavones on PCa cells in the following sections.
1.3.1 Cell-cycle regulators
Cell cycle is regulated at different phases by cyclins, cyclin dependent kinases (CDKs), and cyclin dependent kinase inhibitors (CDKIs). The expression and/or the function of most of these cell cycle regulators are disrupted during cancer development and progression. Thus, these regulators constitute important targets for cancer therapy and prevention.
Many investigators have searched for direct effects of soy isoflavones on the expression of genes that control cell growth. An example is genistein-mediated transcriptional up-regulation of p21, a CDKI, causing cell cycle arrest and apoptosis in both androgen sensitive and androgen independent PCa cell lines LNCaP and PC-3, respectively [42]. In DU145, another androgen independent PCa cell line, genistein treatment resulted in a significant induction of p21 and p27, reduction of CDK4, and a moderate inhibition of CDK2, cyclin D1 and cyclin E [43]. In another study, a combination of genistein and daidzein increased p53 and reduced cyclin B1 protein expression significantly in LNCaP and PC-3 PCa cells [44]. Furthermore, equol, which is a biologically active metabolite of daidzein, has shown cell-cycle regulatory effects on benign and malignant prostate epithelial cells at concentrations that can be obtained naturally through dietary soy consumption [45]. A fair number of studies have shown that these changes in cyclins and CDK inhibitors such as p21 and p27 coincide with soy isoflavone-induced growth arrest [46–48]. FOXO (forkhead box O) proteins are tumor suppressors that control cell proliferation [49]. The potential of soy isoflavones to modify FOXO family gene expression has been demonstrated recently by Christensen et al. [50] where high dietary intake of soy unregulated FOXO3 in the prostate of TRAMP mouse model. The function of several FOXO family members is inhibited by MAPK (mitogen-activated protein kinase) mediated phosphorylation [51]. Soy isoflavones genistein, equol, and daidzein showed an inhibitory effect on MAPK related pathway genes in LNCaP cells [52], which counteracts the MAPK mediated phosphorylation of FOXO proteins and could explain the ability of these isoflavones to increase FOXO protein expression. In support of this cell cycle regulatory role, a global gene expression analysis demonstrated that soy isoflavones target several genes central to cell cycle control in human PCa cell lines (LNCaP, DU145 and PC-3). These genes include cyclin B2, CDK4, CDK7, cyclin-dependent kinase inhibitors 1A and 2A (CDKI-1A and CDKI-2A), P53, and checkpoint kinase 2 (CHEK2) [53]. Collectively, these data suggest that cell cycle regulation offer a mechanistic explanation for the inhibitory effect of soy isoflavones on androgen dependent and independent PCa. Although some of these effects have been obtained using high doses of soy isoflavones [42,44,47,53], low physiological levels were also capable of regulating cell cycle progression in PCa cells [45,46,48,52].
1.3.2 IGF-1 (Insulin-Like Growth Factor-1) signaling
Soy isoflavones may indirectly influence the growth rate of PCa cells by interfering with the growth factors that stimulate the proliferation or increasing growth factors that promote cell differentiation. One of these factors that are modified by soy isoflavones is IGF-1. There is a growing body of experimental and epidemiologic evidence that elevated levels of IGF-1 are linked to prostate cancer progression [54]. IGF-1 can contribute to PCa development by blocking apoptosis and promoting cancer cell proliferation and invasion [55,56]. Most of these effects are mediated through the binding of IGF-1 to its membrane receptor IGF-1R, activating the receptor tyrosine kinase which subsequently phosphorylates intercellular substrates including insulin receptor substrate (IRS-1) which in turn activates a cascade of downstream signals such as PI3K/AKT and RAS/MAPK that stimulate cell proliferation and suppress apoptosis [57]. In PC-3 cells, genistein inhibited the IGF-1 stimulated cell growth, as well as the phosphorylation of its downstream targets Src, Akt, and glycogen synthase kinase-3β (GSk-3β) [58].
A pioneering research by Valentinis et al changed the way the action of IGF-1 was perceived [59,60]. They reported that IGF-1, besides being a mitogenic factor, it may also activate differentiation pathways. Their data demonstrated that the ratio of IGF-1R to IRS-1 controls the predominance of the proliferation pathway over the differentiation pathway or conversely. Low IGF-1R and high IRS-1 levels are required to inhibit the differentiation program while favoring cell proliferation. When IRS-1 is low, a dominant differentiation program is activated.
Wang et al [61] showed that genistein at 10 μmol/L inhibited the IGF-1 stimulated growth in AT6.3 rat PCa cells. Fascinatingly, it also modified IGF-1R/IRS-1 ratio through decreasing the overall cell content of IRS-1 and inhibiting the IGF-1 induced down-regulation of IGF-1R expression. The high IGF-1R and low IRS-1 in PCa cells after exposure to genistein enhanced terminal differentiation pathways and contributed to an eventual apoptotic cell death. Further evidence that IGF-1 is a critical target for the growth inhibitory properties of soy isoflavones comes from studies showing that a pretreatment of LNCaP and PC-3 PCa cells with 20 nmol/L of IGF-I induced a significant increase in the protein levels of both total AKT and phosphorylated AKT (Ser 473) and abolished genistein-mediated inhibition of AKT phosphorylation [47]. Microarray analyses of soy isoflavones treated PCa cells (LNCaP, DU 145, and PC-3) also showed down-regulation of a wide array of IGF-1 induced signaling pathways [52,53]. These data show that the ability of soy isoflavones to inhibit PCa cell proliferation is partially mediated by interference with the growth stimulatory factor, IGF-1 and/or modulating its downstream signaling pathways via adjusting the IGF-1R/IRS-1 ratio.
1.3.3 TGF-β (Transforming Growth Factor-β) signaling pathway
TGF-β is a pleiotropic cytokine that regulates multiple cellular functions. It influences normal prostate growth and differentiation by regulating the balance between epithelial cell proliferation and apoptosis. TGF-β acts as a tumor suppressor and changes roles to become a tumor promoter in the later stages of carcinogenesis [62]. At early stages of carcinogenesis, TGF-β regulates prostate growth by inhibiting epithelial cell proliferation and inducing apoptosis. However, in metastatic PCa, TGF-β serves as a tumor promoter. TGF-β exerts its action through Smad-dependent and Smad-independent mechanisms. Increased production of TGF-β results in immunosuppression, extracellular matrix degradation, epithelial to mesenchymal transition and angiogenesis, which promote tumor cell invasion and metastasis. Accordingly, there should be a balance in the molecular basis for effective therapeutic targeting of TGF-β in a way to inhibit the TGF-β tumor promoter function in advanced, metastatic PCa and at the same time maintain the growth-inhibitory function displayed in early stages of prostate tumorigenesis [63]. In the aggressive highly metastatic PCa cell variant (PC3M), TGF-β treatment resulted in increased phosphorylation of MAPK-Activated Protein Kinase 2 (MAPKAPK2) and MAP kinase p38 that regulate cell adhesion, migration, and invasion and show up regulated expression during PCa progression. Pretreatment with genistein almost totally abolished the TGF-β-induced phosphorylation of these proteins [64]. Matrix Metalloproteinase-2 (MMP-2) and Heat Shock Protein (HSP27) are another group of proteins that are phosphorylated and activated by TGF-β; they play a pivotal role in cell invasion and are often dysregulated in cancer. Genistein, at concentrations associated with dietary consumption, abrogated TGF-β-mediated increase in MMP-2 and HSP27 activity, in PC-3 and DU145 PCa cells, and reduced cell metastatic and invasive properties [64,65]. Gene expression profile studies showed that genistein reduced the mRNA expression of TGF-β and some other genes that are regulated by TGF-β such as; MMPs [66]. These data demonstrate that genistein at both low doses [64] and high doses [65] is capable of inhibiting TGF-β and its downstream signaling pathways in androgen independent PCa cells. Nevertheless, further research needs to be conducted to examine this effect on androgen sensitive PCa models. Also, other soy isoflavones need to be investigated for this effect.
1.3.4 Wnt/β-catenin signaling
An alternative hypothesis has emerged to explain how soy isoflavones mediate cell growth arrest, namely disruption of the function of β-catenin, the terminal mediator of Wnt signaling. In the cytoplasm, β-catenin is found in association with APC (adenomatous polyposis coli), axin, glycogen synthase kinase 3β (GSK3β) and other proteins. β-catenin undergoes phosphorylation by GSK3β leading to ubiquitin-mediated degradation. In the presence of Wnt ligand, frizzled binding causes disheveled (DSH) protein to dissociate glycogen synthase kinase (GSK) from the axin protein, thus blocking phosphorylation of β-catenin and set the later free. This free β-catenin translocates to the nucleus, binds with the transcription factor, T cell factor-4 (TCF-4) and activates transcription of genes that control cell proliferation (c-Myc and cyclin D1) [67,68]. Several studies have shown that Wnt pathway plays a critical role in the development and the progression of human PCa. High levels of β-catenin gene expression and accumulation in the nucleus are associated with advanced PCa. In addition, detection of mutations in β-catenin has been reported in PCa [69–71]. Soy isoflavones down regulate β-catenin expression and block β-catenin mediated gene transcription in PC-3 cells [72]. This mechanism is associated with an increased expression of GSK3β which serves as a downstream regulator of the Wnt/β-catenin pathway. Moreover, genistein plays an essential role in the cross-talk among different signaling pathways; its mediated inhibition of IGF-1 markedly attenuated the IGF-1-induced β-catenin signaling. Wnt/β-catenin inhibition by genistein subsequently resulted in an inhibition of the T-cell factor/lymphoid enhancer factor (TCF/LEF) dependent transcriptional activity and effective inhibition of PCa cell growth [58]. Consistent with these observations, a cross-species gene expression analysis has demonstrated that Wnt/β-catenin is one of the most influential pathways that mediate the effect of soy protein isolate and isoflavones on both human and rat PCa cell growth. Soy and genistein treatment shared the same gene regulation profile in the human PC-3 and the rat PAIII PCa cell lines, which indicates that genistein is likely the main contributor to the effect of soy on PCa cellular pathways [72]. Soy isoflavone-mediated events may also indirectly influence β-catenin function through increased production of E-cadherin, a membrane protein that can bind β-catenin and prevent its nuclear accumulation. Treatment of PC-3 cells with genistein (1–50 μmol/L) increased E-cadherin protein levels which led to a subsequent decrease in nuclear localization of β-catenin [58]. In summary, soy isoflavones are capable of suppressing Wnt/β-catenin pathway in PCa cells. Although some of these studies applied high doses of soy isoflavones (20–100 μmol/L) [72], this effect was quite achievable at lower physiological doses too [58].
1.3.5 Tyrosine kinase inhibition
Tyrosine kinases are enzymes that transfer a phosphate group from ATP (adenosine triphosphate) to a protein in the cell. Phosphorylation of tyrosine regulates many proteins such as epidermal growth factor (EGF), protein kinase B (PKB), MAPK, vascular endothelial growth factor receptor (VEGFR), platelet-derived growth factor receptor (PDGFR), stem cell factor receptor, and colony-stimulating factor-1 (CSF-1) receptor. Tyrosine kinases can sometimes function incorrectly and leads to derangement of the function of certain proteins essential for pathways such as cell proliferation, cell survival and cell adhesion. Reducing the activity of tyrosine kinase enzymes can be used to treat or correct a malfunctioning system. In PCa specifically, growth factor stimulation of receptor tyrosine kinases (RTK) such as insulin receptor, IGF, PDGF, or EGF activate several different pathways such as the PI3K/Akt signaling pathway, ultimately resulting in proliferation and survival of PCa cells. Tryrosine kinase activation is one of the pathways by which PCa can bypass androgen ablation and acquire androgen independence [73–76].
Soy isoflavones act as negative regulators of tyrosine kinase activity. This effect was verified in the androgen responsive (LNCaP) and the androgen unresponsive (PC-3) PCa cell lines. Soy isoflavones suppressed tyrosine kinase activity, reduced the phosphorylation of Akt, GSK-3β and p70S6k and inhibited the Akt/FOXO3a/GSK-3β/AR signaling network, as well as the Akt/mTOR signal transduction. These effects were postulated to mediate genistein-induced growth inhibition and apoptotic cell death in PCa cells [77–79]. In consistence with these findings, a microarray analysis performed on PC-3 and DU145 PCa cells have shown that genistein inhibits he phosphorylation of proteolytic enzymes that play a crucial role in PCa invasion such as urokinase-type plasminogen activator (uPA), MMPs, and various types of cathepsins [80]. This effect was comparable to that obtained by the well-recognized tyrosine kinase inhibitor; AG-1478 and was postulated to be mediated by the tyrosine kinase inhibition properties of genistein. In the above mentioned studies, tyrosine kinase inhibition was obtained in response to high doses of soy isoflavones which is consistent with findings from our laboratory showing that genistein inhibits tyrosine kinase only at the upper limit of physiologically attainable doses (≥ 10μmol/L) in both LAPC-4 and LNCaP cells [41].
1.4. Effects of soy isoflavones on apoptosis
Soy isoflavones increase basal and chemotherapy-induced apoptosis in PCa cells [81]. They promote apoptosis in LNCaP and PC-3 cells through upregulation of phosphatase and tensin homolog (PTEN), a tumor-suppressor gene that suppresses the anti-apoptotic Akt signaling pathway and subsequently downregulates the expression of AR [82]. Genistein, quercetin, and biochanin A, used separately or in a combination at a final concentration of 25μmol/L increased apoptosis in LNCaP cells, which was accompanied by downregulation of the anti-apoptotic protein Bcl-2 and increased Bax (pro-apoptotic) mRNA and protein levels [83]. Genistein at millimolar dose range induced apoptosis in LNCaP and DU145 PCa cells which was associated with enhanced activation and expression of caspase-3 and this effect was significantly inhibited by the caspase-3 inhibitor, N-benzyloxycarbonyl-Val-Asp-fluoromethyl-ketone (z-VAD-fmk) [84]. Another mechanism by which genistein-induced apoptosis is through inhibition of the proteasome. The proteasome is a large protein complex whose function is to degrade unneeded or damaged proteins by proteolysis. Kazi et al. [85] showed that genistein-mediated proteasome inhibition induces apoptosis by disrupting the regulated degradation of pro-growth cell cycle proteins. This effect, however, was significant only in response to higher doses of genistein (50μmol/L). Interestingly, soy extract induced more substantial apoptosis in PCa cells (LNCaP and PC-3) than genistein or daidzein. In non-cancerous BPH-1 cells, while no significant changes in apoptosis were observed in response to treatment with soy extract, genistein and daidzein augmented apoptosis, suggesting that the effects of soy extract may be more tumor cell-specific than the individual soy isoflavones [86]. Transcript profiling studies showed that the mRNA levels for the anti-apoptotic proteins, survivin, and protease activated receptor 2 (PAR-2) were downregulated in PC-3 cells treated with genistein (5–50μmol/L), whereas the mRNA levels for the pro-apoptotic protein elafin was upregulated [87]. Conclusively, soy isoflavones especially genistein have demonstrated a significant pro-apoptotic activity in androgen dependent and independent PCa cells. Soy isoflavone doses that have been used in most of these studies are pharmacological and do not reflect what can be achieved by the people consuming soy rich diet. Using low physiological doses of genistein (1–10 μmol/L), we encountered this pro-apoptotic activity in LAPC-4 cells that have wild type AR. At these low doses, however, we did not find any significant effect on apoptosis in LNCaP cells that have mutant AR and the apoptotic activity in this cell line started to be obvious only at higher doses of genistein (> 10μmol/L) [41]. Our data suggests that effects of genistein on apoptosis in PCa cells are dose sensitive and may vary among different PCa cell lines that possess different AR status which necessitates careful selection of the models to be used in future studies to test this effect of genistein in PCa.
