

Abstract
In Burkholderia cenocepacia H111, the large surface protein BapA plays a crucial role in the formation of highly structured communities, known as biofilms. We have recently demonstrated that quorum sensing (QS) is necessary for the maximal expression of bapA. In this study we identify BapR, a protein from the IclR family of transcriptional regulators that, in conjunction with QS, controls biofilm formation by affecting the expression of bapA. We present evidence that, in addition to bapA, BapR influences the expression of extracellular proteases, swimming motility and has a profound impact in the incidence of persister cells, making this regulator an interesting target for persister cells and biofilm eradication.
Introduction
Burkholderia cenocepacia is a Gram-negative opportunistic pathogen that belongs to the Burkholderia cepacia complex (Bcc), a group that currently comprise 17 bacterial species [1]. Bcc strains show a remarkable ability to thrive in different niches that range from environmental to human clinical settings [2]. Despite having a high potential in biotechnological applications, their use has been restricted due to the emergence of Bcc strains as human opportunistic pathogens, particularly in patients affected by cystic fibrosis [3]–[5].
As part of the mechanisms controlling gene expression, B. cenocepacia utilizes QS, an ubiquitous mechanism among Gram-negative bacteria that relies in the synthesis, diffusion, detection and response to self-generated signals [6]. B. cenocepacia H111 has two QS systems, one based on N-acyl-homoserine lactone (AHL) and a second based on cis-2-dodecenoic acid (BDSF) [7]–[9]. These two systems regulate a specific and an overlapping set of genes [9], modulating the expression of phenotypes as diverse as protease production, swarming motility, pathogenicity and the formation of biofilms. Recently, the role of QS-regulated factors that had an impact on biofilm development was studied in B. cenocepacia H111 [7]. Among a set of 48 genes identified as downregulated in a cepR deficient strain, the lectin cluster BclACB and particularly the large surface protein BapA showed a significant contribution to the development of the biofilm [7]. We sought to extend these findings by looking for additional regulatory elements that could participate in the control of the biofilm phenotype. Here, we identify BapR, a transcriptional regulator of the IclR family that is able to modulate the expression of bapA and thus control biofilm formation. We show that BapR, in conjunction with the AHL-BDSF QS systems, is necessary for maximal expression of bapA and for maximal biofilm formation. Additionally, we provide evidence that BapR plays a role in the expression of other phenotypes like motility, protease production and also in the maintenance of a persister cell subpopulation of B. cenocepacia H111.
Results and Discussion
A mutation in CCE51534 results in lowered PbapA-lacZ expression
We have recently established that cell-to-cell communication in Burkholderia cenocepacia H111, mediated by AHLs and by BDSF, controls the expression of a specific and an overlapping set of genes [7], [9]. One of the genes identified in these studies as controlled by both systems was bapA, which codes for a large surface protein of crucial importance in biofilm formation [7]. The expression of bapA was shown to be diminished in both cepI (AHL biosynthesis) or rpfFBc (BDSF biosynthesis) mutant backgrounds, and was only restored to wild-type levels when the media was supplemented with both AHLs and BDSF in a double cepI rpfFBc mutant background. These results highlighted that expression of bapA requires multiple signals, suggesting a complex regulatory network. In order to extend these findings, we aimed to further characterize the regulation of bapA, looking for additional regulatory elements that may be part of the transcriptional arsenal driving expression of this gene. Here, we performed transposon mutagenesis in a B. cenocepacia cepI rpfFBc double mutant background and screened for diminished activity of PbapA-lacZ fusion in the presence of BDSF (see methods). From about 86000 clones, 19 clones representing 13 different loci had lowered or no PbapA-lacZ activity (Table S1, Figure S1). Among the mutants analyzed, we found 3 transposon insertions in the gene CCE51534. This gene codes for a protein of the IclR family of transcriptional regulators that we re-named to bapR (bapA regulator, see below). Members of this family of proteins can act as transcriptional activators or repressors and may bind cofactor molecules in order to act as DNA-binding regulators [10]. The translated nucleotide sequence of bapR was used to search against the genomic database of current members of the Bcc. We found that BapR is ubiquitously distributed within the Bcc, showing the highest identity with the homolog present in the ET12 lineage strain J2315 (Table 1). Interestingly, in this strain bapR contains a mutation at position 139 (G to T) that results in an early termination codon, suggesting that BapR is not functional in this strain.
Table 1. BapR identity within members of the Bcc.
| Bcc. strain | protein ID | % identitya |
| Burkholderia cenocepacia J2315 | n.d.b | 99 |
| Burkholderia cenocepacia BC7 | U1ZF62 | 99 |
| Burkholderia cenocepacia PC184 | A2VRI9 | 98 |
| Burkholderia cenocepacia MC0-3 | ACA90348.1 | 98 |
| Burkholderia cenocepacia HI2424 | ABK07951.1 | 97 |
| Burkholderia cenocepacia AU 1054 | ABF75630.1 | 97 |
| Burkholderia cenocepacia KC-01 | V4ZZ24 | 98 |
| Burkholderia dolosa AUO158 | A2WBB1 | 96 |
| Burkholderia multivorans ATCC 17616 | BAG43081.1 | 94 |
| Burkholderia multivorans ATCC 17616 | ABX15788.1 | 94 |
| Burkholderia multivorans ATCC BAA-247 | J5BML4 | 95 |
| Burkholderia multivorans CGD2M | B9CGL9 | 95 |
| Burkholderia multivorans CGD2 | B9BXD9 | 95 |
| Burkholderia multivorans CF2 | J5BGZ7 | 95 |
| Burkholderia ambifaria MEX-5 | B1TDI7 | 99 |
| Burkholderia ambifaria IOP40-10 | B1FMA5 | 95 |
| Burkholderia ambifaria MC40-6 | ACB63572.1 | 94 |
| Burkholderia ambifaria AMMD | ABI86640.1 | 93 |
| Burkholderia vietnamiensis G4 | ABO54125.1 | 93 |
| Burkholderia vietnamiensis AU4i | U2HJ07 | 97 |
Identity to full-length predicted protein. The predicted BapR protein sequence from B. cenocepacia H111 was used to search against the Burkholderia Genome Database (www.burkholderia.com) with the TBLASTN algorithm [20].
