

Abstract
Peripheral neuropathy (PN) is the most frequent neurologic complication in individuals infected with human immunodeficiency virus (HIV). It affects over one third of infected patients, including those receiving effective combination antiretroviral therapy. The pathogenesis of HIV-associated peripheral neuropathy (HIV-PN) remains poorly understood. Clinical studies are complicated because both HIV and antiretroviral treatment cause damage to the peripheral nervous system. To study HIV-induced peripheral nervous system (PNS) damage, a unique simian immunodeficiency virus (SIV)/pigtailed macaque model of HIV-PN that enabled detailed morphologic and functional evaluation of the somatosensory pathway throughout disease progression was developed. Studies in this model have demonstrated that SIV induces key pathologic features that closely resemble HIV-induced alterations, including inflammation and damage to the neuronal cell bodies in somatosensory ganglia and decreased epidermal nerve fiber density. Insights generated in the model include: finding that SIV alters the conduction properties of small, unmyelinated peripheral nerves; and that SIV impairs peripheral nerve regeneration. This review will highlight the major findings in the SIV-infected pigtailed macaque model of HIV-PN, and will illustrate the great value of a reliable large animal model to show the pathogenesis of this complex, HIV-induced disorder of the PNS.
Keywords: human immunodeficiency virus (HIV), Macaca nemestrina, macaque, peripheral nervous system (PNS), simian immunodeficiency virus (SIV)
Introduction
Peripheral neuropathy is currently the most common neurologic complication associated with HIV infection, with over one-third of HIV-infected individuals exhibiting abnormalities of the peripheral nervous system (PNS) (Ellis et al. 2010; Ghosh et al. 2012; McArthur et al. 2005). The most frequently reported clinical manifestation of HIV-associated peripheral neuropathy is distal sensory polyneuropathy (DSP), a progressive and often debilitating syndrome characterized by bilaterally symmetrical pain, numbness, and hypersensitivity that is most pronounced on the feet and lower legs (Keswani et al. 2002; Pardo et al. 2001). In severe cases, sensory abnormalities may extend above the knees, or involve the hands (Gonzalez-Duarte et al. 2008). While HIV-PN is not a life-threatening condition, symptomatic treatment with standard analgesic medications is often unsatisfactory. The relentless pain and paresthesias can be detrimental to patients' quality of life (Ellis et al. 2010; Krashin et al. 2012; McArthur 2012; McArthur et al. 2005).
Although the widespread use of effective combination antiretroviral therapy (cART) has drastically improved morbidity and mortality in HIV-infected individuals and reduced the incidence of other neurologic complications such as HIV dementia, the prevalence of HIV-PN remains high in both well-resourced and developing countries (Ellis et al. 2010; Ghosh et al. 2012; Smyth et al. 2007; Wiebe et al. 2011). It is now clear that several of the “d-class” nucleoside reverse transcriptase inhibitors (dNRTIs), including zacitabine, stavudine, and didanosine, are potently neurotoxic, and use of these agents to treat HIV can lead to antiretroviral toxic neuropathy (ATN), a syndrome clinically indistinguishable from primary HIV-PN (Moyle 2000; Simpson and Tagliati 1995). Exposure to protease inhibitors has also been proposed as a risk factor for the development of peripheral neuropathy, although this proposal remains a topic of debate (Ellis et al. 2008; Pettersen et al. 2006). Dose reduction or discontinuation of known neurotoxic antiretroviral drugs is often the first step in clinical management of neuropathic signs in HIV patients, and for many patients, results in symptomatic improvement (Gonzalez-Duarte et al. 2008; Wiebe et al. 2011). However, the frequency of DSP remains unacceptably high, despite declining use of dNRTIs, attesting to the ongoing importance of primary HIV-mediated PN, as well as contributions of other proposed risk factors such as advancing age, impaired immune status, metabolic abnormalities, and genetic predisposition (Canter et al. 2008; Evans et al. 2011; Kamerman et al. 2012b; Lichtenstein et al. 2005; Smyth et al. 2007).
The pathophysiologic mechanisms underlying HIV-PN are poorly understood. While the clinical features are consistent with a length-dependent “dying back” of sensory nerve fibers (Pardo et al. 2001), it is uncertain whether HIV induces damage primarily to the neuron cell body at the level of the dorsal root ganglia (DRG), or if the inciting pathologic change is primarily damage to the peripheral nerve fiber, either of which could ultimately result in a distal axonopathy. Furthermore, while we know that HIV is unable to infect neurons productively, it is unclear whether the neuronal injury arising in HIV-PN is due to a persistent release of neurotoxic products caused by activated macrophages and satellite glial cells (tumor necrosis factor-alpha, IL-1beta, chemokines); expressions of neurotoxic viral proteins, such as glycoprotein (gp)41, gp120, or Tat, by infected peri-neuronal cells; or an additive combination of these processes (Acharjee et al. 2010; Bhangoo et al. 2009; Hahn et al. 2008; Kamerman et al. 2012a; Melli et al. 2006; Smith 2011). These questions are difficult to study in HIV-infected individuals, due to the inability to repeatedly sample multiple components of the somatosensory pathway. The presence of confounding factors, such as treatment of the majority of HIV-infected individuals with cART and prevalent comorbid conditions also make study difficult. Thus, a robust animal model of HIV-PN is critical to dissect the underlying pathogenesis of this disorder and direct the development of new therapeutic strategies.
