

Summary
O-linked glycans on plasma membrane proteins are altered in cancer cells leading to changes in cell adhesive properties and contributing to metastasis. Mechanisms of how these carbohydrates alter tumor spread remain vague. In this issue of Cancer Discovery, Murugaesu and colleagues, using an in vivo functional RNAi metastasis screen, identified sialyltransferase ST6GalNAc2 as a novel metastasis suppressor gene. Aggressive estrogen receptor negative breast cancers with reduced ST6GalNAc2 expression alter O-linked glycans on tumor cell surface increasing soluble lectin galectin-3 binding and tumor cell clustering at metastatic sites.
Metastasis remains the major cause of mortality of solid tumors. There is a critical need for novel therapies but also biomarkers indicative of metastatic disease to help stratify patients towards targeted therapy.
Cancer cell dissemination and establishment at a secondary site is the result of a complex multistep process. Various reports have linked alterations in cell surface glycosylation with increased malignant properties in a number of cancers including breast, prostate and colorectal tumors (1). Specifically, cell surface glycans function in cell-cell and cell-matrix adhesions playing a role in cancer tumor growth and spread. Aberrant cell surface glycosylation is often due to altered expression or activity of glycosyltransferases leading to expression of tumor-associated carbohydrate antigens (TACAs). Cancer cells typically express truncated sialylated or unsialylated (Core 1) O-glycans compared to normal cells that express longer (Core 2) O-glycans chains (1). A number of studies have shown that the expression of the sialyltransferase ST6GalNAc1 (alpha-N-acetylgalactosaminide alpha-2,6-sialytransferase1), responsible for catalyzing the addition of sialic acid on core 1 O-glycans, is elevated in breast cancers and regulated by oncogenes such as Ras (1). Recent studies have shown that the sialyltransferase ST6GalNAc5, which transfers sialic acid residues on gangliosides, is critical for breast cancer metastasis to the brain as its expression allows breast cancer cells to adhere to brain endothelium and pass trough blood-brain barrier (2). However, few studies have addressed mechanistically how changes in sialylation can alter tumor cell behavior and metastasis in vivo.
Cancer-specific O-glycan antigens often recruit carbohydrate-binding proteins, or lectins, that play an important role in metastasis. In particular, galectins are sugar-binding proteins that function in the immune response in normal cells and play a diverse role in cancer cells including survival, invasion and metastasis. Cytoplasmic galectins can have intracellular functions to modulate cell survival and splicing but can also be secreted. Once outside the cell, galectins can mediate communication between cells through binding to cell surface glycoconjugates. Galectins share a highly conserved carbohydrate recognition domains (CRD) and a flexible N-terminal domain and can form monomers in solution. However, in the presence of specific carbohydrate binding, galectin-3 can polymerize and form pentamers through its N-terminal domain and multimerize on surface of cells (3). Galectin-3 in tumor cells can trigger cell homotypic aggregation and heterotypic adhesion to endothelial cell and signaling that protects from anoikis and increase metastasis (4). Indeed, levels of circulating galectin-3 are elevated in patients with metastatic disease and associated with poor patient outcome (4). The exact mechanism by which galectin-3 binding to glycoproteins contributes to metastasis is not clear.
In this issue of Cancer Discovery, Murugaesu and colleagues (5) provide insight into understanding of how O-linked glycans can regulate galectin-3 binding, tumor cell retention and metastatic burden in estrogen receptor (ER) negative breast cancers. The authors use an in vivo enrichment screening of shRNA library to identify genes that restrain metastasis. This screen identified the sialyltransferase ST6GalNAc2 as a novel metastasis suppressor. Depleting ST6GalNAc2 alone enhances metastasis in 4T1 breast cancer metastasis model and overexpression of this gene in metastatic human breast cancer cells impairs tumor spread. In addition, the authors find that low ST6GalNAc2 expression correlates with ER-negative breast cancers and high ST6GalNAc2 levels in this ER-negative cohort is associated with improved overall survival. The authors go on to show that depletion of ST6GalNAc2 changes abundance of O-glycans on tumor cells that promotes binding of galectin-3 and increasing tumor cell retention in lungs and increasing metastatic frequency. Additionally, the authors demonstrate that levels of ST6GalNAc2 expression determine responsiveness to galectin-3 inhibition.