1.5. Effects of soy isoflavones on PCa cell differentiation
Cell differentiation is the process in which unspecialized cells develop into specialized ones in response to a specific prompt from the body or cells themselves. This process occurs during tissue repair and normal cell turnover. Differentiation dramatically changes cell structure and function. These changes are mainly due to highly controlled modifications of gene expression. Cancer cells begin to multiply before they are fully mature and turn to an undifferentiated phenotype. Cancer cells can also lose properties they originally had, such as expression of specific protein; this process is termed dedifferentiation. In cancer pathology, the level of cellular differentiation is used as a measure of the degree of malignancy and “Tumor Grade” is a term that reflects how differentiated cells in that tumor are.
Although the effects of soy isoflavones on PCa cell proliferation have been thoroughly investigated, there are relatively few studies of their impact on PCa cell differentiation. Clubbs and Bomser [88] have shown that eight days of treatment with the soy isoflavones genistein, daidzein, and equol induced the expression of cytokeratin 18, a marker of luminal cell differentiation, in RWPE-1 PCa cells. However, the soy isoflavone glycetin stimulated an opposite effect by reducing the expression of cytokeratin 18 and maintaining expression of the basal epithelial cell markers p63 and cytokeratin 5 in the same cell line. TRAMP mice fed an AIN-76A-based genistein rich diet (containing 100, 250, or 500 mg of genistein per kg) showed a significant dose-dependent reduction in the incidence of poorly differentiated prostate adenocarcinoma relative to controls [89]. These data suggest that soy isoflavones may induce PCa cell differentiation. However, given the scarcity of research conducted to test this effect, further studies are required to elucidate this possible action of soy isoflavones.
1.6. Novel molecular events regulated by soy isoflavones that contribute to anticancer activity
In the past decade, soy isoflavones have been shown to regulate a much wider array of cellular events than previously thought. Below we will discuss some of these processes and their potential relevance to the prevention of cancer.
1.6.1 Autophagy
Autophagy is the process by which cells degrade, or turn-over, their own macromolecules and organelle components through the lysosomal machinery, often in response to damage, in order to provide substrates for energy metabolism and to recycle amino acids, fatty acids, and nucleotides for the biosynthetic needs of the cell. Autophagy is a tightly regulated process that plays a normal role in cell growth, development, and homeostasis. Although autophagy is considered a survival strategy to protect cells during stress (e.g. starvation and pro-oxidant conditions), this process is also regarded as a mode of programmed cell death [90]. Mammalian target of rapamycin (mTOR) is one of the major points of control within the autophagy signaling network [91]. In LNCaP and 22Rv1 PCa cells, soy isoflavones have been shown to suppress anti-autophagic mTOR signaling, potentially through a marked reduction of Threonine (389) phosphorylation of the mTOR substrate, p70 S6 kinase (p70S6K) [78]. Genistein induces autophagy in ovarian carcinoma cell lines (A2780, CaOV3, and ES2) and human colon cancer cells (HT-29) [92,93]. Despite these potentially relevant mechanistic relationships, neither in vitro nor in vivo research has directly connected the induction of autophagy to the anticancer activity of soy isoflavone compounds in PCa.
1.6.2 Antioxidant defense and DNA repair mechanisms
Reactive oxygen species (ROS) are continuously generated in aerobic organisms as a result of normal energy metabolism. However, excessive levels of ROS cause oxidative stress potentially creating a toxic environment to cells. This stressful condition may develop either due to antioxidant depletion or to increased generation of ROS because of exposure to noxious agents and/or pathologic processes. A group of endogenous enzymes is responsible for protecting cells against the excess ROS including superoxide dismutases (SODs), which convert supoeroxide (O2•−) to hydrogen peroxide (H2O2), glutathione peroxidase (GPx) that reduces free H2O2 to water, glutathione reductase (GSR) that reduces glutathione disulfide (GSSG) to the sulfhydryl form (GSH), and catalase that catalyzes the decomposition of H2O2 to water and oxygen [94].
Soy isoflavones induce the expression of several enzymes involved in the antioxidant defense system. Furthermore, they protect cells against free radical-induced DNA damage in both cancerous and non-cancerous tissue [95–98]. In PCa cells (LNCaP and PC-3), genistein induced the expression of the GPx-1 gene, which was accompanied by elevation of GPx enzyme activity [99]. However, this effect was achieved only at a high dose of genistein (100μmol/L) with no significant increase in GPx-1 in response to lower doses (≤5μmol/L). The GPx-1 protein plays a pivotal role in protecting cells from the oxidative damage through detoxifying H2O2 as well as, reducing lipid hydroperoxides to their corresponding alcohols. Genistein also induced the mRNA levels for other essential antioxidant proteins such as glutathione reductase, microsomal glutathione S-transferase 1, and metallothionein in LAPC-4 cells and this was linked to the reduced levels of DNA damage (assessed by comet assay) [100]. It is worth mentioning that some of these effects have been obtained in response to genistein doses as low as 1 μmol/L. Genistein at a dose of 10μmol/L induced manganese-superoxide dismutase (Mn-SOD) and catalase expression in DU-145 PCa cells and this was associated with significantly decreased ROS production [101]. There is some evidence that soy isoflavones regulate genes encoding proteins that protect the genome. Oki et al. [102] found that genistein upregulated mRNA and protein levels of the DNA damage-inducible gene 45 (GADD45) which is a p53 target gene whose product is involved in DNA repair. In DU145 cells, genistein induced G2/M arrest in PCa cells which was lost upon deletion of part of GADD45 promoter, suggesting the critical importance of GADD45 induction in genistein induced effects. Taken together, it appears that genistein directly regulates the expression of a variety of genes whose protein products are involved in DNA damage repair and programmed cell death, thereby offering protection against carcinogenesis.
1.6.3 Prostaglandin metabolism and action
A variety of studies has shown that prostaglandin signaling stimulates PCa cell growth and PCa progression [103,104]. In this context, cyclo-oxygenase-1 (COX-1) and COX2 are the rate-limiting enzymes in prostaglandin synthesis. It has become increasingly apparent that COX-2 overexpression is observed in a variety of tumor types, including colorectal, non–small-cell lung, breast, bladder, and prostate cancers making this pathway a drug target for cancer treatment and prevention [105,106]. The molecular and functional consequences of COX-2 overexpression in neoplasia are not yet entirely elucidated. Increased expression of COX-2 correlates with increased angiogenesis, decreased apoptosis, increased tumor invasiveness, and immunosuppression in a range of cancers [107]. Drugs that specifically target COX2 reduce the risk of PCa in human [108]. Soy isoflavones, including genistein, are negative regulators of prostaglandin synthesis and signaling, as shown by a study using LNCaP and PC-3 cells, as well as primary PCa cells [23]. In these cells, 10 μmol/L of genistein suppressed COX2 mRNA and protein levels and reduced the expression of the prostaglandin receptors EP4 and FP. In addition, genistein induced mRNA levels of 15-hydroxy prostaglandin dehydrogenase, the enzyme that inactivates prostaglandins, which resulted in a significant reduction of secreted prostaglandin-E2 in these cells. A combination of calcitriol and low doses of genistein (10μmol/L) has been shown to act additively to suppress the prostaglandin pathway in PC-3 cells [109]. In PCa patients, soy isoflavones significantly decreased prostate COX-2 mRNA expression [23] suggesting a direct negative regulation of prostaglandin metabolism and signaling by soy isoflavones. These data demonstrate that soy isoflavones, at physiological doses, reduce PCa progression through inhibiting prostaglandins pathway. Still, many gaps exist in our understanding of this chemopreventive mechanism which needs further elucidation.
1.6.4 Cancer stem cells as targets of soy isoflavone action
Most solid tumors have now been reported to contain stem cell-like cells called cancer stem cells (CSCs). These cells possess characteristics associated with normal stem cells, specifically the ability to give rise to all cell types found in the cancer sample. CSCs have the potential of self-renewal and differentiation into different cell phenotypes. Such CSCs persist in tumors as a distinct cell population upon therapy-induced tumor regression and following relapse giving rise to new tumors and metastases. Therefore, CSCs have been attractive targets for cancer prevention and treatment [110,111].
A strong body of evidence supports the existence of stem cells (SCs) in the normal prostate, as has been shown by John Isaac’s classic androgen cycling experiments, using several rounds of castration-induced regression and testosterone-induced regeneration in the adult rodent prostate to identify the existence of a small population of SCs that have the capacity for both self-renewal and differentiation [112]. In prostate cancer, populations of prostate stem/progenitor cells that express high levels of stem cell markers, such as CD44, integrin α2β1, or CD133, have been detected [113]. Interestingly, two recent independent studies on the mouse prostate have identified two distinct populations of SCs. One population expresses CD117 (c-Kit) and is localized in the basal layer [114], and the other population expresses Nkx3.1 and is located in the luminal layer [115]. Using different prostate cancer cell xenografts (DU145, LAPC-4, and LAPC-9), Patrawala et al. have shown that the CD44 positive cell population is enriched in prostate CSCs [116].
Most of the data exploring the molecular role of soy isoflavones has been collected from in vitro cell culture systems, which utilized cancer cell lines or primary cell cultures. Only recently have researchers begun to assess the impact of soy isoflavones directly on CSCs. McCubrey et al. [117] identified cancer initiating cells that have stem cell characteristics from PC-3 cell line by culturing these cells in media containing 25 nmol/L of the chemotherapeutic drug doxorubicin, a method that had been established and published earlier by the same research group [118]. They showed that while PC3 cells were highly sensitive to docetaxel compared with the PC3-derived cancer initiating cells, they did not display significant differences in the sensitivity to genistein which was able, at low physiologic doses, to reduce the IC50 of PC3-derived cancer initiating cells for docetaxel by approximately 200-fold. Interestingly, Ouchi and colleagues [119] found that human telomerase reverse transcriptase (hTERT) mRNA levels in LNCaP cells and hTERT transcriptional activity in DU145 cells were suppressed in response to genistein treatment. hTERT is the catalytic subunit of telomerase that marks a slowly cycling population of stem cells [120]. Thus, soy isoflavones may suppress the expansion of this cell population and protect them from future cancer-causing gene mutations. Research on the impact of soy isoflavones on adult and cancer stem cells is certain to expand in the future.
1.7. Epigenetic effects of soy isoflavones
Epigenetic mechanisms play tremendously important regulatory roles in the expression of genes associated with PCa. These mechanisms include DNA methylation, histone modifications, and regulatory small RNA molecules (miRNA). These epigenetic changes can be modulated by various factors, including dietary components. Soy isoflavones are among these dietary elements that possess potential anticancer epigenetic activity [121].
1.7.1 DNA methylation
DNA methylation is an essential regulator of gene transcription and a large body of evidence has demonstrated that aberrant DNA methylation can result in an unscheduled gene silencing since genes with high levels of 5-methylcytosine in their promoter region are transcriptionally silent. Aberrant DNA methylation patterns have been associated with a large number of human malignancies. Hypermethylation is one of the critical epigenetic modifications that repress transcription of tumor suppressor genes.
A number of studies support the notion that genistein can reverse promoter hypermethylation of tumor suppressor genes in PCa cells. Adjakly et al. [122] showed that soy phytoestrogens; 40 μmol/L of genistein and 110 μmol/L of daidzein, reduced the methylation of breast cancer 1 (BRCA1), glutathione S-transferase P1 (GSTP1) and ephrin B2 (EPHB2) genes in DU-145 and PC-3 cells. This effect on promoter methylation of these tumor suppressor genes accompanied an increase in their protein expression. Genistein also reversed promoter hypermethylation of some other tumor suppressor genes, including Ras association domain family 1 (RASSF1A) [123], B-cell translocation gene 3 (BTG3/ANA/APRO4) [124], PTEN, cyclin D, p53, FOXO3a [125], p21 (WAF1/CIP1) and p16 (INK4a) [125]. Some of these studies showed that this effect of genistein on reducing promoter methylation of tumor suppressor genes was linked to a reduction of the expression and/or the activity of DNA methyltransferase enzymes, and methyl-binding domain proteins [124,126]. Collectively, these data demonstrate that soy isoflavones are capable of reversing promoter hypermethylation in some tumor suppressor genes in PCa cells; however, most of these studies used very high pharmacological doses. Thus, further investigations of these effects using physiological concentrations are required.
1.7.2 Histone modifications
Acetylation of the lysine residues at the N terminus of histone proteins removes positive charges, thereby reducing the affinity between histones and DNA. This allows easier access of RNA polymerase and transcription factors to the promoter region. Therefore, in most cases, histone acetylation enhances transcription, while histone deacetylation represses transcription. Histone acetylation and deacetylation are catalyzed by histone acetyltransferases and histone deacetylases, respectively [127].
Kikuno et al. [125] showed that genistein-induced expression of tumor suppressor genes PTEN, cyclin D, p53 and FOXO3a in both LNCaP and PC-3 PCa cells were mediated by modulation of histone H3-Lysine 9 (H3-K9) methylation and deacetylation. H3-K9 promoter methylation or deacetylation induced by genistein led to inhibition of the AKT signaling pathway. Studies that discuss effects of genistein on histone modifications are limited which opens the window for research to explore this new epigenetic chemopreventive action.
1.7.3 Regulation of mi-RNA
MicroRNAs (miRNA/miR) are a class of short (18–25 nucleotides in length), noncoding RNA that binds mRNAs with complementary sequences and causes translational repression or target degradation and gene silencing. MiRNA expression is frequently altered in cancer in which they can act as either oncogenes (when they target tumor suppressor genes) or as tumor suppressors (when they target oncogenes). The expression of several miRNAs is disrupted in PCa, which may contribute to the initiation and progression of this cancer. Examples of these miRNA are miR-221, miR-222, miR-488, miR-146a, miR-331-3p, miR-21, miR-15a, miR-16-1, miR-125b, miR-145, and let-7c (a comprehensive report has been recently published by Catto et al. [128]). Genes encoding for miRNA can serve also as targets for cancer-induced epigenetic modifications such as promoter methylation and histone modifications [129]. Several studies have identified miRNA as a significant target for cancer treatment and/or chemoprevention. Soy isoflavones, genistein and daidzein at doses of 40 and 110 μmol/L, respectively regulate the expression of certain miRNAs in PC-3, DU145, and LNCaP PCa cells, most of which play a vital role in cell growth and survival [130]. Interestingly, these changes in miRNA expression by soy isoflavones were similar to those obtained by the demethylating agent 5-azacytidine, which suggests that the regulatory effect of soy isoflavones on miRNA expression was mainly mediated by their ability to reverse the promoter methylation of the epigenetically suppressed miRNA. The ability of soy isoflavones to regulate miRNAs was supported by findings of Li et al. [129] showing that soy isoflavones can reduce methylation of the promoter sequence of miR-29a and miR-1256, leading to upregulation of expression of these miRNAs and subsequent degradation and gene silencing of two oncogenes, tripartite motif-containing protein 68 (TRIM68) and phosphoglycerate kinase 1 (PGK-1), which was associated with inhibition of PCa cell growth and invasion. These studies demonstrate a new role of soy isoflavones on suppressing PCa through inhibiting oncogenic miRNAs.