Simian Immunodeficiency Virus-Infected Macaque Model of HIV Peripheral Neuropathy
Simian immunodeficiency virus (SIV) infected macaques have been widely used to study host and viral aspects of HIV infection in humans. The appropriateness of such models is underpinned by the close resemblance of macaque immune responses during SIV infection to those of HIV-infected individuals; and extensive homology shared by SIV and HIV, including the binding of SIV envelope glycoprotein gp120 to the host receptors CD4 and CCR5 for viral entry (Sina et al. 2011). Because SIV models vary widely with respect to progression to fulminant AIDS and development of neurologic disease, a refined pigtailed macaque model was developed to study SIV-induced CNS disease efficiently. This model, which entails simultaneous intravenous inoculation of pigtailed macaques with both the neurovirulent molecular SIV clone SIV/17E-Fr and the immunosuppresive virus strain SIV/DeltaB670, results in an accelerated and highly reproducible progression to AIDS within 84 days post-inoculation, with approximately 90% of SIV-infected macaques also developing encephalitis within this time frame (Flaherty et al. 1997; Mankowski et al. 2002; Mankowski et al. 1997). The encephalitis observed in these animals shares many morphologic similarities with HIV-associated encephalitis, including characteristic multifocal perivascular infiltrates of macrophages and the formation of multinucleated giant cells bearing replicating viruses (McArthur et al. 2005; Zink et al. 1997). Furthermore, this model has been useful to study numerous translationally relevant aspects of SIV-associated CNS disease, including the effectiveness of neuroprotective agents, relevance of biomarkers in the CSF, characterization of neuroprotective MHC class I alleles, persistence of neuroinflammation with suppressive cART, and identification of CNS latency reservoirs (Clements et al. 2002; Follstaedt et al. 2008; Mankowski et al. 2008; Mankowski et al. 2004; Meulendyke et al. 2012; Zink et al. 2010).
Recapitulating the neuropathologic features of HIV-PN in rodent models has proved challenging. Because these species are not susceptible to infection by HIV, or a closely analogous lentivirus, in vitro rodent studies of HIV-PN have required the creation of transgenic mice expressing HIV gp120 in perineuronal cell populations, or direct inoculation of viral proteins into the peripheral nerves (Höke 2012; Keswani et al. 2006; Zheng et al. 2011). To determine whether the SIV/pigtailed macaque model could serve as a reliable model for HIV-PN, key components of the sensory pathway, including samples of skin from the distal leg, sural, and sciatic nerves, and somatosensory ganglia including trigeminal and dorsal root ganglia, were collected from SIV-inoculated pigtailed macaques and examined at the morphologic and molecular level.
Lesions in Somatosensory Ganglia of Simian Immunodeficiency Virus-Infected Macaques
Studies examining the morphologic changes in both the trigeminal ganglia and lumbar dorsal root ganglia of SIV-infected pigtailed macaques revealed lesions that closely resemble those described in patients with HIV-PN (Keswani et al. 2002; Pardo et al. 2001). Hematoxylin and eosin, (H and E)-stained sections of trigeminal ganglia, were evaluated for microscopic lesions by two pathologists and the severity of ganglionitis was scored as none, mild, moderate, or severe based on the degree and presence of infiltrating mononuclear cells, neuronophagia, and neuronal loss. As illustrated in Figure 1, the trigeminal ganglia from SIV-infected macaques contained multifocal perineuronal inflammatory cells, including macrophages and lymphocytes. In animals with moderate to severe ganglionitis there was evidence of neuronophagia, characterized by macrophages within the neuronal compartment of degenerate neurons, as well as nodules of Nageotte, with neuron cell bodies replaced by compact aggregates of satellite glial cells and macrophages (Figure 1, C and D) (Laast et al. 2007). Similar changes were also observed in the lumbar dorsal root ganglia, which house the neuronal cell bodies of sensory nerves in the lower limbs. A histologic grading scheme was not employed in this study (Laast et al. 2011). Neuronal loss in the trigeminal and dorsal root ganglia was evaluated using the area fraction fractionator technique, a method to estimate neuron density. In both locations, significant neuronal loss was identified in SIV-infected macaques at 12 weeks post-infection (p.i.) when compared with uninfected control animals (Figure 1E) (Laast et al. 2011; Laast et al. 2007). In the trigeminal ganglia, decline in neuronal density was inversely proportional with the extent of macrophage infiltration as measured by CD68 immunostaining (Figure 1F) (Laast et al. 2007). Similar patterns of neuropathologic changes and macrophage infiltration into the DRG have been reported in a SIV-infected, CD8 T cell-depleted rhesus macaque model (Macaca mulatta) (Burdo et al. 2012).
Figure 1.
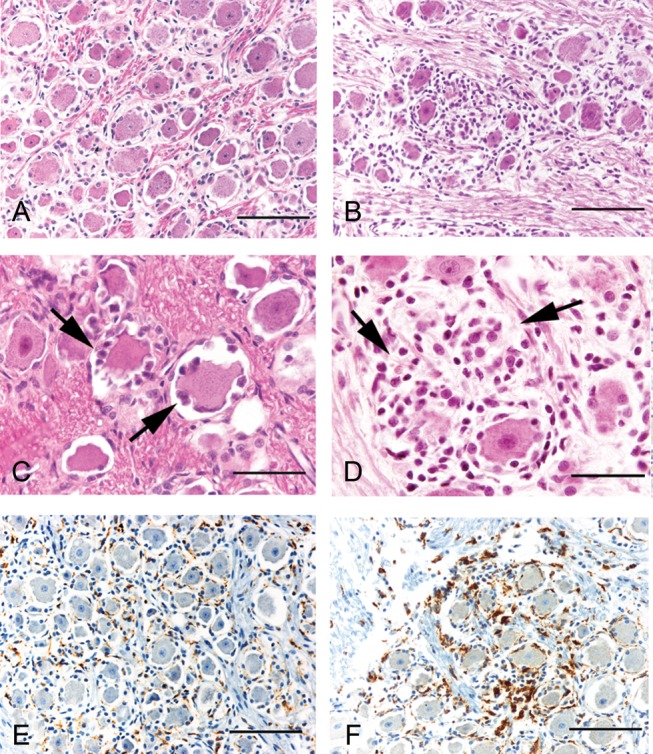
Trigeminal ganglionitis (TG) in SIV-infected pigtailed macaques. Hematoxylin and eosin-stained sections of trigeminal ganglia from an uninfected control macaque (A) and a SIV-infected macaque (B). TG showed evidence of neuronophagia, characterized by macrophages within the neuronal compartment of degenerate neurons (C), and nodules of Nageotte, with neuron cell bodies replaced by aggregates of satellite glial cells and macrophages (D). Sections of TG from control animals showed scattered CD68-immunopositive resident macrophages (E). The TG of SIV-infected animals had increased immunostaining for CD68, likely reflecting a combination of resident macrophage activation and infiltration of circulating macrophages (F). Similar changes were also observed in the DRG (not pictured). Scale bars = (A, B, E, F) 100 µm; (C, D) 50 µm. Reprinted with permission from Laast et al. (2007).
