

Abstract
The neural cell adhesion molecule peptide mimetic fibroblast growth loop (FGL) proved to exert neuroprotective, neurotrophic, and anti-inflammatory effects in different in vitro and in vivo experiments. Based on this beneficial efficacy profile, it is currently in clinical development for neurodegenerative diseases and brain insults. Here, we addressed the hypothesis that the peptide might affect development of seizures in a kindling paradigm, as well as associated behavioral and cellular alterations. Both doses tested, 2 and 10 mg/kg FGL, significantly reduced the number of stimulations necessary to induce a generalized seizure. FGL did not exert relevant effects on the behavioral patterns of kindled animals. As expected, kindling increased the hippocampal cell proliferation rate. Whereas the low dose of FGL did not affect this kindling-associated alteration, 10 mg/kg FGL proved to attenuate the expansion of the doublecortin-positive cell population. These data suggest that FGL administration might have an impact on disease-associated alterations in the hippocampal neuronal progenitor cell population. In conclusion, the effects of the peptide mimetic FGL in the kindling model do not confirm a disease-modifying effect with a beneficial impact on the development or course of epilepsy. The results obtained with FGL rather raise some concern regarding a putative effect, which might promote the formation of a hyperexcitable network. Future studies are required to further assess the risks in models with development of spontaneous seizures.
Keywords: Amygdala kindling, FGL, mimetic peptide, NCAM, neurogenesis, epileptogenesis
Disease-modifying and antiepileptogenic strategies are considered a major goal of drug development in the field of epileptology.1,2 Multiple molecular and cellular alterations are evident during epileptogenesis following an initial brain insult. Among these alterations, neuronal plasticity and inflammatory signaling are discussed as key pathophysiological mechanisms contributing to epilepsy development and hyperexcitability in the chronic epileptic brain.2−4 Evidence exists that neuromodulatory and anti-inflammatory approaches can exert beneficial disease-modifying or preventive effects in chronic rodent models such as the kindling model or post-status epilepticus models.3
The neural cell adhesion molecule (NCAM)-derived peptide mimetic FGL (fibroblast growth loop) seems to constitute a good candidate compound to achieve neuromodulatory and anti-inflammatory effects and to target the development of a hyperexcitable network during epileptogenesis. This hypothesis is based on the fact that FGL proved to act as a neuroprotective, neurotrophic, and anti-inflammatory agent in different in vitro and in vivo experimental studies. FGL has been designed as a 15 amino acid peptide synthesized from the interconnecting loop region of the second fibronectin type III module in the extracellular domain of NCAM.5,6 The peptide proved to bind to and activate the fibroblast growth factor receptor 1 (FGFR1).5 Neuroprotective effects have been observed in vitro using a model of oxygen–glucose deprivation as well as in vivo in a gerbil transient global ischemia model.7 Moreover, FGL affected long-term plasticity, synapse and dendritic spine structure, and synaptogenesis.8−10 Beneficial effects were also reported from a mouse aging model, a chronic stress model, and an Alzheimer’s model indicating neuroprotective and cognition-enhancing properties.11−14 The protective and trophic effects of FGL have been partly attributed to an anti-inflammatory action mediated by an impact on IL-4 secretion from glial cells, which in turn affects neuronal CD200 signaling. Based on an enhancement of these intercellular signaling pathways, the FGL peptide seems to reduce microglia activation and the associated release of pro-inflammatory mediators such as interleukin-1 beta.15 Based on its efficacy profile it is in clinical development for neurodegenerative disorders including Alzheimer’s disease, stroke, traumatic brain injury, and chronic stress. A phase I study already indicated that FGL is well tolerated and safe and exhibits a dose-related pharmacokinetic profile after intranasal administration.16
The experiments were planned to address the following hypotheses: (1) Based on its impact on neuronal plasticity and its anti-inflammatory effects, FGL interferes with the development of a hyperexcitable network and with ictogenesis in the kindling paradigm. (2) Based on its impact on neuronal plasticity including hippocampal neurogenesis,17 FGL might exert effects on kindling-associated alterations in cell proliferation and generation of newborn granule cells.
Results and Discussion
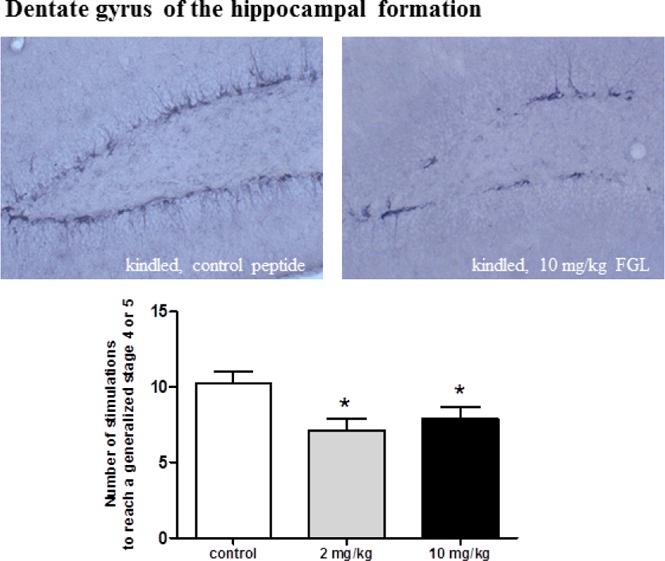
Impact of FGL on Seizure Thresholds and Kindling Progression
The impact of the FGL peptide on kindling progression was assessed at doses of 2 and 10 mg/kg with a pretreatment time of 60 min. In the early phase of kindling acquisition (stimulation day 1–12), neither the low nor the high dose of FGL exerted any effects on seizure severity (Figure 1A), afterdischarge duration (Figure 1B), and seizure duration (data not shown). However, mice treated with the FGL peptide required fewer stimulations to reach a generalized stage 4 or 5 (2 mg/kg, 7.125 ± 0.766; 10 mg/kg, 7.875 ± 0.812) compared with animals treated with the control peptide (10.25 ± 0.75) (Figure 1C,D). At stimulation days 13 to 16, the mean seizure severity tended to be lower in mice treated with 10 mg/kg FGL peptide compared with mice treated with the control peptide and mice treated with 2 mg/kg FGL peptide (Figure 1A). The difference from controls reached significance at stimulation day 13.
