

Abstract
Pathogenic Yersinia species employ the Ysc-Yop type III secretion system (T3SS) encoded by a highly conserved pYV virulence plasmid to export the virulence effectors into host cells. The Ysc-Yop T3SS is tightly regulated by multiple contributing proteins that function at different levels. However, systematic transcriptional regulation analysis of Ysc-Yop T3SS is lacking and the detailed mechanism under this regulation process is still elusive. Aimed at systematically characterizing transcriptional regulations of all T3SS genes in Y. pseudotuberculosis, we amplified 97 non-coding fragments from the pYV plasmid and analyzed transcriptional responses of the T3SS genes under different growth conditions. Transcriptions of T3SS genes were induced at 37°C and genes encoding T3SS effectors were highly induced by further depletion of Ca2+. The temperature induced gene transcription process is mediated by modules encoded on the chromosome, while the Ca2+ depletion-induced process is controlled by the positive regulatory protein LcrF as well as the negative regulatory protein LcrQ. In this process, LcrQ shares the same targets with LcrF and the effect of LcrQ is dependent on the presence of LcrF. Furthermore, over-expression of LcrF showed the same phenotype as that of the lcrQ mutant strain and intracellular amount balance of LcrQ and LcrF is important in T3SS regulation. When the expression level of LcrF exceeds LcrQ, expression of the Ysc-Yop T3SS genes is activated and vice versa. Together, these data support a model in which LcrQ blocks the activation role of LcrF in regulating the transcription of T3SS genes in Yersinia.
Introduction
The genus Yersinia includes three human pathogenic bacterial species, Y. pestis, Y. enterocolitica and Y. pseudotuberculosis. These pathogens harbor a conserved virulence plasmid called pYV (or pCD) which encodes the Ysc-Yop type III secretion system (T3SS) [1]. When contacting with host cells, pathogenic Yersinia can efficiently inject effectors termed Yops (Yersinia outer proteins) into eukaryotic host cells with this secretion system. These translocated Yops enable bacteria to evade host immune responses by modulating host cell signaling pathways and facilitate the following infection [1]–[3].
The Ysc-Yop T3SS in Yersinia is both positively and negatively regulated and is induced by low Ca2+ conditions at 37°C in vitro or by direct contact with host cells in vivo [4], [5]. Several regulators are involved in the negative regulatory process. The LcrQ protein (or the YscM1 and YscM2 homologues in Y. enterocolitica) is an anti-activator which causes the feedback inhibition for Ysc-Yop T3SS gene expression. This protein is secreted outside of cells by the secretory machinery under low Ca2+ conditions at 37°C with the help from a chaperone protein named SycH [6]–[8]. This process relieves the repressive effect of LcrQ and results in the activation of the Ysc-Yop T3SS [7]. We have also recently characterized the role of an export apparatus component named YscV in LcrQ secretion and subsequent Ysc-Yops T3SS regulation [9]. However, the targets and roles of LcrQ during this regulatory process have not yet been fully illustrated. In addition to LcrQ, other negative regulatory elements, including SycN, YscB, YopN, TyeA and the lcrGVH-yopBD operon were also demonstrated to play important roles in regulating T3SS gene expression [10]–[14].
The AraC family activator LcrF (known as VirF in Y. enterocolitica) is the only protein from the pYV plasmid that positively regulates the transcription of T3SS genes [15]. LcrF directly binds to the promoter regions of several yop genes and also binds to RNA polymerase as supposed by the AraC family proteins [16]. With such interactions, LcrF enhances the binding of RNA polymerase to specific promoters to facilitate the transcription initiation. Transcription of a number of genes from the pYV plasmid, e.g., ylpA, yadA, yopE, yopH, virC and lcrGVHyopBD operons, are dependent on LcrF [17]–[20]. The direct bindings of LcrF to yopE, yopH, virC and lcrGVHyopBD have been confirmed and the ∼40 bp AT-rich region in each of these promoters is characterized to be recognized by LcrF [20].
The Ysc-Yop T3SS in Yersinia is regulated at both transcriptional and post-transcriptional levels. LcrF is a classical activator that controls T3SS gene expression at the transcriptional level [20]. YopD is another regulator which binds to the mRNA of several yop genes to accelerate mRNA degradation or inhibit the ribosome binding in gene translation [21], [22], which acts at the post-transcriptional level. The expression of lcrF has also been shown to be regulated at the post-transcriptional level. The mRNA structure around the ribosomal binding region of lcrF transcript is opened when the temperature is shifted from 26°C to 37°C [23], which would facilitate the gene translation. Several other regulators, such as the YopN-TyeA complex, also act at the post-translational level to inhibit T3SS secretion process [13], [24]. Although LcrQ negatively regulates the expression of T3SS genes, it lacks obvious DNA or RNA binding domains, and does not act by obstructing the secretion channel [25], thus the regulatory mechanism of LcrQ is not fully understood.
In this study, we systematically analyzed the responses of each gene on the pYV plasmid at the transcriptional level under different growth conditions in Y. pseudotuberculosis YPIII strain. By comparing the activities of each promoter, the transcriptional responses of the Ysc-Yop T3SS genes were characterized. The role of LcrQ in regulating the Ysc-Yop T3SS genes at the transcriptional level was subsequently analyzed, and its interaction with LcrF in this transcriptional regulatory process was further investigated and discussed.
Materials and Methods
Bacterial strains, plasmids, growth media and oligos
Bacterial strains and plasmids used in this study are summarized in Table 1 . Yersinia pseudotuberculosis YPIII strains were grown in YLB medium (1% tryptone, 0.5% yeast extract, 0.5% NaCl) and E. coli strains were grown in Luria-Bertani (LB) medium. When appropriate, antibiotics were used at the following concentrations: nalidixic acid (Nal) 15 μg ml−1, chloramphenicol 30 μg ml−1, ampicillin 100 μg ml−1, kanamycin 100 μg ml−1. All oligos used in this study are listed in Table S1.