Changes in Levels of Viral Replication and Inflammatory Markers Over the Course of Simian Immunodeficiency Virus Infection
To determine the temporal dynamics of SIV replication and associated neuroinflammatory changes in the PNS during disease progression, somatosensory ganglia from SIV-infected macaques euthanized during acute, asymptomatic, and terminal stages of infection were examined. In the trigeminal ganglia, SIV RNA was detected by in situ hybridization in two of six animals euthanized at seven and ten days p.i., but none (zero of six) of the animals examined at 21 days p.i. showed SVI RNA in the trigeminal ganglia. In contrast, at 56 days p.i., viral replication was only detected in one of six infected animals (Laast et al. 2007). This pattern, in which low levels of SIV replication occurs during acute infection in the somatosensory ganglia, followed by suppression for 21 days, and subsequent recrudescence at 56 days, parallels the trend reported for SIV replication in the CNS of SIV-infected macaques (Clements et al. 2005). In the dorsal root ganglia, SIV replication was measured by qRT-PCR using SIV gag region primers and a probe. This was found to be relatively constant among animals euthanized at six through twelve weeks p.i., demonstrating that SIV replication in the DRG is continuous throughout the late asymptomatic and terminal stages of infection. At the terminal disease time point, approximately twelve weeks p.i., both the TG and DRG of SIV-infected macaques contained abundant SIV-infected cells; this abundance was demonstrated by immunostaining for the transmembrane glycoprotein gp41 (Laast et al. 2011; Laast et al. 2007). Similar to the pattern of HIV replication in human ganglia (Esiri et al. 1993), SIV-infected cells were observed among infiltrating mononuclear cells, and the perineuronal compartment. To identify infected cell types, ganglia were double immunostained for gp41 and the macrophage marker Iba-1. Laser confocal microscopy revealed clear co-localization of SIV gp41 within Iba-1 positive macrophages, confirming that macrophages were the predominant infected cell type (Laast et al. 2011; Laast et al. 2007).
To characterize the severity and composition of inflammatory changes in the somatosensory ganglia during SIV infection, immunohistochemistry and quantitative image analysis on sections of TG and DRG from animals euthanized at multiple time points post-SIV inoculation were performed. Immunostaining revealed a resident population of CD68-positive macrophages distributed diffusely throughout the perineuronal compartment in the uninfected controls and SIV-infected macaques. In the SIV-infected animals, this resident population of perineuronal endogenous macrophages stained more intensely than the controls and was accompanied by additional, randomly distributed infiltrating CD68 positive cells (Figure 1, E and F). Although this immunostaining pattern likely represented concurrent activation of resident macrophages and infiltration by circulating macrophages, there are currently no available immunologic markers that can distinguish between these two populations. Compared to uninfected control animals, the total CD68 immunostaining was found to be significantly elevated in the DRG of SIV-infected macaques by six weeks p.i., and remained elevated at eight and twelve weeks p.i. (Figure 2A) (Laast et al. 2011). CD68 immunostaining was also significantly higher in the trigeminal ganglia of SIV-infected macaques at terminal stages compared to the control animals (Laast et al. 2007). At terminal stages, a strong correlation between the amounts of CD68 immunostaining in the trigeminal ganglia and dorsal root ganglia (p = 0.03, r = 0.64, Spearman's rank coefficient) was found. Contrasted with the increase in macrophages, there was modest decline in the number of cytotoxic T cells (TIA-1 + , CD3+) in the trigeminal ganglia of SIV-infected animals versus the uninfected controls (Laast et al. 2007).
Figure 2.

Alterations in CD68 and GFAP immunostaining in the dorsal root ganglia during SIV infection. (A) Scatter plot depicting the amount of CD68 immunostaining in the DRG of uninfected control macaques (circles) and SIV-infected macaques (triangles) demonstrates a statistically significant increase in macrophage activation and infiltration in the DRG of SIV-infected animals by six weeks p.i. with sustained, elevated levels throughout the twelve week course of infection. (P = 0.008, analysis of variance). (B) GFAP immunostaining of satellite glial cells was similarly increased by six and eight weeks p.i., but declined to control animal levels by twelve weeks p.i. (P = 0.001, analysis of variance). ROI = region of interest. Reprinted with permission from Laast et al. (2011).
Several recent studies have proposed an integral role for glial cell activation in the induction and maintenance of neuropathic pain, including painful HIV-PN (Capuano et al. 2009; Gosselin et al. 2010; Shi et al. 2012; Villa et al. 2010). To determine whether satellite glial cells were immune activated during SIV-infection, DRG sections were immunostained for glial fibrillary acid protein (GFAP) followed by digital image analysis to measure GFAP expression. GFAP expression was significantly increased in SIV-infected macaques at six weeks p.i. and remained upregulated at eight weeks p.i. However, in animals euthanized at twelve weeks p.i., the median GFAP staining had declined to the control animal level (Figure 2B) (Laast et al. 2011). These compelling results suggest that satellite glial cell activation plays a key role in the pathogenesis of SIV-induced damage to the DRG. Further studies in the SIV-infected macaque model may show the functional significance of SGC activation during disease progression, including its association with neuronal viability and sensitivity.