1.8. Non-epithelial cells as targets of soy isoflavones mediated-cancer prevention
Although most cell-based soy isoflavone research has focused upon the impact of isoflavones on either tumor cells or their non-neoplastic progenitors, there is also evidence showing that other cells that exist in the tissue or tumor microenvironment are targets of soy isoflavones action. For example, in the prostate, the communication between epithelial cells and the stromal cells surrounding them is crucial for the growth and progression of cancer in that organ. In the next section, we will discuss two additional examples of non-epithelial cell targets of soy isoflavones: vascular cells and immune cells.
1.8.1 Inhibition of angiogenesis
Tumor angiogenesis is a pathological process through which cancer cells induce new blood vessel formation at the tumor site which is essential for the expansion of tumor growth and tumor cell metastasis. A large number of molecules are involved throughout the process of induction of angiogenesis in cancer, including cytokines and growth factors, such as VEGF (vascular endothelial growth factor), bFGF (basic fibroblast growth factor), and PDGF BB (platelet-derived growth factor BB homodimer), expressed on cancer cells, stromal cells, and tumor-associated macrophages [131].
Soy isoflavones can inhibit the development of tumor angiogenesis. This is mediated by effects exerted on either endothelial or epithelial cells in the tumor microenvironment. Ambra et al. [132] have demonstrated that genistein at a low physiological dose (2.5 μmol/L) down regulated the mRNA level of VEGF receptors 1 and 2 (VEGFR1 and VEGFR2) in the primary human endothelial cells. Genistein also directly inhibits the proliferation of endothelial cells and stops endothelial cell sprouting and elongation induced by VEGF [133]. In addition to such a direct role of genistein signaling in endothelial cells, Guo et al. [134] found that genistein (5–50μmol/L) caused a significant inhibition of both the basal and the hypoxia-stimulated VEGF expression in the human PCa cells, PC-3. This effect was mediated partly by the ability of genistein to reduce nuclear accumulation and activity of the transcription factor hypoxia-inducible factor-1α (HIF-1α) that regulates VEGF expression in response to hypoxia. Moreover, treatment of PC-3 cells with soy isoflavones caused a downregulation of the Apurinic apyrimidinic endonuclease redox effector factor-1 (APE1/Ref-1), which is a multifunctional protein that is responsible for redox-activation of HIF-1α [135,136]. In androgen-independent PC-3 and C4-2B PCa cells transfected with APE1/Ref-1 cDNA, radiation caused a greater increase in HIF-1α and NF-κB activities but this effect was inhibited by pretreatment with genistein (60μmol/L), daidzein (30μmolL) or a combination of soy isoflavones prior to radiation [137].
Tumors from subcutaneously inoculated LNCaP cells in severe combined immune-deficient mice had reduced growth, less vascular volume, and reduced microvessel density in soy rich diet fed mice than mice fed on control diet [138]. Additionally, soy isoflavones reduced the mRNA level of the pro-angiogenic cytokine interleukin-8 (IL-8) in PC-3 cells [139]. IL-8 is a potent pro-angiogenic factor and induces the metastatic potential of PCa as well [140]. Also, it has been shown that the expression of IL-8 is directly correlated with the Gleason score and pathologic tumor stage and could distinguish organ-confined from nonconfined tumors [141,142]. In two independent global gene expression analysis studies, genistein and daidzein down-regulated a set of genes that are necessary for the angiogenesis pathway in LNCaP, PC-3, and DU-145 PCa cells. These genes included platelet-derived endothelial cell growth factor (ECGF1), fibroblast growth factor 1 (FGF1), IGF1, fibroblast growth factor receptor 3 (FGFR3), IL-1β, IL-6, IL-8, platelet/endothelial cell adhesion molecule (CD31 antigen or PECAM1), CXCL10, and many other molecules that are essential to angiogenesis [53,139]. These data support the anti-angiogenic properties of soy isoflavones in both androgen dependent and independent PCa. Although some of these studies used high pharmacological doses of soy isoflavones [137,139], others used a wide range of doses that cover the physiologically attainable range [134].
1.8.2 Inhibition of metastasis
Cancer metastasis poses the biggest obstacle to treatment of the disease and is the leading cause of death in PCa patients. Cancer metastasis is a multistep process involving the spread of malignant cells from a primary tumor to remote sites. This process includes epithelial to mesenchymal transition (EMT), degradation of the extracellular matrix (ECM), invasion of surrounding tissues, cell migration, anchorage-independent growth, apoptosis evasion, intravasation into existing and newly formed blood and lymph vessels, transportation through the vessels and extravasation, mesenchymal to epithelial transition (MET), establishment of surviving cancer cells at distant sites, and outgrowth of secondary tumors [143].
The effect of soy isoflavones on the metastatic properties of PCa cells has been tested both in vitro and in vivo, and there are several proposed mechanisms for the anti-metastatic effects of isoflavones. One of these mechanisms is the ability of soy isoflavones to reverse EMT. Genistein (15 μmol/L) suppresses the invasive growth of IA8-ARCaP human PCa cells through the reversal of EMT [144]. IA8-ARCaP is a highly invasive clone of the parental ARCaP PCa cell line which can undergo EMT [145]. This reversal of EMT by genistein has been verified by cell morphological changes and changes in the expression of EMT-related markers, including reduced expression of E-cadherin and increased expression of vimentin, fibronectin, N-cadherin, Twist, and Snail. Another proposed mechanism for the impact of genistein on metastasis is by inhibiting matrix metalloproteinases (MMPs). MMPs are endopeptidases that degrade the extracellular matrix and basement membrane of cells, which makes them key factors in tumor invasion and metastasis. Genistein (0.5–50 μmol/L) induced a dose-dependent suppression of the total activity of MMPs in both PC3 and LNCaP cell lines [146]. A number of in vitro and animal studies support the notion that genistein down regulates MMPs, reducing the invasiveness of PCa cells [64,65,147]. This effect is mediated by blocking the action of focal adhesion kinase (FAK), p38 MAPK, and heat shock protein 27 (HSP27) by genistein [148]. FAK mediates cell adhesion-associated signals in human PCa [149] while p38 MAPK is required for phosphorylation of HSP27, which in turn is essential for MMP-2 activation, and PCa cell invasion [150].
Genistein has also been shown to induce a dose-dependent inhibitory effect on osteopontin (OPN) mRNA levels in transgenic TRAMP mice with prostate adenocarcinoma [151] and in PC-3 cells [152]. OPN is a protein that is implicated in bone remodeling and highly expressed in PCa cell lines and human PCa specimens. The malignant and metastatic phenotypes of PCa correlate with elevated OPN expression, which suggests that OPN has a role in PCa growth, invasion, and metastasis [153]. These findings point to inhibition of OPN expression as another mechanism by which genistein may exert its anti-metastatic effect.
Cell motility studies have shown that genistein reduces the homo-and heterotypic contact stimulated migration of PC-3 cells [154] as well as the rat prostate carcinoma cell lines, MAT-LyLu, and AT-2 [155]. In addition, dietary supplements of soy protein inhibited metastases to lymph nodes and lungs in male SCID mice orthotopically implanted with LNCaP cells [156]. In summary, soy isoflavones demonstrated anti-metastatic effects against PCa. Proposed mechanisms of this effect include reversal of EMT, inhibition of MMPs and OPN and suppression of cell motility and migration. Most of the studies we reviewed have used low physiological ranges of soy isoflavones.
1.8.3 Regulation of immune cell function by soy isoflavones
The role of the immune system in the context of cancer is complicated. Inflammation was initially believed to be a host response against cancer which leads to cancer suppression, but host immune cells in the tumor microenvironment can also provide the tumor with cytokines and growth that promote tumor growth angiogenesis and progression and lead to unfavorable prognosis. Thus, although therapeutic or prophylactic agents that restrain the inflammation in cancer patients may contribute to tumor regression and improve prognosis, there may be inadvertent negative effects, as well.
Tumor associated macrophages (TAMs) can release a broad range of growth factors, proteolytic enzymes, cytokines, and inflammatory mediators. Many of these factors are key agents in cancer metastasis. TAMs represent one of the targets for the anticancer effects of genistein on PCa. Genistein reduced the number of TAMs and subsequently, the tumor vessel density and the tumor size in rats which were injected subcutaneously with the androgen-independent, anaplastic, and highly metastatic rat PCa cell line, R3327 MAT-Lu. This effect was mediated by the ability of genistein to inhibit TNF-α and granulocyte monocyte-colony stimulating factor [157]. Another possible mechanism by which genistein can interfere with the inflammation pathway is by blocking interleukin-10 (IL-10) signaling in PCa cells [158]. IL-10 is an inflammatory cytokine that is produced primarily by monocytes and to a lesser extent by lymphocytes.
Despite the documented evidence that soy isoflavones have anti-inflammatory effects in many diseases, including but not limited to colitis [159], steatohepatitis [160], chronic renal disease [161], and cancers such as skin cancer [162], the impact of soy isoflavones on regulation of immune response and inflammation in PCa has not been consistent. Genistein treatment led to an increase in radiation-induced giant cells, inflammatory cells, and fibrosis that was observed in nude mice injected orthotopically with PC-3 cells [163]. Furthermore, oral administration of genistein and daidzein rich diet induced inflammation in the rat prostate [164]. Because of these observations, it remains to be determined how soy isoflavones can influence the immune cell function in PCa.
1.9. Increase sensitivity to radiotherapy and chemotherapy
In locally advanced stages of PCa, the recommended treatment is radiotherapy combined with simultaneous application of LHRH-agonists [165]. Genistein at a physiologically achievable dose (10 μmol/L) increased low-dose radiosensitivity in LNCaP cells, expressed as a decreased colony formation and survival and an increased proportion of cells arrested in G2/M phase of the cell cycle [166]. Consistent with these findings, Hillman et al. [167] showed that a similar range of genistein doses caused an inhibition in DNA synthesis, cell growth, and colony formation and enhanced the effect of low dose photon or neutron radiation in PC-3 cells. Consistent with these findings, two separate studies using the PC-3 orthotopic metastatic mouse model indicated that soy isoflavones potentiated the effect of irradiation on inhibiting the primary prostate tumor growth and metastasis [168].
A combination treatment with genistein and perifosine, a chemotherapeutic agent, boosted the inhibitory action of perifosine on the clonogenic potential of PCa cell lines such as LNCaP, LNCaP-R273H, C4-2, Cds1, and PC3. This was evident by a profound reduction in the Akt activity and the sub-G1 cell population indicating apoptosis, as well as suppression of p53 and p21protein expression [81]. Similarly, genistein enhanced the cytotoxic effects of the chemotherapeutic agents, taxol (paclitaxel) [169] and docetaxel and the antiandrogen bicalutamide in human PCa cells [147,170]. All the above-mentioned findings support a potential role of soy isoflavones in enhancing the therapeutic effect of radiation and chemotherapy in PCa.
1.10. Soy isoflavones action may not be uniform across all stages of PCa
Although we focus on the effect that soy isoflavones have on the development of PCa, it is just as crucial to understand the impact that PCa has on soy isoflavones activity. In this section, we will review studies that have demonstrated effects of soy isoflavones on steroid hormone receptors (AR and ER), and how the protective effects of soy isoflavones can be reversed in advanced stages and/or hormonally pretreated PCa, in which mutations can take place in these receptors as a consequence of genomic instability or natural selection.
1.10.1 Androgen receptor
In 1940, Dr. Charles Huggins [171] was the first to recognize that PCa is an androgen-sensitive disease through his pioneering work on castration in the treatment of PCa. This discovery was followed by several studies that confirmed the essential role of androgen in the development and progression of PCa. Many therapeutic modalities are directed toward improved androgen ablation for locally advanced or metastatic PCa and for cases that recur after radical prostatectomy and radiotherapy. Most of the patients who receive androgen ablation therapies show initial clinical and biochemical evidence of improvement (decrease in serum PSA). However, most of those patients relapse with a more aggressive, hormone refractory form of PCa, which is independent of circulating androgen but still dependent on the functional AR for growth and progression.
Although the role of AR in the pathogenesis of PCa development and progression was discovered by Charles Huggins in 1940, it was quite recently that investigators have recognized the role of AR in hormone refractory PCa [172–174]. In the hormone refractory PCa, the AR activity despite the absence of androgens takes place through several mechanisms: androgen receptor gene amplification [175,176], altered expression and function of AR coactivators [177,178] and ligand independent AR activation through stimulation of other signal transduction pathways [179,180]. AR can also obtain mutations that make the receptor either hypersensitive or expand its ligand specificity as those occur in the LBD. These mutations may enable the receptor to be activated by a broad variety of steroids such as estrogens, progestins, adrenal steroids, or even antiandrogens. These mutant receptors with enlarged spectrum to various steroids or synthetic analogs have been called “promiscuous”.
Cumulatively, these data suggest that AR plays a pivotal role in both androgen-dependent and androgen-independent PCa and that it is the principal focus for PCa prevention and treatment. Epidemiological studies suggest an inverse association between soy intake and PCa risk. However, human data regarding the implementation of isoflavones on local AR expression in PCa tissue are remarkably little. Indirect evidence of the effect of isoflavones on AR has been obtained through clinical studies that demonstrated the effects of isoflavones on serum levels of prostate-specific antigen (PSA) in PCa patients. PSA is a clinically relevant AR-responsive gene, which is used to monitor treatment response, prognosis, and progression in patients with PCa [181]. AR regulates PSA transcription via binding to the AR responsive element (ARE) in the promoter region of PSA [182].
Dietary genistein downregulated AR in the prostate of rats fed a soy diet that adjusted genistein concentrations to be comparable to those found in human consuming soy [183]. Similar results have been shown in TRAMP mice [184]. In vitro studies also demonstrated that genistein downregulates AR protein and mRNA expression and transcriptional activity [185]. In addition, genistein activates HDAC6-Hsp90, which mediates AR protein degradation [186]. The ability of genistein to regulate the AR-mediated pathway is considered the most pertinent chemopreventive mechanism for genistein. A microarray analyses for global gene expression in human PCa cells exposed to genistein showed a significant inhibition of genes in the AR pathway [187]. Comparable results have been obtained using glyceollins which is a soy-derived phytoalexins [188]. It has been proposed that the effect of genistein on AR expression is mediated by ER-β [189], and this was evident by the abolishment of that effect when cells were pretreated with the ER-β antagonist (ICI 164 384). These results provide evidence to support the inhibitory effect of genistein on AR expression and/or transcriptional activity. However, some in vitro studies demonstrated stimulatory effects of isoflavones particularly genistein at certain low doses [190–192]. The cause of these controversial results is not clear. Nevertheless, the use of different types of PCa cells with diverse molecular characterization could be the reason for these mixed results. One of the widely used in vitro cell models, LNCaP, has a Threonine to Alanine (T877A) mutation in the ligand binding domain (LBD) of the AR which broadens their binding capacity to cover most of the steroidal structures. This cell line has been used in the above referred studies that encountered stimulatory effects of genistein on AR expression and transcriptional activity. Genistein has a steroid like structure and could be recognized by the mutant AR as a ligand, which could explain the stimulatory response encountered in LNCaP cells to genistein treatment. Data from our recently published report [41], which strongly supports this hypothesis, demonstrate that genistein exerts a pleiotropic effect on PCa cell proliferation and AR activity depending on the AR status of these cells. It inhibited in a dose-dependent manner cell proliferation and AR expression in LAPC-4 cells that have wild-type (WT) AR. However, in LNCaP cells, lower doses of genistein exerted growth stimulatory effects and enhanced AR expression.