n.d., not determined. In B. cenocepacia J2315 there is an ochre, nonsense mutation after codon 46.
bapR expression is not auto-regulated
We engineered a bapR mutant strain and compared its growth to that of the wild-type (WT) strain. As depicted in Figure 1A, no significant differences in growth were detected between these two strains. It has been reported that members of the IclR-family of transcriptional regulators may control their own expression [10] and for this reason we measured the expression of bapR using a promoter fusion with lacZ as reporter gene in various genetic backgrounds including the bapR mutant background. As depicted in Figure 1B, the activity of the bapR promoter fusion was comparable in both the WT and the bapR mutant background, suggesting that bapR is not auto-regulated. Moreover, the activity of the bapR promoter was not significantly altered in the QS-deficient mutants cepIR or rpfFBc (Figure 1B), suggesting that the expression of this gene is not regulated by QS.
Figure 1. Expression of bapR is not auto-regulated.
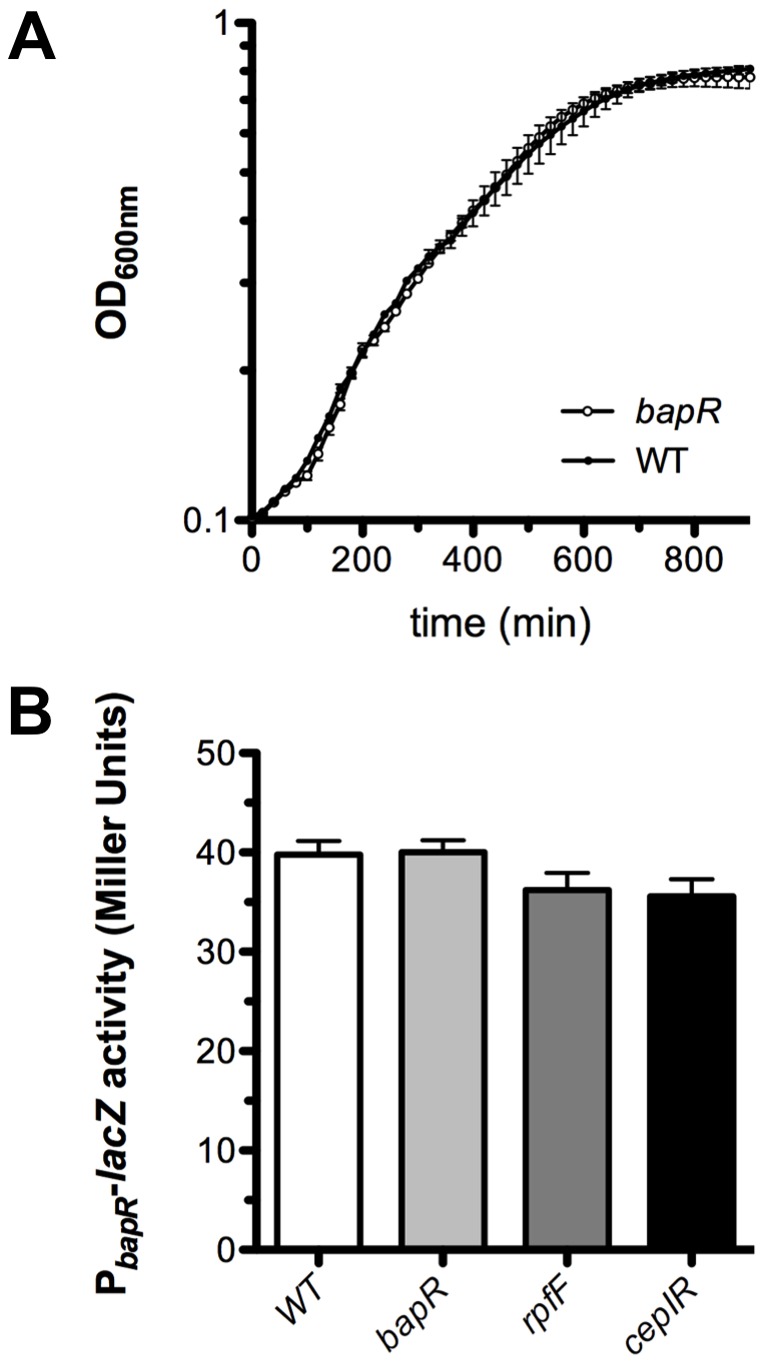
A, growth curves of the bapR mutant and its parent, WT strain. The strains were inoculated on microtiter plates in LB at 37°C and the OD600nm was monitored every 20 min. No significant differences in growth were detected between the strains (Bars, SEM; n = 3). B, the promoter region of bapR was cloned in front of a promoter-less lacZ reporter gene. The activity of the promoter fusion was measured in different genetic backgrounds, as depicted, using samples obtained from late exponential cultures. No significant difference was detected between the samples (ANOVA, p>0.05). Bars, SEM; n = 3.