Simian Immunodeficiency Virus-Induced Functional Alterations in Peripheral Nerves
In contrast to the changes observed in the somatosensory ganglia, little morphologic evidence of neuritis or damage to myelinated fibers was found by examining plastic-embedded, toluidine blue-stained sections of sural and peroneal nerves (Laast et al. 2011). A similar paucity of lesions in sural nerve biopsies has been reported in HIV neuropathy (Gonzalez-Duarte et al. 2008; Gonzalez-Duarte et al. 2007). Possible explanations for this observation include the extensive length of peripheral nerves and multifocal distribution of inflammatory changes, as well the primary involvement of small, unmyelinated fibers. Given the lack of widespread inflammatory lesions in the peripheral nerve, the focus of the investigation of peripheral nerves was concentrated on alterations in neurophysiology, the correlations between these changes, and SIV-induced pathology in the DRG.
Because current clinical methods of measuring peripheral nerve conduction velocity (CV) are relatively insensitive to changes in small unmyelinated fibers, these electrophysiologic studies are often of limited diagnostic value in patients with HIV-PN (Gonzales-Duarte 2007, 2008). To circumvent this limitation, an ex vivo teased fiber technique used to measure CV in single, unmyelinated fibers in the sural nerves from control and SIV-infected macaques was employed. Electrophysiology experiments demonstrated a significant decrease in mean C-fiber conduction velocity in SIV-infected macaques by twelve weeks p.i., during late stages of the disease (Figure 3A). Additionally, as shown in Figure 3B, there was strong inverse correlation between mean C-fiber CV of SIV-infected animals and the degree of CD68 expression in the DRG, indicating that the extent of macrophage infiltration in the DRG corresponded closely with functional changes in C-fiber conduction properties (Laast et al. 2011).
Figure 3.

Decreased C-fiber conduction velocity in SIV-infected macaques strongly correlated with CD68 expression in the lumbar DRG. (A) Mean sural nerve C-fiber CV was significantly slower in SIV-animals evaluated at twelve weeks p.i. when compared to uninfected controls or SIV-infected animals evaluated at six or eight weeks p.i. (P = 0.003, analysis of variance). (B) Comparing the mean C-fiber CV of SIV-infected animals to corresponding levels of CD68 immunostaining in the lumbar DRG demonstrated a strong inverse correlation between slowing of CV in C-fibers and macrophage activation and infiltration in the DRG (P = 0.006, r = -0.97, Pearson correlation coefficient). Reprinted with permission from Laast et al. (2011).
Epidermal Nerve Fiber Density Declines with Simian Immunodeficiency Virus Infection
Skin biopsies have proven to be an invaluable tool in the clinical diagnosis of small fiber neuropathies, and have largely replaced more invasive sural nerve biopsies for the assessment of conditions including diabetic neuropathy and HIV-PN (Ebenezer et al. 2007; Lauria and Lombardi 2012; McArthur 2012). As illustrated in Figure 4A, immunostaining of skin sections with pan-neuronal marker PGP9.5 allows for the visualization and quantification of epidermal nerve fiber density (Wendelschafer-Crabb et al. 2006). In HIV-infected patients, decreased ENF density in the skin of the distal leg has been shown to correlate with lower CD4 counts, higher viral loads, and the presence of neuropathic pain (Polydefkis 2006; Polydefkis 2002). To examine whether a parallel decline in ENF density occurs in SIV-infected macaques, cutaneous biopsies from footpads were collected at necropsy, immunostained for PGP9.5, and analyzed by quantitative image analysis. Compared with uninfected control animals, SIV-infected animals exhibited a progressive decline in ENF density throughout the disease, resulting in a significant decrease by eight weeks p.i. and a marked reduction at twelve weeks p.i. (Figure 4, B and C) (Laast et al. 2011). As a point of note, rhesus macaques inoculated with the same combination of viruses do not develop a significant decline in ENF density, suggesting that additional host factors are involved in PNS damage (unpublished, in-house data).
Figure 4.
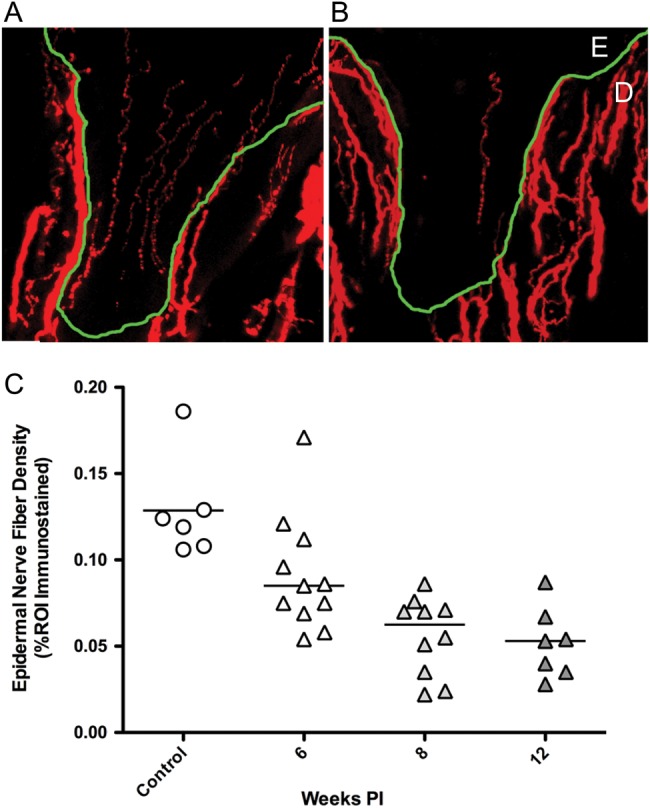
Epidermal nerve fiber density is reduced with SIV-infection. PGP9.5 immunostained sections of footpad skin from an uninfected control (A) and SIV-infected macaques (B) showed marked decline in ENF density. Nerve fibers (red) are present in the epidermis (E) and dermis (D), with the dermal-epidermal junction traced in green. (C) Measurement of ENF density demonstrated reduced ENF density in SIV-infected macaques (triangles) compared to uninfected controls (circles), with significant decrease developing by eight weeks p.i., then declining further at the twelve week time point. (P < 0.001, analysis of variance). Reprinted with permission from Laast et al. 2011).
