

Abstract
Stroke causes life long disabilities where few therapeutic options are available. Using electrical and magnetic stimulation of the brain and physical rehabilitation, recovery of brain function can be enhanced even late after stroke. Animal models support this notion, and housing rodents in an enriched environment (EE) several days after experimental stroke stimulates lost brain function by multisensory mechanisms. We studied the dynamics of functional recovery of rats with a lesion to the fore and hind limb motor areas induced by photothrombosis (PT), and with subsequent housing in either standard (STD) or EE. In this model, skilled motor function is not significantly enhanced by enriched housing, while the speed of recovery of sensori-motor function substantially improves over the 9-week study period. In particular, this stroke lesion completely obliterates the fore and hind limb placing ability when visual and whisker guidance is prevented, a deficit that persists for up to 9 weeks of recovery, but that is markedly restored within 2 weeks by enriched housing. Enriched housing after stroke also leads to a significant loss of perineuronal net (PNN) immunoreactivity; detection of aggrecan protein backbone with AB1031 antibody was decreased by 13–22%, and labelling of a glycan moiety of aggrecan with Cat-315 antibody was reduced by 25–30% in the peri-infarct area and in the somatosensory cortex, respectively. The majority of these cells are parvalbumin/GABA inhibitory interneurons that are important in sensori-information processing. We conclude that damage to the fore and hind limb motor areas provides a model of loss of limb placing response without visual guidance, a deficit also seen in more than 50% of stroke patients. This loss is amenable to recovery induced by multiple sensory stimulation and correlates with a decrease in aggrecan-containing PNNs around inhibitory interneurons. Modulating the PNN structure after ischemic damage may provide new therapies enhancing tactile/proprioceptive function after stroke.
Introduction
Stroke is the leading cause of disability in developed countries [1] and leaves 50% of the patients with motor deficits [2], and 50 to 80% of patients with loss of somatosensory functions, such as sense of limb position and touch discrimination [3]. Although lost neurological function recovers to some extent [4], 30% of stroke patients remain severely disabled through life [5]. Proprioception-based abilities such as locating the limb without looking, directing a limb to a given point, judging structure and weight of an object, are often impaired after stroke and very disabilitating [3], [6]. Transcranial magnetic stimulation, direct current stimulation, physical and cognitive rehabilitative training improve various neurological modalities even when treatment is instituted several months after stroke [7]–[9], demonstrating the innate capacity of the brain to remodel and recover some lost functions [10], [11]. This brain plasticity is considered to be the basis for spontaneous and training promoted recovery after stroke, and encompasses a set of sensori-motor and cognitive mechanisms that promote the brain to adapt to new behavioral challenges.
In the experimental setting, housing animals in an enriched environment (EE) stimulates brain plasticity by multimodal sensory, cognitive and social stimulations [12]. Enriched housing improves performance in healthy animals [13], [14] as well as in models of various central nervous system (CNS) diseases and injuries such as Huntington's disease [15], Alzheimer's disease [16], Parkinson's disease [17], fragile X syndrome [18], Down's syndrome [19] and traumatic brain injury [20].
After stroke, enriched housing improves recovery in various behavioral tests [21], [22] by stimulating brain plasticity mechanisms including synaptogenesis, growth of axons or dendrites, gliogenesis and angiogenesis [23], as well as increasing sensitivity of neurons to activation, i.e. Hebbian or homeostatic plasticity [10]. The detailed mechanisms involved in the functional recovery processes are still poorly understood, but changes in gene expression [24], [25], attenuation of the inflammatory response [26], [27], increase in dendritic branching and spine density [28], [29] have been proposed to be involved.
The extracellular matrix (ECM) is important in regulating brain plasticity [30] but is also a potential hampering factor for recovery after injury [31]–[33]. Perineuronal nets (PNNs) are highly organized, lattice-like macromolecular structures of ECM that enwrap the surface of soma and proximal dendrites of neurons [34], [35], but with distinct “holes” at sites of synaptic contacts [36]. First described by Camillo Golgi, PNNs have since then been a matter of intense research particularly in the context of neural development, axon pathfinding and guidance, plasticity and regeneration after injury (for review, see [32], [37], [38]). The main components of PNNs are hyaluronan and the lectican family of chondroitin sulfate proteoglycans (CSPGs), aggrecan, brevican, neurocan, phosphacan and versican (for review, see [39], [40]), where aggrecan is the main constituent [41]. Only approximately 15% of forebrain neurons are enwrapped by PNNs [42], [43], and these cells are mainly parvalbumin-containing GABAergic (PV/GABA) positive [44]. PNNs are found in most CNS regions, and appear to stabilize synapses [45], [46]. The number of PNNs is reduced after traumatic brain injury [47], [48], or stroke induced by middle cerebral artery occlusion (MCAO) [49], [50] and photothrombosis (PT) [51], but whether this is a detrimental or beneficial event has not been assessed.
The aim of this study was twofold: (1) to investigate how enriched housing influences brain function in a model of chronic stroke with damage to the fore and hind limb motor areas and (2) to study how enriched housing affects aggrecan-containing PNNs in the peri-infarct and somatosensory cortex after experimental stroke.
Materials and Methods
Permanent focal ischemia by PT
All experiments were carried out with the approval of the Malmö-Lund animal review board (ethical permit number: M 25-12) and according to the ARRIVE guidelines. Animals were housed under reverse light conditions, with the testing performed during the dark period when the rats are active. The experiments were carried out on male Sprague Dawley rats (8 weeks, Charles River). The animals were anesthetized by isoflurane (approximately 2% in O2 under spontaneous ventilation) and placed into a stereotaxic frame. Temperature was monitored during surgery using a rectal temperature probe and animals were kept at 37.0–37.5±0.2°C by means of a heating pad with feedback control (Table S1). A sagittal skin incision was made, subcutaneous connective tissue was removed and the skull was dried. Thereafter, the dye Rose Bengal (0.5 mL at 10 mg/mL) was injected in the tail vein. Two minutes after injection, the skull was illuminated with cold light (Schott, KL 1500 LCD) on an area of 8 to 4.5 mm for 15 min (from +4 to −4 mm antero-posterior and from 0.5 to 5 mm on the left from bregma). The tail and the scalp incisions were sutured and the rats transferred to their home cage. The functional deficit was assessed 2d after the onset of the stroke using the limb placement test and then regularly once a week.
Randomization and EE
On day 2 after PT, animals were randomized and kept either in pairs in standard laboratory cages (STD) or all together (7 animals) in EE cages for 9 weeks as described previously [26], [52] (Fig. 1). During the time of the experiments, the neurological function was evaluated by different behavioral tests as described below. No animal died during the surgeries and all the animals exhibited a sufficient deficit two days after stroke. In total, 14 rats were used in this study.
Figure 1. Experimental design.
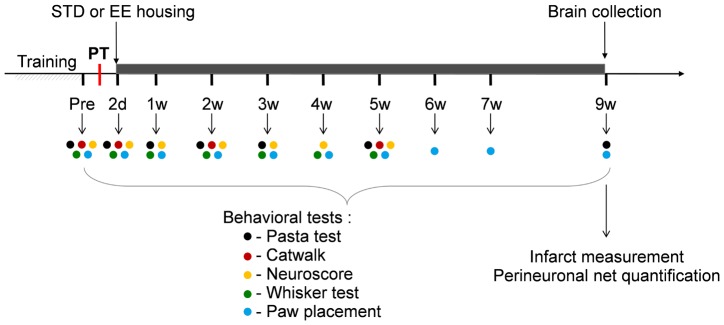
Rats were trained, and baseline values set for the different functional tests. Rats were then subjected to PT and the functional deficits assessed 2 days later. Subsequently, rats were placed either in standard (STD) or enriched environment (EE). Functional tests were regularly performed during 9 weeks following the onset of the stroke (the time of the different tests are represented by color dots). At 9 weeks, rats were perfusion fixed and brains were analyzed by immunohistochemistry.