BapR influences the expression of bapA
In order to study and characterize the function of BapR we measured the activity of a PbapA-lacZ promoter fusion in a bapR mutant background. In agreement with the results obtained from the transposon library screening, the activity of the promoter fusion PbapA-lacZ was diminished in the bapR mutant background (Figure 2A). We next tested biofilm formation and found that the bapR mutant produced less biofilms compared to the WT (Figure 2B). Based on these results we hypothesized that BapR is necessary for expression of bapA. To confirm this hypothesis, we created conditional mutants of bapR and bapA in which the expression of these genes were induced only upon addition of rhamnose to the media (see methods). We first tested biofilm formation in presence or absence of rhamnose. As depicted in Figure 3, biofilm formation was recovered after expressing bapR by the addition of rhamnose. Importantly, no biofilm formation was observed when bapR expression was induced in a bapA mutant background. We next tested whether the biofilm phenotype persisted when expressing bapA in a bapR mutant background. In this case, we found that biofilm formation was near WT levels, suggesting that the regulatory action of BapR over bapA can be uncoupled and that expression of bapA was necessary and sufficient for the formation of biofilms under the conditions tested.
Figure 2. A bapR mutation affects bapA expression and biofilm formation.
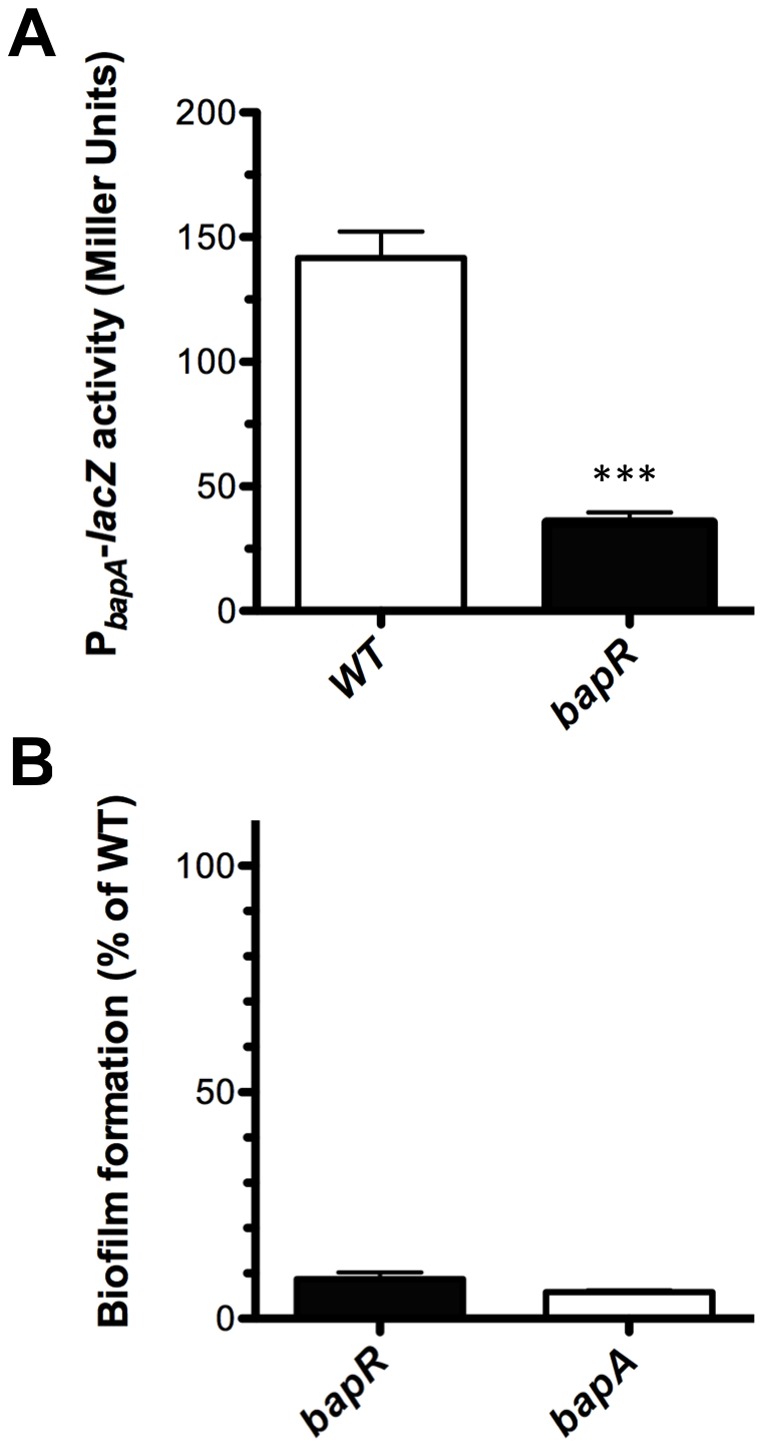
A, the activity of a PbapA-lacZ promoter fusion was measured in both the WT and the bapR genetic backgrounds from samples taken from late exponential cultures. B, biofilm formation in 96-well plates was measured after 20 h, staining the biomass adhered to the wells with crystal violet. Error bars, SEM. Asterisks denote significant differences in bapA expression between WT and the bapR mutant strain (t-test, *** p<0.0001, n≥3).
Figure 3. BapR controls biofilm formation via BapA.

The defect in biofilm formation of a bapR mutant was reverted by expressing bapR from a rhamnose-inducible promoter. No biofilm was detected when bapR expression was induced in a bapA genetic background. Expression of bapA is necessary and sufficient to restore the biofilm defect of a bapR mutant. White bars are biofilms grown in the absence of rhamnose. Black bars are biofilms grown in the presence of 0.2% rhamnose. Error bars, SEM. Asterisks denote significant differences in biofilm formation when rhamnose was used to induce the expression of bapR or bapA (2-way ANOVA, *** p<0.001, n≥3).