Simian Immunodeficiency Virus-induced Impairment of Neurovascular Regeneration
In addition to simulating HIV-induced damage to the peripheral nervous system, the SIV-infected macaque model has also been used to study the impact of infection on the capacity of peripheral nerves to regenerate following injury. Similar to techniques previously described in humans (Rajan et al. 2003), an excisional intracutaneous axotomy model in macaques to study both the normal features of nerve regeneration in this species and the effect of SIV infection on this process was utilized. Briefly, cutaneous axotomies were performed at two-week intervals following SIV inoculation using 3mm circular skin punches to transect the epidermis and dermis. On the 70th day after the initial axotomy, all 3mm punch incision sites were harvested using a 5mm skin punch, producing skin samples containing excision sites from 14, 28, 42, 56, and 70 days post-axotomy. Samples were then immunostained for the pan-neuronal marker PGP9.5, and the length of the regenerating epidermal nerve fiber sprouts within the axotomy site was compared to the length of normal epidermal nerve fibers located outside of the initial axotomy zone. In both SIV-infected and uninfected macaques, epidermal reinnervation was rapid and complete by 56 days post-axotomy, which is in contrast to the slower, incomplete reinnervation observed in humans following this technique (Rajan et al. 2003). However, in SIV-infected animals epidermal nerve fiber regeneration was delayed at every time point post-axotomy when compared to uninfected controls (Figure 5) (Ebenezer et al. 2009).
Figure 5.

SIV infection results in delayed epidermal nerve fiber regeneration following cutaneous axotomy. When compared to uninfected control macaques, ENF length was significantly lower in SIV-infected animals at days 14 (P = 0.01, Mann-Whitney test) and 70 post axotomy, indicative of delayed ENF regrowth at the axotomy site. Reprinted with permission from Ebenezer et al. (2012).
Because Schwann cells play a crucial role in guiding axonal regrowth following injury (Albers and Davis 2007; Bentley and Lee 2000; Song et al. 2006; Thompson and Buettner 2006), the degree of Schwann cell migration into denervated epidermis by immunostaining for Schwann cell marker p75 nerve growth factor receptor was assessed. In both control and SIV-infected animals, Schwann cell density was highest at the earliest time points (post-axotomy), and declined at a similar rate in both groups. At every time point post-axotomy, the SIV-infected animals consistently had lower mean Schwann cell density measurements than the uninfected controls (Ebenezer et al. 2009). It remains to be determined whether the Schwann cell density decrease detected post-axotomy in SIV-infected macaques represents impaired Schwann cell proliferation, reduced migration, or a combination of these factors.
Another key contributor to axonal regeneration following injury is the regrowth of blood vessels, which travel in close morphologic approximation with nerves forming a well-organized network of neurovascular bundles. Further studies investigating neurovascular repair in the pigtailed macaque model revealed that while SIV-infection does not affect blood vessel regrowth at cutaneous biopsy sites, expression of vascular endothelial growth factor (VEGF) around regenerating neurovascular units, measured by immunostaining, is lower in SIV-infected animals compared to controls. Additionally, a strong positive correlation was demonstrated between VEGF expression and the extent of ENF regeneration (Ebenezer et al. 2012). Downregulation of VEGF expression may alter the perivascular extracellular matrix such that it is less conducive to nerve regrowth. Taken together, these results indicate that SIV-infection leads to deficient epidermal nerve fiber regeneration and that this impairment is mediated, at least in part, by altered Schwann cell migration and decreased VEGF expression along neurovascular tracts. In addition, these studies illustrate that the macaque model can accelerate study of human nerve regrowth biology.
Conclusion
SIV-infected pigtailed macaques share key features of HIV-associated peripheral neuropathy, making this model invaluable for understanding the complex pathophysiologic mechanisms of HIV-PN, and showing new therapeutic targets. By sampling and examining key components of the peripheral nervous system at multiple time points throughout SIV infection, it has been found that the virus infects and immune-activates perineuronal cells of the somatosensory ganglia during early stages of infection. This sets the scene for insidious neuronal damage that becomes evident later in the disease by decreased neuronal density in somatosensory ganglia, altered C-fiber conduction properties, and decreased epidermal nerve fiber density. SIV-infection also impairs the regenerative capacity of small epidermal nerve fibers, a factor that will be imperative to address when designing therapeutic strategies for HIV-PN. Another notable outcome of these studies in the pigtailed macaque model is the development and refinement of various techniques to assess peripheral nervous system integrity. These tools are applicable to studies of HIV-PN, and will be instrumental for studies involving non-human primate models of other clinically relevant small fiber neuropathies including those associated with diabetes mellitus and cancer chemotherapy.
Acknowledgments
These studies were supported by NIH NS055651, NIH MH070306, National Center for Research Resources and the Office of Research Infrastructure Programs P40 OD013117, and NIH T32 OD011089.