Accordingly, it became imminent to search for a reliable in vitro model for better verification of the AR mediated effects of genistein on PCa. This model should include PCa cells that are hormone naive and expressing wild type androgen and estrogen receptors. Nevertheless, we must value the utilization of the currently used models such as LNCaP cells to identify any potential harmful effects of genistein exposure on patients with late stage or hormonal pretreated PCa who are more prone to have AR mutations which could cause alteration of their response to genistein or other soy isoflavones.
1.10.2 Estrogen receptor
Estrogen is an effective hormonal therapy for androgen-dependent PCa. Paradoxically, estrogens might be involved in the development and progression of PCa [193]. The effects of estrogen were once considered to be mediated by ER-α only. Now, the relatively newly discovered ER-β provided a different insight for understanding the mechanism of action of estrogen in prostate. In normal prostate, ERα is expressed in stromal cells and the basal cell layer, while ERβ is predominately expressed in luminal cells. These receptors undergo an apparent remodeling during PCa development and progression either in their level of expression or cellular distribution [194–197].
ER-α expression is upregulated and shifted to the epithelial cells in high-grade prostatic intraepithelial neoplasia (HGPIN), and most likely mediates the effects of estradiol in promoting prostate carcinogenesis as demonstrated in animal models [197]. This role of ER-α has been supported by the ability of the ER-α antagonist toremifene to reduce the incidence of PCa in men with HGPIN [198]. In two animal models, Sprague-Dawley rats and TRAMP mice, genistein rich diet reduced the expression of ER-α, and its responsive gene, progesterone receptor (PR) [183,184]. There is a significant potential in the use of ER-α antagonists to prevent PCa and delay disease progression. However, there is a lack of studies that examined the effectiveness of soy isoflavones in modifying ER-α expression in PCa.
ER-β gene is expressed in prostate epithelial cells and has a role in the cellular homeostasis that is anti-proliferative [199], pro-differentiative [200], and pro-apoptotic [201]. Most studies on ER-β expression reported that levels declined in localized PCa with increasing grade from PIN through low to high Gleason scores [202,203]. This expression pattern supports the tumor suppressor role of ER-β. ER-β is differentially expressed in normal tissue and PCa [202]. Therefore, ER-β agonists might be a potential chemopreventive and therapeutic agents for treatment of PCa.
Soy isoflavones have stimulatory effects upon the expression of ER-β. Combined dietary soy isoflavones genistein, quercetin, and biochanin-A, at their physiologically achievable concentrations exert a synergistic effect on reducing PCa cell proliferation. This effect was mediated by modulation of genes that control cell cycle and apoptosis in LNCaP cells. The combination use of these agents led to up-regulation of ER-β mRNA expression more effectively than individual use [83]. Although the affinity of soy isoflavones is much higher for ER-β than for ER-α, it is possible the lack of ER-β expression or the unbalanced ratio of ER-α/ER-β could lead to modification of the expected effects of genistein in PCa. Similarly, Single Nucleotide Polymorphisms (SNPs) or mutations in ER-α or ER-β can modulate the agonistic potential of soy isoflavones to the receptor and subsequently, alter the chemopreventive effects of these isoflavones. Examples of these SNPs that distorted the association between soy isoflavones consumption and risk of PCa are those shown by Sonoda et al. [204] in Japanese population. They found that TC or CC alleles of rs2077647 in ER-α showed a high risk of PCa despite intake of greater than or equal to 60 mg isoflavones/day. However, the CC alleles showed a lower risk despite intake of less than 60mg isoflavones/day. A role of ER-β gene polymorphisms in modifying the association between isoflavone intake and breast cancer risk has been demonstrated by Iwasaki et al. [205]. This role needs to be investigated in PCa in order to identify any genetic variations that may influence the cancer-preventive effects of soy isoflavones.
2. Conclusion
There is a fair amount of population-based evidence showing that soy isoflavones can protect against PCa. Unfortunately, the mechanisms supporting this association have not yet been resolved. Nonetheless, mechanistic explanations for how soy isoflavones can reduce cancer risk or disrupt the biology of transformed cells are increasing. Although the role of soy isoflavones in PCa has traditionally been linked with the suppression of proliferation and the induction of apoptosis, recently there is a compelling evidence that soy isoflavones regulate other cancer-related cellular processes. For example, a reasonable number of reports suggests that the antioxidant actions of soy isoflavones are worth exploring for cancer prevention. The breadth of these potential mechanisms by which soy isoflavones prevent or suppress cancer is given in table 2. Still, many gaps remain beyond understanding of how soy isoflavones regulates cell biology relevant to PCa prevention and treatment. Most of the studies performed to assess the protective effects of soy isoflavones on PCa have used cells with mutant AR such as LNCaP, or cells that are devoid of AR such as PC-3 and DU-145. In addition, most of these studies used high pharmacological doses of genistein that do not reflect what can be achieved by people consuming soy rich diet. Similarly, there are few controlled animal studies that warrant the validity of proposed molecular mechanisms for soy isoflavone mediated cancer prevention using the ranges of soy isoflavones concentrations that are observed in human populations. Therefore, further studies must be conducted to determine if these mechanisms are different within the context of wild type AR and low physiological concentrations. Recent advances in global profiling of transcription factor-binding sites with techniques such as ChIP-seq should overcome this uncertainty, and help researchers to examine the differential effects of these isoflavones in the context of these pervasive AR mutations. Even with all of these gaps in our understanding, this is an unprecedented opportunity for research to explore mechanisms of action of soy isoflavones in preventing PCa.
Table 2.
The Molecular Mechanisms of the Chemopreventive Effects of Soy Isoflavones in Prostate Cancer.
| Molecular Event | Target Genes/Proteins | Soy Isoflavones | Invitro/invivo System |
|---|---|---|---|
| Cell-cycle regulators | p21, p27, p53, cyclin B1, cyclin B2*, cyclin D1, Cyclin E, CDK2, CDK4, CDK7*, FOXO1, MAPK, CDKI-1A*, CDKI-2A*, CHEK2* | Genistein, daidzein and equol | LNCaP, PC-3, DU-145, LAPC-4 |
| IGF-1 (Insulin-Like Growth Factor-1) signaling | p-IGF-1R, p-Src, p-Akt, p-GSk-3β, IRS-1 | Genistein, daidzein and equol | PC-3, AT6.3, LNCaP and DU-145 |
| TGFβ (transforming growth factor β) signaling pathway | TGFβ, MAPK-Activated Protein Kinase 2*, p38 MAPK, HSP27*, MMP2 | Genistein | PC3 and PC3-M |
| Wnt/β-catenin signaling | β-catenin, E-cadherin | Genistein | PC-3 |
| Tyrosine kinase inhibition | p-EGFR*, p-Akt*, p-GSK-3β*, mTOR-p70S6k*, p-FOXO3a*, p-Src* | Genistein, daidzein and glycetin | LNCaP, LNCaP-p53(GOF), 22Rv1, and C4-2B |
| Apoptosis | PTEN, Bcl-2*, Bax*, caspase-3*, survivin, elafin, PAR-2, NFκB, proteasome* | Genistein, daidzein and glycetin | LNCaP, PC-3, DU145 and BPH-1 |
| cell differentiation | cytokeratin 18*, cytokeratin 5* and P63* | Genistein, daidzein, glycetin and equol | RWPE-1 cells line and TRAMP mouse model |
| Autophagy | mTOR-p70S6k* | Genistein | LNCaP and 22Rv1 |
| Antioxidant defense and DNA repair | (GPx)-1*, glutathione reductase, microsomal glutathione S-transferase 1, metallothionein, MnSOD, catalase, GADD45*, p53 | Genistein | LNCaP, PC-3, LAPC-4 and DU-145 |
| Prostaglandin metabolism and action | PGE(2), COX2, prostaglandin receptors EP4 and FP, PGDH (hydroxyprostaglandin dehydrogenase) | Genistein | LNCaP and PC-3 |
| Cancer stem cells | hTERT (human telomerase reverse transcriptase) | Genistein | PC3/(CIC), LNCaP and DU-145 |
| DNA methylation | BRCA1*, GSTP1*, EPHB2*, RASSF1A*, (BTG3/ANA/APRO4), PTEN*, CYLD*, p53*, FOXO3a*, p21* (WAF1/CIP1/KIP1)*, p16* INK4a*, DNMT and methyl-binding domain proteins | Genistein and daidzein | LNCaP, DU-145, DuPro and PC-3 |
| Histone modifications | H3-K9*, Akt*, PTEN*, Cyclin D*, p53* and FOXO3a* | Genistein | LNCaP and PC-3 |
| Regulation of mi-RNA | miR-29a and miR-1256 | Genistein and daidzein | LNCaP, VCaP, PC-3, C4–2B, ARCaP cell lines and in human tumor tissues |
| Inhibition of angiogenesis | VEGF, HIF-1, NF-kappaB, angiopoietin 2 (ANGPT2), PD-ECGF*, IL6*, IL-8*, endoglin*, CD31*, FGF1*, IGF1*, FGFR3*, PECAM1*, CXCL10* | Genistein and daidzein | LNCap, PC-3, DU-145 and C4-2B cell lines and LNCaP xenograft mouse model |
| Inhibition of metastasis | MMP-2, Focal adhesion kinase (FAK), p38 MAPK, and HSP27, Osteopontin (OPN)* | Genistein | PC3 and LNCaP cell lines, TRAMP mouse model and LNCaP xenograft SCID mouse model |
| Inhibition of EMT | E-Cadherin, N-Cadherin, Vimentin, Fibronectin, Twist, Snail | Genistein | IA8-ARCaP |
| Regulation of immune cell function | TNF-α*, granulocyte monocyte-colony stimulating factor (GM-CSF)*, IL-10 | Genistein | PC-3/PI cell line and R3327 MAT-Lu xenograft rat model |
| Increase sensitivity to radio and chemotherapy | Akt*, p53*, p21*, PARP*, Bcl-xL*, Bax*, survivin*, HIF1-α* | Genistein | LNCaP, PC-3, LNCaP-R273H, C4-2 and Cds1 cell lines and PC-3 orthotopic metastatic mouse model, |
| Androgen receptor mediated pathway | AR, PSA, HDAC6-Hsp90 | Genistein | LNCaP cell line, TRAMP mouse model and rat model |
| Estrogen receptor mediated pathway | ER-α, ER-β | Genistein | LNCaP and PC-3 cell lines, Sprague-Dawley rats and TRAMP mice |
The asterisk indicates that high pharmacological doses were applied to obtain the effect on target genes/proteins.
Highlights.
We reviewed of all anti-carcinogenic effects of soy isoflavones in prostate cancer
We discussed the traditional growth and apoptotic pathways in prostate cancer cells
Other new-fangled mechanisms were discussed e.g. autophagy, telomerase, epigenetics
Effect on cancer milieu e.g. immune, stem and endothelial cells were also discussed
These are discussed along with considerations about doses and preclinical models
Acknowledgments
This work was supported by Grant No. CA 116195, Department of Pathology at University of Illinois at Chicago and by Grant No. GM 0842 from the Egyptian Ministry of Higher Education.