Maximal expression of bapA and biofilm formation requires BapR and QS
In a previous study we determined that maximal expression of bapA (measured as PbapA-lacZ activity) was detected only when both AHL and BDSF were present in the medium [9]. We explored whether the regulatory action of BapR over bapA required the presence of QS. To do this, we expressed bapR from the rhamnose-inducible promoter in a cepI rpfFBc double mutant background harbouring a PbapA-lacZ promoter fusion. As depicted in Figure 4A, maximum activity of the reporter (close to WT levels) was only obtained when bapR was induced and both AHL and BDSF were present in the media. We next tested biofilm formation in these conditions. In agreement with the pattern of expression of bapA, maximal biofilm formation was obtained only after induction of bapR and addition of C8-AHL and BDSF to the media (Figure 4B).
Figure 4. BapR and QS are required for maximal expression of bapA and for maximal biofilm formation.
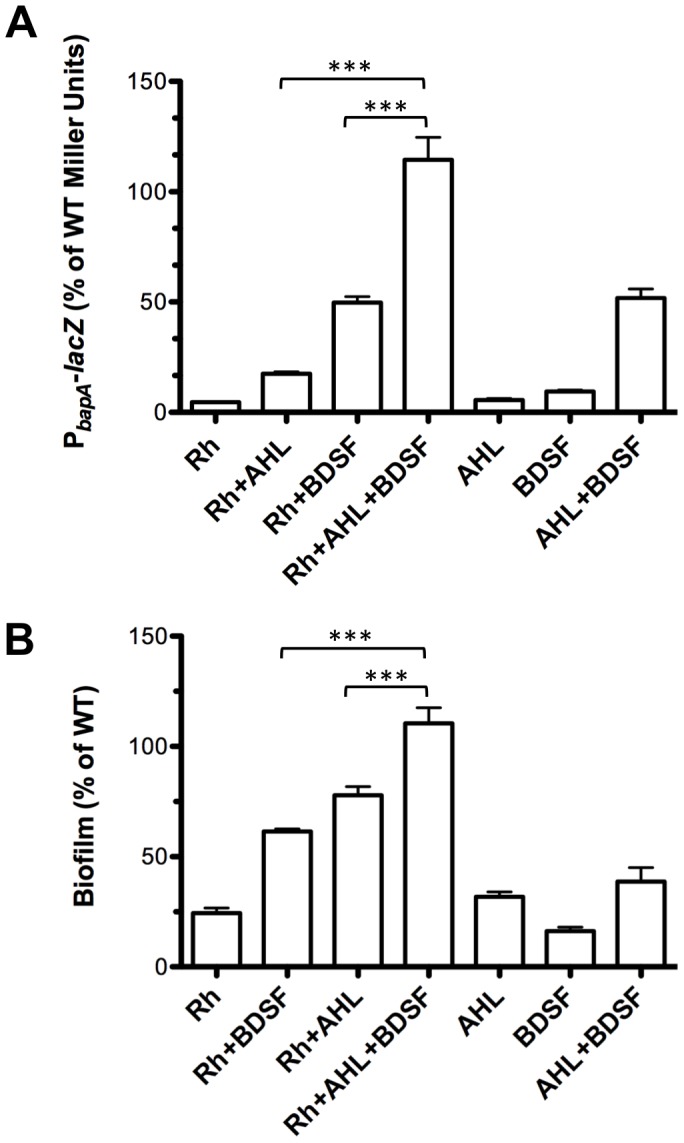
A, a mutant cepI rpfFBc, Prha-bapR and harboring a PbapA-lacZ promoter fusion was grown in the presence of rhamnose, C8-AHL, BDSF or the combination of these molecules. Expression of bapA was determined measuring the activity of the reporter gene. B, biofilm formation was quantified in a mutant cepI rpfFBc, Prha-bapR after the addition of rhamnose, C8-AHL, BDSF or the combination of these molecules. Error bars, SEM. Asterisks denote significant differences in bapA expression or in biofilm formation between the samples after supplementing with rhamnose, C8-AHL, BDSF or the combination of these molecules (1-way ANOVA, *** p<0.0001, n≥3).