Table 1. The strains and plasmids used in this work.
| Strain or plasmid Description* Source | |||
| Bacteria | |||
| Y. pesudotuberculosis | |||
| YPIII | Parental strain Y. pseudotuberculosis YPIII, Nalr | Lab collection | |
| ΔpYV | YPIII without virulence plasmid, Nalr | This study | |
| ΔlcrF | YPIII with the deletion of lcrF gene, Nalr | This study | |
| ΔlcrQ | YPIII with the deletion of lcrQ gene, Nalr | [9] | |
| ΔyscV 618–644 | YPIII with deletion of a 81 bp fragment encoding amino acid 618–644 in yscV gene, Nalr | [9] | |
| E. coli | |||
| DH5α | Used for Cloning construction | Lab collection | |
| BL21(DE3) | Used for protein expression | Novagen | |
| S17-1 (λ-pir) | λ-pir lysogen of S17-1, thi pro hsdR hsdM + recA RP4 2-Tc::Mu-Kan::Tn7 | Lab collection | |
| BTH101 | F−,cya-99, araD139, galE15, galK16, rpsL1+, hsdR2, mcrA1, mcrB1 | Lab collection | |
| Plasmid | |||
| pDM4 | Suicide vector, mobRK2, oriR6K, sacBR, Cmr | [27] | |
| pDM4-lcrFm | Suicide plasmid for ΔlcrF construction, Cmr | This study | |
| pOVR200 | Plasmid for over-expressing proteins, lac promoter, Ampr | This study | |
| pOVR-LcrF | pOVR200 carrying lcrF gene, Ampr | This study | |
| pOVR-LcrQ | pOVR200 carrying lcrQ gene, Ampr | This study | |
| pOVR-GST-LcrQ | pOVR200 carrying GST-lcrQ fusion, Ampr | This study | |
| pKT100 | Cloning vector, p15A replicon, Kanr | [26] | |
| pKT-LcrF | pKT100 carrying lcrF gene, Kanr | This study | |
| pKT-LcrQ | pKT100 carrying lcrQ gene, Kanr | This study | |
| pKT-GST-LcrQ | pKT100 carrying GST-lcrQ fusion, Kanr | This study | |
| pZT100 | Promoter cloning vector, transcriptional lacZ fusion, Kanr | [9] | |
| pKT25 | Two-hybrid vector, for T25 fragment fusion, Kanr | [30] | |
| pKT25-LcrF | pKT25 carrying lcrF gene, Kanr | This study | |
| pKT25-SycH | pKT25 carrying sycH gene, Kanr | This study | |
| pUT18 | Two-hybrid vector, T18 fragment fusion, Ampr | [30] | |
| pUT18-LcrF | pUT18 carrying lcrF gene, Ampr | This study | |
| pUT18-LcrQ | pUT18 carrying lcrQ gene, Ampr | This study | |
| pET28a | Protein expression vector, Kanr | Novagen | |
| pET28a-LcrF | pET28a carrying lcrF gene from YPIII, Kanr | This study | |
* Nalr, Cmr, Kanr and Ampr represent resistance to naladixic acid, chloramphenicol, kanamycin and ampicillin respectively.
Promoter cloning and β-galactosidase assay
The sequence of the pYV virulence plasmid in Y. pseudotuberculosis YPIII strain is not available. We collected and analyzed available sequences of the virulence plasmids of different Y. pseudotuberculosis strains from the NCBI database, e.g., pYV from IP 32953 (NC_006153.2) and IP 31758 (NC_009704.1), pYPTS01 from PB1+ strain (NC_010635.1). There are 99 annotated genes on the pYV plasmid from IP32953, 66 genes from IP31758, and 87 genes on pYPTS01 from PB1+ strain. To cover all possible genes, we chose the sequence of the pYV plasmid from IP32953 as the template to design primers (Table S1) for cloning of the non-coding fragments from the pYV plasmid in YPIII. All these amplified non-coding fragments were cloned into the promoter cloning vector pZT100 [9]. These clones were respectively transformed into YPIII or its derivative strains and the β-galactosidase assay was performed at least in triplicate as previously described [26] at 26°C, or 37°C or 37°C with Ca2+ depletion.
RNA isolation and real-time PCR
Total RNA of Yersinia strains cultured at 26°C, or 37°C or 37°C with Ca2+ depletion was extracted with TRIzol reagent (Invitrogen) according to the manufacturer’s instruction. After treatment with RNase-free DNase I (Promerga), cDNA was obtained by reverse transcription (Random 9 mers, TaKaRa) using 2 μg of each RNA sample. Real-time PCR was performed in reactions containing 1×iTaq™ Universal SYBR Green Supermix Reagent (Bio-Rad), cDNA templates and the gene specific primers using the CFX Connect Bio-Rad system. All these assays were tested in duplicates. The mRNA level in each sample was normalized to the level of 16S rRNA gene transcript and amplification specificity was assessed using melting curve analysis. The degrees of expression change were calculated using the 2−ΔΔCt method.
Gel-retardation assay
The gel-retardation was performed as described previously [26]. The LcrF protein was expressed in E. coli BL21(DE3) by pET28a plasmid carrying the lcrF gene from YPIII (named pET28a-LcrF), and purified by Ni-NTA resin. About 400 ng of each DNA fragment, which was amplified from the YPIII genomic DNA, was incubated with various amounts of purified LcrF protein in 20 μl of binding buffer (20 mM Tris-HCl pH 7.4, 4 mM MgCl2, 100 mM NaCl, 1 mM dithiothreitol, 10% glycerol, and 100 ng bovine serum albumin). A ∼200 bp fragment amplified from the non-coding region upstream of the pYV0053 gene was used as a negative control. Probe binding were performed at 37°C for 1 h, and were then loaded onto a 6.5% native polyacrylamide gel. Electrophoresis was performed in 0.5×TBE buffer on ice. The gel was stained with ethidium bromide for 20 min and scanned using a Syngene GeneGenius gel documentation system.
Mutant construction and complementation
For ΔlcrF construction, two DNA fragments flanking the lcrF gene were amplified by PCR from the genomic DNA of YPIII using two pairs of primers: named LcrF-in-upF/R, and LcrF-in-downF/R (Table S1). These two fragments were mixed and then used as the template DNA in a second round of PCR with primers LcrF-in-upF and LcrF-in-downR. The PCR product was digested with SalI and BglII and inserted into the suicide plasmid pDM4 [27]. This recombinant plasmid was transformed into E. coli S17-1 and transconjugation was performed as described [27] to obtain the ΔlcrF strain. To complement ΔlcrF, a plasmid named pOVR200 was constructed, which contains the pMB1 replicon, the bla gene and the lacI gene from pET21a (Novagen) and a lac promoter upstream of the multiple cloning site. The lcrF gene was PCR amplified from YPIII and was cloned into pOVR200 to obtain pOVR-LcrF, which was transformed into ΔlcrF. The pOVR200 plasmid was also transformed into ΔlcrF as a control.