Although an in vivo RNAi screen to identify metastasis suppressors has been described previously (6), Murugaesu and colleagues (5) go one step further performing next generation sequencing to pinpoint ST6GalNAc2 as a key player in modulation of late stage metastasis. Combination of a pooled shRNA library screen with next generation sequencing allows for rapid identification of changes in representation of different shRNAs in metastatic tumor populations. Large-scale screening using the 4T1 mouse breast cancer cells in experimental model of metastasis (i.e. tail-vein injection) identified 4 genes contributing to tumor metastasis using in vivo bioluminescence imaging as a readout. The enzyme ST6GalNAc2, a glycosyltransferase responsible for the addition of sialic acid on O-glycans on the cell surface, was one of the most enriched genes identified. Interestingly, stable ST6GalNAc2 knockdown in 4T1 cells resulted in increased metastatic tumor burden without affecting primary tumor growth. To further establish role of ST6GalNAc2 as metastasis suppressor, they performed in vivo gain of function experiments using triple-negative MDA-MB-231 cells that normally contain low expression levels of ST6GalNAc2 and demonstrate that overexpression of this glycosyltransferase can indeed function as a suppressor of breast cancer metastasis. In expanding their findings to human clinical data, the authors demonstrate that low levels of ST6GalNAc2 expression are associated with the ER-negative breast cancer subtype and that within this patient cohort, higher expression of ST6GalNAc2 significantly correlates with improved survival outcome. Together, the combination of in vivo RNAi genetic screening with functional target validation unveiled a novel metastatic tumor suppressor that may be contributing to the progression of aggressive ER-negative breast cancers.
Increased attention has been focused on tumor-associated sialylated O-glycans, and the enzymes responsible for these modifications, as potential biomarkers of breast cancer (1). Alterations in sialyltransferases have been shown to alter O-glycan modifications on the cell surface, thereby affecting cell-cell and cell-substrate interactions, particularly lectin binding. Here the authors demonstrate a mechanism whereby the reduction in ST6GalNAc2 expression alters the tumor cell surface O-glycan profile thus promoting galectin-3 binding and metastasis. Specifically, the authors first performed mass spectrometric analysis of cell surface O-glycans in response to ST6GalNAc2 silencing and observed variations in O-glycan abundance. Among these alterations, they were able to detect increased levels of the unmodified core 1 O-glycan or core 1 T-antigen (Galβ1-3GalNac), while decreasing the α2-3,α2-6 disialyl core 1 O-glycan. Although unmodified Tn antigen is a well known TACA (1), they did not see changes in this carbohydrate in ST6GalNAc2 knockdown cancer cells. Since galectin-3 is known to bind core 1 glycan present on tumor cell surface glycoproteins, the authors investigated changes in galectin-3 binding to cells with reduced ST6GalNAc expression. Reduction in ST6GalNAc expression enhanced galectin-3 binding on cancer cell surface in vitro and promoted tumor cell aggregation and lung retention in vivo. Mechanistically, they show this effect requires galectin-3 as co-inhibition of ST6GalNAc2 and galectin-3 via RNAi in 4T1 cells breast cancer cells impaired tumor cell seeding in the lungs. Because galectin-3 has been shown to promote tumor-endothelial cell adhesion to increase tumor retention, the authors next investigated whether ST6GalNAc downregulation increased breast cancer cell adhesion to endothelial cells. In vitro experiments showed that reducing expression of ST6GalNAc results in increased adhesion of tumor cells to human umbilical vein endothelial cells and that this adhesion is attenuated with the use of galectin-3 inhibitors or RNAi in both static and dynamic flow conditions.
Importantly, galectin-3 RNAi alone had no effect on metastasis of 4T1 cells. In MDA-MB-231 cells, which contain low levels of ST6GalNAc2, RNAi against galectin-3 alone inhibited tumor seeding in lungs but had no impact on MDA-MB-231 cells with stable ectopic ST6GalNAc2 expression. Thus, suggesting that galectin-3 effects on metastatic colonization may be dependent on ST6GalNAc2 activity. To explore this idea further, Murugaesu and colleagues (5) treated mice injected with 4T1 cells containing RNAi against ST6GalNAc2 with the galectin-3 inhibitor, GCS-100 (7). Breast cancer cells expressing RNAi targeting ST6GalNAc treated with GCS-100 demonstrated an inhibition of metastasis. However, GCS-100 treatment had no effect on metastatic colonization of control RNAi 4T1 cells, suggesting that galectin-3 inhibition is effective only if tumor cells have low levels of ST6GalNAc2 levels.