BapR affects motility, extracellular protease activity and persisters abundance
Throughout this study we have provided evidence showing that BapR is an important regulatory element controlling biofilm formation. Since B. cenocepacia is an important opportunistic human pathogen, we were interested in exploring the possibility that BapR could also be influencing other aspects of the physiology of this bacterium. For this reason, we performed a transcriptome analysis using RNA-Seq (see Supplementary Methods and Figure S2) which allowed us to compare the global expression of a bapR mutant to its respective WT parent strain. The results of this analysis further confirmed that bapA, together with all the members of the operon (CCE53117-120) were under the control of BapR (Table S2). Among the top-regulated genes, we found 253 genes with altered expression (116 down-regulated and 119 up-regulated, p<0.1, see Table S2). These genes were classified into different categories and the analysis of these categories suggest that BapR is mainly involved in the control of functions related to amino-acid transport and metabolism (19 genes), cell motility (28 genes), cell wall/membrane/envelope biogenesis (14 genes) and other, non-classified functions (96 genes). Genes showing a diminished expression in a bapR mutant background included the bap operon (CCE53117-120), the bclACB lectins (CCE46720-22), the extracellular protease zmpB (CCE52940) and the nematocidal protein aidA (CCE52108), which have all been previously described as QS-regulated (Table S2). To further validate the results obtained from this global analysis we chose two conditions and tested for changes in their respective phenotypes. Since ZmpB was found to be regulated by BapR (fold change in expression of -4 between bapR mutant and WT, see Table S2) we measured the extracellular protease activity of the bapR mutant strain and found that there is in fact a significant decrease (Figure 5A). We also detected that the expression of at least 28 genes involved in cell motility was up-regulated in the bapR mutant background. In agreement with these results, the bapR mutant displayed a significant increase in motility when tested in a swimming assay (Figure 5B). Interestingly, the expression of a gene coding for an isocitrate lyase (aceA, CCE52795, BCAL2118) was down-regulated in the bapR mutant background. We validated this result by quantitative PCR, and we could measure a fold-change of −3.1±0.3 in the bapR mutant background compared to WT (not shown). A recent study suggested a link between aceA and the maintenance of the persister subpopulation of cells in Bcc strains [11]. In the clinical setting, chronic, biofilm-related infections are typically refractory to the treatment with antibiotics and it is believed that persister cells are responsible for this recalcitrance [12]. In this context, AceA would be of importance for persister cell maintenance since it helps cells to reduce the production of reactive oxygen species (ROS) by utilizing the glyoxylate shunt instead of the TCA cycle [11]. Motivated by this finding, we next tested whether BapR influenced the abundance of persisters cells in cultures of B. cenocepacia H111. As aminoglycoside antibiotics are implicated in the formation of ROS in the cell, we investigated the effect of aminoglycoside antibiotics (gentamicin and amikacin) on the abundance of persister cells in a bapR mutant compared to its parent strain. As shown in Figure 6, a mutation in bapR has in fact a strong impact on the abundance of persisters upon treatment with aminoglycoside antibiotics, showing a dose-dependent effect that caused a reduction of the persister subpopulation of over 10-fold. These results strongly suggest that in B. cenocepacia H111, aceA is at least partially responsible for the maintenance of a persister cell sub-population. While we cannot exclude that other genes, in addition to aceA, may contribute to the abundance of persister cells, these findings suggest that BapR may represent an interesting drug target for biofilm and persister eradication in the clinic.
Figure 5. BapR acts positively on extracellular protease activity and negatively on swimming motility.
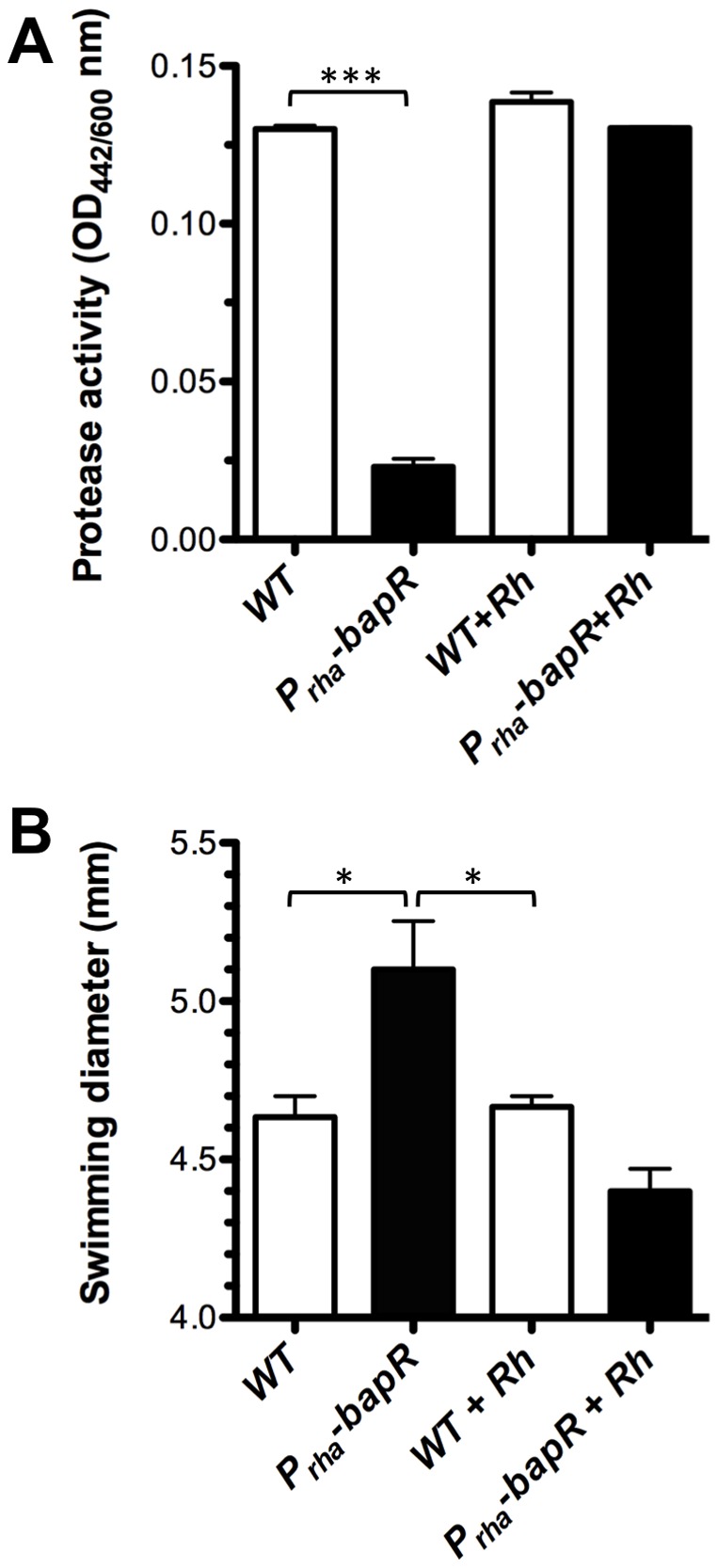
A, extracellular protease was measured in the supernatant of WT or bapR mutant strains grown in NYG. B, swimming motility was measured after incubation of WT or bapR mutant strains in LB media fortified with 0.3% agar. Error bars, SEM. Asterisks denote significant differences in protease production or swimming motility between WT and Prha-bapR mutant strain (1-way ANOVA, * p<0.01, *** p<0.0001, n≥3).