Over-expression and co-expression of LcrF and LcrQ
The pOVR-LcrF plasmid obtained above was transformed into YPIII to over-express LcrF protein. To over-express LcrQ protein, lcrQ gene was amplified from YPIII and cloned into pOVR200 to obtain pOVR-LcrQ, which was transformed into YPIII. For co-expression of LcrF and LcrQ (or GST-LcrQ), the low copy plasmid named pKT100 [26], which contains the pOVR200 compatible P15A replicon and a promoter region of the chloramphenicol resistance gene, was used to express the target proteins together with the pOVR200 plasmid.
Yops analysis
Overnight cultures of Yersinia strains grown in Ca2+ rich medium (YLB with 2.5 mM Ca2+) were diluted (1:20) into fresh Ca2+-depleted medium (YLB containing 5 mM EGTA and 20 mM MgCl2) [28] and incubated at 26°C for 1 h, then shifted to 37°C to induce Yops expression and secretion. When required, isopropyl β-D-thiogalactoside (IPTG) at a final concentration of 1 mM was added into the cultures before temperature changes. After 4 h of incubation at 37°C, 20 ml of each culture was harvested and centrifuged. The wet weight of the bacterial cell pellet was measured and the supernatant was collected and filtered by 0.22 μm filter (Millipore). Proteins from the supernatant were precipitated overnight at 4°C with 10% trichloroacetic acid as described [9]. The proteins from both supernatants and pellets were dissolved in the SDS loading buffer according to the wet weight of the bacteria and resolved by SDS-PAGE. Western-blot was carried out to detect the proteins following SDS-PAGE. The horseradish peroxidase-labeled anti-rabbit antibodies and a chemiluminescence detection kit (Beyotime Institute of Biotechnology) were used to develop the reaction following the protocol provided by the manufacturer.
pYV plasmid curing
The pYV plasmid in YPIII was cured as described by others [29]. Briefly, YPIII strain was grown on YLB plate supplemented with 20 mM of sodium oxalate, 20 mM MgCl2, and 0.05% Congo Red Dye at 37°C. White colonies were picked up and cultured overnight in YLB medium and then spread on Congo Red plate. After overnight culture at 37°C, a single white colony (loss of the functional pYV plasmid results in white colonies) was chosen for PCR to further confirm that the pYV plasmid was cured using five pairs of primers, which paired to different regions on the pYV plasmid.
Bacterial two-hybrid assay
The adenylate cyclase based bacterial two-hybrid system [30] was applied in analyzing the interaction between LcrF and LcrQ proteins. Briefly, pKT25 plasmid carrying T25 fragment was used to clone lcrF and sycH genes to obtain T25-LcrF and T25-SycH. The pUT18 carrying T18 fragment was applied to clone lcrF and lcrQ genes to obtain T18-LcrF and T18-LcrQ. These two pairs of plasmids were co-transformed into E. coli BTH101 and were cultured in LB medium with 1 mM IPTG at 30°C for 2 days. The β-galactosidase activities were determined as described [26].
Statistical analysis
All data for the β-galactosidase activity assays were shown as mean ± standard deviation (SD) of the results of multiple independent experiments. Student’s T test was used to compare the data between two relevant groups.
Results
Transcriptional responses of T3SS genes under inducible conditions
To systematically characterize the transcriptional responses of Ysc-Yop T3SS genes in YPIII under T3SS inducible conditions, we compared the promoter activities of all genes from the pYV plasmid. Using primers designed from Y. pseudotuberculosis IP 32953 strain, we successfully amplified 96 non-coding regions from the pYV plasmid in YPIII. Only three regions upstream of pYV0042, pYV0043 and pYV0044 could not be obtained. We next sequenced the region from pYV0041 to pYV0045, a deletion at the C-terminal of the coding region of pYV0041 and a full deletion of the pYV0042 and pYV0043 genes were found in YPIII. Using this new sequence, we designed another pair of primers to amplify the non-coding region of pYV0044. In total, 97 non-coding fragments were amplified and cloned into a lacZ reporter vector to obtain non-coding fragment-lacZ fusions. The promoter activities of these non-coding fragments under T3SS non-inducible (26°C and 37°C) and inducible (37°C with Ca2+ depletion) conditions were subsequently analyzed. As shown in Fig. 1A , 68 out of the 97 non-coding fragments showed promoter activities compared with the pZT100 plasmid (202±12, p<0.01) at least under one condition. We also compared the promoter activities of these 68 non-coding fragments at 26°C and 37°C, which is one of the key factors that induces T3SS in vitro. As shown in Fig. 1B , 34 out of 68 promoters showed higher (>2 folds) activities at 37°C, suggesting some factors activate the expression of T3SS genes when bacterial cells were shifted from 26°C to 37°C.
Figure 1. Promoter activity analyses of genes on the pYV plasmid in YPIII.

(A) The promoter activities of 99 non-coding fragments at 26°C, 37°C or 37°C with Ca2+ depletion. Fragments with relative promoter activities higher than the pZT100 plasmid (202±12, p<0.01) at one of these conditions were marked with red dot. (B) & (C) Relative promoter activities of selected fragments in YPIII and ΔpYV strains at 26°C and 37°C (B) or at 37°C with Ca2+ depletion versus at 37°C (C).
To test if these factors are encoded by genes on the pYV plasmid, we next compared this activation in YPIII parent strain with the pYV plasmid cured strain (ΔpYV). Our results showed that induced folds at 37°C for all these promoters are similar between YPIII parent and ΔpYV strains ( Fig. 1B ), which suggests that the activation at 37°C is largely mediated by factors encoded on the chromosome. To fully characterize the transcriptional response during the T3SS inducing process, we further compared the promoter activities of these 68 fragments at 37°C and at 37°C with Ca2+ depletion. As shown in Fig. 1C , the activities of 9 promoters were induced more than 2-fold when Ca2+ was depleted. On the contrary to temperature changes, this activation is dependent on factors encoded on the pYV plasmid, since all these activations were strongly decreased in the ΔpYV strain ( Fig. 1C ). To further confirm the lacZ fusions results, we applied qRT-PCR to test the mRNA levels of lcrG and lcrQ genes in YPIII under different conditions. Being consistent with the β-galactosidase activity assay, the mRNA level of lcrG gene was highly induced at 37°C with Ca2+ depletion, and lcrQ gene is induced at 37°C (Fig. S1).