References
- Acharjee S, Noorbakhsh F, Stemkowski PL, Olechowski C, Cohen EA, Ballyani K, Kerr B, Pardo C, Smith PA, Power C. HIV-1 viral protein R causes peripheral nervous system injury associated with in vivo neuropathic pain. FASEB J. 2010;24:4343–4353. doi: 10.1096/fj.10-162313. [DOI] [PubMed] [Google Scholar]
- Albers KM, Davis BM. The skin as a neurotrophic organ. NRO. 2007;13:371–382. doi: 10.1177/10738584070130040901. [DOI] [PubMed] [Google Scholar]
- Bentley CA, Lee KF. p75 is important for axon growth and schwann cell migration during development. Journal of Neurosci. 2000;20:7706–7715. doi: 10.1523/JNEUROSCI.20-20-07706.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bhangoo SK, Ripsch MS, Buchanan DJ, Miller RG, White FA. Increased chemokine signaling in a model of HIV1-associated peripheral neuropathy. Molecular Pain. 2009;5 doi: 10.1186/1744-8069-5-48. 48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burdo TH, Orzechowski K, Knight HL, Miller AD, Williams K. Dorsal Root Ganglia Damage in SIV-Infected Rhesus Macaques: An Animal Model of HIV-Induced Sensory Neuropathy. Am J pathol. 2012;180:1–8. doi: 10.1016/j.ajpath.2011.12.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Canter JA, Haas DW, Kallianpur AR, Ritchie MD, Robbins GK, Schafer RW, Clifford DB, Murdock DG, Hulgan T. The mitochondrial pharmacogenomics of haplogroup T: MTND2*LHON4917G and antiretroviral therapy-associated peripheral neuropathy. Pharmacogenomics Journal. 2008;8:71–77. doi: 10.1038/sj.tpj.6500470. [DOI] [PubMed] [Google Scholar]
- Capuano A, De Corato A, Lisi L, Tringali G, Navarra P, Dello Russo P. Proinflammatory-activated trigeminal satellite cells promote neuronal sensitization: Relevance for migraine pathology. Molecular Pain. 2009;5 doi: 10.1186/1744-8069-5-43. 43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clements JE, Babas T, Mankowski JL, Suryanarayana K, Piatak M, Tarwater PK, Lifson JD, Zink MC. The central nervous system as a reservoir for simian immunodeficiency virus (SIV): Steady-state levels of SIV DNA in brain from acute through asymptomatic infection. J Infect Dis. 2002;186:905–913. doi: 10.1086/343768. [DOI] [PubMed] [Google Scholar]
- Clements JE, Li M, Gama L, Bullock B, Carruth CM, Mankowski JL, Zink MC. The central nervous system is a viral reservoir in simian immunodeficiency virus–infected macaques on combined antiretroviral therapy: a model for human immunodeficiency virus patients on highly active antiretroviral therapy. J Neuroviro. 2005;11:180–189. doi: 10.1080/13550280590922748-1. [DOI] [PubMed] [Google Scholar]
- Ebenezer G, Laast V, Dearman B, Hauer P., Tarwater PM, Adams RJ, Zink MC, McArthur JC, Mankowski JL. Altered cutaneous nerve regeneration in a simian immunodeficiency virus/macaque intracutaneous axotomy model. J Comp Neurol. 2009;514:272–283. doi: 10.1002/cne.22019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ebenezer GJ, Hauer P, Gibbons C, McArthur JC, Polydefkis M. Assessment of epidermal nerve fibers: A new diagnostic and predictive tool for peripheral neuropathies. J Neuropatho Exp Neurol. 2007;66:1059–1073. doi: 10.1097/nen.0b013e31815c8989. [DOI] [PubMed] [Google Scholar]
- Ebenezer GJ, McArthur JC, Polydefkis M, Dorsey JL, O'Donnel R, Hauer P, Adams RJ, Mankowski JL. SIV-induced impairment of neurovascular repair: a potential role for VEGF. J Neuroviro. 2012;18:222–230. doi: 10.1007/s13365-012-0102-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ellis RJ, Marquie-Beck J, Delaney P, Alexander T, Clifford DB, McArthur JC, Simpson DM, Ake C, Collier AC, Gelman BS, McCutchan JA, Morgello S, Grant I CHARTER Study Group. Human immunodeficiency virus protease inhibitors and risk for peripheral neuropathy. Ann Neurol. 2008;64:566–572. doi: 10.1002/ana.21484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ellis RJ, Rosario D, Clifford DB, McArthur JC, Simpson D, Alexander T, Gelman BS, Vaida F, Collier A, Marra CM, Ances B, Atkinson JH, Dworkin RH, Morgello S, Grant I CHARTER Study Group. Continued high prevalence and adverse clinical impact of human immunodeficiency virus-associated sensory neuropathy in the era of combination antiretroviral therapy: The CHARTER Study. Arch Neurol. 2010;67:552–558. doi: 10.1001/archneurol.2010.76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Esiri MM, Morris CS, Millard PR. Sensory and sympathetic ganglia in HIV-1 infection: Immunocytochemical demonstration of HIV-1 viral antigens, increased MHC class II antigen expression and mild reactive inflammation. J Neurol Sci. 1993;114:178–187. doi: 10.1016/0022-510x(93)90295-a. [DOI] [PubMed] [Google Scholar]
- Evans SR, Ellis RJ, Chen H, Tzu-min Y, Lee AJ, Schiffito G, Wu K, Bosch RJ, McArthur JC, Simpson DM, Clifford DB. Peripheral neuropathy in HIV: Prevalence and risk factors. AIDS. 2011;25:919–928. doi: 10.1097/QAD.0b013e328345889d. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flaherty MT, Hauer DA, Mankowski JL, Zink MC, Clements JE. Molecular and biological characterization of a neurovirulent molecular clone of simian immunodeficiency virus. J Virol. 1997;71:5790–5798. doi: 10.1128/jvi.71.8.5790-5798.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Follstaedt SC, Barber SA, Zink MC. Mechanisms of minocycline-induced suppression of simian immunodeficiency virus encephalitis: Inhibition of apoptosis signal-regulating kinase 1. J Neurovirol. 2008;14:376–388. doi: 10.1080/13550280802199898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghosh S, Chandran A, Jansen JP. Epidemiology of HIV-related neuropathy: A systematic literature review. AIDS Res Hum Retroviruses. 2012;28:36–48. doi: 10.1089/AID.2011.0116. [DOI] [PubMed] [Google Scholar]