Various reports implicate galectin-3 as a promoter of tumor progression and clinical trials are currently underway utilizing GCS-100 as a treatment for multiple myeloma with low, tolerated side effects (7). However, Murugaesu and colleagues (5) suggests that galectin-3 itself may not be a good prognostic factor using gene expression profiling since they found that galectin-3 had no significant association with ER status in human breast cancers and no association with survival outcome within ER-negative cohort. Importantly, this study provides strong evidence that ST6GalNAc2 may be used as a biomarker in ER-negative breast cancer to identify patients who may benefit from treatment with galectin-3 inhibitors.
Changes in glycosylation are known to be associated with malignancy, with increases in surface sialic acids being one of the most common (1). However, this study is the first to show that loss of a sialyltransferase can result in reduced branching and increase in Core 1 glycans. Changes in this terminal glycan moiety create a functional advantage to tumor cells by altering cellular adhesion with neighboring cells (3). It will be interesting to determine whether other sialyltransferase found to be overexpressed in cancer, notably ST6GalNAc1, can also alter galectin-3 binding. These modifications in surface O-glycan expression on tumor cells can also confer an immune escape mechanism, interfering with antigen presentation to T-cells (8). Specifically, altered glycans have the ability to interact with T-cell receptors and NK cells eliciting an inhibitory response (3). Additionally, galectin binding has been shown to promote integrin clustering to activate T-cell apoptosis and stimulate tumor cell invasion, angiogenesis and proliferation (9). Future studies should evaluate the role decreased ST6GalNAc2 expression plays as an immune escape mechanism in ER-negative breast cancer cells. Does ST6GlcNAc2 reduction-induced surface O-glycan expression prevent tumor antigen-presentation and immune attack in an immune-competent model?
In summary, Murugaesu and colleagues (5) present a model (Figure 1) in which ER-negative breast cancer cells that have lost ST6GalNAc2 expression, contain increased unmodified core 1 O-glycan (core1/T antigen) while decreasing disialyl core O-glycan levels, promoting galectin-3 binding to tumor cells, increase tumor cell aggregation and facilitating tumor cell retention in lung vasculature. This increase in tumor cell seeding in the lungs allows for increased extravasation and increased frequency of metastatic events. Currently, clinical trials employing galectin-3 inhibitors (GCS-100) are underway for the treatment of lymphomas and leukemias (clinicaltrials.gov). GCS-100 has revealed low toxicity profiles and efficacy in tumor patients both as a single agent and as a combination therapy in preclinical and clinical trials, with ability to restore T-cell immune response (10). Thus, ST6GalNAc2 expression may help identify a population of breast cancer patients that could benefit from galectin inhibitors.
Figure 1. Breast cancer alterations in ST6GalNAc2 levels contribute to changes in O-glycans, galectin-3 binding and cellular aggregation at metastasis site.