Comparing Fig. 1B and Fig. 1C , we conclude that most of the promoters activated between 37°C and the Ca2+ depleted conditions do not overlap, suggesting the Ysc-Yop T3SS activation process induced by 37°C and low Ca2+ is different. As summarized in Table S2, the genes induced at 37°C could be divided into four categories according to their functions: I) pYV plasmid replication; II) secretion machinery components; III) effector proteins; and IV) hypothetical proteins. On the contrary, the genes induced by Ca2+-depletion mainly encode effectors for Ysc-Yop T3SS (Table S3). Based on these analyses, we conclude that the Ysc-Yop T3SS inducing process first involves genes encoded from the YPIII chromosome, which activate the transcription of most of the secretion machinery components when the culture temperature is shifted from 26°C to 37°C, and full activation requires other key factors encoded on the pYV plasmid to maximally express the effector proteins.
Ca2+-depletion induced genes are regulated by LcrF
Since LcrF protein is the only characterized transcriptional regulator from the pYV plasmid, we next aimed to explore the role of LcrF in Ca2+-depletion induced gene expression. To systematically analyze the regulatory targets of LcrF, We compared the promoter activities from the pYV plasmid under T3SS inducible conditions in YPIII parent and the lcrF mutant (ΔlcrF) strains. Interestingly, activities of those nine Ca2+-depletion induced promoters, including promoters upstream of sycO (pYV0002), yadA (pYV0013), yopE (pYV0025), yopK (pYV0040), lcrG (pYV0058), yopN (pYV0065), yscN (pYV0067) and yopH (pYV0094), were all dramatically decreased in ΔlcrF ( Fig 2A ), which suggests all these genes are possible targets of LcrF. To further confirm these promoters are indeed regulated by LcrF, we also compared the promoter activities in the ΔlcrF complementary strain. As shown in Fig. 2B , expression of LcrF in ΔlcrF successfully restored the activities of these promoters.
Figure 2. Role of LcrF in controlling the transcription of T3SS genes.

(A) Relative promoter activities of genes in lcrF mutant (ΔlcrF) compared with YPIII parent strain under T3SS inducible conditions. Promoters with decreased activities were marked with red dot. (B) & (C) Effect of LcrF over-expression (pOVR-LcrF) on the promoter activities of its targets in ΔlcrF strain at 37°C with or without Ca2+. ** p<0.01.
Transcriptional regulation of LcrF to T3SS is not influenced by Ca2+
Since transcription of all the targets of LcrF is induced by Ca2+-depletion, we next compared the regulatory effects of LcrF under Ca2+ rich and depleted conditions. As shown in Fig. 2C , over-expression of LcrF in ΔlcrF induced the expression of Yops genes in the presence of Ca2+, which suggests the regulatory activity of LcrF is not influenced by Ca2+. To confirm this, we next tested the regulatory role of LcrF to its targets in the ΔpYV strain. Our results showed that expression of LcrF also increased the activities of these promoters both in Ca2+ rich and depleted conditions (Fig. S2). These data are inconsistent with the results from Y. enterocolitica in which the role of LcrF is not affected by Ca2+ [31]. However, our data demonstrated that transcription of all the promoters targeted by LcrF are strongly induced by depletion of Ca2+ at 37°C, which suggests other co-factor(s) may sense the change of Ca2+ concentration and coordinate with LcrF in regulating the Ysc-Yop T3SS.
Role of LcrQ in the regulatory process of LcrF
The LcrQ protein globally regulates the Ysc-Yop T3SS in respond to Ca2+ concentration changes, but the mechanism under this regulatory role has not been illustrated. To test whether LcrQ is involved in controlling the regulatory effect of LcrF, we next screened the targets of LcrQ by comparing the promoter activities in YPIII parent and LcrQ over-expressed strains using our promoter library. Interestingly, all and only the targets of LcrF showed decreased promoter activities in the LcrQ over-expressed strain compared with those in YPIII parent strain ( Fig. 3A ), which suggests LcrQ shares the same targets with LcrF in regulating the Ysc-Yop T3SS.
Figure 3. Role of LcrQ in regulating the transcription of T3SS genes.

(A) Comparison of relative promoter activities of T3SS genes under T3SS inducible conditions in LcrQ over-expressed strain with YPIII carrying the pOVR200 plasmid. Promoters repressed by LcrQ were marked with red dot. (B) Secretion (Supernatant) and expression (Pellet) of T3SS effectors in LcrQ over-expressed and ΔlcrF strains under T3SS inducible conditions. RpoA was used as a loading control. (C) Secretion and expression of T3SS effectors in ΔlcrQ and LcrF over-expressed strains at 37°C with or without Ca2+. (D) & (E) Effects of LcrQ over-expression on the promoter activities of three LcrQ targets (yopE, lcrG and yopH) in YPIII ΔpYV and ΔlcrF strains.
To further analyze the relationship between LcrF and LcrQ in regulating the Ysc-Yop T3SS, we next compared the phenotypes of ΔlcrF with LcrQ over-expressed strain and LcrF over-expressed strain with ΔlcrQ. The expression of Yops in the LcrQ over-expressed strain was the same as that in ΔlcrF, which were both decreased under T3SS inducible conditions ( Fig. 3B , lanes 2&4), and Yops secretion was also decreased as confirmed by Western blot analysis ( Fig. 3B ). Over-expression of LcrF and deletion of the lcrQ gene both activated Yops expression and secretion under T3SS inducible conditions ( Fig. 3C ). Moreover, deletion of the lcrQ gene successfully secreted LcrV and YopD proteins in the presence of Ca2+ at 37°C as reported by others [32], and over-expression of LcrF also promote the secretion of these two proteins ( Fig. 3C ). Together, these data suggest LcrQ and LcrF are closely related during the regulatory process.
We next compared the regulatory effects of LcrQ to its targets in YPIII parent and ΔpYV strains. As shown in Fig. 3D , promoter activities of the LcrQ targets were at basal levels in ΔpYV strain and over-expression of LcrQ could not repress these promoters in ΔpYV strain; however, these promoter activities were obviously inhibited by over-expressing LcrQ in YPIII parent strain (Fig. S3), suggesting that LcrQ represses the promoter activities activated by pYV-encoded protein. To test whether this factor is LcrF, we tested the repressive effect of LcrQ in ΔlcrF strain and found no repressive role to these promoters ( Fig. 3E ). It is worthy to note that the activities of these promoters in LcrQ over-expressed YPIII strain were at similar levels as that in ΔlcrF strain, indicating that LcrQ only represses the LcrF-activated but not the basal level promoter activities. Together, these data suggest that the repressive effect of LcrQ requires LcrF.