Behavioral tests
Neuroscore
The functional deficit of the animals was assessed using a 28-point neuroscore test as described previously [53], [54]. This neuroscore uses eleven tests with a cumulative maximum score of 28: (1) circling behavior (maximum 4 points), (2) motility (maximum 3 points), (3) general condition (maximum 3 points), (4) righting reflex when placed on the back (maximum 1 point), (5) paw placement of each limb onto a table top (maximum 4 points), (6) ability to pull self up on a horizontal bar (maximum 3 points), (7) climbing on an inclined platform (maximum 3 points), (8) grip strength (maximum 2 points), (9) contralateral reflex (maximum 1 point) and (10) contralateral rotation when held by the base of tail (maximum 2 points), and (11) visual fore paw reaching (maximum 2 points). Scoring was determined on a scale starting from 0 for severe impairment to the maximum score for healthy function.
Paw Placement (PP)
This test provides information on the tactile/proprioceptive response to limb stimulation. Animals were placed with all 4 paws on the table top, and the paws to be tested along the edge. The rat was moved over the edge so the paws to be tested loose contact with the table surface. The ability of the animals to place the limb back onto the table surface when the rat was moved towards the edge was evaluated [55]. Importantly, the head was held at 45° angle, so that visual and whisker/snout stimulation was prevented. A prompt placement of the limb onto the table was scored as 1. Incomplete placing of the limb was scored as 0.5, and extension of the limb and paw was scored as 0.
Whisker paw placement test (WP)
The whisker paw placement test was performed as described in [56]. The fore limb placing assesses the ability of rats to sense tactile stimulus from the vibrissae and subsequently elicits a motor response of fore limb placing [57]. In this test, the animal's body was held gently such that the limbs were suspended off the experimenter's hand. The animal was then brought laterally to a tabletop, while the vibrissae brush against the edge of the table, allowing the animal to reach the edge with its fore paw. This was repeated 10 times on each side. The number of successful limb placements onto the edge of the table was recorded. A cross-midline variation to the test was also performed. Here, the animal was held on its side and successful placements of the opposite limb to the stimulated whisker were recorded.
Skilled reaching test (Pasta test)
Rats were kept under restricted feeding [58]. The aim of restricted feeding was to keep the rats 10% under the normal age/weight curve. Prior to stroke, the animals were weighted every day at the same time and, depending on their weight, they received between 22–30 g of pellet/2 rats in a cage. At 2 days after stroke, the animals received pellets (11–15 g/rat) placed at 4–5 different places in the EE cages. The STD housed rats received 11–15 g of pellet/rat, while weight was controlled three times/week. On the day before testing the amount of pellet/rat was reduced by 50%. For habituation to the pasta, four small pieces (4–5 cm long) were given to the rats in the cage, daily for 5–7 days, before initiating the training period. Animals were trained for 12 days prior to the test. For the test, the animals were placed into a Plexiglas cage with a small opening giving access to the pasta plate. The pasta plate included 10 rows of 14 holes filled with 3.5 mm long pasta pieces (2.5 cm available for grabbing). The plate was placed in front of the opening on the side of the lesion so that only pasta pieces could be reached with the paw contralateral to the lesion. The total number of pieces removed after 20 min was counted. In between tests, pasta was given twice a week into the cage.
Gait
Gait analysis was performed using the Catwalk automated gait analysis system (Noldus Information Technology, Wageningen, The Netherlands). Rats were trained on the walkway until 3 compliant (speed variation less than 55%, and between 15 and 115 cm.s−1) runs were successfully performed and recorded according to the manufacturers instructions. Thereafter, the recorded data were analyzed using the Catwalk XT 9.0 software. Each footprint was imaged and gait parameters were subsequently calculated to provide data on (1) time course of movement of each individual paw, (2) size and intensity of the contact of each individual paw, and (3) coordination (temporal and spatial) among the paws (Table S2).
Tissue collection
Rats were deeply anesthetized (isoflurane) and perfusion-fixed with 4% paraformaldehyde (PFA) solution. Brains were collected and immersed in 4% PFA solution for 4 h, then transferred into a 25% sucrose phosphate buffer solution (0.1 M NaH2PO4, 0.1 M Na2HPO4) and stored at 4°C. Brains were then cut in 30 μm slices using a microtome and one slice was kept each 1 mm for infarct size measurement. The others were kept for immunohistochemistry and immunofluorescence. All the slices were stored in anti-freeze solution (30% ethylene glycol, 30% glycerol, 0.01 M NaH2PO4, 0.03 M Na2HPO4) at -20°C.
Immunohistochemistry
Free-floating brain slices were rinsed three times in PBS (phosphate buffer saline) and quenched in 3% H2O2 and 10% MetOH for 15 minutes. As enhancing step for the AB1031 aggrecan staining (AB1031, Millipore), chondroitinase ABC (C3667, Sigma) digestion of the slices was first performed for 60 min at 37°C in Tris 50 mM, pH 8.0, sodium acetate 50 mM, 0.02% bovine serum albumin, containing 20 mU/mL lyase. After washing in PBS, the sections were blocked with blocking solution (5% normal donkey serum, Jackson ImmunoResearch, UK and 0.25% Triton X-100 in PBS) for one hour at room temperature. The sections were incubated overnight at 4°C with either of the following two primary antibodies: polyclonal rabbit anti-aggrecan (1∶1000, AB1031, Millipore), monoclonal mouse anti-chondroitin sulfate proteoglycan (1∶1000, protein core epitope, Cat-315/MAB1581, Millipore) diluted in blocking solution. Following three rinses with 1% normal donkey serum and 0.25% Triton X-100 in PBS, the sections were incubated with appropriate secondary biotinylated antibodies (donkey anti-rabbit/mouse, Vector Laboratories, USA) at a dilution of 1∶400 in blocking solution for 90 minutes at room temperature (RT). Visualization was achieved via the Vectorstain ABC kit (Vector) using 3,3-diaminobenzidine-tetrahydrochlorid (DabSafe, Saveen Werner, Sweden), 8% NiCl and 3% H2O2. Bright field pictures were acquired using an Olympus BX60 microscope (Solna, Sweden).
Immunofluorescence
Free-floating brain slices were rinsed three times in PBS and digested with chondroitinase ABC (C3667, Sigma; for details, see immunohistochemistry) as enhancing step for the AB1031 aggrecan staining (AB1031, Millipore). The sections were then blocked with blocking solution (5% normal donkey serum, Jackson ImmunoResearch, UK and 0.25% Triton X-100 in PBS) for one hour at RT. The sections were incubated overnight at 4°C with a primary antibody (polyclonal rabbit anti-aggrecan 1∶1000, AB1031, Millipore; monoclonal mouse anti-chondroitin sulfate proteoglycan 1∶1000, protein core epitope, Cat-315/MAB1581, Millipore; monoclonal goat anti-parvalbumin, 1∶2500, PV235, Swant) diluted in blocking solution. Following three rinses with 2% normal donkey serum and 0.25% Triton X-100 in PBS, the sections were incubated with appropriate secondary antibodies (donkey anti-rabbit/mouse/goat antibody) conjugated with either Cy3 or DyLight 488 fluorescent dyes (Jackson ImmunoResearch, UK) at a dilution of 1∶400 in blocking solution for 90 minutes at RT. Fluorescent dyes were imaged using a confocal laser-scanning microscope (Zeiss LSM 510, Jena, Germany).
Cell counting
Aggrecan-containing PNNs (AB1031 and Cat-315 positive) were visualized by immunohistochemistry in bright field. A 4× magnification objective was used to draw the boundaries of the regions of interest and to perform the actual cell counts by an investigator blinded to the group identity. AB1031 and Cat-315 positive PNNs were counted in the peri-infarct cortex (defined as 0.5 mm lateral from the ischemic core on the ipsilateral hemisphere and the correspondent region in the contralateral hemisphere) and in the somatosensory cortex (comprised between the peri-infarct cortex and 3.5 mm from the top of the slice on both hemispheres). Distances were defined using an optical grid. Two coronal brain sections for each animal of the study were used (level +1.20 mm in relation to bregma).