Figure 1.

Development of seizure severity (A) and afterdischarge duration (ADD) (B) during the kindling procedure. Data are shown for mice treated with the control peptide (n = 8), with 2 mg/kg of the FGL peptide (day 1–10, n = 8; day 11–16, n = 7), or with 10 mg/kg (n = 8) of the FGL peptide. Statistical analysis of data by Kruskal–Wallis followed by Dunn’s post hoc test indicated that FGL treatment did not influence kindling progression in the early phase (days 1–12). From day 13 to 16, treatment with 10 mg/kg of the FGL peptide tended to decrease the mean seizure severity reaching significance on day 13 compared with mice treated with the control peptide or with 2 mg/kg of the FGL peptide. The number of stimulations to reach a generalized stage 4 or 5 of mice treated with the control peptide (n = 8), with 2 mg/kg of the FGL peptide (n = 8), or with 10 mg/kg (n = 8) of the FGL peptide is given in panels C and D. Analyzed by one-way ANOVA followed by Newman–Keuls post hoc test, data from mice treated with 2 mg/kg or 10 mg/kg of the FGL peptide significantly differed from that for mice treated with the control peptide (p < 0.05). Significant differences are indicated by asterisks (p < 0.05). Data are given as means ± SEM.
Impact of FGL on Hippocampal Cell Proliferation and Neurogenesis in the Amygdala Kindling Model
Doublecortin (DCX) proved to be one of the key markers of hippocampal neuronal progenitor cells and early postmitotic neurons.18 By analysis of DCX immunohistochemistry 1 week following the last kindled seizure, we obtained information about the effects of kindling and treatment with the FGL peptide on this cell population in the dentate gyrus. As expected, kindling resulted in a significant rise in the number of neuronal progenitor cells (Figure 2C,D,F). In kindled mice of all treatment groups, the number of DCX-positive cells exceeded that in nonkindled mice. In animals receiving the control peptide, the expansion of this cell population reached 241% in kindled mice compared with nonkindled mice. Whereas 2 mg/kg of the FGL peptide did not exert significant effects, the higher dose of the FGL peptide efficaciously attenuated the kindling-associated increase in the number of DCX-positive cells in a significant manner (Figure 2E). The fraction of DCX-positive cells with hilar basal dendrites (HBDs) did not differ between nonkindled and kindled animals (Figure 2G). A comparison of the number of DCX-positive cells with HBDs among the three kindled groups did not reveal any significant differences (Figure 2G).
Figure 2.

Impact of FGL peptide on hippocampal neuronal progenitor cells. (A–E) Doublecortin (DCX) immunostaining in the dentate gyrus shown for representative mice from the different groups (nonkindled mice, (A) control peptide, (B) 10 mg/kg FGL peptide; kindled mice, (C) control peptide, (D) 2 mg/kg FGL peptide, (E) 10 mg/kg FGL peptide). Scale bar = 50 μm. (F) Analysis of DCX-positive cells in the dentate gyrus showed a significant increase in the number of DCX-positive cells in kindled mice of all treatment groups compared with nonkindled mice. In animals receiving the control peptide, the expansion of this cell population reached 241% in kindled mice compared with nonkindled mice. Whereas 2 mg/kg of the FGL peptide did not exert significant effects, the number of DCX-positive cells in kindled mice was significantly decreased by 10 mg/kg FGL compared with kindled mice treated with the control peptide and with 2 mg/kg FGL. (G) Analysis of the subset of DCX-positive cells with persistent hilar basal dendrites (HBDs) in the dentate gyrus did not indicate any significant group differences. Data are given as means ± SEM. Significant differences are indicated by asterisks (p < 0.05). Data in panels F and G were analyzed by two-way ANOVA followed by post hoc testing for individual differences by the Student’s t-test [nonkindled mice (control peptide, 10 mg/kg FGL) and kindled mice (control peptide, 10 mg/kg FGL)]. Kindled mice treated with 2 mg/kg FGL were compared separately with nonkindled mice treated with the control peptide and with kindled mice treated with the control peptide or 10 mg/kg FGL and analyzed by using Student’s t-test.
Kindling significantly increased the number of proliferating cells that incorporated BrdU during the stimulation phase as well as the number of newborn BrdU/NeuN-labeled neurons (Figure 3A–D) and BrdU/calbindin-labeled granule cells (Figure 4A–D). Neither the low nor the high dose of the FGL peptide prevented these cellular alterations in the dentate gyrus (Figure 3C,D; 4D).
Figure 3.