Intracellular LcrQ inhibits the regulatory role of LcrF
Judging from the data we have obtained above, we hypothesized that intracellular LcrQ blocks the regulatory role of LcrF. If so, the expression levels of these two proteins are important for the regulatory process. To confirm this hypothesis, we over-expressed LcrF using the pOVR200 plasmid in the YPIII strain carrying pKT-LcrQ, in which LcrQ protein was also over-expressed by the pKT100 plasmid. As shown in Fig. 4A , over-expression of LcrF released the negative regulatory effect of LcrQ in this strain (compared lane 4 with lane 3). In agreement with this observation, the amount of intracellular LcrQ was obviously decreased in LcrF over-expressed strain ( Fig. 4A ). Therefore, these results can be explained by intracellular amount balance of LcrF and LcrQ proteins. When the amount of LcrF exceeds LcrQ, LcrF dominates the regulatory effect. To prove our hypothesis, we next exchanged the plasmid to over-express these two proteins. pOVR200, which contains a strong promoter, was used to over-express LcrQ and pKT100 was used to express LcrF. In this case, the amount of intracellular LcrQ protein was slightly increased ( Fig. 4A , compared lane 7 with lane 4), which in turn greatly inhibited the positive effect of LcrF ( Fig. 4A , compared lane 7 with lane 5). All together, these results suggest that the intracellular amount of LcrQ is important for blocking the regulatory role of LcrF.
Figure 4. Effects of co-expression of LcrF and LcrQ on T3SS.

(A) Secretion and expression of Yops and LcrQ in strains with overexpression of LcrF or LcrQ or co-expression of these two proteins. (B) Effects of co-expression of GST-fused LcrQ (G-Q) with LcrF on Yops and LcrQ expression and secretion. (C) Influences of over-expressing LcrF in an yscV mutant (ΔyscV 618–644) on expression of Yops. (D) Promoter activities of LcrF targets in ΔyscV 618–644 strains carrying pOVR-LcrF or pOVR200.
To further confirm our hypothesis, we used the pOVR200 plasmid to express GST-LcrQ (which is known to abolish transport of LcrQ [6], [33]) in YPIII strain carrying the pKT-LcrF plasmid. As anticipated, GST-LcrQ is not secreted ( Fig. 4B ). And over-expression of LcrF by the pKT100 plasmid in this background did not show obvious effect on the expression or secretion of Yops ( Fig. 4B , compared lane 7 with lane 5). In our previous work, we showed that mutation of the yscV gene (ΔyscV 618–644), which encodes a component of secretion machinery, accumulates the intracellular LcrQ protein and therefore represses the expression of Yops [9]. We next over-expressed LcrF protein in this background and found that both Yops expression and the promoter activities of its targets were all activated comparing with ΔyscV 618–644 carrying the control plasmid pOVR200 ( Fig. 4C&D ). Taken together, our data suggest that the amount of intracellular LcrQ and LcrF is vital for the regulation of Ysc-Yop T3SS. We hypothesize that intracellular LcrQ abolishes the role of LcrF at 37°C under Ca2+ rich conditions, and this inhibition is released when LcrQ is secreted outside of cells under Ca2+ depleted conditions.
LcrQ does not directly interact with LcrF
The regulatory mechanism for the LcrQ protein has not been fully understood. No DNA binding domain can be found on this protein [25], suggesting LcrQ does not directly interact with its target DNA. To explore how LcrQ functions in the regulatory process of LcrF, we applied the Bacterial Adenylate Cyclase Two-Hybrid (BACTH) system to detect possible interaction between these two proteins. As shown in Fig. 5 , LcrF can form dimer as reported for the AraC family proteins [16], and LcrQ can interact with its chaperone protein SycH [6], [7], which suggest the expression of these two proteins in the BACTH system is stable. However, we could not detect direct interaction between LcrF and LcrQ ( Fig. 5 ), suggesting that other factor(s) is also required for LcrQ to interact with LcrF to block its regulatory role.
Figure 5. Interaction between LcrQ and LcrF by a bacterial two-hybrid assay.
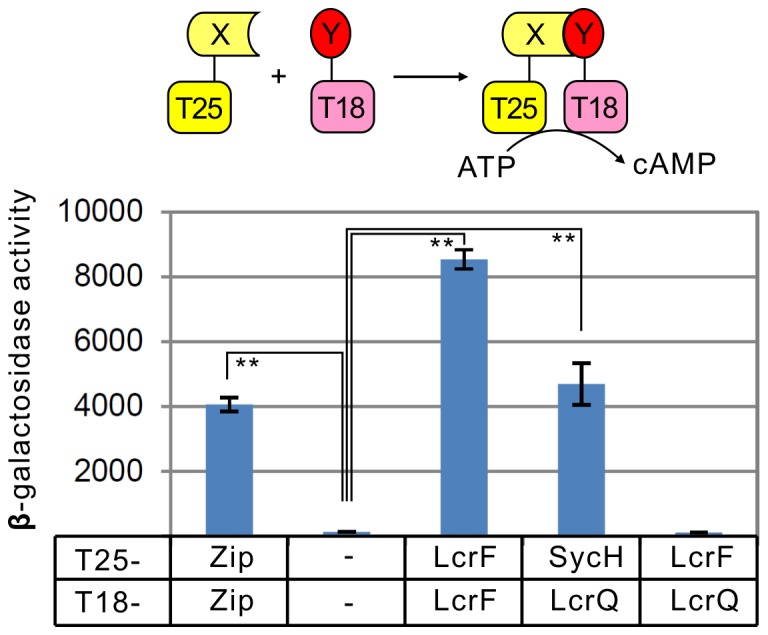
Model for bacterial adenylate cyclase two-hybrid assay is shown in the upper panel. Interaction between LcrQ and LcrF as detected by bacterial two-hybrid system is shown in the lower panel. ** p<0.01.