Infarct measurement
The brain slices were stained following the immunohistochemistry protocol described above using a monoclonal mouse anti-NeuN antibody (A60/MAB377, Millipore) at the dilution of 1∶1500. All slices were scanned, the non-lesioned area of the infarcted hemisphere and the non-lesioned contralateral hemisphere were outlined and measured using ImageJ. The infarct volume was then calculated by integration of the different areas [59].
Statistical analysis
For the analysis of PP, WP test and neuroscore, the Mann-Whitney test was used. For the analysis of skilled reaching test, the results were compared using two-factor ANOVA with post-hoc Tukeýs test. Regarding the analysis of infarct volume and cell counting, Student's t-test (unpaired) was used to compare the STD and EE groups. For the analysis of the effect of stroke on the gait (Catwalk) parameters, Student's t-test has been employed between values before stroke and values at 2 days of recovery (paired). For the analysis of differences in gait parameters between housing conditions during recovery, two-factor ANOVA was used, followed by the Student's t-test (unpaired) and Bonferroni corrections. All data are presented as mean ± SEM if not stated differently. Data and statistical processing were performed using Microsoft Excel and GraphPad Prism Software.
Results
Enriched housing does not affect infarct size after PT
Figures 2A and 2B show the location of the cortical infarct 9 weeks after induction of a PT lesion as displayed in NeuN stained coronal sections of a rat brain, along its rostro-caudal axis. The lesion extended 8 mm caudally from the frontal cortex, and included the fore and hind paw motor areas. The medial aspects, the cingulate and retrospinal cortices, were left intact, as were large parts of the somatosensory cortex. In all animals, the lesion extended from the cortical surface down to the corpus callosum, but did not transect the white matter. There was no difference in localization of the lesion between the STD (Fig. 2A) and EE (Fig. 2B) animals. Also, the mean size of the infarct in the STD animals at 9 weeks after PT stroke was 53±5 mm3, while in the animals housed in EE the mean infarct size was 52±7 mm3, and the fraction damaged cortex was the same between the groups (Fig. 2C). Hence, enriched housing starting 2 days after PT and continuing for 9 weeks did not influence the size of the infarct.
Figure 2. Infarct Volume.

Infarct volume was measured at 9 weeks after PT, and estimated by integration of the lesioned area on brain slices taken every 1-caudal brain axis. NeuN staining was performed to discriminate between intact and infarcted tissue. (A) Successive coronal slides of a representative brain infarct with their respective distances from the bregma for the STD housed animals, and (B) EE housed animals. (C) Infarct volume expressed as percentage of the hemisphere. Values are presented in box plots: median; 1st quartile to 3rd quartile; minimum and maximum value. (n = 7 for each housing condition; STD: standard environment, EE: enriched environment).
Enriched housing does not improve skilled reaching and does not affect gait
Skilled reaching test
A lesion to the motor cortex impairs paw functions in particular behavioral tests. In the pasta matrix reaching task, a test of skilled motor function [58], the number of pasta bars reached and grabbed during 20 min, before induction of stroke, was 41±6 in the STD group and 38±5 in the EE group (Fig. 3A). At 2 days after stroke, the reaching was markedly impaired and decreased by approximately 50% in both groups. At 1 week of recovery, i.e. 5 days in either STD or EE cages, the impairment remained significantly depressed and still at around 50% of the baseline values in both groups (p<0.001, Tukeýs test). Skilled motor function started to recover at 2 weeks and by 9 weeks recovered to 68% and 81% of baseline values in the STD and EE groups, respectively. The deficiency in reaching was mainly seen in the ability to reach the most distant rows of pasta pieces (Fig. 3B). Importantly, there was no significant difference between STD and EE groups at any time point of recovery.
Figure 3. Skilled reaching test.

Skilled reaching test at different times of recovery after PT stroke for animals housed in standard (STD) and enriched environment (EE). (A) The total number of pasta pieces reached during 20 minutes prior to PT (pre) and at different time points after PT. (#: p<0.05 when compared to the respective group before stroke (pre). (B) Map of the reaching area presenting the number of animals (as indicated by colors) that reached the pasta pieces (n = 7 for each housing condition).
Gait
Gait reflects the relative position of the body as seen in limb movement and positioning during locomotion over solid ground. Studies of gait following stroke also provide information on limb coordination and compensatory movements due to deficits in sensori-motor function. The Catwalk software provides 32 parameters describing the gait of the individual rat and thereby generates a large amount of data (Table S2). We analyzed gait at 2 days after PT in all animals prior to moving them into different housing conditions. Hence, at two days after stroke, animals performed with a reduced average speed and exhibited shifts in paw support and coordination (Table S3). Moreover, stroke induced asymmetry in the gait, especially in the hind paws (Table S4). The animals suffered from a lack of support on the paretic side and this results in an increased support on the non-paretic side (larger print area) but also on an accelerated swing, where the weight of the animal was on the paretic paw (shorter swing period, higher swing speed). This lack of support on the paretic paw, led to a longer support time on the non-paretic side before lifting up the affected paw (longer terminal dual stance). Similar but less pronounced changes in the initial/terminal dual stance and in the coupling of front paw were also seen (Fig. 4). We analyzed if there was a difference in gait parameters between the STD and EE groups at 2 and 5 weeks of recovery. No differences between the STD and EE groups were found (Table S5).
Figure 4. Effect of stroke on gait.

Presentation of gait parameters and the changes induced by PT. (A) Principle of the Catwalk. Rats run on a glass plate illuminated from the side. When the paws press the plate, light is diffracted and captured by a camera below the glass plate. The calculation of the gait parameters require the following steps: 1) the runs are recorded, 2) the light spots are associated with their respective paws, 3) the sequence of paw contact with the glass plate over time is determined and 4) the different gait parameters are calculated. (B) Stroke induces changes in different gait parameters. Here are summarized the general effect of stroke on gait two days after stroke. (C) Stroke also induces an asymmetry between left and right side in some gait parameters. The colored boxes represent time fractions when the paws are in contact with the surface of the glass plate. The black lines represent the values before stroke and the red lines the direction as well as increase or decrease of the values at 2 days after stroke (LH: left hind limb, RH: right hind limb). For details see Table S3.
Enriched housing enhances sensori-motor function after lesions to motor cortex
Neuroscore
General neurological function was assessed using the 28-point neuroscore. On day 2 after stroke, the median score was 23 in the two groups destined subsequently to either STD or EE (Fig. 5). Hence, the deficit was similar among the experimental groups before initiating the enriched housing. Also, a rapid recovery of most behavioral parameters was evident at 2 days after stroke in this test. Among the 11 tests, only three tests were affected by the stroke at 2 days of recovery: the paw placement (see below), the grip strength and the fore paw reaching test. The animals housed in the STD group showed a persistent deficit up to the 5th week, while the EE animals recovered almost completely at this time point.
Figure 5. Neuroscore.

The normal neuroscore before stroke is 28. Values at different times after PT in rats subsequently housed in standard (STD) and enriched environment (EE) are presented in box plot: median; 1st quartile to 3rd quartile, minimum and maximum value. (n = 7 for each housing condition) (*: p<0.05 when STD compared to EE; #: p<0.05 when STD and EE where compared to the respective values before stroke).
Whisker-paw test
The WP test showed a marked deficit at 2 days after PT in both the STD and EE groups of rats (score 0), (Fig. 6). By 1 to 2 weeks of recovery and housing in enriched condition, the animals regained full score on this test (score 10). In the STD housing group, recovery was delayed by one week. Hence, by 3 weeks of recovery, these animals also regained the motor response to whisker stimulation, first in the ipsilateral WP stimulation paradigm and a week later in the cross-line stimulation.
Figure 6. The whisker-paw test.

(A) Ipsilateral whisker test, and (B) cross-midline whisker test of rats subjected to PT at various times of recovery. Animals were placed in a standard (STD) or an enriched (EE) environment at 2 days after stroke. The normal score before stroke for rats is 10. Each dot corresponds to a rat value and the black line corresponds to the median value for the group. (n = 7 for each housing condition) (*: p<0.05 when STD compared to EE; #: p<0.05 when STD and EE where compared to the respective values before stroke).