- Gonzalez-Duarte A, Cikurel K, Simpson DM. Managing HIV peripheral neuropathy. Curr HIV/AIDS Rep. 2007;4:114–118. doi: 10.1007/s11904-007-0017-6. [DOI] [PubMed] [Google Scholar]
- Gonzalez-Duarte A, Robinson-Papp J, Simpson DM. Diagnosis and management of HIV-associated neuropathy. Neurol Clin. 2008;26:821–32. doi: 10.1016/j.ncl.2008.04.001. [DOI] [PubMed] [Google Scholar]
- Gosselin R, Suter M, Ji R, Decosterd I. Glial cells and chronic pain. Neuroscientist. 2010;16:519–531. doi: 10.1177/1073858409360822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hahn K, Robinson B, Anderson C, Li W, Pardo CA, Morgello S, Simpson D, Nath A. Differential effects of HIV infected macrophages on dorsal root ganglia neurons and axons. Exp Neurol. 2008;210:30–40. doi: 10.1016/j.expneurol.2007.06.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Höke A. Animal models of peripheral neuropathies. Neurotherapeutics. 2012;9:262–269. doi: 10.1007/s13311-012-0116-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kamerman PR, Moss PJ, Weber J, Wallace VCJ, Rice ASC, Huang W. Pathogenesis of HIV-associated sensory neuropathy: Evidence from in vivo and in vitro experimental models. J Peripher Nerv Syst. 2012a;17:19–31. doi: 10.1111/j.1529-8027.2012.00373.x. [DOI] [PubMed] [Google Scholar]
- Kamerman PR, Wadley AL, Cherry CL. HIV-associated sensory neuropathy: Risk factors and genetics. Curr Pain Headache Rep. 2012b;16:226–236. doi: 10.1007/s11916-012-0257-z. [DOI] [PubMed] [Google Scholar]
- Keswani SC, Jack C, Zhou C, Höke A. Establishment of a rodent model of HIV-associated sensory neuropathy. J Neurol Sci. 2006;26:10299–10304. doi: 10.1523/JNEUROSCI.3135-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keswani SC, Pardo CA, Cherry CL, Höke A, McArthur JC. HIV-associated sensory neuropathies. AIDS. 2002;16:2105–2117. doi: 10.1097/00002030-200211080-00002. [DOI] [PubMed] [Google Scholar]
- Krashin DL, Merrill JO, Trescot AM. Opioids in the management of HIV-related pain. Pain Physician. 2012;15 ES157–168. [PubMed] [Google Scholar]
- Laast VA, Pardo CA, Tarwater PM, Queen SE, Reinhart TA, Ghosh M, Adams RJ, Zink MC, Mankowski JL. Pathogenesis of simian immunodeficiency virus-induced alterations in macaque trigeminal ganglia. J Neuropatho Exp Neurol. 2007;66:26–34. doi: 10.1097/nen.0b013e31802c398d. [DOI] [PubMed] [Google Scholar]
- Laast VA, Shim B, Johanek LM, Dorsey JL, Hauer PE, Pardko CA, McArthur JC, Ringkamp M, Mankowski JL. Macrophage-mediated dorsal root ganglion damage precedes altered nerve conduction in SIV-infected macaques. Am J Pathol. 2011;179:2337–2345. doi: 10.1016/j.ajpath.2011.07.047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lauria G, Lombardi R. Skin biopsy in painful and immune-mediated neuropathies. J Peripher Nerv Syst. 2012;17(Suppl 3):38–45. doi: 10.1111/j.1529-8027.2012.00430.x. [DOI] [PubMed] [Google Scholar]
- Lichtenstein KA, Armon C, Baron A, Moorman AC, Wood KC, Holmberg SD HIV Outpatient Study Investigators. Modification of the incidence of drug-associated symmetrical peripheral neuropathy by host and disease factors in the HIV outpatient study cohort. Clin Infect Dis. 2005;40:148–157. doi: 10.1086/426076. [DOI] [PubMed] [Google Scholar]
- Mankowski JL, Clements JE, Zink MC. Searching for clues: Tracking the pathogenesis of human immunodeficiency virus central nervous system disease by use of an accelerated, consistent simian immunodeficiency virus macaque model. J Infect Dis. 2002;186(Suppl) doi: 10.1086/344938. S199–208. [DOI] [PubMed] [Google Scholar]
- Mankowski JL, Flaherty MT, Spelman JP, Hauer DA, Didier PJ, Amedee AM, Murphey-Corb M., Kirstein LM, Munoz A, Clements JE, Zink MC. Pathogenesis of simian immunodeficiency virus encephalitis: Viral determinants of neurovirulence. J Virol. 1997;71:6055–6060. doi: 10.1128/jvi.71.8.6055-6060.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mankowski JL, Queen SE, Clements JE, Zink MC. Cerebrospinal fluid markers that predict SIV CNS disease. J Neuroimmunol. 2004;157:66–70. doi: 10.1016/j.jneuroim.2004.08.031. [DOI] [PubMed] [Google Scholar]
- Mankowski JL, Queen SE, Fernandez CS, Tarwater PM, Karper JM, Adams RJ, Kent SJ. Natural host genetic resistance to lentiviral CNS disease: A neuroprotective MHC class I allele in SIV-infected macaques. PloS one. 2008;3 doi: 10.1371/journal.pone.0003603. e3603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McArthur JC. Painful small fiber neuropathies. Continuum (Minneap Minn) 2012;18:106–25. doi: 10.1212/01.CON.0000411570.79827.25. [DOI] [PubMed] [Google Scholar]