Discussion
The Ysc-Yop T3SS is a strictly regulated system and several modules have been demonstrated to play important roles in this regulatory process [34]. However, this regulation at the transcriptional level has not been systematically analyzed. In this study, we have cloned 97 non-coding fragments from the pYV plasmid into the lacZ fusion plasmid and compared the promoter activities of all these fragments under T3SS inducible and non-inducible conditions. The Ysc-Yop T3SS is induced by low Ca2+ signal at 37°C in vitro [4]. In agreement with this phenotype, transcription of genes on the pYV plasmid is also regulated by low Ca2+ signal at 37°C. Based on the promoter activity analyses, we proposed a model for transcriptional regulation of Ysc-Yop T3SS in Yersinia. As shown in Fig. 6 , factors encoded on the chromosome first sense the temperature changes. At 37°C, these factors will activate the transcription of most genes on the pYV plasmid, including the secretion machinery components and two regulators LcrF and LcrQ. Under this condition, LcrQ blocks the role of LcrF. Therefore, the expression of T3SS genes could not be massively activated by LcrF at 37°C when LcrQ and LcrF are both intracellularly located. However, the low Ca2+ signal will trigger the secretion of LcrQ, which subsequently release the repressive effect of LcrQ, the positive regulatory role of LcrF will thus be exerted.
Figure 6. Proposed model for the transcriptional responses of the T3SS genes in Yersinia.

LcrF and LcrQ are two characterized regulators for Ysc-Yop T3SS in Yersinia. LcrF is a classical transcriptional regulator that can directly binds to several promoters on the pYV plasmid [20]. In this study, we systematically characterized the targets of LcrF and identified 9 targets in YPIII. Being consistent with other reports in Y. enterocolitica [20], yopE, yopH, yopQ/yopK, yadA, lcrGVHyopBD operon and virC are all activated by LcrF. VirF is not essential for transcription of both virA and virB operons in Y. enterocolitica [15], but these two operons were both regulated by LcrF in Y. pseudotuberculosis. Binding of LcrF to these two promoter fragments were confirmed by in vitro gel-retardation assay (Fig. S4), which suggests these two operons are also the direct targets of LcrF. The regulation of LcrF to its targets is not affected by Ca2+, but the transcription of all these genes in YPIII are induced by low Ca2+ signal. These data suggest that some other factors are also involved in activating the role of LcrF under low Ca2+ conditions. The role of LcrQ in T3SS regulation has not been documented. LcrQ has been proposed to form a complex with YopD-LcrH proteins [35]. A recent study has shown that YscM1 (a homologue of LcrQ in Y. enterocolitica) binds to ribosomes together with YopD-LcrH and subsequently inhibits translation of the yop genes [36]. Our current study suggests another mechanism for the regulatory role of LcrQ to block the role of LcrF. This blocking process is not in a direct way since we did not observe any direct interaction between these two proteins by a customized bacterial two-hybrid assay, which is in agreement with a previous study showing that LcrQ does not act as an anti-LcrF protein [25]. We suppose that other uncharacterized proteins may act as a linker between these two proteins in this process ( Fig. 6 ). Functional homologues of LcrF and LcrQ are found in other pathogens. For example, the T3SS in Pseudomonas aeruginosa is triggered by low Ca2+ at 37°C and is also globally regulated by two proteins with opposing effects: the AraC family positive regulator ExsA and the secreted negative regulator ExsE [37]. Similar to LcrQ and LcrF, ExsE blocks the role of ExsA by indirect interactions with two intermediate proteins ExsC and ExsD [37]. Further studies are needed to characterize functional homologues of ExsC and ExsD in Yersinia and find the binding proteins of LcrF and LcrQ, which will provide information to detail the mechanism of transcriptional regulation to the Ysc-Yop T3SS.
In addition to regulators encoded by the pYV plasmid, several chromosome encoded proteins are also involved in the regulation of the Ysc-Yop T3SS. For example, YmoA, a chromosome-encoded histone-like protein, directly binds to the promoter region of the lcrF gene to inhibit its transcription. The ATP-dependent ClpXP and Lon proteases degrade YmoA when the environment temperature achieved at 37°C, which in turn releases the transcription of lcrF gene and subsequently activates the T3SS in Yersinia [38]. And the LysR family regulatory protein YtxR competes with LcrF in binding with the promoter regions of yopE and yopH genes in regulating T3SS [39], [40]. All these factors regulate the Ysc-Yop T3SS indirectly through LcrF protein. In our study, we found that transcription of most of the pYV genes are activated by the chromosome-encoded factors at 37°C and these pYV-encoded genes are not all belongs to the targets of LcrF. These data suggest that chromosome-encoded factor is not one of the above proteins. Previous studies have detected that supercoiling of the pYV plasmid changes at 30°C and 37°C and supposed that this DNA topology change may be the temperature-sensing mechanism for T3SS gene expression [41], [42]. Whether the uncharacterized factors are proteins participated in DNA architecture changes need further studies.
As stated in the introduction, the Ysc-Yop T3SS is regulated at both transcriptional and post-transcriptional levels. In this study, our proposed model is only for the transcriptional regulation of the Ysc-Yop T3SS, which could not explain all the phenotypes we have observed in our study. For example, as suggested in our model, deletion of the lcrQ gene would facilitate the role of LcrF to activate the Yops at 37°C in the presence of Ca2+, as demonstrated by the fact that blockage to the role of LcrF by LcrQ is diminished in the ΔlcrQ strain. However, as shown in Fig. 3C , the secretion pattern of the ΔlcrQ strain under Ca2+ rich conditions is different from the normal secretion pattern, which suggests some other factors may also participate in controlling the secretion pathway of Ysc-Yop T3SS. In addition, the promoter activities of yopE and yopH remains high in ΔlcrF background ( Fig. 3E ), but Western blot results showed the expression of these proteins are undetectable in this strain ( Fig. 3B ), which indicate the regulation also occurs at the post-transcriptional level. Moreover, we found that the thermal induction of yopEp-lacZ is not dependent on the pYV plasmid, which is different from the results using a yopE-lacZ fusion (the yopE covers a promoter and a ∼700 bp coding region in this fusion) [19]. These suggest some pYV-encoded elements may inhibit the expression of yopE at the post-transcriptional level, e.g., YopD protein [21]. The expression of LcrQ protein was increased in LcrF over-expressed strain ( Fig. 4 & Fig. S5) but its promoter activity was not activated by LcrF protein. Transcription is only the first step for gene expression. Regulation at other levels should be considered together to fully clarify the complicated regulatory process of Ysc-Yop T3SS.