Abbreviations
- APC
Adenomatous Polyposis Coli
- APE1/Ref
Apurinic Apyrimidinic Endonuclease Redox Effector Factor
- APK
Activated Protein Kinase
- BTG
B-cell translocation gene
- CSC
Cancer Stem Cells
- CSF
Colony-Stimulating Factor
- DSH
Disheveled
- ECGF1
Endothelial Cell Growth Factor
- EPHB
Ephrin B
- FAK
Focal Adhesion Kinase
- GADD
Growth Arrest and DNA Damage
- GM-CSF
Granulocyte Monocyte-Colony Stimulating Factor
- GPx
Glutathione Peroxidase
- GSk-3β
Glycogen synthase kinase-3β
- GSK3β
Glycogen Synthase kinase 3β
- GSR
Glutathione Reductase
- GSTP
Glutathione S-Transferase P
- HAT
Histone Acetyltransferases
- LEF
Lymphoid Enhancer Factor
- LHRH
Luteinizing Hormone Releasing Hormone
- RASSF
Ras Association Domain Family
- TAMs
Tumor Associated Macrophages
- TRIM
Tripartite Motif-Containing Protein
- uPA
Urokinase-Type Plasminogen Activator
- z-VAD-fmk
N-benzyloxycarbonyl-Val-Asp-fluoromethyl-ketone
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Siegel R, Desantis C, Virgo K, Stein K, Mariotto A, et al. Cancer treatment and survivorship statistics, 2012. CA Cancer J Clin. 2012;62:220–241. doi: 10.3322/caac.21149. [DOI] [PubMed] [Google Scholar]
- 2.Jemal A, Bray F, Center MM, Ferlay J, Ward E, et al. Global cancer statistics. CA Cancer J Clin. 2011;61:69–90. doi: 10.3322/caac.20107. [DOI] [PubMed] [Google Scholar]
- 3.Kimura T. East meets West: ethnic differences in prostate cancer epidemiology between East Asians and Caucasians. Chin J Cancer. 2011 doi: 10.5732/cjc.011.10324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Adlercreutz H, Markkanen H, Watanabe S. Plasma concentrations of phyto-oestrogens in Japanese men. Lancet. 1993;342:1209–1210. doi: 10.1016/0140-6736(93)92188-y. [DOI] [PubMed] [Google Scholar]
- 5.Morton MS, Chan PS, Cheng C, Blacklock N, Matos-Ferreira A, et al. Lignans and isoflavonoids in plasma and prostatic fluid in men: samples from Portugal, Hong Kong, and the United Kingdom. Prostate. 1997;32:122–128. doi: 10.1002/(sici)1097-0045(19970701)32:2<122::aid-pros7>3.0.co;2-o. [DOI] [PubMed] [Google Scholar]
- 6.Hwang YW, Kim SY, Jee SH, Kim YN, Nam CM. Soy food consumption and risk of prostate cancer: a meta-analysis of observational studies. Nutr Cancer. 2009;61:598–606. doi: 10.1080/01635580902825639. [DOI] [PubMed] [Google Scholar]
- 7.Maskarinec G, Morimoto Y, Hebshi S, Sharma S, Franke AA, et al. Serum prostate-specific antigen but not testosterone levels decrease in a randomized soy intervention among men. Eur J Clin Nutr. 2006;60:1423–1429. doi: 10.1038/sj.ejcn.1602473. [DOI] [PubMed] [Google Scholar]
- 8.Adams KF, Chen C, Newton KM, Potter JD, Lampe JW. Soy isoflavones do not modulate prostate-specific antigen concentrations in older men in a randomized controlled trial. Cancer Epidemiol Biomarkers Prev. 2004;13:644–648. [PubMed] [Google Scholar]
- 9.Jenkins DJ, Kendall CW, D’Costa MA, Jackson CJ, Vidgen E, et al. Soy consumption and phytoestrogens: effect on serum prostate specific antigen when blood lipids and oxidized low-density lipoprotein are reduced in hyperlipidemic men. J Urol. 2003;169:507–511. doi: 10.1097/01.ju.0000046060.59113.57. [DOI] [PubMed] [Google Scholar]
- 10.Hamilton-Reeves JM, Rebello SA, Thomas W, Kurzer MS, Slaton JW. Effects of soy protein isolate consumption on prostate cancer biomarkers in men with HGPIN, ASAP, and low-grade prostate cancer. Nutr Cancer. 2008;60:7–13. doi: 10.1080/01635580701586770. [DOI] [PubMed] [Google Scholar]
- 11.Urban D, Irwin W, Kirk M, Markiewicz MA, Myers R, et al. The effect of isolated soy protein on plasma biomarkers in elderly men with elevated serum prostate specific antigen. J Urol. 2001;165:294–300. doi: 10.1097/00005392-200101000-00082. [DOI] [PubMed] [Google Scholar]
- 12.deVere White RW, Tsodikov A, Stapp EC, Soares SE, Fujii H, et al. Effects of a high dose, aglycone-rich soy extract on prostate-specific antigen and serum isoflavone concentrations in men with localized prostate cancer. Nutr Cancer. 2010;62:1036–1043. doi: 10.1080/01635581.2010.492085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Kumar NB, Cantor A, Allen K, Riccardi D, Besterman-Dahan K, et al. The specific role of isoflavones in reducing prostate cancer risk. Prostate. 2004;59:141–147. doi: 10.1002/pros.10362. [DOI] [PubMed] [Google Scholar]
- 14.Bosland MC, Kato I, Zeleniuch-Jacquotte A, Schmoll J, Enk Rueter E, et al. Effect of soy protein isolate supplementation on biochemical recurrence of prostate cancer after radical prostatectomy: a randomized trial. JAMA. 2013;310:170–178. doi: 10.1001/jama.2013.7842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Napora JK, Short RG, Muller DC, Carlson OD, Odetunde JO, et al. High-dose isoflavones do not improve metabolic and inflammatory parameters in androgen-deprived men with prostate cancer. J Androl. 2011;32:40–48. doi: 10.2164/jandrol.110.010983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.deVere White RW, Hackman RM, Soares SE, Beckett LA, Li Y, et al. Effects of a genistein-rich extract on PSA levels in men with a history of prostate cancer. Urology. 2004;63:259–263. doi: 10.1016/j.urology.2003.09.061. [DOI] [PubMed] [Google Scholar]
- 17.Kwan W, Duncan G, Van Patten C, Liu M, Lim J. A phase II trial of a soy beverage for subjects without clinical disease with rising prostate-specific antigen after radical radiation for prostate cancer. Nutr Cancer. 2010;62:198–207. doi: 10.1080/01635580903305318. [DOI] [PubMed] [Google Scholar]
- 18.Pendleton JM, Tan WW, Anai S, Chang M, Hou W, et al. Phase II trial of isoflavone in prostate-specific antigen recurrent prostate cancer after previous local therapy. BMC Cancer. 2008;8:132. doi: 10.1186/1471-2407-8-132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Grainger EM, Schwartz SJ, Wang S, Unlu NZ, Boileau TW, et al. A combination of tomato and soy products for men with recurring prostate cancer and rising prostate specific antigen. Nutr Cancer. 2008;60:145–154. doi: 10.1080/01635580701621338. [DOI] [PubMed] [Google Scholar]
- 20.Lazarevic B, Boezelijn G, Diep LM, Kvernrod K, Ogren O, et al. Efficacy and safety of short-term genistein intervention in patients with localized prostate cancer prior to radical prostatectomy: a randomized, placebo-controlled, double-blind Phase 2 clinical trial. Nutr Cancer. 2011;63:889–898. doi: 10.1080/01635581.2011.582221. [DOI] [PubMed] [Google Scholar]
- 21.Dalais FS, Meliala A, Wattanapenpaiboon N, Frydenberg M, Suter DA, et al. Effects of a diet rich in phytoestrogens on prostate-specific antigen and sex hormones in men diagnosed with prostate cancer. Urology. 2004;64:510–515. doi: 10.1016/j.urology.2004.04.009. [DOI] [PubMed] [Google Scholar]
- 22.Hussain M, Banerjee M, Sarkar FH, Djuric Z, Pollak MN, et al. Soy isoflavones in the treatment of prostate cancer. Nutr Cancer. 2003;47:111–117. doi: 10.1207/s15327914nc4702_1. [DOI] [PubMed] [Google Scholar]
- 23.Swami S, Krishnan AV, Moreno J, Bhattacharyya RS, Gardner C, et al. Inhibition of prostaglandin synthesis and actions by genistein in human prostate cancer cells and by soy isoflavones in prostate cancer patients. Int J Cancer. 2009;124:2050–2059. doi: 10.1002/ijc.24161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hamilton-Reeves JM, Rebello SA, Thomas W, Slaton JW, Kurzer MS. Isoflavone-rich soy protein isolate suppresses androgen receptor expression without altering estrogen receptor-beta expression or serum hormonal profiles in men at high risk of prostate cancer. J Nutr. 2007;137:1769–1775. doi: 10.1093/jn/137.7.1769. [DOI] [PubMed] [Google Scholar]
- 25.Joannou GE, Kelly GE, Reeder AY, Waring M, Nelson C. A urinary profile study of dietary phytoestrogens. The identification and mode of metabolism of new isoflavonoids. J Steroid Biochem Mol Biol. 1995;54:167–184. doi: 10.1016/0960-0760(95)00131-i. [DOI] [PubMed] [Google Scholar]
- 26.Takimoto CH, Glover K, Huang X, Hayes SA, Gallot L, et al. Phase I pharmacokinetic and pharmacodynamic analysis of unconjugated soy isoflavones administered to individuals with cancer. Cancer Epidemiol Biomarkers Prev. 2003;12:1213–1221. [PubMed] [Google Scholar]
- 27.Setchell KD, Faughnan MS, Avades T, Zimmer-Nechemias L, Brown NM, et al. Comparing the pharmacokinetics of daidzein and genistein with the use of 13C-labeled tracers in premenopausal women. Am J Clin Nutr. 2003;77:411–419. doi: 10.1093/ajcn/77.2.411. [DOI] [PubMed] [Google Scholar]
- 28.Gardner CD, Oelrich B, Liu JP, Feldman D, Franke AA, et al. Prostatic soy isoflavone concentrations exceed serum levels after dietary supplementation. Prostate. 2009;69:719–726. doi: 10.1002/pros.20922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Bloedon LT, Jeffcoat AR, Lopaczynski W, Schell MJ, Black TM, et al. Safety and pharmacokinetics of purified soy isoflavones: single-dose administration to postmenopausal women. Am J Clin Nutr. 2002;76:1126–1137. doi: 10.1093/ajcn/76.5.1126. [DOI] [PubMed] [Google Scholar]
- 30.Kuiper GG, Lemmen JG, Carlsson B, Corton JC, Safe SH, et al. Interaction of estrogenic chemicals and phytoestrogens with estrogen receptor beta. Endocrinology. 1998;139:4252–4263. doi: 10.1210/endo.139.10.6216. [DOI] [PubMed] [Google Scholar]
- 31.Morito K, Hirose T, Kinjo J, Hirakawa T, Okawa M, et al. Interaction of phytoestrogens with estrogen receptors alpha and beta. Biol Pharm Bull. 2001;24:351–356. doi: 10.1248/bpb.24.351. [DOI] [PubMed] [Google Scholar]
- 32.An J, Tzagarakis-Foster C, Scharschmidt TC, Lomri N, Leitman DC. Estrogen receptor beta-selective transcriptional activity and recruitment of coregulators by phytoestrogens. J Biol Chem. 2001;276:17808–17814. doi: 10.1074/jbc.M100953200. [DOI] [PubMed] [Google Scholar]
- 33.Zhao L, Mao Z, Brinton RD. A select combination of clinically relevant phytoestrogens enhances estrogen receptor beta-binding selectivity and neuroprotective activities in vitro and in vivo. Endocrinology. 2009;150:770–783. doi: 10.1210/en.2008-0715. [DOI] [PubMed] [Google Scholar]
- 34.Moiseeva EP, Almeida GM, Jones GD, Manson MM. Extended treatment with physiologic concentrations of dietary phytochemicals results in altered gene expression, reduced growth, and apoptosis of cancer cells. Mol Cancer Ther. 2007;6:3071–3079. doi: 10.1158/1535-7163.MCT-07-0117. [DOI] [PubMed] [Google Scholar]
- 35.Touny LH, Banerjee PP. Identification of both Myt-1 and Wee-1 as necessary mediators of the p21-independent inactivation of the cdc-2/cyclin B1 complex and growth inhibition of TRAMP cancer cells by genistein. Prostate. 2006;66:1542–1555. doi: 10.1002/pros.20495. [DOI] [PubMed] [Google Scholar]
- 36.Li Y, Bhuiyan M, Sarkar FH. Induction of apoptosis and inhibition of c-erbB-2 in MDA-MB-435 cells by genistein. Int J Oncol. 1999;15:525–533. doi: 10.3892/ijo.15.3.525. [DOI] [PubMed] [Google Scholar]
- 37.Ruiz-Larrea MB, Mohan AR, Paganga G, Miller NJ, Bolwell GP, et al. Antioxidant activity of phytoestrogenic isoflavones. Free Radic Res. 1997;26:63–70. doi: 10.3109/10715769709097785. [DOI] [PubMed] [Google Scholar]
- 38.Akiyama T, Ishida J, Nakagawa S, Ogawara H, Watanabe S, et al. Genistein, a specific inhibitor of tyrosine-specific protein kinases. J Biol Chem. 1987;262:5592–5595. [PubMed] [Google Scholar]
- 39.Li Y, Sarkar FH. Inhibition of nuclear factor kappaB activation in PC3 cells by genistein is mediated via Akt signaling pathway. Clin Cancer Res. 2002;8:2369–2377. [PubMed] [Google Scholar]
- 40.Davis JN, Kucuk O, Sarkar FH. Genistein inhibits NF-kappa B activation in prostate cancer cells. Nutr Cancer. 1999;35:167–174. doi: 10.1207/S15327914NC352_11. [DOI] [PubMed] [Google Scholar]
- 41.Mahmoud AM, Zhu T, Parray A, Siddique HR, Yang W, et al. Differential effects of genistein on prostate cancer cells depend on mutational status of the androgen receptor. PLoS One. 2013;8:e78479. doi: 10.1371/journal.pone.0078479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Seo YJ, Kim BS, Chun SY, Park YK, Kang KS, et al. Apoptotic effects of genistein, biochanin-A and apigenin on LNCaP and PC-3 cells by p21 through transcriptional inhibition of polo-like kinase-1. J Korean Med Sci. 2011;26:1489–1494. doi: 10.3346/jkms.2011.26.11.1489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Agarwal R. Cell signaling and regulators of cell cycle as molecular targets for prostate cancer prevention by dietary agents. Biochem Pharmacol. 2000;60:1051–1059. doi: 10.1016/s0006-2952(00)00385-3. [DOI] [PubMed] [Google Scholar]
- 44.Wang BF, Wang JS, Lu JF, Kao TH, Chen BH. Antiproliferation effect and mechanism of prostate cancer cell lines as affected by isoflavones from soybean cake. J Agric Food Chem. 2009;57:2221–2232. doi: 10.1021/jf8037715. [DOI] [PubMed] [Google Scholar]
- 45.Hedlund TE, Johannes WU, Miller GJ. Soy isoflavonoid equol modulates the growth of benign and malignant prostatic epithelial cells in vitro. Prostate. 2003;54:68–78. doi: 10.1002/pros.10137. [DOI] [PubMed] [Google Scholar]
- 46.Shen JC, Klein RD, Wei Q, Guan Y, Contois JH, et al. Low-dose genistein induces cyclin-dependent kinase inhibitors and G(1) cell-cycle arrest in human prostate cancer cells. Mol Carcinog. 2000;29:92–102. doi: 10.1002/1098-2744(200010)29:2<92::aid-mc6>3.0.co;2-q. [DOI] [PubMed] [Google Scholar]
- 47.Zhao R, Xiang N, Domann FE, Zhong W. Effects of selenite and genistein on G2/M cell cycle arrest and apoptosis in human prostate cancer cells. Nutr Cancer. 2009;61:397–407. doi: 10.1080/01635580802582751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Raffoul JJ, Wang Y, Kucuk O, Forman JD, Sarkar FH, et al. Genistein inhibits radiation-induced activation of NF-kappaB in prostate cancer cells promoting apoptosis and G2/M cell cycle arrest. BMC Cancer. 2006;6:107. doi: 10.1186/1471-2407-6-107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Burgering BM. A brief introduction to FOXOlogy. Oncogene. 2008;27:2258–2262. doi: 10.1038/onc.2008.29. [DOI] [PubMed] [Google Scholar]