Paw Placement test
A remarkable difference in behavioral recovery between the housing groups was found in the PP test under conditions that prevented visual and whisker/chin stimulation during testing. In this test, the fore and hind paws on the side of the lesion were unaffected in both groups. The contralateral paws in the STD housed group recovered slowly, and by 9 weeks, 50% of the animals still displayed a deficit in right fore paw function and all but one animal had the lowest score in the hind paw function (Fig. 7). In contrast, all animals housed in the EE group displayed a rapid dynamic recovery. Fore paw placement recovered completely by 2 weeks (but not at 1 week) of recovery and the recovery persisted up to the 9-week time point. The hind paw placement recovered slower, but by 9 weeks 70% of the animals showed recovery, a time point when only 1 animal displayed recovery in the STD housed group.
Figure 7. Paw placement test.

Limb placing ability of rats subjected to PT under conditions that prevented visual and whisker/chin stimulation during testing. (A) Fore paw placement, and (B) hind paw placement. The normal score before stroke for rats is 1. Each dot corresponds to a rat value and the black line corresponds to the median value for the group. (n = 7 for each housing condition) (*: p<0.05 when STD compared to EE; #: p<0.05 when STD and EE where compared to the respective values before stroke; STD: standard environment, EE: enriched environment).
Enriched housing promotes a decrease in aggrecan-containing PNNs around PV/GABA neurons
To establish a structural correlate to the recovery of somatosensory function promoted by enriched housing, we investigated the integrity of PNNs. It has earlier been shown that the number of PNNs is affected by stroke, after both MCAO [49], [50] and PT [51], but, to our knowledge, there are no data linking these observations in the cerebrum with differential housing conditions and behavior. Since the true complexity of the extracellular matrix can only be appreciated if several protein epitopes are studied, we determined the stroke-induced modulation/distribution of perineuronal net-bearing cells by means of two antibodies recognizing different epitopes of aggrecan, the major constituent of PNNs in the adult rat brain [41].
Two antibodies were used to target different epitopes on the aggrecan molecule: AB1031 [60], [61], which recognizes an epitope within the central protein domain of aggrecan at chondroitin sulfate glycosaminoglycan binding sites, and Cat-315, which labels a glycan moiety of aggrecan in the adult [41], [43], [62]. In the following, we will refer to the AB1031 immunopositive and Cat-315 immunopositive cells as AB1031+ and Cat-315+ respectively. We examined the number of AB1031+ and Cat-315+ PNNs in the peri-infarct and somatosensory cortex of both hemispheres of rats kept in either housing condition after PT (Fig. 8, 9).
Figure 8. Environmental stimulation decreases the number of AB1031+ PV/GABA neurons in the rat cerebral cortex after PT.

(A) Confocal images of parvalbumin expressing cells (green) enwrapped by AB1031+ PNNs (red) in the somatosensory cortex of a representative STD animal, scale bar 20 μm. (B and C) Representative bright-field micrographs of AB1031+ PNNs in the rat cerebral cortex ipsilateral to the lesion, scale bar 100 μm; (B1 and C1) higher magnification, scale bar 20 μm. AB1031 immunoreactivity in (B) STD and (C) EE conditions. Quantification of cortical neurons bearing AB1031+ PNNs in the peri-infarct cortex (D), the corresponding area contralateral to the lesion (E), the ipsilateral somatosensory cortex (F) and the contralateral somatosensory cortex (G). (n = 7 for each housing condition; 9 weeks after stroke; D with *p = 0.04; E with p = 0.3613; F with *p = 0.007; G with p = 0.0963; STD: standard environment, EE: enriched environment, PI: peri-infarct cortex, SS: somatosensory cortex, IPSI: ipsilateral, CONTRA: contralateral).
Figure 9. Enriched housing decreases the number of Cat-315+ PV/GABA neurons in the rat cerebral cortex after PT.

(A) Confocal images of a parvalbumin (PV) expressing cell (green) enwrapped by a Cat-315+ PNN (red) in the somatosensory cortex of a representative STD animal; Z-stack demonstrates close proximity of the Cat-315 aggrecan antibody and PV, scale bar 20 μm. (B and C) Representative bright-field micrographs of Cat-315+ PNNs in the rat cerebral cortex ipsilateral to the lesion, scale bar 100 μm; (B1 and C1) higher magnification, scale bar 20 μm. Cat-315 immunoreactivity denotes a critical difference between (B) STD and (C) EE conditions. Quantification of cortical neurons bearing Cat-315+ PNNs in the peri-infarct cortex (D), the corresponding area contralateral to the lesion (E), the ipsilateral somatosensory cortex (F) and the contralateral somatosensory cortex (G). (n = 7 for each housing condition; 9 weeks after stroke; D with *p = 0.04; E with p = 0.06; F with *p = 0.0007; G with p = 0.1; STD: standard environment, EE: enriched environment, PI: peri-infarct cortex, SS: somatosensory cortex, IPSI: ipsilateral, CONTRA: contralateral).
The aggrecan-positive PNNs surround neurons containing the calcium-binding protein parvalbumin [44], a widely accepted marker for GABAergic interneurons. We confirmed this notion with a double fluorescent staining for PV and both aggrecan antibodies (Fig. 8A, 9A), where we observed that the majority of PV+ neurons were also positive for either AB1031 or Cat-315 markers. We also confirmed the localization of PNN+ neurons in layers II, III, IV, V and VI (AB1031 staining, Fig. 8B, C) and in layers IV and VI (Cat-315 staining, Fig. 9B, C) in both the STD and EE groups.
In the injured hemisphere of animals housed in an EE, we found a significantly 13% reduction in the number of AB1031+ PNNs in the peri-infarct cortex (95±4 cells) when compared with animals housed in STD conditions (110±5 cells; Fig. 8D). In the homotopic area of the contralateral hemisphere there was no significant differences between the two groups (Fig. 8E; STD: 145±6 cells and EE: 138±5 cells). A significant 22% reduction in the number of AB1031+ PNNs was observed in the ipsilateral somatosensory cortex of the EE animals (433±19 cells) when compared to STD housing conditions (337±23 cells; Fig. 8F) while there was no difference in the contralateral hemisphere (Fig. 8G; STD: 542±16 cells and EE: 499±17 cells).
With regard to the Cat-315+ PNNs, we found a significant 30% decrease in the number of Cat-315+ PNNs (19±2 cells) in the peri-infarct area of EE animals (Fig. 9D) compared to the STD housed group (27±3 cells). In the homotopic area of the contralateral hemisphere, enriched housing also appeared to affect the number of Cat-315+ PNNs although the changes were not statistically significant (35±4 cells and 26±2 cells in the STD and EE groups respectively, p = 0.06, Fig. 9E). This tendency was also seen throughout the somatosensory cortex, where a 25% decrease of Cat-315+ PNNs in the EE group (146±5 cells) compared to the STD group (195±10 cells) was found (Fig. 9F). Again, no statistical significance was found in the homotopic region of the contralateral hemisphere, where we counted 263±14 cells in the STD conditions and 235±6 cells in the EE conditions (Fig. 9G).
Discussion
In the present investigation we show that housing rats in an EE following an ischemic lesion to the fore and hind limb motor areas, enhances the speed of neurological recovery, particularly the recovery of the marked and persistent loss of tactile/proprioceptive limb placing response. The recovery-enhancing effect of EE is not due to a decrease in infarct size and sparing of brain tissue, but correlates with a robust reduction of aggrecan-containing PNNs surrounding parvalbumin-containing GABAergic neurons in the ipsilateral surviving cortical tissue. In the following discussion we will focus on the enhancement of somatosensory functions by enriched housing, the possible underlying biological mechanisms and its clinical implications.