Among the 97 cloned non-coding fragments, 68 have promoter activities (summarized in Fig. S6), and several of these non-coding fragments located inside an operon, e.g., the non-coding fragment upstream of yopD gene (pYV0054), which is located in the lcrGVH-yopBD operon, were detected with promoter activity, suggesting the transcriptional regulation of pYV-encoded T3SS is complicated. On the other hand, we did not observe promoter activities of some genes located as the first gene in an operon. This may due to the fact that some fragments we chose do not cover the full promoter regions, since open reading frame annotation is only based on bioinformatics analysis from the NCBI, which may introduce mis-annotation. Nevertheless, the promoter bank we have constructed in this study will facilitate further transcriptional regulation analysis in Yersinia and results obtained in this study would provide information for further transcriptional regulation analyses related to the Ysc-Yop T3SS.
Supporting Information
The mRNA levels of lcrG and lcrQ genes in YPIII at 26°C, 37°C or 37°C with Ca2+ depletion.
(TIF)
Regulatory role of LcrF to its targets in the ΔpYV strain in the absence (A) or presence (B) of Ca2+ at 37°C.
(TIF)
Repressive effect of LcrQ over-expression on promoter activities of yopE p, lcrG p and yopH p.
(TIF)
The binding of LcrF protein to lcrG p (A), pYV0053p (B), yopN p (C) and sycN p (D). LcrF protein was added at final concentrations of 0, 0.025, 0.05, 0.1, 0.25, 0.5 and 1 μM respectively.
(TIF)
Relative level of LcrQ protein in LcrF over-expressed strains (quantified from Figure 4A ).
(TIF)
Location of promoters on the pYV plasmid. The non-coding fragments with promoter activities detected in this study were labeled with black “P”, and those located upstream of the first gene in an operon but showed no promoter activity in our test were labelled with grey “P”.
(TIF)
Primers used in this study.
(XLS)
Genes induced at 37°C on the pYV plasmid.
(DOC)
Genes induced by Ca2+ depletion at 37°C on the pYV plasmid.
(DOC)
Funding Statement
Funded by the National Science Foundation of China (#31170133). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Cornelis GR, Boland A, Boyd AP, Geuijen C, Iriarte M, et al. (1998) The virulence plasmid of Yersinia, an antihost genome. Microbiol Mol Biol Rev 62: 1315–1352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Viboud GI, Bliska JB (2005) Yersinia outer proteins: role in modulation of host cell signaling responses and pathogenesis. Annu Rev Microbiol 59: 69–89. [DOI] [PubMed] [Google Scholar]
- 4. Yother J, Chamness TW, Goguen JD (1986) Temperature-controlled plasmid regulon associated with low calcium response in Yersinia pestis . J Bacteriol 165: 443–447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Juris SJ, Shao F, Dixon JE (2002) Yersinia effectors target mammalian signalling pathways. Cell Microbiol 4: 201–211. [DOI] [PubMed] [Google Scholar]
- 6. Cambronne ED, Cheng LW, Schneewind O (2000) LcrQ/YscM1, regulators of the Yersinia yop virulon, are injected into host cells by a chaperone-dependent mechanism. Mol Microbiol 37: 263–273. [DOI] [PubMed] [Google Scholar]
- 7. Wulff-Strobel CR, Williams AW, Straley SC (2002) LcrQ and SycH function together at the Ysc type III secretion system in Yersinia pestis to impose a hierarchy of secretion. Mol Microbiol 43: 411–423. [DOI] [PubMed] [Google Scholar]
- 8. Cambronne ED, Sorg JA, Schneewind O (2004) Binding of SycH chaperone to YscM1 and YscM2 activates effector yop expression in Yersinia enterocolitica . J Bacteriol 186: 829–841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Li Y, Li L, Huang L, Francis MS, Hu Y, et al. (2014) Yersinia Ysc-Yop type III secretion feedback inhibition is relieved through YscV-dependent recognition and secretion of LcrQ. Mol Microbiol 91: 494–507. [DOI] [PubMed] [Google Scholar]
- 10. Nilles ML, Williams AW, Skrzypek E, Straley SC (1997) Yersinia pestis LcrV forms a stable complex with LcrG and may have a secretion-related regulatory role in the low-Ca2+ response. J Bacteriol 179: 1307–1316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Cheng LW, Schneewind O (2000) Yersinia enterocolitica TyeA, an intracellular regulator of the type III machinery, is required for specific targeting of YopE, YopH, YopM, and YopN into the cytosol of eukaryotic cells. J Bacteriol 182: 3183–3190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Day JB, Ferracci F, Plano GV (2003) Translocation of YopE and YopN into eukaryotic cells by Yersinia pestis yopN, tyeA, sycN, yscB and lcrG deletion mutants measured using a phosphorylatable peptide tag and phosphospecific antibodies. Mol Microbiol 47: 807–823. [DOI] [PubMed] [Google Scholar]
- 13. Joseph SS, Plano GV (2007) Identification of TyeA residues required to interact with YopN and to regulate Yop secretion. Adv Exp Med Biol 603: 235–245. [DOI] [PubMed] [Google Scholar]
- 14. Costa TR, Edqvist PJ, Broms JE, Ahlund MK, Forsberg A, et al. (2010) YopD self-assembly and binding to LcrV facilitate type III secretion activity by Yersinia pseudotuberculosis . J Biol Chem 285: 25269–25284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Lambert de Rouvroit C, Sluiters C, Cornelis GR (1992) Role of the transcriptional activator, VirF, and temperature in the expression of the pYV plasmid genes of Yersinia enterocolitica . Mol Microbiol 6: 395–409. [PubMed] [Google Scholar]
- 16. Schleif R (2010) AraC protein, regulation of the l-arabinose operon in Escherichia coli, and the light switch mechanism of AraC action. FEMS Microbiol Rev 34: 779–796. [DOI] [PubMed] [Google Scholar]
- 17. Skurnik M, Toivanen P (1992) LcrF is the temperature-regulated activator of the yadA gene of Yersinia enterocolitica and Yersinia pseudotuberculosis . J Bacteriol 174: 2047–2051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. China B, Michiels T, Cornelis GR (1990) The pYV plasmid of Yersinia encodes a lipoprotein, YlpA, related to TraT. Mol Microbiol 4: 1585–1593. [DOI] [PubMed] [Google Scholar]