Figure 6. A mutation in bapR decreases the number of persister cells.

A, Percentage of surviving cells after treatment with increasing concentration of gentamicin after 20 h. B, Percentage of surviving cells after treatment with increasing concentration of amikacin after 20 h. White bars, WT; black bars, bapR mutant. Error bars, SEM. Asterisk denote significant differences in persister cell abundance between WT and bapR mutant after treatment with aminoglycoside antibiotics (2-way ANOVA, ** p<0.005, n = 3).
In summary, we present evidence that BapR, a novel IclR-family regulator, controls the expression of bapA and as a consequence biofilm formation. By means of RNA-seq analysis we show that, in addition to bapA, BapR controls a set of approximately 253 genes, including those necessary for swimming, protease production and aceA, which plays a role in the maintenance of a persister-cell subpopulation. At the moment it is not known whether BapR controls the expression of target genes directly or via downstream regulators. Some of the BapR-controlled phenotypes could be explained by the down-regulation of the QS systems by BapR, like.g. in the case of the BclACB lectins or BapA. However, based on the RNA-seq data, we did not obtained evidence for such a control. We therefore favor the idea that BapR and the QS systems synergistically control biofilm formation and bapA expression. Since BapA is a remarkably large protein (over 2700 amino-acid residues), the regulation of bapA expression is probably under tight control in the cell. For example, the bapA promoter has a long non-translated region of approximately 600 bp which might contain regulatory elements that contribute to bapA expression in addition to BapR (Figure S3). The precise regulatory mechanism of BapR and its synergy with the QS system is the subject of ongoing research.
Materials and Methods
Bacterial strains, plasmids and growth conditions
Strains and plasmids used in this study are listed in Table S3. Bacterial strains were grown aerobically at 37°C in LB broth, Lennox (BD, Sparks, MD, USA) or AB minimal medium [13] supplemented with 10 mM sodium citrate. Antibiotics were added as required at final concentrations of 100 μg/ml ampicillin, 50 μg/ml kanamycin, 10 μg/ml gentamicin, 100 μg/ml trimethoprim, 80 μg/ml chloramphenicol. Growth was spectrophotometrically monitored by measurement of optical density (OD) at 600 nm.
DNA manipulations, conjugative plasmid transfer and nucleotide sequencing
Routine DNA molecular techniques were performed using standard methods [14]. Plasmid DNA was isolated with a miniprep kit (Qiagen, Hilden, Germany), chromosomal DNA of B. cenocepacia strains was isolated by the sarkosyl-pronase method [15]. Triparental matings from E. coli to B. cenocepacia were performed with helper strains E. coli (pRK600) or E.coli (pRK2013) as previously described [16]. Sequencing reactions were performed with the ABI 3730 DNA analyzer using the ABI BigDye Terminator Cycle Sequencing kit (Applied Biosystems, Foster City, USA).
Transposon mutagenesis and screening
A transposon mutagenesis was performed as previously described [17], using a cepI rpfFBc double mutant of B. cenocepacia as genetic background and the transposon delivery vector pUT/mini-Tn5 Km [18]. Approximately 40000 independent transposon insertion mutants were obtained. Aliquots of the library were saved and stored at -80C. To perform the screening, the vector pPbapA-lacZ [7] was introduced into the transposon library by conjugation using E.coli S17-1 (pPbapA-lacZ) as parent strain, selecting for trimethoprim resistant colonies in plates supplemented with X-gal (100 μg/ml) in the presence of BSDF. We supplemented only with BDSF in order to reduce complexity of the system and also since we have determined that addition of this molecule is sufficient to recover approximately 50% of the expression of the promoter fusion in double cepI rpfFBc background [9]. Approximately 86000 clones were screened for diminished expression of the PbapA-lacZ promoter fusion. A total of 19 insertions in 13 different loci were found and selected for further analysis. To identify the loci interrupted by the transposon, arbitrary PCR was performed as previously described [17].
Construction of an insertional mutant in bapR
To generate an insertional mutant in bapR, a 288 bp internal fragment of CCE51534 was amplified by PCR using oligonucleotides bapR_F and bapR_R (Table S4) and cloned in pGEMT-easy (Promega). The fragment was then sub-cloned into the suicide vector pEX18Gm as BamHI/HindIII fragment, generating pNS-bapR. The plasmid was transferred to B. cenocepacia by triparental mating as described, selecting for gentamicin-resistant colonies. The integrity of the insertion was verified by PCR using oligonucleotides bapR-check and pEXcheck_F (Table S4).
β-galactosidase activity determination
β-galactosidase activity obtained by the product of the lacZ reporter gene was quantified as described elsewhere [7]. The oligonucleotides used to create the different promoter fusions are listed in Table S4. Miller Units were obtained with the following formula, which includes normalization by cell growth: MU = (1000*OD420/OD540)/(time[min]*V[ml]*OD600).
Construction of a rhamnose-inducible bapR strain
Using the vector pSC200 [19], bapR was engineered for induction of expression upon addition of rhamnose to the media. The vector pSC200 was first digested with NdeI (New England Biolabs) and then blunt-ended with Klenow enzyme (Promega). The first 507 bp of bapR were amplified by PCR using oligonucleotides CA202 and CA203 with Pfu polymerase (Promega) and then cloned into the blunt-ended vector pSC200. The resulting plasmid, in which a rhamnose-inducible promoter controlled the expression of bapR, was transferred to the B. cenocepacia H111 by triparental mating [17] and the exconjugants were selected on PIA plates supplemented with trimethoprim. To induce the expression of the promoter, the media was supplemented with 0.2% rhamnose.