- 50.Christensen MJ, Quiner TE, Nakken HL, Lephart ED, Eggett DL, et al. Combination effects of dietary soy and methylselenocysteine in a mouse model of prostate cancer. Prostate. 2013;73:986–995. doi: 10.1002/pros.22646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Roy SK, Srivastava RK, Shankar S. Inhibition of PI3K/AKT and MAPK/ERK pathways causes activation of FOXO transcription factor, leading to cell cycle arrest and apoptosis in pancreatic cancer. J Mol Signal. 2010;5:10. doi: 10.1186/1750-2187-5-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Takahashi Y, Lavigne JA, Hursting SD, Chandramouli GV, Perkins SN, et al. Molecular signatures of soy-derived phytochemicals in androgen-responsive prostate cancer cells: a comparison study using DNA microarray. Mol Carcinog. 2006;45:943–956. doi: 10.1002/mc.20247. [DOI] [PubMed] [Google Scholar]
- 53.Rabiau N, Kossai M, Braud M, Chalabi N, Satih S, et al. Genistein and daidzein act on a panel of genes implicated in cell cycle and angiogenesis by polymerase chain reaction arrays in human prostate cancer cell lines. Cancer Epidemiol. 2010;34:200–206. doi: 10.1016/j.canep.2009.12.018. [DOI] [PubMed] [Google Scholar]
- 54.Renehan AG, Zwahlen M, Minder C, O’Dwyer ST, Shalet SM, et al. Insulin-like growth factor (IGF)-I, IGF binding protein-3, and cancer risk: systematic review and meta-regression analysis. Lancet. 2004;363:1346–1353. doi: 10.1016/S0140-6736(04)16044-3. [DOI] [PubMed] [Google Scholar]
- 55.Ozkan EE. Plasma and tissue insulin-like growth factor-I receptor (IGF-IR) as a prognostic marker for prostate cancer and anti-IGF-IR agents as novel therapeutic strategy for refractory cases: a review. Mol Cell Endocrinol. 2011;344:1–24. doi: 10.1016/j.mce.2011.07.002. [DOI] [PubMed] [Google Scholar]
- 56.Gennigens C, Menetrier-Caux C, Droz JP. Insulin-Like Growth Factor (IGF) family and prostate cancer. Crit Rev Oncol Hematol. 2006;58:124–145. doi: 10.1016/j.critrevonc.2005.10.003. [DOI] [PubMed] [Google Scholar]
- 57.Werner H, Le Roith D. New concepts in regulation and function of the insulin-like growth factors: implications for understanding normal growth and neoplasia. Cell Mol Life Sci. 2000;57:932–942. doi: 10.1007/PL00000735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Lee J, Ju J, Park S, Hong SJ, Yoon S. Inhibition of IGF-1 signaling by genistein: modulation of E-cadherin expression and downregulation of beta-catenin signaling in hormone refractory PC-3 prostate cancer cells. Nutr Cancer. 2012;64:153–162. doi: 10.1080/01635581.2012.630161. [DOI] [PubMed] [Google Scholar]
- 59.Valentinis B, Baserga R. IGF-I receptor signalling in transformation and differentiation. Mol Pathol. 2001;54:133–137. doi: 10.1136/mp.54.3.133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Valentinis B, Romano G, Peruzzi F, Morrione A, Prisco M, et al. Growth and differentiation signals by the insulin-like growth factor 1 receptor in hemopoietic cells are mediated through different pathways. J Biol Chem. 1999;274:12423–12430. doi: 10.1074/jbc.274.18.12423. [DOI] [PubMed] [Google Scholar]
- 61.Wang S, DeGroff VL, Clinton SK. Tomato and soy polyphenols reduce insulin-like growth factor-I-stimulated rat prostate cancer cell proliferation and apoptotic resistance in vitro via inhibition of intracellular signaling pathways involving tyrosine kinase. J Nutr. 2003;133:2367–2376. doi: 10.1093/jn/133.7.2367. [DOI] [PubMed] [Google Scholar]
- 62.Diener KR, Need EF, Buchanan G, Hayball JD. TGF-beta signalling and immunity in prostate tumourigenesis. Expert Opin Ther Targets. 2010;14:179–192. doi: 10.1517/14728220903544507. [DOI] [PubMed] [Google Scholar]
- 63.Jones E, Pu H, Kyprianou N. Targeting TGF-beta in prostate cancer: therapeutic possibilities during tumor progression. Expert Opin Ther Targets. 2009;13:227–234. doi: 10.1517/14728220802705696. [DOI] [PubMed] [Google Scholar]
- 64.Huang X, Chen S, Xu L, Liu Y, Deb DK, et al. Genistein inhibits p38 map kinase activation, matrix metalloproteinase type 2, and cell invasion in human prostate epithelial cells. Cancer Res. 2005;65:3470–3478. doi: 10.1158/0008-5472.CAN-04-2807. [DOI] [PubMed] [Google Scholar]
- 65.Xu L, Bergan RC. Genistein inhibits matrix metalloproteinase type 2 activation and prostate cancer cell invasion by blocking the transforming growth factor beta-mediated activation of mitogen-activated protein kinase-activated protein kinase 2-27-kDa heat shock protein pathway. Mol Pharmacol. 2006;70:869–877. doi: 10.1124/mol.106.023861. [DOI] [PubMed] [Google Scholar]
- 66.Li Y, Sarkar FH. Down-regulation of invasion and angiogenesis-related genes identified by cDNA microarray analysis of PC3 prostate cancer cells treated with genistein. Cancer Lett. 2002;186:157–164. doi: 10.1016/s0304-3835(02)00349-x. [DOI] [PubMed] [Google Scholar]
- 67.Yost C, Torres M, Miller JR, Huang E, Kimelman D, et al. The axis-inducing activity, stability, and subcellular distribution of beta-catenin is regulated in Xenopus embryos by glycogen synthase kinase 3. Genes Dev. 1996;10:1443–1454. doi: 10.1101/gad.10.12.1443. [DOI] [PubMed] [Google Scholar]
- 68.MacDonald BT, Tamai K, He X. Wnt/beta-catenin signaling: components, mechanisms, and diseases. Dev Cell. 2009;17:9–26. doi: 10.1016/j.devcel.2009.06.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Patriarca C, Petrella D, Campo B, Colombo P, Giunta P, et al. Elevated E-cadherin and alpha/beta-catenin expression after androgen deprivation therapy in prostate adenocarcinoma. Pathol Res Pract. 2003;199:659–665. doi: 10.1078/0344-0338-00477. [DOI] [PubMed] [Google Scholar]
- 70.Morita N, Uemura H, Tsumatani K, Cho M, Hirao Y, et al. E-cadherin and alpha-, beta- and gamma-catenin expression in prostate cancers: correlation with tumour invasion. Br J Cancer. 1999;79:1879–1883. doi: 10.1038/sj.bjc.6690299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Jaggi M, Johansson SL, Baker JJ, Smith LM, Galich A, et al. Aberrant expression of E-cadherin and beta-catenin in human prostate cancer. Urol Oncol. 2005;23:402–406. doi: 10.1016/j.urolonc.2005.03.024. [DOI] [PubMed] [Google Scholar]
- 72.Liss MA, Schlicht M, Kahler A, Fitzgerald R, Thomassi T, et al. Characterization of soy-based changes in Wnt-frizzled signaling in prostate cancer. Cancer Genomics Proteomics. 2010;7:245–252. [PubMed] [Google Scholar]
- 73.Graff JR, Konicek BW, McNulty AM, Wang Z, Houck K, et al. Increased AKT activity contributes to prostate cancer progression by dramatically accelerating prostate tumor growth and diminishing p27Kip1 expression. J Biol Chem. 2000;275:24500–24505. doi: 10.1074/jbc.M003145200. [DOI] [PubMed] [Google Scholar]
- 74.Murillo H, Huang H, Schmidt LJ, Smith DI, Tindall DJ. Role of PI3K signaling in survival and progression of LNCaP prostate cancer cells to the androgen refractory state. Endocrinology. 2001;142:4795–4805. doi: 10.1210/endo.142.11.8467. [DOI] [PubMed] [Google Scholar]
- 75.Kreisberg JI, Malik SN, Prihoda TJ, Bedolla RG, Troyer DA, et al. Phosphorylation of Akt (Ser473) is an excellent predictor of poor clinical outcome in prostate cancer. Cancer Res. 2004;64:5232–5236. doi: 10.1158/0008-5472.CAN-04-0272. [DOI] [PubMed] [Google Scholar]
- 76.Uzgare AR, Isaacs JT. Enhanced redundancy in Akt and mitogen-activated protein kinase-induced survival of malignant versus normal prostate epithelial cells. Cancer Res. 2004;64:6190–6199. doi: 10.1158/0008-5472.CAN-04-0968. [DOI] [PubMed] [Google Scholar]
- 77.Li Y, Wang Z, Kong D, Li R, Sarkar SH, et al. Regulation of Akt/FOXO3a/GSK-3beta/AR signaling network by isoflavone in prostate cancer cells. J Biol Chem. 2008;283:27707–27716. doi: 10.1074/jbc.M802759200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Tepper CG, Vinall RL, Wee CB, Xue L, Shi XB, et al. GCP-mediated growth inhibition and apoptosis of prostate cancer cells via androgen receptor-dependent and -independent mechanisms. Prostate. 2007;67:521–535. doi: 10.1002/pros.20548. [DOI] [PubMed] [Google Scholar]
- 79.Oh HY, Leem J, Yoon SJ, Yoon S, Hong SJ. Lipid raft cholesterol and genistein inhibit the cell viability of prostate cancer cells via the partial contribution of EGFR-Akt/p70S6k pathway and down-regulation of androgen receptor. Biochem Biophys Res Commun. 2010;393:319–324. doi: 10.1016/j.bbrc.2010.01.133. [DOI] [PubMed] [Google Scholar]
- 80.Skogseth H, Follestad T, Larsson E, Halgunset J. Transcription levels of invasion-related genes in prostate cancer cells are modified by inhibitors of tyrosine kinase. APMIS. 2006;114:364–371. doi: 10.1111/j.1600-0463.2006.apm_370.x. [DOI] [PubMed] [Google Scholar]
- 81.Vinall RL, Hwa K, Ghosh P, Pan CX, Lara PN, Jr, et al. Combination treatment of prostate cancer cell lines with bioactive soy isoflavones and perifosine causes increased growth arrest and/or apoptosis. Clin Cancer Res. 2007;13:6204–6216. doi: 10.1158/1078-0432.CCR-07-0600. [DOI] [PubMed] [Google Scholar]
- 82.Cao F, Jin TY, Zhou YF. Inhibitory effect of isoflavones on prostate cancer cells and PTEN gene. Biomed Environ Sci. 2006;19:35–41. [PubMed] [Google Scholar]
- 83.Kumar R, Verma V, Jain A, Jain RK, Maikhuri JP, et al. Synergistic chemoprotective mechanisms of dietary phytoestrogens in a select combination against prostate cancer. J Nutr Biochem. 2011;22:723–731. doi: 10.1016/j.jnutbio.2010.06.003. [DOI] [PubMed] [Google Scholar]
- 84.Kumi-Diaka J, Sanderson NA, Hall A. The mediating role of caspase-3 protease in the intracellular mechanism of genistein-induced apoptosis in human prostatic carcinoma cell lines, DU145 and LNCaP. Biol Cell. 2000;92:595–604. doi: 10.1016/s0248-4900(00)01109-6. [DOI] [PubMed] [Google Scholar]
- 85.Kazi A, Daniel KG, Smith DM, Kumar NB, Dou QP. Inhibition of the proteasome activity, a novel mechanism associated with the tumor cell apoptosis-inducing ability of genistein. Biochem Pharmacol. 2003;66:965–976. doi: 10.1016/s0006-2952(03)00414-3. [DOI] [PubMed] [Google Scholar]
- 86.Hsu A, Bray TM, Helferich WG, Doerge DR, Ho E. Differential effects of whole soy extract and soy isoflavones on apoptosis in prostate cancer cells. Exp Biol Med (Maywood) 2010;235:90–97. doi: 10.1258/ebm.2009.009128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Li Y, Sarkar FH. Gene expression profiles of genistein-treated PC3 prostate cancer cells. J Nutr. 2002;132:3623–3631. doi: 10.1093/jn/132.12.3623. [DOI] [PubMed] [Google Scholar]
- 88.Clubbs EA, Bomser JA. Basal cell induced differentiation of noncancerous prostate epithelial cells (RWPE-1) by glycitein. Nutr Cancer. 2009;61:390–396. doi: 10.1080/01635580802582728. [DOI] [PubMed] [Google Scholar]
- 89.Mentor-Marcel R, Lamartiniere CA, Eltoum IE, Greenberg NM, Elgavish A. Genistein in the diet reduces the incidence of poorly differentiated prostatic adenocarcinoma in transgenic mice (TRAMP) Cancer Res. 2001;61:6777–6782. [PubMed] [Google Scholar]
- 90.Singletary K, Milner J. Diet, autophagy, and cancer: a review. Cancer Epidemiol Biomarkers Prev. 2008;17:1596–1610. doi: 10.1158/1055-9965.EPI-07-2917. [DOI] [PubMed] [Google Scholar]
- 91.Alers S, Loffler AS, Wesselborg S, Stork B. Role of AMPK-mTOR-Ulk1/2 in the regulation of autophagy: cross talk, shortcuts, and feedbacks. Mol Cell Biol. 2012;32:2–11. doi: 10.1128/MCB.06159-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Nakamura Y, Yogosawa S, Izutani Y, Watanabe H, Otsuji E, et al. A combination of indol-3-carbinol and genistein synergistically induces apoptosis in human colon cancer HT-29 cells by inhibiting Akt phosphorylation and progression of autophagy. Mol Cancer. 2009;8:100. doi: 10.1186/1476-4598-8-100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Gossner G, Choi M, Tan L, Fogoros S, Griffith KA, et al. Genistein-induced apoptosis and autophagocytosis in ovarian cancer cells. Gynecologic Oncology. 2007;105:23–30. doi: 10.1016/j.ygyno.2006.11.009. [DOI] [PubMed] [Google Scholar]
- 94.Valko M, Rhodes CJ, Moncol J, Izakovic M, Mazur M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem Biol Interact. 2006;160:1–40. doi: 10.1016/j.cbi.2005.12.009. [DOI] [PubMed] [Google Scholar]
- 95.Pugalendhi P, Manoharan S, Panjamurthy K, Balakrishnan S, Nirmal MR. Antigenotoxic effect of genistein against 7,12-dimethylbenz[a]anthracene induced genotoxicity in bone marrow cells of female Wistar rats. Pharmacol Rep. 2009;61:296–303. doi: 10.1016/s1734-1140(09)70035-0. [DOI] [PubMed] [Google Scholar]
- 96.Leung HY, Yung LH, Poon CH, Shi G, Lu AL, et al. Genistein protects against polycyclic aromatic hydrocarbon-induced oxidative DNA damage in non-cancerous breast cells MCF-10A. Br J Nutr. 2009;101:257–262. doi: 10.1017/S0007114508998457. [DOI] [PubMed] [Google Scholar]
- 97.Khan N, Afaq F, Mukhtar H. Cancer chemoprevention through dietary antioxidants: progress and promise. Antioxid Redox Signal. 2008;10:475–510. doi: 10.1089/ars.2007.1740. [DOI] [PubMed] [Google Scholar]
- 98.Foti P, Erba D, Riso P, Spadafranca A, Criscuoli F, et al. Comparison between daidzein and genistein antioxidant activity in primary and cancer lymphocytes. Arch Biochem Biophys. 2005;433:421–427. doi: 10.1016/j.abb.2004.10.008. [DOI] [PubMed] [Google Scholar]
- 99.Suzuki K, Koike H, Matsui H, Ono Y, Hasumi M, et al. Genistein, a soy isoflavone, induces glutathione peroxidase in the human prostate cancer cell lines LNCaP and PC-3. Int J Cancer. 2002;99:846–852. doi: 10.1002/ijc.10428. [DOI] [PubMed] [Google Scholar]