- 19. Hoe NP, Minion FC, Goguen JD (1992) Temperature sensing in Yersinia pestis: regulation of yopE transcription by LcrF. J Bacteriol 174: 4275–4286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Wattiau P, Cornelis GR (1994) Identification of DNA sequences recognized by VirF, the transcriptional activator of the Yersinia yop regulon. J Bacteriol 176: 3878–3884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Chen Y, Anderson DM (2011) Expression hierarchy in the Yersinia type III secretion system established through YopD recognition of RNA. Mol Microbiol 80: 966–980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Schiano CA, Lathem WW (2012) Post-transcriptional regulation of gene expression in Yersinia species. Front Cell Infect Microbiol 2: 129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Bohme K, Steinmann R, Kortmann J, Seekircher S, Heroven AK, et al. (2012) Concerted actions of a thermo-labile regulator and a unique intergenic RNA thermosensor control Yersinia virulence. PLoS Pathog 8: e1002518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Amer AA, Costa TR, Farag SI, Avican U, Forsberg A, et al. (2013) Genetically engineered frameshifted YopN-TyeA chimeras influence type III secretion system function in Yersinia pseudotuberculosis . PLoS One 8: e77767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Stainier I, Iriarte M, Cornelis GR (1997) YscM1 and YscM2, two Yersinia enterocolitica proteins causing downregulation of yop transcription. Mol Microbiol 26: 833–843. [DOI] [PubMed] [Google Scholar]
- 26. Hu Y, Lu P, Wang Y, Ding L, Atkinson S, et al. (2009) OmpR positively regulates urease expression to enhance acid survival of Yersinia pseudotuberculosis . Microbiology 155: 2522–2531. [DOI] [PubMed] [Google Scholar]
- 27. OToole R, Milton DL, WolfWatz H (1996) Chemotactic motility is required for invasion of the host by the fish pathogen Vibrio anguillarum . Mol Microbiol 19: 625–637. [DOI] [PubMed] [Google Scholar]
- 28. Straley SC, Bowmer WS (1986) Virulence Genes Regulated at the Transcriptional Level by Ca2+ in Yersinia pestis Include Structural Genes for Outer Membrane Proteins. Infect Immun 51: 445–454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Balada-Llasat JM, Mecsas J (2006) Yersinia has a tropism for B and T cell zones of lymph nodes that is independent of the type III secretion system. PLoS Pathog 2: e86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Karimova G, Pidoux J, Ullmann A, Ladant D (1998) A bacterial two-hybrid system based on a reconstituted signal transduction pathway. Proc Natl Acad Sci U S A 95: 5752–5756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Cornelis G, Sluiters C, de Rouvroit CL, Michiels T (1989) Homology between virF, the transcriptional activator of the Yersinia virulence regulon, and AraC, the Escherichia coli arabinose operon regulator. J Bacteriol 171: 254–262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Rimpilainen M, Forsberg A, Wolf-Watz H (1992) A novel protein, LcrQ, involved in the low-calcium response of Yersinia pseudotuberculosis shows extensive homology to YopH. J Bacteriol 174: 3355–3363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Lee VT, Anderson DM, Schneewind O (1998) Targeting of Yersinia Yop proteins into the cytosol of HeLa cells: one-step translocation of YopE across bacterial and eukaryotic membranes is dependent on SycE chaperone. Mol Microbiol 28: 593–601. [DOI] [PubMed] [Google Scholar]
- 34. Dewoody RS, Merritt PM, Marketon MM (2013) Regulation of the Yersinia type III secretion system: traffic control. Front Cell Infect Microbiol 3: 4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Cambronne ED, Schneewind O (2002) Yersinia enterocolitica type III secretion: yscM1 and yscM2 regulate yop gene expression by a posttranscriptional mechanism that targets the 5' untranslated region of yop mRNA. J Bacteriol 184: 5880–5893. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Kopaskie KS, Ligtenberg KG, Schneewind O (2013) Translational regulation of Yersinia enterocolitica mRNA encoding a type III secretion substrate. J Biol Chem 288: 35478–35488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Urbanowski ML, Lykken GL, Yahr TL (2005) A secreted regulatory protein couples transcription to the secretory activity of the Pseudomonas aeruginosa type III secretion system. Proc Natl Acad Sci U S A 102: 9930–9935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Jackson MW, Silva-Herzog E, Plano GV (2004) The ATP-dependent ClpXP and Lon proteases regulate expression of the Yersinia pestis type III secretion system via regulated proteolysis of YmoA, a small histone-like protein. Mol Microbiol 54: 1364–1378. [DOI] [PubMed] [Google Scholar]
- 39. Axler-Diperte GL, Miller VL, Darwin AJ (2006) YtxR, a conserved LysR-like regulator that induces expression of genes encoding a putative ADP-ribosyltransferase toxin homologue in Yersinia enterocolitica . J Bacteriol 188: 8033–8043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Axler-DiPerte GL, Hinchliffe SJ, Wren BW, Darwin AJ (2009) YtxR acts as an overriding transcriptional off switch for the Yersinia enterocolitica Ysc-Yop type 3 secretion system. J Bacteriol 191: 514–524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Rohde JR, Fox JM, Minnich SA (1994) Thermoregulation in Yersinia enterocolitica is coincident with changes in DNA supercoiling. Mol Microbiol 12: 187–199. [DOI] [PubMed] [Google Scholar]
- 42. Rohde JR, Luan XS, Rohde H, Fox JM, Minnich SA (1999) The Yersinia enterocolitica pYV virulence plasmid contains multiple intrinsic DNA bends which melt at 37 degrees C. . J Bacteriol 181: 4198–4204. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
The mRNA levels of lcrG and lcrQ genes in YPIII at 26°C, 37°C or 37°C with Ca2+ depletion.
(TIF)
Regulatory role of LcrF to its targets in the ΔpYV strain in the absence (A) or presence (B) of Ca2+ at 37°C.
(TIF)
Repressive effect of LcrQ over-expression on promoter activities of yopE p, lcrG p and yopH p.
(TIF)
The binding of LcrF protein to lcrG p (A), pYV0053p (B), yopN p (C) and sycN p (D). LcrF protein was added at final concentrations of 0, 0.025, 0.05, 0.1, 0.25, 0.5 and 1 μM respectively.
(TIF)
Relative level of LcrQ protein in LcrF over-expressed strains (quantified from Figure 4A ).
(TIF)
Location of promoters on the pYV plasmid. The non-coding fragments with promoter activities detected in this study were labeled with black “P”, and those located upstream of the first gene in an operon but showed no promoter activity in our test were labelled with grey “P”.
(TIF)
Primers used in this study.
(XLS)
Genes induced at 37°C on the pYV plasmid.
(DOC)
Genes induced by Ca2+ depletion at 37°C on the pYV plasmid.
(DOC)