- McArthur JC, Brew BJ, Nath A. Neurological complications of HIV infection. Lancet Neurol. 2005;4:543–555. doi: 10.1016/S1474-4422(05)70165-4. [DOI] [PubMed] [Google Scholar]
- Melli G, Keswani SC, Fischer A, Chen W, Höke A. Spatially distinct and functionally independent mechanisms of axonal degeneration in a model of HIV-associated sensory neuropathy. Brain. 2006;129:1330–1338. doi: 10.1093/brain/awl058. [DOI] [PubMed] [Google Scholar]
- Meulendyke KA, Pletnikov MV, Engle EL, Tarwater PM, Graham DR, Zink MC. Early minocycline treatment prevents a decrease in striatal dopamine in an SIV model of HIV-associated neurological disease. J Neuroimmune Pharmacol. 2012;7:454–464. doi: 10.1007/s11481-011-9332-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moyle G. Clinical manifestations and management of antiretroviral nucleoside analog-related mitochondrial toxicity. Clin Ther. 2000;22 doi: 10.1016/S0149-2918(00)80064-8. 911–936; discussion 898. [DOI] [PubMed] [Google Scholar]
- Pardo CA, McArthur JC, Griffin JW. HIV neuropathy: Insights in the pathology of HIV peripheral nerve disease. J Peripher Nerv Syst. 2001;6:21–27. doi: 10.1046/j.1529-8027.2001.006001021.x. [DOI] [PubMed] [Google Scholar]
- Pettersen JA, Jones G, Worthington C, Krentz HB, Keppler OT, Höke A, Gill MJ, Power C. Sensory neuropathy in human immunodeficiency virus/acquired immunodeficiency syndrome patients: Protease inhibitor-mediated neurotoxicity. Ann Neuro. 2006;59:816–824. doi: 10.1002/ana.20816. [DOI] [PubMed] [Google Scholar]
- Polydefkis M. Skin biopsy findings predict development of symptomatic neuropathy in patients with HIV. Nat Clin Pract Neurol. 2006;2:650–651. doi: 10.1038/ncpneuro0353. [DOI] [PubMed] [Google Scholar]
- Polydefkis MJ. Peripheral neuropathy and HIV. Hopkins HIV Rep. 2002;14:6–7. [PubMed] [Google Scholar]
- Rajan B, Polydefkis M, Hauer P, Griffin JW, McArthur JC. Epidermal reinnervation after intracutaneous axotomy in man. J Comp Neurol. 2003;457:24–36. doi: 10.1002/cne.10460. [DOI] [PubMed] [Google Scholar]
- Shi Y, Gelman BB, Lisinicchia JG, Tang S-J. Chronic-pain-associated astrocytic reaction in the spinal cord dorsal horn of human immunodeficiency virus-infected patients. J Neurol Sci. 2012;32:10833–10840. doi: 10.1523/JNEUROSCI.5628-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simpson DM, Tagliati M. Nucleoside analogue-associated peripheral neuropathy in human immunodeficiency virus infection. J Acquir Immune Defic Syndr Hum Retrovirol. 1995;9:153–161. [PubMed] [Google Scholar]
- Sina ST, Ren W, Cheng-Mayer C. Coreceptor use in nonhuman primate models of HIV infection. J Transl Med. 2011;9(Suppl 1) doi: 10.1186/1479-5876-9-S1-S7. S7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith HS. Treatment considerations in painful HIV-related neuropathy. Pain Physician. 2011;14 E505–524. [PubMed] [Google Scholar]
- Smyth K, Affandi JS, McArthur JC, Bowtell-Harris C, Mijch AM, Watson K, Costello K, Woolley IL, Price P, Wesselingh SL, Cherry CL. Prevalence of and risk factors for HIV-associated neuropathy in Melbourne, Australia 1993-2006. HIV Med. 2007;8:367–373. doi: 10.1111/j.1468-1293.2007.00478.x. [DOI] [PubMed] [Google Scholar]
- Song XY, Zhou FH, Zhong JH, Wu LL, Zhao XF. Knockout of p75(NTR) impairs re-myelination of injured sciatic nerve in mice. J Neurochem. 2006;96:833–842. doi: 10.1111/j.1471-4159.2005.03564.x. [DOI] [PubMed] [Google Scholar]
- Thompson DM, Buettner HM. Neurite outgrowth is directed by schwann cell alignment in the absence of other guidance cues. Ann Biomed Eng. 2006;34:161–168. doi: 10.1007/s10439-005-9013-4. [DOI] [PubMed] [Google Scholar]
- Villa G, Fumagalli M, Verderio C, Abbrachi MP, Ceruti S. Expression and contribution of satellite glial cells purinoceptors to pain transmission in sensory ganglia: An update. Neuron Glia Biol. 2010;6:31–42. doi: 10.1017/S1740925X10000086. [DOI] [PubMed] [Google Scholar]
- Wendelschafer-Crabb G, Kennedy WR, Walk D. Morphological features of nerves in skin biopsies. J Neurol Sci. 2006;242:15–21. doi: 10.1016/j.jns.2005.11.010. [DOI] [PubMed] [Google Scholar]
- Wiebe LA, Phillips TJ, Li JM, Allen JA, Shetty K. Pain in HIV: An evolving epidemic. J Pain. 2011;12:619–624. doi: 10.1016/j.jpain.2011.03.006. [DOI] [PubMed] [Google Scholar]
- Zheng W, Ouyang H, Zheng X, Liu S, Mata M, Fink DJ, Hao S. Glial TNFα in the spinal cord regulates neuropathic pain induced by HIV gp120 application in rats. Mol Pain. 2011;7 doi: 10.1186/1744-8069-7-40. 40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zink MC, Amedee AM, Mankowski JL, Craig LE, Didier P, Carter DL, Munoz A, Murphy-Corb M, Clements JE. Pathogenesis of SIV encephalitis. Selection and replication of neurovirulent SIV. Am J Pathol. 1997;151:793–803. [PMC free article] [PubMed] [Google Scholar]
- Zink MC, Brice AK, Kelly KM, Queen SE, Gama L, Li M, Adams RJ, Bartizal C, Varrone J, Rabi SA, Graham DR, Tarwater PM, Mankowski JL, Clements JE. Simian immunodeficiency virus-infected macaques treated with highly active antiretroviral therapy have reduced central nervous system viral replication and inflammation but persistence of viral DNA. J Infect Dis. 2010;202:161–170. doi: 10.1086/653213. [DOI] [PMC free article] [PubMed] [Google Scholar]