A model for somatosensory neglect and dysfunction
The PT model of stroke is advantageous in studying long-term recovery mechanisms after experimental stroke, since the localization and size of the lesion can be controlled with little variability among animals. With this model, plasticity processes affecting various brain regions, functions and cellular elements, which range from small infarcts induced by single vessel PT occlusions [63] to large brain lesions involving partial or entire functional entities such as the somatosensory cortex [64], [65], can be studied. Hence, depending on the size and the localization of the lesion, variable degree and speed of recovery is observed. With PT stroke, deficits in laterality, sensory or motor function and learning have been reported using the cylinder test, adhesive removal test [66], beam walking test, beam balanced test, corner test [67], rotarod [68], gap crossing test [69], Catwalk [70], skilled reaching, horizontal ladder test [71], neurological severity score [72], Morris water maze [73], plantar test [74], spontaneous activity and grip strength [75].
We induced the PT lesion in the fore and hind limb M1 areas [76], [77] leaving the medial agranular cortex and the somatosensory cortex essentially intact [78]. This lesion caused a deficit in several sensori-motor tests, albeit to varying degree and persistency. We confirm a deficit in skilled reaching that persisted throughout the 9 weeks of recovery [58]. Gait was affected 2 days post stroke, with changes in coordination, speed of movement as well as asymmetry in the support of the paws. The sensori-motor response to whisker stimulation, that was lost at day 2 after stroke, recovered within 2-4 weeks. The most dramatic effect on behavioral outcome was the deficit in tactile/proprioceptive response when visual stimuli and whisker/snout contact were prevented, as reported earlier [55], [78]. Here we found that the hind limb response was persistently depressed, while the fore limb function partially recovered by the 9th week of recovery. Evidently, the extensive lesion to M1 caused marked and persistent deficits in fine motor skills due to the damage of the cortico-spinal tract (CST) and loss of somatosensory functions with a variable degree of recovery. When visual and whisker/chin stimulations were prevented, the recovery of limb placement was markedly depressed (fore limb) or prevented (hind limb). Therefore, this cortical PT lesion appears to induce a specific somatosensory neglect [55], [78].
Enriched housing reverses sensory neglect and dysfunction
Using the EE paradigm, it is possible to assess the recovery potential of various lost brain functions following brain damage such as stroke. We initiated enriched housing at 2 days after PT, a time when the infarct can be considered fully matured. Subsequent enriched housing for 9 weeks did not influence infarct size, in line with earlier studies using this model [71]. Improved performance during enriched housing must therefore be associated with recovery of neuronal functions, connectivity and post-stroke brain plasticity.
Over the 9-week period studied, the animals housed in the EE did not improve skilled reaching. This is in accord with earlier findings using PT, where enriched housing promoted use of compensatory movements while fine motor skills essentially did not improve [71]. Presumably, the PT lesion damages the CST to such extent that little tissue is left to become substrate for network remodeling and recovery over the 9-week period studied. Longer periods of recovery could involve activation of the contralateral motor cortex [29], but this is not likely in a lesion of the present size [79]. Also, different results were obtained after transient and permanent occlusion of the middle cerebral artery, where the CST is largely preserved and amenable to plasticity and recovery [80], [81].
In contrast to the limited enhancement of fine skilled motor recovery by enriched housing, the speed of recovery of the sensori-motor deficit seen in the whisker tests at 1–2 weeks of recovery was markedly enhanced. Likewise, the tactile proprioceptive response (limb placement) where visual and whisker cues are hidden, displays an even more striking recovery enhancement induced by enriched housing. The lost fore limb placement fully recovered after 12 days of enriched housing. The hind limb placement response that never recovered in STD housed animals clearly responded after enriched housing.
Hence, we observe a beneficial effect of the enriched housing when using tests of sensory abilities and proprioception but no marked effect when using tests of skilled motor functions or locomotion. This suggests that the STD housed animals fail to integrate sensory stimuli to trigger a motor response, particularly when visual and whisker stimuli are absent. The EE, stimulating the somatosensory network involved in the sensori-motor response, evidently promotes cellular and network plasticity and provides improved recovery. The peri-infarct area has been proposed to constitute a growth permissive region that promotes axonal outgrowth and synaptogenesis [82]. The recovery of tactile sensation and proprioception could be envisaged to involve sensori-motor cortex also remote from the lesion. We therefore investigated changes in the integrity of PNNs, known to be affected by enriched housing [83], [84].
Enriched housing enhances reduction of aggrecan-containing PNNs after experimental stroke
We show here for the first time that the multisensory stimulation provided by enriched housing after experimental stroke reduces the number of aggrecan-containing PNNs in the peri-lesional area, as well as in the ipsilateral somatosensory cortex remote to the lesion.
CSPGs are generally up-regulated in the peri-infarct regions after experimental stroke as part of the glial scar formed to contain the infarct [50], [85]. However, in areas remote from the ischemic lesion, CSPGs are reduced [49], contributing to the formation of a growth-permissive region surrounding the infarct after injury [86]. Our data suggest that the multisensory stimulation provided by enriched housing further expands this window of experience-dependent plasticity in space and time after stroke, including more remote somatosensory cortical areas, or even the contralateral hemisphere, several months after stroke. This is in line with recent reports that degradation of CSPGs with chondroitinase ABC improves functional recovery after experimental stroke [87]–[89].
We also found that the Cat-315+ PNNs, staining a glycan moiety of aggrecan, are more reduced than the AB1031+ PNNs, staining the aggrecan protein backbone. This would indicate that a subset of neurons (Cat-315+) is particularly sensitive to enriched housing conditions after experimental stroke [62]. Earlier studies have demonstrated that sensory deprivation reduces the number of Cat-315+ PNNs [43], [90]. Still, we do not know if the reduction of aggrecan-containing PNNs in our study is due to enhanced degradation or a reduced synthesis of aggrecan after experimental stroke. The Cat-315+ neurons enwrap mainly parvalbumin-containing GABAergic neurons [43], [44], of crucial importance for processing of sensori-information [91] and hence also of importance in functional recovery after stroke. Indeed, the wide spread reduction of aggrecan-containing PNNs in the somatosensory cortex suggests changes in sensory processing enhanced by enriched housing and involving neural networks remote from the lesion.
Conclusions
The PT stroke lesion to fore and hind paw regions of the rat brain provides a tool and model to study the mechanisms of somatosensory dysfunction and recovery after stroke. We propose that multisensory stimulation provided by enriched housing reduces the density of PNNs surrounding PV/GABA neurons, improving cortical plasticity and sensori-information processing. Elucidating the mechanisms and pharmacology of PNN turnover after experimental stroke may provide new therapies supporting rehabilitation.
Supporting Information
Animal data. Parameters of the rats subjected to PT with subsequent housing in STD or EE.
(TIF)
Explanation of the Catwalk parameters. (a.u.: arbitrary units).
(TIF)
Gait parameters affected by stroke. The pre-stroke values and the values 2 days after stroke are presented (n = 14) as well as the percentage of variation between the time points, the p-value of the Student's t-test (pre vs. 2d), the Pearson's correlation coefficient with the speed and weight with the respective associated p-values. (RF: right front, RH: right hind, LF: left front, LH: left hind, a.u.: arbitrary unit).
(TIF)
Asymmetry in parameters of gait induced by stroke. Gait parameters where differences were seen between paretic and non-paretic side at 2 days after stroke. The pre-stroke values and the values 2 days after stroke are presented (n = 14) as well as the percentage of variation between the two side, the respective p-value of the Student's t-test (L vs. R) for both time and the p-value of the Student's t-test on the difference (Right-Left) before and 2d after stroke. (F: front, H: hind, L: left, R: right).
(TIF)
Recovery in parameters of the gait affected by stroke. Gait parameter values affected by stroke (general) and the difference between paretic and non-paretic side induced by stroke (asymmetry) at 2 and 5 weeks in rats housed in either STD or EE. (RF: right front, RH: right hind, LF: left front, LH: left hind, F: front, H: hind).
(TIF)
Acknowledgments
The authors wish to acknowledge Kerstin Beirup for excellent technical assistance in the immunohistochemistry and Marco Maccarana for advice on the methods.