Biofilm quantification
Biofilm formation by B. cenocepacia H111 was quantified in a microtiter dish assay as described by Huber et al. 2001 with some modifications. Briefly, overnight cultures were normalized to an OD600 = 0.05 in AB media supplemented with 10 mM citrate and then used to inoculate a 96-well microtiter dish, incubating statically at 30°C for 20 h. Biofilms were stained by adding 100 μl of a 1% crystal violet solution and incubating for 30 min at room temperature. After the incubation period, the microtiter dishes were inverted to remove the contents of the wells and then they were washed gently and thoroughly using distilled water. The plates were allowed to dry at room temperature. The crystal violet adhered to the wells was resuspended in 120 μl of DMSO and the color was quantified at 550 nm in a Synergy HT microplate reader (Biotek, Luzern, Switzerland).
Extracellular proteolytic activity determination
Proteolytic activity was quantified as described previously [9]. Briefly, bacteria were grown at 37°C with vigorous shaking to late exponential growth phase in NYG medium (0.5% peptone, 0.3% yeast extract, 2% glycerol) and the OD600 was recorded. To start the reaction, 100 μl of a solution of azocasein (5 mg/ml, in 50 mM Tris-Cl, pH 8) was incubated with 100 μl of cell-free supernatant for 60 min at 37°C. After this, 400 μl of 10% TCA were added, followed by centrifugation. The supernatant was mixed with 750 μl of 525 mM NaOH and the absorbance at 442 nm was recorded. Protease activity was expressed as the ratio OD442/OD600.
Quantification of persister cells
To quantify the persister cell sub-population, overnight cultures were normalized to an OD600 of 0.05 and grown in LB broth with constant shaking until late stationary phase. Planktonic cells were harvested, washed twice with saline solution (0.9% NaCl) and normalized to an OD600 of 1 in saline solution. Cells were incubated without shaking at 37°C in the presence or in the absence of antibiotics for 20 h. After the antibiotic treatment, cells were washed twice with saline solution, serial diluted and seeded on LB plates fortified with 1.5% agar for quantification.
Statistical analysis of data
Histograms, curves and statistical analyses were performed with Prism, V.5a (www.graphpad.com). ANOVA analyses were conducted using Bonferroni post-test with 95% confidence intervals.
RNA-seq analysis
A full description of the RNA-seq, sequencing data analysis and quantitative PCR methodologies used in this study is presented in Supplementary methods. The RNA-Seq raw data files are accessible through the GEO Series accession number GSE52769.
Supporting Information
Identification of a mini-Tn5 transposon insertion in the IclR-type regulator bapR. A, from the genetic screening, three clones showed a diminished expression of the reporter PbapA-lacZ at exponential, early and late stationary phases of growth. B, Using arbitrary PCR, all three mini-Tn5 transposon insertion were mapped to a genetic locus coding for an IclR-type transcriptional regulator that we re-named bapR.
(TIF)
Differential gene expression in the bapR mutant compared to WT. MA plot showing the log2 fold change in expression observed in a bapR versus B. cenocepacia H111. The top regulated genes are shown in color: genes with increased expression in the bapR mutant are indicated in red, whereas genes whose expression was down-regulated are shown in green. Highlighted are bapA, the type 1 secretion genes necessary for BapA export (BCAM2142-40) and the protease zmpB.
(TIF)
Analysis of the bapA promoter region. A, the region upstream of bapA was systematically analyzed for promoter activity using the lacZ reporter gene. Black arrows represent the location and names of the primers used to generate the different promoter fusions, drawn to scale. B, β-galactosidase activity of each of the promoter fusions generated using the fragments depicted in A, named after the pair of oligonucleotides used in each case. White bars show the activity of the fusion in the WT background. Black bars show the activity of the fusion in the bapR mutant background. Error bars, SEM, n = 3.
(TIF)
(DOCX)
Genes identified by transposon mutagenesis in B. cenocepacia H111 displaying a diminished activity of the PbapA-lacZ reporter.
(DOCX)
Classification of 235 B. cenocepacia H111 genes that showed differential expression in a bapR mutant strain compared to the wild-type.
(DOCX)
Bacterial strains and plasmids used in this study.
(DOCX)
Oligonucleotides used in this study.
(DOCX)
Acknowledgments
We thank Alexander Grunau for excellent technical assistance. We are grateful to Ulrich Omasits and Christian Ahrens for advice on DE-Seq analysis.