Impact of FGL peptide on hippocampal neurogenesis. (A, B) BrdU (red) and NeuN (green) immunostaining in the dentate gyrus shown for (A) nonkindled mice (control peptide) and (B) kindled mice (control peptide). (C) Analysis of BrdU-positive cells and (D) of BrdU/NeuN-double labeled cells showed a significant increase in kindled compared with nonkindled mice treated with the control peptide. Neither the low nor the high dose of FGL peptide prevented these cellular alterations in the dentate gyrus. Data are given as means ± SEM. Significant differences are indicated by asterisks (p < 0.05). Scale bar = 50 μm. Data in panels C and D were analyzed by two-way ANOVA followed by post hoc testing for individual differences by the Student’s t-test [nonkindled mice (control peptide, 10 mg/kg FGL) and kindled mice (control peptide, 10 mg/kg FGL)]. Kindled mice treated with 2 mg/kg FGL were compared separately with nonkindled mice treated with the control peptide and with kindled mice treated with the control peptide or 10 mg/kg FGL and analyzed by using Student’s t-test.
Figure 4.

Fluorescence micrographs. Calbindin (green) and BrdU (red) immunostaining is shown in an animal from the nonkindled group treated with the control peptide (A) and in an animal from the kindled group treated with the control peptide (B). A BrdU/calbindin double-labeled cell in an animal from the kindled group treated with the control peptide is shown in panel C. Scale bar is 20 μm in high magnification and 100 μm in the rest of the images. (D) Analysis of BrdU/calbindin double-labeled cells revealed that the cell numbers in kindled animals exceed those in nonkindled animals. Kindling significantly increased the number of BrdU/calbindin double-labeled cells in all treatment groups with no effect of the low or the high dose of the FGL peptide. Data are given as means ± SEM. Significant differences are indicated by asterisks (p < 0.05). Data were analyzed by two-way ANOVA followed by post hoc testing for individual differences by the Student’s t-test [nonkindled mice (control peptide, 10 mg/kg FGL) and kindled mice (control peptide, 10 mg/kg FGL)]. Kindled mice treated with 2 mg/kg FGL were compared separately with nonkindled mice treated with the control peptide and with kindled mice treated with the control peptide or 10 mg/kg FGL and analyzed by using Student’s t-test.
The prescence of newly generated astrocytes in the subgranular zone was confirmed by double-labeling the BrdU-positive cells with antiglial fibrillary acidic protein (GFAP). No noticeable differences in the number of GFAP-positive cells or in the number of BrdU/GFAP double-labeled cells among all groups (Figure 5A, B) were detected by visual inspection.
Figure 5.

Fluorescence micrographs. GFAP (green) and BrdU (red) immunostaining is shown exemplarily for an animal from the nonkindled group treated with the control peptide (A) and for an animal from the kindled group treated with the control peptide (B). No difference was evident among all treatment groups either in the number of GFAP-positive cells or in the number of BrdU/GFAP-positive cells. Scale bar is 5 μm in high magnification micrographs and 50 μm in the rest of the images.
The NCAM-derived peptide mimetic and FGFR1 ligand FGL is discussed as an interesting therapeutic.6,13,14 Pharmacokinetic studies have confirmed a relevant brain penetration rate of the peptide in rats and humans. FGL concentrations proved to be detectable in the cerebrospinal fluid of rats within 10 min following subcutaneous administration resulting in stable concentrations up to 5 h.19 In humans, concentrations in the cerebrospinal fluid have been analyzed following intranasal administration, thus confirming relevant brain access in translational studies.16
FGL proved to exert beneficial effects in different models of neurological or psychiatric diseases.6,7,11−14,20 In the present study, repeated administration of the peptide mimetic FGL affected kindling progression as well as associated alterations in the number of neuronal progenitor cells and early postmitotic cells. The impact of FGL on kindling progression is indicated by a reduced number of stimulations necessary to induce a generalized seizure. These data might suggest that FGL can promote the development of an epileptogenic network with FGF receptor mediated plasticity playing a role. In this context, it is of interest that FGF and its receptor can be regulated following seizure activity.21−25 A proconvulsant effect has been suggested by a study with genetic modulation of FGF2 expression. Whereas FGF2 knockout did not affect seizure susceptibility, FGF2 overexpression resulted in increased kainate-induced seizure severity in FGF2 overexpressing mice.25 More recently, it was reported that the FGF family members FGF22 and FGF7 can affect differentiation of excitatory and inhibitory synapses in distinct ways thereby also affecting seizure susceptibility in contrasting ways.26 In these experiments, FGF22-deficient mice exhibited resistance to epileptic seizures, whereas FGF7-deficient mice are prone to seizures.
When the currently ongoing clinical development of FGL as a therapeutic for central nervous system diseases is continued,16 a putative pro-epileptogenic potency should be kept in mind. However, experimental proof from a model with development of spontaneous seizures would be required to further confirm a respective risk. Moreover, the course of kindling acquisition in nontreated and treated animals also indicated a trend toward a biphasic effect. Whereas the data from days 5–8 would indicate an acceleration of kindling in response to FGL treatment, a trend toward a reduced seizure severity is indicated by data from the late kindling phase (days 13–16). Thus, FGL might enhance excitability in the naïve brain but might exert anticonvulsant effects in the epileptic brain once a hyperexcitable network has been generated.
Kindling has been repeatedly described to be associated with alterations in hippocampal cell proliferation, neuronal differentiation, and neurogenesis.27−30 This effect was confirmed in the present kindling experiment with significant increases in cell proliferation, the number of newborn neurons as well as the number of doublecortin-expressing neuronal progenitor cells and early postmitotic neurons. Whereas the low dose of FGL did not exert relevant effects on kindling associated increases in these cell populations, 10 mg/kg FGL proved to attenuate the expansion of the doublecortin-positive cell population. Considering that this was not accompanied by an effect on the number of BrdU-labeled cells, the effect of FGL seems to be rather related to an impact on neuronal differentiation and not to an impact on cell proliferation. In addition, it needs to be considered that related to the time course of neuronal progenitor development as well as our experimental planning, doublecortin expression rates will rather indicate effects of FGL during the second week of kindling. In contrast, BrdU was administered every other day during the whole kindling phase, so effects that might be evident during a selected time window might be diluted.