Funding Statement
This study was supported by the Swedish Research Council grants no. 2011-2652 (TW) and no. 2011-2684 (KR), the EU 7th work program through the European Stroke Network grant no. 201024, the Hans-Christian and Alice Wachtmeister Foundation (TW, KR), the Swedish Brain Fund (TW), the Greta och Johan Kocks Stiftelser (KR), the Thorsten och Elsa Segerfalk Stiftelse (KR) and the Crafoord Foundation (KR). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Roger VL, Go AS, Lloyd Jones DM, Benjamin EJ, Berry JD, et al. (2012) Heart disease and stroke statistics--2012 update: a report from the American Heart Association. Circulation 125: e2–e220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Jørgensen HS, Nakayama H, Raaschou HO, Vive Larsen J, Støier M, et al. (1995) Outcome and time course of recovery in stroke. Part I: Outcome. The Copenhagen Stroke Study. Arch Phys Med Rehabil 76: 399–405. [DOI] [PubMed] [Google Scholar]
- 3. Carey LM, Oke LE, Matyas TA (1996) Impaired limb position sense after stroke: a quantitative test for clinical use. Arch Phys Med Rehabil 77: 1271–1278. [DOI] [PubMed] [Google Scholar]
- 4. Duncan PW, Goldstein LB, Matchar D, Divine GW, Feussner J (1992) Measurement of motor recovery after stroke. Outcome assessment and sample size requirements. Stroke 23: 1084–1089. [DOI] [PubMed] [Google Scholar]
- 5. Muntner P, Garrett E, Klag MJ, Coresh J (2002) Trends in stroke prevalence between 1973 and 1991 in the US population 25 to 74 years of age. Stroke 33: 1209–1213. [DOI] [PubMed] [Google Scholar]
- 6. Carey LM, Abbott DF, Harvey MR, Puce A, Seitz RJ, et al. (2011) Relationship between touch impairment and brain activation after lesions of subcortical and cortical somatosensory regions. Neurorehabil Neural Repair 25: 443–457. [DOI] [PubMed] [Google Scholar]
- 7. Kitago T, Liang J, Huang VS, Hayes S, Simon P, et al. (2013) Improvement after constraint-induced movement therapy: recovery of normal motor control or task-specific compensation? Neurorehabil Neural Repair 27: 99–109. [DOI] [PubMed] [Google Scholar]
- 8. Edwardson MA, Lucas TH, Carey JR, Fetz EE (2013) New modalities of brain stimulation for stroke rehabilitation. Exp Brain Res 224: 335–358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Schulz R, Gerloff C, Hummel FC (2013) Non-invasive brain stimulation in neurological diseases. Neuropharmacology 64: 579–587. [DOI] [PubMed] [Google Scholar]
- 10. Murphy TH, Corbett D (2009) Plasticity during stroke recovery: from synapse to behaviour. Nat Rev Neurosci 10: 861–872. [DOI] [PubMed] [Google Scholar]
- 11. Dancause N (2006) Neurophysiological and anatomical plasticity in the adult sensorimotor cortex. Rev Neurosci 580: 561–580. [DOI] [PubMed] [Google Scholar]
- 12. Nithianantharajah J, Hannan AJ (2006) Enriched environments, experience-dependent plasticity and disorders of the nervous system. Nat Rev Neurosci 7: 697–709. [DOI] [PubMed] [Google Scholar]
- 13. Hebb DO (1947) The effects of early experience on problem solving at maturity. Am Psychol 2: 306–307. [Google Scholar]
- 14. Van Praag H, Kempermann G, Gage FH (2000) Neural consequences of environmental enrichment. Nat Rev Neurosci 1: 191–198. [DOI] [PubMed] [Google Scholar]
- 15. Hockly E, Cordery PM, Woodman B, Mahal A, van Dellen A, et al. (2002) Environmental enrichment slows disease progression in R6/2 Huntington's disease mice. Ann Neurol 51: 235–242. [DOI] [PubMed] [Google Scholar]
- 16. Arendash GW, Garcia MF, Costa DA, Cracchiolo JR, Wefes IM, et al. (2004) Environmental enrichment improves cognition in aged Alzheimer's transgenic mice despite stable beta-amyloid deposition. Neuroreport 15: 1751–1754. [DOI] [PubMed] [Google Scholar]
- 17. Bezard E, Dovero S, Belin D, Duconger S, Jackson Lewis V, et al. (2003) Enriched environment confers resistance to 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine and cocaine: involvement of dopamine transporter and trophic factors. J Neurosci 23: 10999–11007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Restivo L, Ferrari F, Passino E, Sgobio C, Bock J, et al. (2005) Enriched environment promotes behavioral and morphological recovery in a mouse model for the fragile X syndrome. Proc Natl Acad Sci U S A 102: 11557–11562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Martínez Cué C, Rueda N, García E, Davisson MT, Schmidt C, et al. (2005) Behavioral, cognitive and biochemical responses to different environmental conditions in male Ts65Dn mice, a model of Down syndrome. Behav Brain Res 163: 174–185. [DOI] [PubMed] [Google Scholar]
- 20. Kovesdi E, Gyorgy AB, Kwon SK, Wingo DL, Kamnaksh A, et al. (2011) The effect of enriched environment on the outcome of traumatic brain injury; a behavioral, proteomics, and histological study. Front Neurosci 5: 1–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Janssen H, Bernhardt J, Collier JM, Sena ES, McElduff P, et al. (2010) An enriched environment improves sensorimotor function post-ischemic stroke. Neurorehabil Neural Repair 24: 802–813. [DOI] [PubMed] [Google Scholar]
- 22. Ohlsson AL, Johansson BB (1995) Environment influences functional outcome of cerebral infarction in rats. Stroke 26: 644–649. [DOI] [PubMed] [Google Scholar]
- 23. Wieloch T, Nikolich K (2006) Mechanisms of neural plasticity following brain injury. Curr Opin Neurobiol 16: 258–264. [DOI] [PubMed] [Google Scholar]
- 24. Rampon C, Jiang CH, Dong H, Tang YP, Lockhart DJ, et al. (2000) Effects of environmental enrichment on gene expression in the brain. Proc Natl Acad Sci 97: 12880–12884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Keyvani K, Sachser N, Witte OW, Paulus W (2004) Gene expression profiling in the intact and injured brain following environmental enrichment. J Neuropathol Exp Neurol 63: 598–609. [DOI] [PubMed] [Google Scholar]
- 26. Ruscher K, Johannesson E, Brugiere E, Erickson A, Rickhag M, et al. (2009) Enriched environment reduces apolipoprotein E (ApoE) in reactive astrocytes and attenuates inflammation of the peri-infarct tissue after experimental stroke. J Cereb Blood Flow Metab 29: 1796–1805. [DOI] [PubMed] [Google Scholar]
- 27. Inácio AR, Ruscher K, Wieloch T (2011) Enriched environment downregulates macrophage migration inhibitory factor and increases parvalbumin in the brain following experimental stroke. Neurobiol Dis 41: 270–278. [DOI] [PubMed] [Google Scholar]
- 28. Johansson BB, Belichenko PV (2002) Neuronal plasticity and dendritic spines: effect of environmental enrichment on intact and postischemic rat brain. J Cereb Blood Flow Metab 22: 89–96. [DOI] [PubMed] [Google Scholar]
- 29. Biernaskie J, Corbett D (2001) Enriched rehabilitative training promotes improved forelimb motor function and enhanced dendritic growth after focal ischemic injury. J Neurosci 21: 5272–5280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Frischknecht R, Gundelfinger ED (2012) The brain's extracellular matrix and its role in synaptic plasticity. Adv Exp Med Biol 970: 153–171. [DOI] [PubMed] [Google Scholar]
- 31. Rosell A, Lo EH (2008) Multiphasic roles for matrix metalloproteinases after stroke. Curr Opin Pharmacol 8: 82–89. [DOI] [PubMed] [Google Scholar]
- 32. Soleman S, Filippov MA, Dityatev A, Fawcett JW (2013) Targeting the neural extracellular matrix in neurological disorders. Neuroscience 253: 194–213. [DOI] [PubMed] [Google Scholar]