- 100.Raschke M, Rowland IR, Magee PJ, Pool-Zobel BL. Genistein protects prostate cells against hydrogen peroxide-induced DNA damage and induces expression of genes involved in the defence against oxidative stress. Carcinogenesis. 2006;27:2322–2330. doi: 10.1093/carcin/bgl082. [DOI] [PubMed] [Google Scholar]
- 101.Park CE, Yun H, Lee EB, Min BI, Bae H, et al. The antioxidant effects of genistein are associated with AMP-activated protein kinase activation and PTEN induction in prostate cancer cells. J Med Food. 2010;13:815–820. doi: 10.1089/jmf.2009.1359. [DOI] [PubMed] [Google Scholar]
- 102.Oki T, Sowa Y, Hirose T, Takagaki N, Horinaka M, et al. Genistein induces Gadd45 gene and G2/M cell cycle arrest in the DU145 human prostate cancer cell line. FEBS Lett. 2004;577:55–59. doi: 10.1016/j.febslet.2004.09.085. [DOI] [PubMed] [Google Scholar]
- 103.Hussain T, Gupta S, Mukhtar H. Cyclooxygenase-2 and prostate carcinogenesis. Cancer Lett. 2003;191:125–135. doi: 10.1016/s0304-3835(02)00524-4. [DOI] [PubMed] [Google Scholar]
- 104.Badawi AF. The role of prostaglandin synthesis in prostate cancer. BJU Int. 2000;85:451–462. doi: 10.1046/j.1464-410x.2000.00507.x. [DOI] [PubMed] [Google Scholar]
- 105.Koki AT, Masferrer JL. Celecoxib: a specific COX-2 inhibitor with anticancer properties. Cancer Control. 2002;9:28–35. doi: 10.1177/107327480200902S04. [DOI] [PubMed] [Google Scholar]
- 106.Otto JC, Smith WL. Prostaglandin endoperoxide synthases-1 and -2. J Lipid Mediat Cell Signal. 1995;12:139–156. doi: 10.1016/0929-7855(95)00015-i. [DOI] [PubMed] [Google Scholar]
- 107.Fosslien E. Review: molecular pathology of cyclooxygenase-2 in cancer-induced angiogenesis. Ann Clin Lab Sci. 2001;31:325–348. [PubMed] [Google Scholar]
- 108.Aparicio Gallego G, Diaz Prado S, Jimenez Fonseca P, Garcia Campelo R, Cassinello Espinosa J, et al. Cyclooxygenase-2 (COX-2): a molecular target in prostate cancer. Clin Transl Oncol. 2007;9:694–702. doi: 10.1007/s12094-007-0126-0. [DOI] [PubMed] [Google Scholar]
- 109.Swami S, Krishnan AV, Moreno J, Bhattacharyya RB, Peehl DM, et al. Calcitriol and genistein actions to inhibit the prostaglandin pathway: potential combination therapy to treat prostate cancer. J Nutr. 2007;137:205S–210S. doi: 10.1093/jn/137.1.205S. [DOI] [PubMed] [Google Scholar]
- 110.Alison MR, Lim SM, Nicholson LJ. Cancer stem cells: problems for therapy? J Pathol. 2011;223:147–161. doi: 10.1002/path.2793. [DOI] [PubMed] [Google Scholar]
- 111.Barclay WW, Axanova LS, Chen W, Romero L, Maund SL, et al. Characterization of adult prostatic progenitor/stem cells exhibiting self-renewal and multilineage differentiation. Stem Cells. 2008;26:600–610. doi: 10.1634/stemcells.2007-0309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.English HF, Santen RJ, Isaacs JT. Response of glandular versus basal rat ventral prostatic epithelial cells to androgen withdrawal and replacement. Prostate. 1987;11:229–242. doi: 10.1002/pros.2990110304. [DOI] [PubMed] [Google Scholar]
- 113.Tang DG, Patrawala L, Calhoun T, Bhatia B, Choy G, et al. Prostate cancer stem/progenitor cells: identification, characterization, and implications. Mol Carcinog. 2007;46:1–14. doi: 10.1002/mc.20255. [DOI] [PubMed] [Google Scholar]
- 114.Leong KG, Wang BE, Johnson L, Gao WQ. Generation of a prostate from a single adult stem cell. Nature. 2008;456:804–808. doi: 10.1038/nature07427. [DOI] [PubMed] [Google Scholar]
- 115.Wang X, Kruithof-de Julio M, Economides KD, Walker D, Yu H, et al. A luminal epithelial stem cell that is a cell of origin for prostate cancer. Nature. 2009;461:495–500. doi: 10.1038/nature08361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Patrawala L, Calhoun T, Schneider-Broussard R, Li H, Bhatia B, et al. Highly purified CD44+ prostate cancer cells from xenograft human tumors are enriched in tumorigenic and metastatic progenitor cells. Oncogene. 2006;25:1696–1708. doi: 10.1038/sj.onc.1209327. [DOI] [PubMed] [Google Scholar]
- 117.McCubrey JA, Chappell WH, Abrams SL, Franklin RA, Long JM, et al. Targeting the cancer initiating cell: the Achilles’ heel of cancer. Adv Enzyme Regul. 2011;51:152–162. doi: 10.1016/j.advenzreg.2010.09.002. [DOI] [PubMed] [Google Scholar]
- 118.McCubrey JA, Abrams SL, Stadelman K, Chappell WH, Lahair M, et al. Targeting signal transduction pathways to eliminate chemotherapeutic drug resistance and cancer stem cells. Adv Enzyme Regul. 2010;50:285–307. doi: 10.1016/j.advenzreg.2009.10.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Ouchi H, Ishiguro H, Ikeda N, Hori M, Kubota Y, et al. Genistein induces cell growth inhibition in prostate cancer through the suppression of telomerase activity. Int J Urol. 2005;12:73–80. doi: 10.1111/j.1442-2042.2004.00973.x. [DOI] [PubMed] [Google Scholar]
- 120.Montgomery RK, Carlone DL, Richmond CA, Farilla L, Kranendonk ME, et al. Mouse telomerase reverse transcriptase (mTert) expression marks slowly cycling intestinal stem cells. Proc Natl Acad Sci U S A. 2011;108:179–184. doi: 10.1073/pnas.1013004108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Molinie B, Georgel P. Genetic and epigenetic regulations of prostate cancer by genistein. Drug News Perspect. 2009;22:247–254. doi: 10.1358/dnp.2009.22.5.1378633. [DOI] [PubMed] [Google Scholar]
- 122.Adjakly M, Bosviel R, Rabiau N, Boiteux JP, Bignon YJ, et al. DNA methylation and soy phytoestrogens: quantitative study in DU-145 and PC-3 human prostate cancer cell lines. Epigenomics. 2011;3:795–803. doi: 10.2217/epi.11.103. [DOI] [PubMed] [Google Scholar]
- 123.Vardi A, Bosviel R, Rabiau N, Adjakly M, Satih S, et al. Soy phytoestrogens modify DNA methylation of GSTP1, RASSF1A, EPH2 and BRCA1 promoter in prostate cancer cells. In Vivo. 2010;24:393–400. [PubMed] [Google Scholar]
- 124.Majid S, Dar AA, Shahryari V, Hirata H, Ahmad A, et al. Genistein reverses hypermethylation and induces active histone modifications in tumor suppressor gene B-Cell translocation gene 3 in prostate cancer. Cancer. 2010;116:66–76. doi: 10.1002/cncr.24662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Kikuno N, Shiina H, Urakami S, Kawamoto K, Hirata H, et al. Genistein mediated histone acetylation and demethylation activates tumor suppressor genes in prostate cancer cells. Int J Cancer. 2008;123:552–560. doi: 10.1002/ijc.23590. [DOI] [PubMed] [Google Scholar]
- 126.Fang MZ, Chen D, Sun Y, Jin Z, Christman JK, et al. Reversal of hypermethylation and reactivation of p16INK4a, RARbeta, and MGMT genes by genistein and other isoflavones from soy. Clin Cancer Res. 2005;11:7033–7041. doi: 10.1158/1078-0432.CCR-05-0406. [DOI] [PubMed] [Google Scholar]
- 127.Bulger M. Hyperacetylated chromatin domains: lessons from heterochromatin. J Biol Chem. 2005;280:21689–21692. doi: 10.1074/jbc.R500004200. [DOI] [PubMed] [Google Scholar]
- 128.Catto JW, Alcaraz A, Bjartell AS, De Vere White R, Evans CP, et al. MicroRNA in prostate, bladder, and kidney cancer: a systematic review. Eur Urol. 2011;59:671–681. doi: 10.1016/j.eururo.2011.01.044. [DOI] [PubMed] [Google Scholar]
- 129.Li Y, Kong D, Ahmad A, Bao B, Dyson G, et al. Epigenetic deregulation of miR-29a and miR-1256 by isoflavone contributes to the inhibition of prostate cancer cell growth and invasion. Epigenetics. 2012;7 doi: 10.4161/epi.21236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Rabiau N, Trraf HK, Adjakly M, Bosviel R, Guy L, et al. miRNAs differentially expressed in prostate cancer cell lines after soy treatment. In Vivo. 2011;25:917–921. [PubMed] [Google Scholar]
- 131.Bergers G, Benjamin LE. Tumorigenesis and the angiogenic switch. Nat Rev Cancer. 2003;3:401–410. doi: 10.1038/nrc1093. [DOI] [PubMed] [Google Scholar]
- 132.Ambra R, Rimbach G, de Pascual Teresa S, Fuchs D, Wenzel U, et al. Genistein affects the expression of genes involved in blood pressure regulation and angiogenesis in primary human endothelial cells. Nutr Metab Cardiovasc Dis. 2006;16:35–43. doi: 10.1016/j.numecd.2005.03.003. [DOI] [PubMed] [Google Scholar]
- 133.Yu X, Zhu J, Mi M, Chen W, Pan Q, et al. Anti-angiogenic genistein inhibits VEGF-induced endothelial cell activation by decreasing PTK activity and MAPK activation. Med Oncol. 2012;29:349–357. doi: 10.1007/s12032-010-9770-2. [DOI] [PubMed] [Google Scholar]
- 134.Guo Y, Wang S, Hoot DR, Clinton SK. Suppression of VEGF-mediated autocrine and paracrine interactions between prostate cancer cells and vascular endothelial cells by soy isoflavones. J Nutr Biochem. 2007;18:408–417. doi: 10.1016/j.jnutbio.2006.08.006. [DOI] [PubMed] [Google Scholar]
- 135.Pines A, Perrone L, Bivi N, Romanello M, Damante G, et al. Activation of APE1/Ref-1 is dependent on reactive oxygen species generated after purinergic receptor stimulation by ATP. Nucleic Acids Res. 2005;33:4379–4394. doi: 10.1093/nar/gki751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Raffoul JJ, Banerjee S, Singh-Gupta V, Knoll ZE, Fite A, et al. Down-regulation of apurinic/apyrimidinic endonuclease 1/redox factor-1 expression by soy isoflavones enhances prostate cancer radiotherapy in vitro and in vivo. Cancer Res. 2007;67:2141–2149. doi: 10.1158/0008-5472.CAN-06-2147. [DOI] [PubMed] [Google Scholar]
- 137.Singh-Gupta V, Zhang H, Banerjee S, Kong D, Raffoul JJ, et al. Radiation-induced HIF-1alpha cell survival pathway is inhibited by soy isoflavones in prostate cancer cells. Int J Cancer. 2009;124:1675–1684. doi: 10.1002/ijc.24015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Zhou JR, Gugger ET, Tanaka T, Guo Y, Blackburn GL, et al. Soybean phytochemicals inhibit the growth of transplantable human prostate carcinoma and tumor angiogenesis in mice. J Nutr. 1999;129:1628–1635. doi: 10.1093/jn/129.9.1628. [DOI] [PubMed] [Google Scholar]
- 139.Handayani R, Rice L, Cui Y, Medrano TA, Samedi VG, et al. Soy isoflavones alter expression of genes associated with cancer progression, including interleukin-8, in androgen-independent PC-3 human prostate cancer cells. J Nutr. 2006;136:75–82. doi: 10.1093/jn/136.1.75. [DOI] [PubMed] [Google Scholar]
- 140.Lehrer S, Diamond EJ, Mamkine B, Stone NN, Stock RG. Serum interleukin-8 is elevated in men with prostate cancer and bone metastases. Technol Cancer Res Treat. 2004;3:411. doi: 10.1177/153303460400300501. [DOI] [PubMed] [Google Scholar]
- 141.Uehara H, Troncoso P, Johnston D, Bucana CD, Dinney C, et al. Expression of interleukin-8 gene in radical prostatectomy specimens is associated with advanced pathologic stage. Prostate. 2005;64:40–49. doi: 10.1002/pros.20223. [DOI] [PubMed] [Google Scholar]
- 142.Inoue K, Slaton JW, Eve BY, Kim SJ, Perrotte P, et al. Interleukin 8 expression regulates tumorigenicity and metastases in androgen-independent prostate cancer. Clin Cancer Res. 2000;6:2104–2119. [PubMed] [Google Scholar]
- 143.Klein CA. Cancer. The metastasis cascade. Science. 2008;321:1785–1787. doi: 10.1126/science.1164853. [DOI] [PubMed] [Google Scholar]
- 144.Zhang LL, Li L, Wu DP, Fan JH, Li X, et al. A novel anti-cancer effect of genistein: reversal of epithelial mesenchymal transition in prostate cancer cells. Acta Pharmacol Sin. 2008;29:1060–1068. doi: 10.1111/j.1745-7254.2008.00831.x. [DOI] [PubMed] [Google Scholar]
- 145.Xu J, Wang R, Xie ZH, Odero-Marah V, Pathak S, et al. Prostate cancer metastasis: role of the host microenvironment in promoting epithelial to mesenchymal transition and increased bone and adrenal gland metastasis. Prostate. 2006;66:1664–1673. doi: 10.1002/pros.20488. [DOI] [PubMed] [Google Scholar]
- 146.Kumi-Diaka JK, Hassanhi M, Merchant K, Horman V. Influence of genistein isoflavone on matrix metalloproteinase-2 expression in prostate cancer cells. J Med Food. 2006;9:491–497. doi: 10.1089/jmf.2006.9.491. [DOI] [PubMed] [Google Scholar]
- 147.Li Y, Kucuk O, Hussain M, Abrams J, Cher ML, et al. Antitumor and antimetastatic activities of docetaxel are enhanced by genistein through regulation of osteoprotegerin/receptor activator of nuclear factor-kappaB (RANK)/RANK ligand/MMP-9 signaling in prostate cancer. Cancer Res. 2006;66:4816–4825. doi: 10.1158/0008-5472.CAN-05-3752. [DOI] [PubMed] [Google Scholar]
- 148.Lakshman M, Xu L, Ananthanarayanan V, Cooper J, Takimoto CH, et al. Dietary genistein inhibits metastasis of human prostate cancer in mice. Cancer Res. 2008;68:2024–2032. doi: 10.1158/0008-5472.CAN-07-1246. [DOI] [PubMed] [Google Scholar]
- 149.Liu Y, Kyle E, Lieberman R, Crowell J, Kellof G, et al. Focal adhesion kinase (FAK) phosphorylation is not required for genistein-induced FAK-beta-1-integrin complex formation. Clin Exp Metastasis. 2000;18:203–212. doi: 10.1023/a:1006729106034. [DOI] [PubMed] [Google Scholar]
- 150.Xu L, Chen S, Bergan RC. MAPKAPK2 and HSP27 are downstream effectors of p38 MAP kinase-mediated matrix metalloproteinase type 2 activation and cell invasion in human prostate cancer. Oncogene. 2006;25:2987–2998. doi: 10.1038/sj.onc.1209337. [DOI] [PubMed] [Google Scholar]
- 151.Mentor-Marcel R, Lamartiniere CA, Eltoum IA, Greenberg NM, Elgavish A. Dietary genistein improves survival and reduces expression of osteopontin in the prostate of transgenic mice with prostatic adenocarcinoma (TRAMP) J Nutr. 2005;135:989–995. doi: 10.1093/jn/135.5.989. [DOI] [PubMed] [Google Scholar]
- 152.El Touny LH, Banerjee PP. Identification of a biphasic role for genistein in the regulation of prostate cancer growth and metastasis. Cancer Res. 2009;69:3695–3703. doi: 10.1158/0008-5472.CAN-08-2958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Thalmann GN, Sikes RA, Devoll RE, Kiefer JA, Markwalder R, et al. Osteopontin: possible role in prostate cancer progression. Clin Cancer Res. 1999;5:2271–2277. [PubMed] [Google Scholar]