Earlier data have rather suggested a proneurogenic effect of FGF and its receptor.31,32 In view of these data, the findings with FGL indicate that this effect might be affected by pathophysiology or that the difference is rather related to the selective action of the FGL peptide sequence.
In conclusion, the effects of the peptide mimetic FGL in the kindling model do not confirm a disease-modifying effect with a beneficial impact on the development or course of epilepsy. The results obtained with FGL rather raise some concern regarding a putative effect, which might promote the formation of a hyperexcitable network. Future studies are required to further assess the risks in models with development of spontaneous seizures.
Material and Methods
Male NMRI mice were purchased at a bodyweight of 20–25 g (Harlan Winkelmann, An Venray, The Netherlands). Prior to surgery, animals were housed in groups of five animals for seven days under controlled environmental conditions (24–25 °C, 50–60% humidity, 12-h light/dark cycle) with free access to water and standard feed. To minimize the impact of circadian variation, all experiments were performed at the same time of the day (8 am to 1 pm). During the experimental phase, animals had a body weight between 30 and 50 g and were housed separately. Experimental procedures were performed according to the European Communities Council Directive of 24 November 1986 (86/609/EEC) and were approved by the responsible governmental administration of Upper Bavaria. All efforts were made to minimize pain or discomfort of the animals used.
Electrode Implantation
For implantation of kindling electrodes, mice (n = 45) were anesthetized with chloral hydrate (400 mg/kg ip). Bupivacaine was applied subcutaneously (sc) for additional local anesthesia before exposure of the skull surface. For analgesia, meloxicam (Metacam, Boehringer-Ingelheim, Ingelheim, Germany) was administered (1 mg/kg sc) 30 min prior to and 24 h postsurgery. The skull surface was exposed, and a Teflon-isolated bipolar stainless steel electrode was stereotactically implanted into the right amygdala (stereotaxic coordinates in millimeters relative to bregma: AP, −1.0; L, −3.2; and DV, −5.3). One screw, placed above the left parietal cortex, served as the indifferent reference electrode. Anchor screws were fixed to the skull to secure all mounted material, and the basolateral amygdala electrode was embedded in dental acrylic cement. Two mice died during surgery, three mice were euthanized following surgery due to alterations in their general condition.
Kindling Procedure
Kindling of mice (n = 24) was initiated following a postsurgical recovery period of 2 weeks. Kindling stimulations were performed at the same time of the day (between 9 am and 1 pm) to avoid intraday variance between animals. Initially, the stimulation threshold for eliciting afterdischarges (initial ADT, afterdischarge threshold) was determined for each animal using an ascending stair-step procedure as described previously.33,34 In 1 min intervals, the current (initially 8 μA) was increased in steps of approximately 20% of the previous current until afterdischarges were elicited. Twenty-four hours later, kindling was started using constant current stimulations (700 μA, 1 ms, monophasic square-wave pulses; 50 Hz for 1 s). Electrical stimulation of the amygdala was performed once daily and five times per week and was continued for 16 days until generalized stage 4 or 5 seizures were elicited in the majority of mice treated with the control peptide (Figure 6). Mice were not kindled or treated over the weekends. Following each amygdala stimulation, seizure severity was scored according to Racine:35 stage 1, immobility, eye closure, ear twitching, twitching of vibrissae, sniffing, facial clonus; stage 2, head nodding associated with more severe facial clonus; stage 3, clonus of one forelimb; stage 4, bilateral clonus of forelimbs accompanied by rearing; stage 5, rearing and falling accompanied by generalized clonic seizures. In addition to seizure severity, seizure duration and afterdischarge duration were recorded following each amygdala stimulation. A separate group of electrode-implanted control mice (n = 16) was not included in the kindling procedure, but was subjected to all handling and experimental procedures associated with the kindling procedure (defined as nonkindled mice). One mouse treated with 2 mg/kg of the FGL peptide died during kindling on the 12th day of stimulation. One nonkindled mouse treated with 10 mg/kg of the FGL peptide had to be euthanized due to alterations in the general condition before behavioral experiments were started. In both animals, no evidence was obtained that death or the alterations in the general condition are linked to FGL adverse effects.
Figure 6.

Time line of the study protocol indicating the timing of electrode implantation, kindling procedure, and treatment scheme. Following the kindling procedure, mice were used for behavioral tests and immunohistochemistry.
Treatment with the Neural Cell Adhesion Molecule-Derived Fibroblast Growth Factor Receptor Agonist Peptide FGL
The 15-amino-acid peptide, termed FGL (from FG loop) and a respective scrambled control peptide (control peptide) were synthesized by GL Biochem Ltd. (Shanghai, China). The sequence of FGL was derived from human NCAM (UniProtKB entry P13591). The FGL peptide (pentadecapeptid EVYVVAENQQGKSKA) and the control peptide (comprising the same amino acids as FGL but two glutamine residues were substituted with alanines) were created as a tetrameric dendrimer composed of four monomers coupled to a lysine backbone. The purity of both peptides reached more than 99% as estimated by high-performance liquid chromatography. Kindled mice were treated subcutaneously (sc) with either 10 mg/kg of the FGL control peptide (n = 8), 2 mg/kg of the FGL peptide (n = 8), or 10 mg/kg of the FGL peptide (n = 8) at each stimulation day (following determination of initial kindling afterdischarge thresholds) 60 min prior to kindling stimulation (Figure 6). Nonkindled mice received sc injections of the control peptide (10 mg/kg; n= 8) or the FGL peptide (10 mg/kg; n = 8) in accordance with this treatment protocol.