Funding Statement
This work was supported by the Swiss National Science Foundation (Project 31003A_143773). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Vanlaere E, Lipuma JJ, Baldwin A, Henry D, De Brandt E, et al. (2008) Burkholderia latens sp. nov., Burkholderia diffusa sp. nov., Burkholderia arboris sp. nov., Burkholderia seminalis sp. nov. and Burkholderia metallica sp. nov., novel species within the Burkholderia cepacia comple. Int J Syst Evol Microbiol 58: 1580–1590. [DOI] [PubMed] [Google Scholar]
- 2. Coenye T, Vandamme P (2003) Diversity and significance of Burkholderia species occupying diverse ecological niches. Env Microbiol 5: 719–729. [DOI] [PubMed] [Google Scholar]
- 3. Parke JL, Gurian-Sherman D (2001) Diversity of the Burkholderia cepacia complex and implications for risk assessment of biological control strains. Annu Rev Phytopathol 39: 225–258. [DOI] [PubMed] [Google Scholar]
- 4. Vandamme P, Opelt K, Knochel N, Berg C, Schonmann S, et al. (2007) Burkholderia bryophila sp. nov. and Burkholderia megapolitana sp. nov., moss-associated species with antifungal and plant-growth-promoting properties. Int J Syst Evol Microbiol 57: 2228–2235. [DOI] [PubMed] [Google Scholar]
- 5. Mahenthiralingam E, Urban TA, Goldberg JB (2005) The multifarious, multireplicon Burkholderia cepacia complex. Nat Rev Microbiol 3: 144–156. [DOI] [PubMed] [Google Scholar]
- 6. Lewenza S, Conway B, Greenberg EP, Sokol PA (1999) Quorum sensing in Burkholderia cepacia: identification of the LuxRI homologs CepRI. J Bacteriol 181: 748–756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Inhülsen S, Aguilar C, Schmid N, Suppiger A, Riedel K, et al. (2012) Identification of functions linking quorum sensing with biofilm formation in Burkholderia cenocepacia H111. Microbiologyopen 1: 225–242 10.1002/mbo3.24 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Suppiger A, Schmid N, Aguilar C, Pessi G, Eberl L (2013) Two quorum sensing systems control biofilm formation and virulence in members of the Burkholderia cepacia complex. Virulence 4: 400–409 10.4161/viru.25338 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Schmid N, Deng Y, Pessi G, Aguilar C, Carlier AL, et al. (2012) The AHL- and BDSF-dependent Quorum Sensing Systems control specific and overlapping sets of genes in Burkholderia cenocepacia H111. PLoS One 7: e49966. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Molina-Henares A, Krell T, Eugenia Guazzaroni M, Segura A, Ramos J (2006) Members of the IclR family of bacterial transcriptional regulators function as activators and/or repressors. FEMS Microbiol Rev 30: 157–186 10.1111/j.1574-6976.2005.00008.x [DOI] [PubMed] [Google Scholar]
- 11. Van Acker H, Sass A, Bazzini S, De Roy K, Udine C, et al. (2013) Biofilm-grown Burkholderia cepacia complex cells survive antibiotic treatment by avoiding production of reactive oxygen species. PLoS One 8: e58943 10.1371/journal.pone.0058943 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Lewis K (2010) Persister cells. Annu Rev Microbiol 64: 357–372 10.1146/annurev.micro.112408.134306 [DOI] [PubMed] [Google Scholar]
- 13. Clark DJ, Maaløe O (1967) DNA replication and the division cycle in Escherichia coli . J Mol Biol 23: 99–112. [Google Scholar]
- 14.Sambrook J, Russel D (2001) Molecular Cloning: a Laboratory Manual, third edition. Cold Spring Harbor Laboratory.
- 15. Better M, Lewis B, Corbin D, Ditta G, Helinski DR (1983) Structural relationships among Rhizobium meliloti symbiotic promoters. Cell 35: 479–485. [DOI] [PubMed] [Google Scholar]
- 16. Huber B, Riedel K, Hentzer M, Heydorn A, Gotschlich A, et al. (2001) The cep quorum-sensing system of Burkholderia cepacia H111 controls biofilm formation and swarming motility. Microbiology 147: 2517–2528. [DOI] [PubMed] [Google Scholar]
- 17. Huber B, Riedel K, Köthe M, Givskov M, Molin S, et al. (2002) Genetic analysis of functions involved in the late stages of biofilm development in Burkholderia cepacia H111. Mol Microbiol 46: 411–426. [DOI] [PubMed] [Google Scholar]
- 18. De Lorenzo V, Timmis KN (1994) Analysis and construction of stable phenotypes in gram-negative bacteria with Tn5- and Tn10-derived minitransposons. Methods Enzym 235: 386–405. [DOI] [PubMed] [Google Scholar]
- 19. Ortega XP, Cardona ST, Brown AR, Loutet SA, Flannagan RS, et al. (2007) A putative gene cluster for aminoarabinose biosynthesis is essential for Burkholderia cenocepacia viability. J Bacteriol 189: 3639–3644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Winsor GL, Khaira B, Van Rossum T, Lo R, Whiteside MD, et al. (2008) The Burkholderia Genome Database: facilitating flexible queries and comparative analyses. Bioinformatics 24: 2803–2804. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Identification of a mini-Tn5 transposon insertion in the IclR-type regulator bapR. A, from the genetic screening, three clones showed a diminished expression of the reporter PbapA-lacZ at exponential, early and late stationary phases of growth. B, Using arbitrary PCR, all three mini-Tn5 transposon insertion were mapped to a genetic locus coding for an IclR-type transcriptional regulator that we re-named bapR.
(TIF)
Differential gene expression in the bapR mutant compared to WT. MA plot showing the log2 fold change in expression observed in a bapR versus B. cenocepacia H111. The top regulated genes are shown in color: genes with increased expression in the bapR mutant are indicated in red, whereas genes whose expression was down-regulated are shown in green. Highlighted are bapA, the type 1 secretion genes necessary for BapA export (BCAM2142-40) and the protease zmpB.
(TIF)
Analysis of the bapA promoter region. A, the region upstream of bapA was systematically analyzed for promoter activity using the lacZ reporter gene. Black arrows represent the location and names of the primers used to generate the different promoter fusions, drawn to scale. B, β-galactosidase activity of each of the promoter fusions generated using the fragments depicted in A, named after the pair of oligonucleotides used in each case. White bars show the activity of the fusion in the WT background. Black bars show the activity of the fusion in the bapR mutant background. Error bars, SEM, n = 3.
(TIF)
(DOCX)
Genes identified by transposon mutagenesis in B. cenocepacia H111 displaying a diminished activity of the PbapA-lacZ reporter.
(DOCX)
Classification of 235 B. cenocepacia H111 genes that showed differential expression in a bapR mutant strain compared to the wild-type.
(DOCX)
Bacterial strains and plasmids used in this study.
(DOCX)
Oligonucleotides used in this study.
(DOCX)