- 154.Bouchal J, Santer FR, Hoschele PP, Tomastikova E, Neuwirt H, et al. Transcriptional coactivators p300 and CBP stimulate estrogen receptor-beta signaling and regulate cellular events in prostate cancer. Prostate. 2011;71:431–437. doi: 10.1002/pros.21257. [DOI] [PubMed] [Google Scholar]
- 155.Miekus K, Madeja Z. Genistein inhibits the contact-stimulated migration of prostate cancer cells. Cell Mol Biol Lett. 2007;12:348–361. doi: 10.2478/s11658-007-0007-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Zhou JR, Yu L, Zhong Y, Nassr RL, Franke AA, et al. Inhibition of orthotopic growth and metastasis of androgen-sensitive human prostate tumors in mice by bioactive soybean components. Prostate. 2002;53:143–153. doi: 10.1002/pros.10141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Joseph IB, Isaacs JT. Macrophage role in the anti-prostate cancer response to one class of antiangiogenic agents. J Natl Cancer Inst. 1998;90:1648–1653. doi: 10.1093/jnci/90.21.1648. [DOI] [PubMed] [Google Scholar]
- 158.Wang M, Hu Y, Shima I, Stearns ME. IL-10/IL-10 receptor signaling regulates TIMP-1 expression in primary human prostate tumor lines. Cancer Biol Ther. 2002;1:556–563. doi: 10.4161/cbt.1.5.222. [DOI] [PubMed] [Google Scholar]
- 159.Seibel J, Molzberger AF, Hertrampf T, Laudenbach-Leschowski U, Diel P. Oral treatment with genistein reduces the expression of molecular and biochemical markers of inflammation in a rat model of chronic TNBS-induced colitis. Eur J Nutr. 2009;48:213–220. doi: 10.1007/s00394-009-0004-3. [DOI] [PubMed] [Google Scholar]
- 160.Ji G, Yang Q, Hao J, Guo L, Chen X, et al. Anti-inflammatory effect of genistein on non-alcoholic steatohepatitis rats induced by high fat diet and its potential mechanisms. Int Immunopharmacol. 2011;11:762–768. doi: 10.1016/j.intimp.2011.01.036. [DOI] [PubMed] [Google Scholar]
- 161.Velasquez MT, Bhathena SJ. Dietary phytoestrogens: a possible role in renal disease protection. Am J Kidney Dis. 2001;37:1056–1068. doi: 10.1016/s0272-6386(05)80025-3. [DOI] [PubMed] [Google Scholar]
- 162.Afaq F, Katiyar SK. Polyphenols: skin photoprotection and inhibition of photocarcinogenesis. Mini Rev Med Chem. 2011;11:1200–1215. doi: 10.2174/13895575111091200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Hillman GG, Wang Y, Kucuk O, Che M, Doerge DR, et al. Genistein potentiates inhibition of tumor growth by radiation in a prostate cancer orthotopic model. Mol Cancer Ther. 2004;3:1271–1279. [PubMed] [Google Scholar]
- 164.Hong SJ, Kim SI, Kwon SM, Lee JR, Chung BC. Comparative study of concentration of isoflavones and lignans in plasma and prostatic tissues of normal control and benign prostatic hyperplasia. Yonsei Med J. 2002;43:236–241. doi: 10.3349/ymj.2002.43.2.236. [DOI] [PubMed] [Google Scholar]
- 165.Bolla M, Collette L, Blank L, Warde P, Dubois JB, et al. Long-term results with immediate androgen suppression and external irradiation in patients with locally advanced prostate cancer (an EORTC study): a phase III randomised trial. Lancet. 2002;360:103–106. doi: 10.1016/s0140-6736(02)09408-4. [DOI] [PubMed] [Google Scholar]
- 166.Hermann RM, Wolff HA, Jarry H, Thelen P, Gruendker C, et al. In vitro studies on the modification of low-dose hyper-radiosensitivity in prostate cancer cells by incubation with genistein and estradiol. Radiat Oncol. 2008;3:19. doi: 10.1186/1748-717X-3-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Hillman GG, Forman JD, Kucuk O, Yudelev M, Maughan RL, et al. Genistein potentiates the radiation effect on prostate carcinoma cells. Clin Cancer Res. 2001;7:382–390. [PubMed] [Google Scholar]
- 168.Raffoul JJ, Banerjee S, Che M, Knoll ZE, Doerge DR, et al. Soy isoflavones enhance radiotherapy in a metastatic prostate cancer model. Int J Cancer. 2007;120:2491–2498. doi: 10.1002/ijc.22548. [DOI] [PubMed] [Google Scholar]
- 169.Ping SY, Hour TC, Lin SR, Yu DS. Taxol synergizes with antioxidants in inhibiting hormal refractory prostate cancer cell growth. Urol Oncol. 2010;28:170–179. doi: 10.1016/j.urolonc.2008.07.003. [DOI] [PubMed] [Google Scholar]
- 170.Burich RA, Holland WS, Vinall RL, Tepper C, White RW, et al. Genistein combined polysaccharide enhances activity of docetaxel, bicalutamide and Src kinase inhibition in androgen-dependent and independent prostate cancer cell lines. BJU Int. 2008;102:1458–1466. doi: 10.1111/j.1464-410X.2008.07826.x. [DOI] [PubMed] [Google Scholar]
- 171.Huggins C. Effect of Orchiectomy and Irradiation on Cancer of the Prostate. Ann Surg. 1942;115:1192–1200. doi: 10.1097/00000658-194206000-00030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Grossmann ME, Huang H, Tindall DJ. Androgen receptor signaling in androgen-refractory prostate cancer. J Natl Cancer Inst. 2001;93:1687–1697. doi: 10.1093/jnci/93.22.1687. [DOI] [PubMed] [Google Scholar]
- 173.Debes JD, Tindall DJ. Mechanisms of androgen-refractory prostate cancer. N Engl J Med. 2004;351:1488–1490. doi: 10.1056/NEJMp048178. [DOI] [PubMed] [Google Scholar]
- 174.Agoulnik IU, Weigel NL. Androgen receptor action in hormone-dependent and recurrent prostate cancer. J Cell Biochem. 2006;99:362–372. doi: 10.1002/jcb.20811. [DOI] [PubMed] [Google Scholar]
- 175.Koivisto P, Kononen J, Palmberg C, Tammela T, Hyytinen E, et al. Androgen receptor gene amplification: a possible molecular mechanism for androgen deprivation therapy failure in prostate cancer. Cancer Res. 1997;57:314–319. [PubMed] [Google Scholar]
- 176.Gregory CW, He B, Johnson RT, Ford OH, Mohler JL, et al. A mechanism for androgen receptor-mediated prostate cancer recurrence after androgen deprivation therapy. Cancer Res. 2001;61:4315–4319. [PubMed] [Google Scholar]
- 177.Li P, Yu X, Ge K, Melamed J, Roeder RG, et al. Heterogeneous expression and functions of androgen receptor co-factors in primary prostate cancer. Am J Pathol. 2002;161:1467–1474. doi: 10.1016/S0002-9440(10)64422-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Linja MJ, Porkka KP, Kang Z, Savinainen KJ, Janne OA, et al. Expression of androgen receptor coregulators in prostate cancer. Clin Cancer Res. 2004;10:1032–1040. doi: 10.1158/1078-0432.ccr-0990-3. [DOI] [PubMed] [Google Scholar]
- 179.Chen T, Wang LH, Farrar WL. Interleukin 6 activates androgen receptor-mediated gene expression through a signal transducer and activator of transcription 3-dependent pathway in LNCaP prostate cancer cells. Cancer Res. 2000;60:2132–2135. [PubMed] [Google Scholar]
- 180.Craft N, Shostak Y, Carey M, Sawyers CL. A mechanism for hormone-independent prostate cancer through modulation of androgen receptor signaling by the HER-2/neu tyrosine kinase. Nat Med. 1999;5:280–285. doi: 10.1038/6495. [DOI] [PubMed] [Google Scholar]
- 181.Kupelian P, Katcher J, Levin H, Zippe C, Klein E. Correlation of clinical and pathologic factors with rising prostate-specific antigen profiles after radical prostatectomy alone for clinically localized prostate cancer. Urology. 1996;48:249–260. doi: 10.1016/S0090-4295(96)00167-7. [DOI] [PubMed] [Google Scholar]
- 182.Luke MC, Coffey DS. Human androgen receptor binding to the androgen response element of prostate specific antigen. J Androl. 1994;15:41–51. [PubMed] [Google Scholar]
- 183.Fritz WA, Wang J, Eltoum IE, Lamartiniere CA. Dietary genistein down-regulates androgen and estrogen receptor expression in the rat prostate. Mol Cell Endocrinol. 2002;186:89–99. doi: 10.1016/s0303-7207(01)00663-3. [DOI] [PubMed] [Google Scholar]
- 184.Wang J, Eltoum IE, Lamartiniere CA. Genistein chemoprevention of prostate cancer in TRAMP mice. J Carcinog. 2007;6:3. doi: 10.1186/1477-3163-6-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Davis JN, Kucuk O, Sarkar FH. Expression of prostate-specific antigen is transcriptionally regulated by genistein in prostate cancer cells. Mol Carcinog. 2002;34:91–101. doi: 10.1002/mc.10053. [DOI] [PubMed] [Google Scholar]
- 186.Basak S, Pookot D, Noonan EJ, Dahiya R. Genistein down-regulates androgen receptor by modulating HDAC6-Hsp90 chaperone function. Mol Cancer Ther. 2008;7:3195–3202. doi: 10.1158/1535-7163.MCT-08-0617. [DOI] [PubMed] [Google Scholar]
- 187.Takahashi Y, Lavigne JA, Hursting SD, Chandramouli GV, Perkins SN, et al. Using DNA microarray analyses to elucidate the effects of genistein in androgen-responsive prostate cancer cells: identification of novel targets. Mol Carcinog. 2004;41:108–119. doi: 10.1002/mc.20045. [DOI] [PubMed] [Google Scholar]
- 188.Payton-Stewart F, Schoene NW, Kim YS, Burow ME, Cleveland TE, et al. Molecular effects of soy phytoalexin glyceollins in human prostate cancer cells LNCaP. Mol Carcinog. 2009;48:862–871. doi: 10.1002/mc.20532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Bektic J, Berger AP, Pfeil K, Dobler G, Bartsch G, et al. Androgen receptor regulation by physiological concentrations of the isoflavonoid genistein in androgen-dependent LNCaP cells is mediated by estrogen receptor beta. Eur Urol. 2004;45:245–251. doi: 10.1016/j.eururo.2003.09.001. discussion 251. [DOI] [PubMed] [Google Scholar]
- 190.Lazarevic B, Karlsen SJ, Saatcioglu F. Genistein differentially modulates androgen-responsive gene expression and activates JNK in LNCaP cells. Oncol Rep. 2008;19:1231–1235. [PubMed] [Google Scholar]
- 191.Gao S, Liu GZ, Wang Z. Modulation of androgen receptor-dependent transcription by resveratrol and genistein in prostate cancer cells. Prostate. 2004;59:214–225. doi: 10.1002/pros.10375. [DOI] [PubMed] [Google Scholar]
- 192.Maggiolini M, Vivacqua A, Carpino A, Bonofiglio D, Fasanella G, et al. The mutant androgen receptor T877A mediates the proliferative but not the cytotoxic dose-dependent effects of genistein and quercetin on human LNCaP prostate cancer cells. Mol Pharmacol. 2002;62:1027–1035. doi: 10.1124/mol.62.5.1027. [DOI] [PubMed] [Google Scholar]
- 193.Bosland MC. The role of estrogens in prostate carcinogenesis: a rationale for chemoprevention. Rev Urol. 2005;7(Suppl 3):S4–S10. [PMC free article] [PubMed] [Google Scholar]
- 194.Bonkhoff H, Fixemer T, Hunsicker I, Remberger K. Estrogen receptor expression in prostate cancer and premalignant prostatic lesions. Am J Pathol. 1999;155:641–647. doi: 10.1016/S0002-9440(10)65160-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Fixemer T, Remberger K, Bonkhoff H. Differential expression of the estrogen receptor beta (ERbeta) in human prostate tissue, premalignant changes, and in primary, metastatic, and recurrent prostatic adenocarcinoma. Prostate. 2003;54:79–87. doi: 10.1002/pros.10171. [DOI] [PubMed] [Google Scholar]
- 196.Leav I, Lau KM, Adams JY, McNeal JE, Taplin ME, et al. Comparative studies of the estrogen receptors beta and alpha and the androgen receptor in normal human prostate glands, dysplasia, and in primary and metastatic carcinoma. Am J Pathol. 2001;159:79–92. doi: 10.1016/s0002-9440(10)61676-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Ricke WA, McPherson SJ, Bianco JJ, Cunha GR, Wang Y, et al. Prostatic hormonal carcinogenesis is mediated by in situ estrogen production and estrogen receptor alpha signaling. FASEB J. 2008;22:1512–1520. doi: 10.1096/fj.07-9526com. [DOI] [PubMed] [Google Scholar]
- 198.Price D, Stein B, Sieber P, Tutrone R, Bailen J, et al. Toremifene for the prevention of prostate cancer in men with high grade prostatic intraepithelial neoplasia: results of a double-blind, placebo controlled, phase IIB clinical trial. J Urol. 2006;176:965–970. doi: 10.1016/j.juro.2006.04.011. discussion 970–961. [DOI] [PubMed] [Google Scholar]
- 199.Weihua Z, Makela S, Andersson LC, Salmi S, Saji S, et al. A role for estrogen receptor beta in the regulation of growth of the ventral prostate. Proc Natl Acad Sci U S A. 2001;98:6330–6335. doi: 10.1073/pnas.111150898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Imamov O, Morani A, Shim GJ, Omoto Y, Thulin-Andersson C, et al. Estrogen receptor beta regulates epithelial cellular differentiation in the mouse ventral prostate. Proc Natl Acad Sci U S A. 2004;101:9375–9380. doi: 10.1073/pnas.0403041101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.McPherson SJ, Hussain S, Balanathan P, Hedwards SL, Niranjan B, et al. Estrogen receptor-beta activated apoptosis in benign hyperplasia and cancer of the prostate is androgen independent and TNFalpha mediated. Proc Natl Acad Sci U S A. 2010;107:3123–3128. doi: 10.1073/pnas.0905524107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Zhu X, Leav I, Leung YK, Wu M, Liu Q, et al. Dynamic regulation of estrogen receptor-beta expression by DNA methylation during prostate cancer development and metastasis. Am J Pathol. 2004;164:2003–2012. doi: 10.1016/s0002-9440(10)63760-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203.Chang WY, Prins GS. Estrogen receptor-beta: implications for the prostate gland. Prostate. 1999;40:115–124. doi: 10.1002/(sici)1097-0045(19990701)40:2<115::aid-pros7>3.0.co;2-3. [DOI] [PubMed] [Google Scholar]
- 204.Sonoda T, Suzuki H, Mori M, Tsukamoto T, Yokomizo A, et al. Polymorphisms in estrogen related genes may modify the protective effect of isoflavones against prostate cancer risk in Japanese men. Eur J Cancer Prev. 2010;19:131–137. doi: 10.1097/CEJ.0b013e328333fbe2. [DOI] [PubMed] [Google Scholar]
- 205.Iwasaki M, Hamada GS, Nishimoto IN, Netto MM, Motola J, Jr, et al. Isoflavone, polymorphisms in estrogen receptor genes and breast cancer risk in case-control studies in Japanese, Japanese Brazilians and non-Japanese Brazilians. Cancer Sci. 2009;100:927–933. doi: 10.1111/j.1349-7006.2009.01118.x. [DOI] [PMC free article] [PubMed] [Google Scholar]