BrdU Labeling
As a thymidine analogue, 5′-bromodeoxyuridine (BrdU) is incorporated into the DNA during the S phase of the cell cycle.36 Each injection of BrdU labels those proliferating cells that are in the DNA-synthetic phase of the cell cycle (S phase). BrdU labeling gives information about only a limited time window. BrdU is available for about 30 min and thus labels a proportion of dividing cells that are in the S phase during this period.37 During the kindling procedure, all mice (kindled and nonkindled mice) received ip injections of 50 mg/kg BrdU three times per week every other day (Monday, Wednesday, Friday) immediately following the kindling stimulation. BrdU administrations did not influence the state or the performance of the animals.
Behavioral Evaluation
From day 20 to 22 (day 5–7 after the last kindled seizure, Figure 6), the behavior of the animals was evaluated in different paradigms including elevated plus maze (day 20), black–white box (day 21), and open field (day 22) using a video-tracking system with EthoVision software from Noldus (Wageningen, Netherlands). The behavioral analyses were of interest as we hypothesized that treatment with the FGL peptide might affect epilepsy-associated psychiatric comorbidities. Therefore, behavior was evaluated in kindled mice [treated with the control peptide (10 mg/kg, n = 8), 2 mg/kg of the FGL peptide (n = 7) or 10 mg/kg of the FGL peptide (n = 8)] and nonkindled mice [treated with the control peptide (10 mg/kg, n= 7) or the FGL peptide (10 mg/kg, n = 8)]. In behavioral analyses, FGL did not show any relevant effects. For this reason, data are not shown.
Tissue Preparation
Following the behavioral paradigms, mice were deeply anesthetized by pentobarbital overdose and transcardially perfused with saline followed by 4% paraformaldehyde in 0.1 M phosphate buffered saline (pH 7.4). The brains were removed and cut in 40 μm coronal sections on a freezing microtome. Sections were stored at −80 °C in cryoprotecting solution (glycerol and 0.1 M phosphate buffer, pH 7.4, 1:1 in volume). Electrode localizations were verified in thionin-stained sections.
Immunohistochemistry of Brain Sections
For immunostaining of the neuronal progenitor marker doublecortin, free-floating 40 μm thick sections were washed, preincubated, and then treated with a primary antiserum overnight at 4 °C. The primary antibody used in this study was polyclonal goat anti-doublecortin (Santa Cruz Biotechnology, Santa Cruz, California, USA), 1:300.
Sections were washed and placed in biotin-labeled secondary antiserum (biotinylated donkey anti-goat 1:550; Jackson Immunoresearch Laboratories, West Grove, Pennsylvania, USA). Sections were then rinsed in TBS and incubated for 60 min in horseradish peroxidase-labeled streptavidin (1:4000; Dianova, Hamburg, Germany). The immunohistochemical staining was finalized with the nickel-intensified diaminobenzidine reaction (0.05% 3.3-diaminobenzidine, 0.01% nickel ammonium sulfate, both from Sigma, and 0.01% H2O2). All sections were washed, mounted onto glass slides, air-dried, dehydrated, and covered with a coverslip with Entellan (Merck, Darmstadt, Germany). Due to damage of some brain sections, one nonkindled animal treated with the control peptide had to be excluded from the evaluation of doublecortin-labeled cells and of doublecortin-positive cells with hilar basal dendrites.
Newborn neurons were identified by double-staining for BrdU and NeuN. After washing in TBS, all sections were incubated in formamid (Sigma, Taufkirchen, Germany) for 2 h at 65 °C. Sections were then incubated in 2 N HCl for 30 min at 37 °C and washed in 0.1 M borate buffer (pH 8.5) for 10 min.40−42 Blocking was performed as described above (by using donkey serum). Sections were incubated in an antibody mixture containing rat anti-BrdU (Serotec, Kidlington, Oxfordshire, UK), 1:30, and anti-NeuN (Millipore, Billerica, Massachusetts), 1:500, at 4 °C overnight. Biotinylated donkey anti-mouse 1:500 (DAKO, Hamburg, Germany) and carbocyanin 3-labeled donkey anti-rat (Jackson Immunoresearch Laboratories, West Grove, Pennsylvania, USA), 1:1000, were applied for 60 min. The sections were washed and incubated in carbocyanin 2-labeled streptavidin (Jackson Immunoresearch Laboratories, West Grove, Pennsylvania, USA), 1:2000, for 60 min. For covering with coverslips, sections were treated as described above. Newborn granule neurons were identified by BrdU/calbindin double-labeling. Pretreatment was performed as described above. Primary antibody mixture contained rat anti-BrdU (AbD Serotec, UK; 1:300) and rabbit anti-calbindin (Abcam, Germany; 1:500). Carbocyanin 3-labeled donkey anti-rat antibody (Jackson Immunoresearch Laboratories, USA; 1:1000) and biotinylated donkey anti-rabbit (Jackson Immunoresearch Laboratories, USA; 1:2000) followed by carbocyanin 2-labeled streptavidin (Jackson Immunoresearch Laboratories, 1:2000) were used for detection. Covering with coverslips was performed as described above.
Astrocytic proliferation was verified by BrdU/GFAP double-labeling. Pretreatment was performed as described above. Primary antibody mixture contained rat anti-BrdU (AbD Serotec, UK; 1:300) and mouse anti-GFAP (Millipore, Germany; 1:500). Carbocyanin 3-labeled donkey anti-rat antibody (Jackson Immunoresearch Laboratories, USA; 1:1000) and biotinylated donkey anti-mouse (Jackson Immunoresearch Laboratories, USA; 1:500) followed by carbocyanin 2-labeled streptavidin (Jackson Immunoresearch Laboratories, 1:2000) were used for detection. Covering with coverslips was performed as described above.