- 33. Lo EH, Dalkara T, Moskowitz MA (2003) Mechanisms, challenges and opportunities in stroke. Nat Rev Neurosci 4: 399–415. [DOI] [PubMed] [Google Scholar]
- 34. Viapiano MS, Matthews RT (2006) From barriers to bridges: chondroitin sulfate proteoglycans in neuropathology. Trends Mol Med 12: 488–496. [DOI] [PubMed] [Google Scholar]
- 35. Wang D, Fawcett J (2012) The perineuronal net and the control of CNS plasticity. Cell Tissue Res 349: 147–160. [DOI] [PubMed] [Google Scholar]
- 36. Hockfield S, Mckay RD (1983) A surface antigen expressed by a subset of neurons in the vertebrate central nervous system. Proc Natl Acad Sci U S A 80: 5758–5761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Celio MR, Spreafico R, De Biasi S, Vitellaro Zuccarello L (1998) Perineuronal nets: past and present. Trends Neurosci 21: 510–515. [DOI] [PubMed] [Google Scholar]
- 38. Morawski M, Brückner G, Arendt T, Matthews RT (2012) Aggrecan: beyond cartilage and into the brain. Int J Biochem Cell Biol 44: 690–693. [DOI] [PubMed] [Google Scholar]
- 39. Bandtlow CE, Zimmermann DR (2000) Proteoglycans in the developing brain: new conceptual insights for old proteins. Physiol Rev 80: 1267–1290. [DOI] [PubMed] [Google Scholar]
- 40. Yamaguchi Y (2000) Lecticans: organizers of the brain extracellular matrix. Cell Mol Life Sci 57: 276–289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Matthews RT, Kelly GM, Zerillo CA, Gray G, Tiemeyer M, et al. (2002) Aggrecan glycoforms contribute to the molecular heterogeneity of perineuronal nets. J Neurosci 22: 7536–7547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Guimarães A, Zaremba S, Hockfield S (1990) Molecular and morphological changes in the cat lateral geniculate nucleus and visual cortex induced by visual deprivation are revealed by monoclonal antibodies Cat-304 and Cat-301. J Neurosci 10: 3014–3024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. McRae PA, Rocco MM, Kelly G, Brumberg JC, Matthews RT (2007) Sensory deprivation alters aggrecan and perineuronal net expression in the mouse barrel cortex. J Neurosci 27: 5405–5413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Härtig W, Brauer K, Bigl V, Brückner G (1994) Chondroitin sulfate proteoglycan-immunoreactivity of lectin-labeled perineuronal nets around parvalbumin-containing neurons. Brain Res 635: 307–311. [DOI] [PubMed] [Google Scholar]
- 45. Hockfield S, Kalb RG, Zaremba S, Fryer H (1990) Expression of neural proteoglycans correlates with the acquisition of mature neuronal properties in the mammalian brain. Cold Spring Harb Symp Quant Biol 55: 505–514. [DOI] [PubMed] [Google Scholar]
- 46. Pizzorusso T, Medini P, Berardi N, Chierzi S, Fawcett JW, et al. (2002) Reactivation of ocular dominance plasticity in the adult visual cortex. Science 298: 1248–1251. [DOI] [PubMed] [Google Scholar]
- 47. Harris NG, Carmichael ST, Hovda DA, Sutton RL (2009) Traumatic brain injury results in disparate regions of chondroitin sulfate proteoglycan expression that are temporally limited. J Neurosci Res 87: 2937–2950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Harris NG, Mironova YA, Hovda DA, Sutton RL (2010) Pericontusion axon sprouting is spatially and temporally consistent with a growth-permissive environment after traumatic brain injury. J Neuropathol Exp Neurol 69: 139–154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Hobohm C, Günther A, Grosche J, Rossner S, Schneider D, et al. (2005) Decomposition and long-lasting downregulation of extracellular matrix in perineuronal nets induced by focal cerebral ischemia in rats. J Neurosci Res 80: 539–548. [DOI] [PubMed] [Google Scholar]
- 50. Carmichael ST, Archibeque I, Luke L, Nolan T, Momiy J, et al. (2005) Growth-associated gene expression after stroke: evidence for a growth-promoting region in peri-infarct cortex. Exp Neurol 193: 291–311. [DOI] [PubMed] [Google Scholar]
- 51. Karetko Sysa M, Skangiel Kramska J, Nowicka D (2011) Disturbance of perineuronal nets in the perilesional area after photothrombosis is not associated with neuronal death. Exp Neurol 231: 113–126. [DOI] [PubMed] [Google Scholar]
- 52. Ruscher K, Shamloo M, Rickhag M, Ladunga I, Soriano L, et al. (2011) The sigma-1 receptor enhances brain plasticity and functional recovery after experimental stroke. Brain 134: 732–746. [DOI] [PubMed] [Google Scholar]
- 53. Encarnacion A, Horie N, Keren Gill H, Bliss TM, Steinberg GK, et al. (2011) Long-term behavioral assessment of function in an experimental model for ischemic stroke. J Neurosci Methods 196: 247–257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Lenzlinger PM, Saatman KE, Hoover RC, Cheney JA, Bareyre FM, et al. (2004) Inhibition of vascular endothelial growth factor receptor (VEGFR) signaling by BSF476921 attenuates regional cerebral edema following traumatic brain injury in rats. Restor Neurol Neurosci 22: 73–79. [PubMed] [Google Scholar]
- 55. De Ryck M, Van Reempts J, Duytschaever H, Van Deuren B, Clincke G (1992) Neocortical localization of tactile/proprioceptive limb placing reactions in the rat. Brain Res 573: 44–60. [DOI] [PubMed] [Google Scholar]
- 56. Hua Y, Schallert T, Keep RF, Wu J, Hoff JT, et al. (2002) Behavioral tests after intracerebral hemorrhage in the rat. Stroke 33: 2478–2484. [DOI] [PubMed] [Google Scholar]
- 57. Jones TA, Schallert T (1992) Subcortical deterioration after cortical damage: effects of diazepam and relation to recovery of function. Behav Brain Res 51: 1–13. [DOI] [PubMed] [Google Scholar]
- 58. Ballermann M, Metz GA, McKenna JE, Klassen F, Whishaw IQ (2001) The pasta matrix reaching task: a simple test for measuring skilled reaching distance, direction, and dexterity in rats. J Neurosci Methods 106: 39–45. [DOI] [PubMed] [Google Scholar]
- 59. Swanson RA, Morton MT, Tsao Wu G, Savalos RA, Davidson C, et al. (1990) A semiautomated method for measuring brain infarct volume. J Cereb Blood Flow Metab 10: 290–293. [DOI] [PubMed] [Google Scholar]
- 60. Giamanco KA, Morawski M, Matthews RT (2010) Perineuronal net formation and structure in aggrecan knockout mice. Neuroscience 170: 1314–1327. [DOI] [PubMed] [Google Scholar]
- 61. Lendvai D, Morawski M, Négyessy L, Gáti G, Jäger C, et al. (2013) Neurochemical mapping of the human hippocampus reveals perisynaptic matrix around functional synapses in Alzheimer's disease. Acta Neuropathol 125: 215–229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Dino MR, Harroch S, Hockfield S, Matthews RT (2006) Monoclonal antibody Cat-315 detects a glycoform of receptor protein tyrosine phosphatase beta/phosphacan early in CNS development that localizes to extrasynaptic sites prior to synapse formation. Neuroscience 142: 1055–1069. [DOI] [PubMed] [Google Scholar]
- 63. Sigler A, Goroshkov A, Murphy TH (2008) Hardware and methodology for targeting single brain arterioles for photothrombotic stroke on an upright microscope. J Neurosci Methods 170: 35–44. [DOI] [PubMed] [Google Scholar]
- 64. Moon SK, Alaverdashvili M, Cross AR, Whishaw IQ (2009) Both compensation and recovery of skilled reaching following small photothrombotic stroke to motor cortex in the rat. Exp Neurol 218: 145–153. [DOI] [PubMed] [Google Scholar]
- 65. Starkey ML, Bleul C, Zörner B, Lindau NT, Mueggler T, et al. (2012) Back seat driving: hindlimb corticospinal neurons assume forelimb control following ischaemic stroke. Brain 135: 3265–3281. [DOI] [PubMed] [Google Scholar]