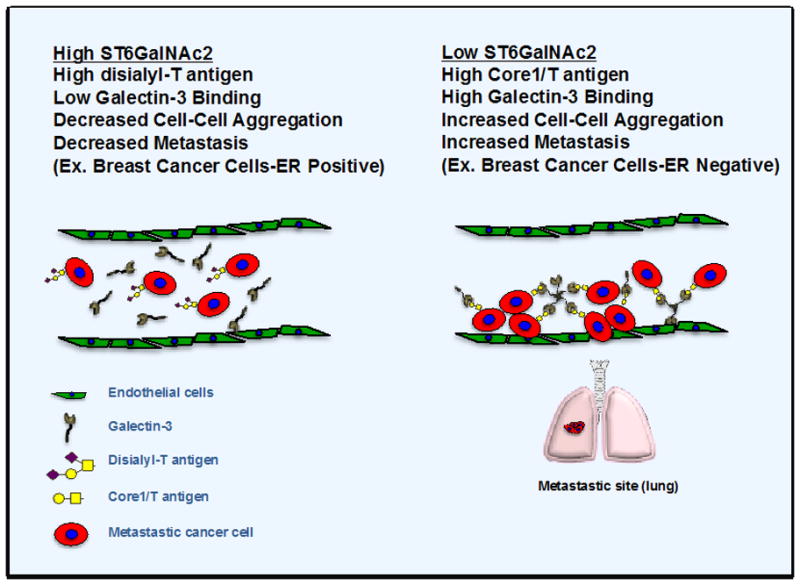
ER-positive breast cancer cells contain overall higher levels of ST6GalNAc2 expression and therefore modified O-glycans on the cell surface, with decreased galectin-3 binding and metastasis. In contrast, ER-negative breast cancer cells with low ST6GalNAc expression display increased unmodified core 1 O-glycans (T antigen) on the cell surface and galectin-3 binding resulting in higher endothelial cell binding and metastasis.
Footnotes
The authors disclose no potential conflicts of interest.
References
- 1.Cazet A, Julien S, Bobowski M, Burchell J, Delannoy P. Tumour-associated carbohydrate antigens in breast cancer. Breast Cancer Res. 2010;12:204. doi: 10.1186/bcr2577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Bos PD, Zhang XH, Nadal C, Shu W, Gomis RR, Nguyen DX, et al. Genes that mediate breast cancer metastasis to the brain. Nature. 2009;459:1005–1009. doi: 10.1038/nature08021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Rabinovich GA, Toscano MA. Turning ‘sweet’ on immunity: galectin-glycan interactions in immune tolerance and inflammation. Nat Rev Immunol. 2009;9:338–352. doi: 10.1038/nri2536. [DOI] [PubMed] [Google Scholar]
- 4.Newlaczyl AU, Yu LG. Galectin-3--a jack-of-all-trades in cancer. Cancer Lett. 2011;313:123–128. doi: 10.1016/j.canlet.2011.09.003. [DOI] [PubMed] [Google Scholar]
- 5.Murugaesu N, Iravani M, van Weverwijk A, Ivetic A, Johnson DA, Antonopoulos A, et al. An in vivo functional screen identifies ST6GalNAc2 sialyltransferase as a breast cancer metastasis suppressor gene. Cancer Discov. 2014 doi: 10.1158/2159-8290.CD-13-0287. [DOI] [PubMed] [Google Scholar]
- 6.Gumireddy K, Li A, Gimotty PA, Klein-Szanto AJ, Showe LC, Katsaros D, et al. KLF17 is a negative regulator of epithelial-mesenchymal transition and metastasis in breast cancer. Nat Cell Biol. 2009;11:1297–1304. doi: 10.1038/ncb1974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Chauhan D, Li G, Podar K, Hideshima T, Neri P, He D, et al. A novel carbohydrate-based therapeutic GCS-100 overcomes bortezomib resistance and enhances dexamethasone-induced apoptosis in multiple myeloma cells. Cancer Res. 2005;65:8350–8358. doi: 10.1158/0008-5472.CAN-05-0163. [DOI] [PubMed] [Google Scholar]
- 8.Tsuboi S, Sutoh M, Hatakeyama S, Hiraoka N, Habuchi T, Horikawa Y, et al. A novel strategy for evasion of NK cell immunity by tumours expressing core2 O-glycans. EMBO J. 2011;30:3173–3185. doi: 10.1038/emboj.2011.215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Muller I, Jenner J, Handgretinger R, Riberdy J, Kerst G. Glycosylation and lectins-examples of immunesurveillance and immune evasion. Histol Histopathol. 2004;19:527–533. doi: 10.14670/HH-19.527. [DOI] [PubMed] [Google Scholar]
- 10.Demotte N, Wieers G, Van Der Smissen P, Moser M, Schmidt C, Thielemans K, et al. A galectin-3 ligand corrects the impaired function of human CD4 and CD8 tumor-infiltrating lymphocytes and favors tumor rejection in mice. Cancer Res. 2010;70:7476–7488. doi: 10.1158/0008-5472.CAN-10-0761. [DOI] [PubMed] [Google Scholar]