Histological Evaluation of Brain Sections
The number of doublecortin-labeled cells as well as the number of doublecortin-labeled cells with hilar basal dendrites in the dentate gyrus of the hippocampus was quantified by stereological analysis using the computer-assisted imaging system StereoInvestigator 6.0 (Microbrightfield Europe, Magdeburg, Germany). The hardware consists of a Leica DMLB microscope (Leica, Bensheim, Germany), a Plan-Neofluar lens (Leica, Bensheim, Germany), a single chip charge coupled device color camera (CX9000, Microbrightfield Europe, Magdeburg, Germany), and an AMD Athlon (tm) 64 Processor. An experimenter unaware of the treatment conditions traced the extent of the dentate hilus and the hippocampal dentate gyrus and performed the counting of cells using the optical fractionator method. In slide-mounted sections (five sections per animal with interval 200 μm), the area of the dentate hilus as well as the dentate gyrus was traced, and within each contour, a step grid was placed. Counting frames were automatically and randomly placed along the grid. The thickness of the counting frame was equal to the thickness of the section (minus guard zones from the top and bottom of the section). Only those immunoreactive cells that appeared within the counting frame and came into focus were counted. Cell counts of fluorescent signals from double-staining for BrdU and NeuN and for BrdU and calbindin were performed at 24× magnification in an area encompassing the entire dentate granule cell layer (superior and inferior blades) and extending approximately two cell body widths deep into the hilus in at least three sections per animal. Double-labeling was verified by careful analysis of the confocal z-series of multiple cells per animal. Double-labeled cells of the dentate hilus of the hippocampal formation were counted separately, using the same method and the same sections. The hilus was defined as the inner border of the granule cell layer and two straight lines connecting the tips of the granule cell layer and the proximal end of the Cornu Ammonis (CA) 3c region. To ensure that the antibody actually penetrated the entire section, we used the free floating method and confirmed penetration by microscope z-scanning. BrdU/calbindin double-labeled cells were counted in kindled mice [treated with the control peptide (10 mg/kg, n = 4) or 2 mg/kg (n = 5) or 10 mg/kg of the FGL peptide (n = 4)] and nonkindled mice [treated with the control peptide (10 mg/kg, n= 5) or the FGL peptide (10 mg/kg, n = 4)].
Statistical Analysis
Statistical analysis was performed using GraphPad Prism 5. Differences between groups in kindling progression experiments (seizure severity, afterdischarge duration) were calculated by Kruskal–Wallis followed by Dunn’s post hoc test. For statistical analysis of the number of stimulations to reach a generalized seizure and the number of stage 4 or 5 seizures, we used one-way ANOVA followed by Newman–Keuls post hoc test. Statistical differences in immunohistological data (proliferation rates and neurogenesis) between nonkindled mice (control peptide, 10 mg/kg FGL) and kindled mice (control peptide, 10 mg/kg FGL) were calculated by two-way analysis of variance (ANOVA) followed by post hoc testing for individual differences by the Student’s t-test. Differences in immunohistological data between kindled mice treated with 2 mg/kg FGL and nonkindled mice treated with the control peptide as well as kindled mice treated with the control peptide or 10 mg/kg FGL were compared and analyzed by using Student’s t-test. All statistical tests were performed two-sided. Data are expressed as mean ± SEM. A value of p < 0.05 was considered statistically significant.
Acknowledgments
This research was supported by a grant of the Deutsche Forschungsgemeinschaft (No. DFG FOR 1103; PO-681/5-2). The authors thank Marion Fisch, Barbara Kohler, Carmen Meyer, Claudia Siegl, Angela Vicidomini, and Andrea Wehmeyer for their technical assistance.
The authors declare no competing financial interest.
References
- Pitkanen A. (2010) Therapeutic approaches to epileptogenesis--hope on the horizon. Epilepsia 51(Suppl 3), 2–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loscher W.; Brandt C. (2010) Prevention or modification of epileptogenesis after brain insults: Experimental approaches and translational research. Pharmacol. Rev. 62, 668–700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ravizza T.; Noe F.; Zardoni D.; Vaghi V.; Sifringer M.; Vezzani A. (2008) Interleukin converting enzyme inhibition impairs kindling epileptogenesis in rats by blocking astrocytic IL-1beta production. Neurobiol. Dis. 31, 327–333. [DOI] [PubMed] [Google Scholar]