- 66. Diederich K, Quennet V, Bauer H, Müller HD, Wersching H, et al. (2012) Successful regeneration after experimental stroke by granulocyte-colony stimulating factor is not further enhanced by constraint-induced movement therapy either in concurrent or in sequential combination therapy. Stroke 43: 185–192. [DOI] [PubMed] [Google Scholar]
- 67. Jin Z, Wu J, Oh SY, Kim KW, Shin BS (2010) The effect of stress on stroke recovery in a photothrombotic stroke animal model. Brain Res 1363: 191–197. [DOI] [PubMed] [Google Scholar]
- 68. Minnerup J, Seeger FH, Kuhnert K, Diederich K, Schilling M, et al. (2010) Intracarotid administration of human bone marrow mononuclear cells in rat photothrombotic ischemia. Exp Transl Stroke Med 2: 3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Jablonka JA, Burnat K, Witte OW, Kossut M (2010) Remapping of the somatosensory cortex after a photothrombotic stroke: dynamics of the compensatory reorganization. Neuroscience 165: 90–100. [DOI] [PubMed] [Google Scholar]
- 70. Vandeputte C, Taymans JM, Casteels C, Coun F, Ni Y, et al. (2010) Automated quantitative gait analysis in animal models of movement disorders. BMC Neurosci 11: 92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Knieling M, Metz GA, Antonow Schlorke I, Witte OW (2009) Enriched environment promotes efficiency of compensatory movements after cerebral ischemia in rats. Neuroscience 163: 759–769. [DOI] [PubMed] [Google Scholar]
- 72. Müller HD, Hanumanthiah KM, Diederich K, Schwab S, Schäbitz WR, et al. (2008) Brain-derived neurotrophic factor but not forced arm use improves long-term outcome after photothrombotic stroke and transiently upregulates binding densities of excitatory glutamate receptors in the rat brain. Stroke 39: 1012–1021. [DOI] [PubMed] [Google Scholar]
- 73. Rogers DC, Hunter AJ (1997) Photothrombotic lesions of the rat cortex impair acquisition of the water maze. Pharmacol Biochem Behav 56: 747–754. [DOI] [PubMed] [Google Scholar]
- 74. Schäbitz WR, Berger C, Kollmar R, Seitz M, Tanay E, et al. (2004) Effect of brain-derived neurotrophic factor treatment and forced arm use on functional motor recovery after small cortical ischemia. Stroke 35: 992–997. [DOI] [PubMed] [Google Scholar]
- 75. Wood NI, Sopesen BV, Roberts JC, Pambakian P, Rothaul AL, et al. (1996) Motor dysfunction in a photothrombotic focal ischaemia model. Behav Brain Res 78: 113–120. [DOI] [PubMed] [Google Scholar]
- 76. Donoghue JP, Sanes JN (1994) Motor areas of the cerebral cortex. J Clin Neurophysiol 11: 382–396. [PubMed] [Google Scholar]
- 77. Hall RD, Lindholm EP (1974) Organization of motor and somatosensory neocortex in the albino rat. Brain Res 66: 23–38. [Google Scholar]
- 78. De Ryck M, Van Reempts J, Borgers M, Wauquier A, Janssen PA (1989) Photochemical stroke model: flunarizine prevents sensorimotor deficits after neocortical infarcts in rats. Stroke 20: 1383–1390. [DOI] [PubMed] [Google Scholar]
- 79. Shanina EV, Schallert T, Witte OW, Redecker C (2006) Behavioral recovery from unilateral photothrombotic infarcts of the forelimb sensorimotor cortex in rats: role of the contralateral cortex. Neuroscience 139: 1495–1506. [DOI] [PubMed] [Google Scholar]
- 80. Wang Y, Bontempi B, Hong SM, Mehta K, Weinstein PR, et al. (2008) A comprehensive analysis of gait impairment after experimental stroke and the therapeutic effect of environmental enrichment in rats. J Cereb Blood Flow Metab 28: 1936–1950. [DOI] [PubMed] [Google Scholar]
- 81. Dahlqvist P, Rönnbäck A, Risedal A, Nergårdh R, Johansson IM, et al. (2003) Effects of postischemic environment on transcription factor and serotonin receptor expression after permanent focal cortical ischemia in rats. Neuroscience 119: 643–652. [DOI] [PubMed] [Google Scholar]
- 82. Overman JJ, Carmichael ST (2014) Plasticity in the injured brain: more than molecules matter. Neuroscientist 20: 15–28. [DOI] [PubMed] [Google Scholar]
- 83. Sale A, Maya Vetencourt JF, Medini P, Cenni MC, Baroncelli L, et al. (2007) Environmental enrichment in adulthood promotes amblyopia recovery through a reduction of intracortical inhibition. Nat Neurosci 10: 679–681. [DOI] [PubMed] [Google Scholar]
- 84. Foscarin S, Ponchione D, Pajaj E, Leto K, Gawlak M, et al. (2011) Experience-dependent plasticity and modulation of growth regulatory molecules at central synapses. PLoS One 6: e16666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85. Katsman D, Zheng J, Spinelli K, Carmichael ST (2003) Tissue microenvironments within functional cortical subdivisions adjacent to focal stroke. J Cereb Blood Flow Metab 23: 997–1009. [DOI] [PubMed] [Google Scholar]
- 86. Carmichael ST (2006) Cellular and molecular mechanisms of neural repair after stroke: making waves. Ann Neurol 59: 735–742. [DOI] [PubMed] [Google Scholar]
- 87. Hill JJ, Jin K, Mao XO, Xie L, Greenberg DA (2012) Intracerebral chondroitinase ABC and heparan sulfate proteoglycan glypican improve outcome from chronic stroke in rats. Proc Natl Acad Sci U S A 109: 9155–9160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88. Soleman S, Yip PK, Duricki DA, Moon LD (2012) Delayed treatment with chondroitinase ABC promotes sensorimotor recovery and plasticity after stroke in aged rats. Brain 135: 1210–1223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89. Gherardini L, Gennaro M, Pizzorusso T (2013) Perilesional treatment with chondroitinase ABC and motor training promote functional recovery after stroke in rats. Cereb Cortex. [DOI] [PubMed] [Google Scholar]
- 90. Lander C, Kind P, Maleski M, Hockfield S (1997) A family of activity-dependent neuronal cell-surface chondroitin sulfate proteoglycans in cat visual cortex. J Neurosci 17: 1928–1939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91. Freund TF, Katona I (2007) Perisomatic inhibition. Neuron 56: 33–42. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Animal data. Parameters of the rats subjected to PT with subsequent housing in STD or EE.
(TIF)
Explanation of the Catwalk parameters. (a.u.: arbitrary units).
(TIF)
Gait parameters affected by stroke. The pre-stroke values and the values 2 days after stroke are presented (n = 14) as well as the percentage of variation between the time points, the p-value of the Student's t-test (pre vs. 2d), the Pearson's correlation coefficient with the speed and weight with the respective associated p-values. (RF: right front, RH: right hind, LF: left front, LH: left hind, a.u.: arbitrary unit).
(TIF)
Asymmetry in parameters of gait induced by stroke. Gait parameters where differences were seen between paretic and non-paretic side at 2 days after stroke. The pre-stroke values and the values 2 days after stroke are presented (n = 14) as well as the percentage of variation between the two side, the respective p-value of the Student's t-test (L vs. R) for both time and the p-value of the Student's t-test on the difference (Right-Left) before and 2d after stroke. (F: front, H: hind, L: left, R: right).
(TIF)
Recovery in parameters of the gait affected by stroke. Gait parameter values affected by stroke (general) and the difference between paretic and non-paretic side induced by stroke (asymmetry) at 2 and 5 weeks in rats housed in either STD or EE. (RF: right front, RH: right hind, LF: left front, LH: left hind, F: front, H: hind).
(TIF)