- Vezzani A.; Friedman A. (2011) Brain inflammation as a biomarker in epilepsy. Biomarkers Med. 5, 607–614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kiselyov V. V.; Skladchikova G.; Hinsby A. M.; Jensen P. H.; Kulahin N.; Soroka V.; Pedersen N.; Tsetlin V.; Poulsen F. M.; Berezin V.; Bock E. (2003) Structural basis for a direct interaction between FGFR1 and NCAM and evidence for a regulatory role of ATP. Structure 11, 691–701. [DOI] [PubMed] [Google Scholar]
- Berezin V.; Bock E. (2010) NCAM mimetic peptides: An update. Adv. Exp. Med. Biol. 663, 337–353. [DOI] [PubMed] [Google Scholar]
- Skibo G. G.; Lushnikova I. V.; Voronin K. Y.; Dmitrieva O.; Novikova T.; Klementiev B.; Vaudano E.; Berezin V. A.; Bock E. (2005) A synthetic NCAM-derived peptide, FGL, protects hippocampal neurons from ischemic insult both in vitro and in vivo. Eur. J. Neurosci. 22, 1589–1596. [DOI] [PubMed] [Google Scholar]
- Popov V. I.; Medvedev N. I.; Kraev I. V.; Gabbott P. L.; Davies H. A.; Lynch M.; Cowley T. R.; Berezin V.; Bock E.; Stewart M. G. (2008) A cell adhesion molecule mimetic, FGL peptide, induces alterations in synapse and dendritic spine structure in the dentate gyrus of aged rats: A three-dimensional ultrastructural study. Eur. J. Neurosci. 27, 301–314. [DOI] [PubMed] [Google Scholar]
- Cambon K.; Hansen S. M.; Venero C.; Herrero A. I.; Skibo G.; Berezin V.; Bock E.; Sandi C. (2004) A synthetic neural cell adhesion molecule mimetic peptide promotes synaptogenesis, enhances presynaptic function, and facilitates memory consolidation. J. Neurosci. 24, 4197–4204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dallerac G.; Perronnet C.; Chagneau C.; Leblanc-Veyrac P.; Samson-Desvignes N.; Peltekian E.; Danos O.; Garcia L.; Laroche S.; Billard J. M.; Vaillend C. (2011) Rescue of a dystrophin-like protein by exon skipping normalizes synaptic plasticity in the hippocampus of the mdx mouse. Neurobiol. Dis. 43, 635–641. [DOI] [PubMed] [Google Scholar]
- Borcel E.; Perez-Alvarez L.; Herrero A. I.; Brionne T.; Varea E.; Berezin V.; Bock E.; Sandi C.; Venero C. (2008) Chronic stress in adulthood followed by intermittent stress impairs spatial memory and the survival of newborn hippocampal cells in aging animals: Prevention by FGL, a peptide mimetic of neural cell adhesion molecule. Behav. Pharmacol. 19, 41–49. [DOI] [PubMed] [Google Scholar]
- Klementiev B.; Novikova T.; Novitskaya V.; Walmod P. S.; Dmytriyeva O.; Pakkenberg B.; Berezin V.; Bock E. (2007) A neural cell adhesion molecule-derived peptide reduces neuropathological signs and cognitive impairment induced by Abeta25–35. Neuroscience 145, 209–224. [DOI] [PubMed] [Google Scholar]
- Ojo B.; Rezaie P.; Gabbott P. L.; Cowely T. R.; Medvedev N. I.; Lynch M. A.; Stewart M. G. (2011) A neural cell adhesion molecule-derived peptide, FGL, attenuates glial cell activation in the aged hippocampus. Exp. Neurol. 232, 318–328. [DOI] [PubMed] [Google Scholar]
- Ojo B.; Rezaie P.; Gabbott P. L.; Davies H.; Colyer F.; Cowley T. R.; Lynch M.; Stewart M. G. (2012) Age-related changes in the hippocampus (loss of synaptophysin and glial-synaptic interaction) are modified by systemic treatment with an NCAM-derived peptide, FGL. Brain, Behav., Immun. 26, 778–788. [DOI] [PubMed] [Google Scholar]
- Downer E. J.; Cowley T. R.; Lyons A.; Mills K. H.; Berezin V.; Bock E.; Lynch M. A. (2010) A novel anti-inflammatory role of NCAM-derived mimetic peptide, FGL. Neurobiol. Aging 31, 118–128. [DOI] [PubMed] [Google Scholar]
- Anand R.; Seiberling M.; Kamtchoua T.; Pokorny R. (2007) Tolerability, safety and pharmacokinetics of the FGLL peptide, a novel mimetic of neural cell adhesion molecule, following intranasal administration in healthy volunteers. Clin. Pharmacokinet. 46, 351–358. [DOI] [PubMed] [Google Scholar]
- Ojo B.; Gabbott P. L.; Rezaie P.; Corbett N.; Medvedev N. I.; Cowley T. R.; Lynch M. A.; Stewart M. G. (2013) An NCAM mimetic, FGL, alters hippocampal cellular morphometry in young adult (4 month-old) rats. Neurochem. Res. 38, 1208–1218. [DOI] [PubMed] [Google Scholar]
- Couillard-Despres S.; Winner B.; Schaubeck S.; Aigner R.; Vroemen M.; Weidner N.; Bogdahn U.; Winkler J.; Kuhn H. G.; Aigner L. (2005) Doublecortin expression levels in adult brain reflect neurogenesis. Eur. J. Neurosci. 21, 1–14. [DOI] [PubMed] [Google Scholar]
- Secher T.; Novitskaia V.; Berezin V.; Bock E.; Glenthoj B.; Klementiev B. (2006) A neural cell adhesion molecule-derived fibroblast growth factor receptor agonist, the FGL-peptide, promotes early postnatal sensorimotor development and enhances social memory retention. Neuroscience 141, 1289–1299. [DOI] [PubMed] [Google Scholar]
- Pedersen M. V.; Helweg-Larsen R. B.; Nielsen F. C.; Berezin V.; Bock E.; Penkowa M. (2008) The synthetic NCAM-derived peptide, FGL, modulates the transcriptional response to traumatic brain injury. Neurosci. Lett. 437, 148–153. [DOI] [PubMed] [Google Scholar]
- Bugra K.; Pollard H.; Charton G.; Moreau J.; Ben-Ari Y.; Khrestchatisky M. (1994) aFGF, bFGF and flg mRNAs show distinct patterns of induction in the hippocampus following kainate-induced seizures. Eur. J. Neurosci. 6, 58–66. [DOI] [PubMed] [Google Scholar]
- Gomez-Pinilla F.; van der Wal E. A.; Cotman C. W. (1995) Possible coordinated gene expressions for FGF receptor, FGF-5, and FGF-2 following seizures. Exp. Neurol. 133, 164–174. [DOI] [PubMed] [Google Scholar]
- Humpel C.; Lippoldt A.; Chadi G.; Ganten D.; Olson L.; Fuxe K. (1993) Fast and widespread increase of basic fibroblast growth factor messenger RNA and protein in the forebrain after kainate-induced seizures. Neuroscience 57, 913–922. [DOI] [PubMed] [Google Scholar]
- Van Der Wal E. A.; Gomez-Pinilla F.; Cotman C. W. (1994) Seizure-associated induction of basic fibroblast growth factor and its receptor in the rat brain. Neuroscience 60, 311–323. [DOI] [PubMed] [Google Scholar]
- Zucchini S.; Barbieri M.; Simonato M. (2005) Alterations in seizure susceptibility and in seizure-induced plasticity after pharmacologic and genetic manipulation of the fibroblast growth factor-2 system. Epilepsia 46(Suppl 5), 52–58. [DOI] [PubMed] [Google Scholar]
- Terauchi A.; Johnson-Venkatesh E. M.; Toth A. B.; Javed D.; Sutton M. A.; Umemori H. (2010) Distinct FGFs promote differentiation of excitatory and inhibitory synapses. Nature 465, 783–787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jafari M.; Soerensen J.; Bogdanovic R. M.; Dimou L.; Gotz M.; Potschka H. (2012) Long-term genetic fate mapping of adult generated neurons in a mouse temporal lobe epilepsy model. Neurobiol. Dis. 48, 454–463. [DOI] [PubMed] [Google Scholar]
- Parent J. M.; Janumpalli S.; McNamara J. O.; Lowenstein D. H. (1998) Increased dentate granule cell neurogenesis following amygdala kindling in the adult rat. Neurosci. Lett. 247, 9–12. [DOI] [PubMed] [Google Scholar]
- Pekcec A.; Lupke M.; Baumann R.; Seifert H.; Potschka H. (2011) Modulation of neurogenesis by targeted hippocampal irradiation fails to affect kindling progression. Hippocampus 21, 866–876. [DOI] [PubMed] [Google Scholar]
- Scott B. W.; Wang S.; Burnham W. M.; De Boni U.; Wojtowicz J. M. (1998) Kindling-induced neurogenesis in the dentate gyrus of the rat. Neurosci. Lett. 248, 73–76. [DOI] [PubMed] [Google Scholar]
- Mudo G.; Bonomo A.; Di Liberto V.; Frinchi M.; Fuxe K.; Belluardo N. (2009) The FGF-2/FGFRs neurotrophic system promotes neurogenesis in the adult brain. J. Neural Transm. 116, 995–1005. [DOI] [PubMed] [Google Scholar]
- Zhao M.; Li D.; Shimazu K.; Zhou Y. X.; Lu B.; Deng C. X. (2007) Fibroblast growth factor receptor-1 is required for long-term potentiation, memory consolidation, and neurogenesis. Biol. Psychiatry 62, 381–390. [DOI] [PubMed] [Google Scholar]
- Fischborn S. V.; Soerensen J.; Potschka H. (2010) Targeting the prostaglandin E2 EP1 receptor and cyclooxygenase-2 in the amygdala kindling model in mice. Epilepsy Res. 91, 57–65. [DOI] [PubMed] [Google Scholar]
- Pekcec A.; Unkruer B.; Schlichtiger J.; Soerensen J.; Hartz A. M.; Bauer B.; van Vliet E. A.; Gorter J. A.; Potschka H. (2009) Targeting prostaglandin E2 EP1 receptors prevents seizure-associated P-glycoprotein up-regulation. J. Pharmacol. Exp. Ther. 330, 939–947. [DOI] [PubMed] [Google Scholar]
- Racine R. J. (1972) Modification of seizure activity by electrical stimulation. I. After-discharge threshold. Electroencephalogr. Clin. Neurophysiol. 32, 269–279. [DOI] [PubMed] [Google Scholar]
- del Rio J. A.; Soriano E. (1989) Immunocytochemical detection of 5′-bromodeoxyuridine incorporation in the central nervous system of the mouse. Dev. Brain Res. 49, 311–317. [DOI] [PubMed] [Google Scholar]
- Packard D. S. Jr.; Menzies R. A.; Skalko R. G. (1973) Incorportaiton of thymidine and its analogue, bromodeoxyuridine, into embryos and maternal tissues of the mouse. Differentiation 1, 397–404. [DOI] [PubMed] [Google Scholar]
- Chatterjee S.; Mizar P.; Cassel R.; Neidl R.; Selvi B. R.; Mohankrishna D. V.; Vedamurthy B. M.; Schneider A.; Bousiges O.; Mathis C.; Cassel J. C.; Eswaramoorthy M.; Kundu T. K.; Boutillier A. L. (2013) A novel activator of CBP/p300 acetyltransferases promotes neurogenesis and extends memory duration in adult mice. J. Neurosci. 33, 10698–10712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kohman R. A.; Bhattacharya T. K.; Kilby C.; Bucko P.; Rhodes J. S. (2013) Effects of minocycline on spatial learning, hippocampal neurogenesis and microglia in aged and adult mice. Behav. Brain Res. 242, 17–24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ouchi Y.; Banno Y.; Shimizu Y.; Ando S.; Hasegawa H.; Adachi K.; Iwamoto T. (2013) Reduced adult hippocampal neurogenesis and working memory deficits in the Dgcr8-deficient mouse model of 22q11.2 deletion-associated schizophrenia can be rescued by IGF2. J. Neurosci. 33, 9408–9419. [DOI] [PMC free article] [PubMed] [Google Scholar]