

Abstract
The clinical relevance of the anti-inflammatory properties of beta-2 agonists remains contentious possibly due to differences in their molecular structures and agonist activities. The current study has compared the effects of 3 different categories of β2-agonists, namely, salbutamol (short-acting), formoterol (long-acting) and indacaterol (ultra-long-acting), at concentrations of 1–1000 nM, with human blood neutrophils in vitro. Neutrophils were activated with either N-formyl-L-methionyl-L-leucyl-L-phenylalanine (fMLP, 1 µM) or platelet-activating factor (PAF, 200 nM) in the absence and presence of the β2-agonists followed by measurement of the generation of reactive oxygen species and leukotriene B4, release of elastase, and expression of the β2-integrin, CR3, using a combination of chemiluminescence, ELISA, colorimetric, and flow cytometric procedures respectively. These were correlated with alterations in the concentrations of intracellular cyclic-AMP and cytosolic Ca2+. At the concentrations tested, formoterol and indacaterol caused equivalent, significant (P < 0.05 at 1–10 nM) dose-related inhibition of all of the pro-inflammatory activities tested, while salbutamol was much less effective (P < 0.05 at 100 nM and higher). Suppression of neutrophil reactivity was accompanied by elevations in intracellular cAMP and accelerated clearance of Ca2+ from the cytosol of activated neutrophils. These findings demonstrate that β2-agonists vary with respect to their suppressive effects on activated neutrophils.
1. Introduction
Beta-2-adrenoreceptor (β2ARs) agonists (β2-agonists) are used primarily as bronchodilators in the therapy of obstructive airways disorders, especially bronchial asthma and chronic obstructive pulmonary disease (COPD) [1, 2]. These agents also possess secondary anti-inflammatory properties, affecting cells of both the innate and adaptive immune systems and their inflammatory mediators in vitro, as well as in some animal models of experimental airway inflammation [3–5]. As with bronchodilatory activity, anti-inflammatory activity is achieved via sequential β2AR-mediated activation of adenylyl cyclase, elevated intracellular concentrations of 3′-5′-cyclic adenosine monophosphate (cAMP), and activation of cAMP-dependent protein kinase (PKA) [3–5]. While the therapeutic efficacy of the bronchodilatory actions of β2-agonists is undisputed, the clinical relevance of the anti-inflammatory activities of these agents remains uncertain [3, 5]. This contention is underscored by data from recently reported large, well-controlled clinical trials demonstrating lack of efficacy of aerosolized or intravenous salbutamol in the therapy of acute lung injury [6, 7]. Several mechanisms may underpin the apparent lack of anti-inflammatory potency of β2-agonists in the clinical setting. These include (i) lower numbers of β2-ARs on immune and inflammatory cells relative to airway smooth muscle cells, rendering them more susceptible to receptor desensitization [8], and (ii) possible variability in anti-inflammatory potency between different types of β2-agonists resulting from differences in molecular structure.
In the current study, we have compared the anti-inflammatory activities of two commonly used representatives of the short- and long-acting categories of β2-agonists (SABA, LABA), namely, salbutamol and formoterol, respectively, with those of the recently introduced ultralong-acting (ultra LABA), indacaterol, used for maintenance therapy of airflow limitation in patients with COPD [9, 10]. The study is focused on the effects of these agents on the proinflammatory activities of the human neutrophil, a cell type which not only expresses relatively high numbers of β2ARs in comparison with other types of immune and inflammatory cells [5, 11], but which is also intimately involved in the immunopathogenesis of COPD [12, 13]. In addition, while several recent studies have addressed the anti-inflammatory interactions of indacaterol with human lung fibroblasts [14] and mast cells [15], the effects of this agent on human neutrophils have not been described.
2. Materials and Methods
2.1. Chemicals and Reagents
Formoterol fumarate dehydrate and salbutamol hydrochloride were purchased from Sigma-Aldrich (Pty) Ltd. (Johannesburg, South Africa), while indacaterol maleate was provided by Novartis International AG, Basel, Switzerland. All 3 agents were dissolved to a stock concentration of 10 mM in dimethylsulfoxide (DMSO) and used at final concentrations of 1, 10, and 100 nM in most of the assays described below and at 1–1000 nM in others. The local concentration of formoterol following inhalation of 24 μg of this agent has been estimated to be approximately 11 nM [16]. The final concentration of DMSO in each assay was 0.1% and appropriate solvent controls were included with each experimental system. Unless indicated, all other chemicals and reagents were purchased from Sigma-Aldrich.
Neutrophils were activated with the chemoattractant, N-formyl-L-methionyl-L-leucyl-L-phenylalanine (fMLP, 1 μM final) either alone or in combination with cytochalasin B (CB, 1 μM final) in assays of oxygen consumption and degranulation. An additional chemoattractant, platelet-activating factor (PAF, 200 nM final) was used in a more limited series of experiments.
2.2. Preparation of Neutrophils
The study was approved by the Faculty of Health Sciences Research Ethics Committee of the University of Pretoria, Pretoria, South Africa, and prior informed consent was obtained from all blood donors. Neutrophils were isolated from heparinized venous blood (5 units of preservative-free heparin per mL of blood) from nonsmoking healthy adult volunteers. Each subject completed a detailed health questionnaire and underwent a health check (including measurement of blood pressure) by an experienced, qualified nursing sister prior to venepuncture. Neutrophils were separated from mononuclear leucocytes by centrifugation on Histopaque-1077 (Sigma Diagnostics, St. Louis, MO, USA) cushions at 400 ×g for 25 min at room temperature. The resultant pellets were suspended in PBS (0.15 M, pH 7.4) and sedimented with 3% gelatin to remove most of the erythrocytes. Following centrifugation (280 ×g at 10°C for 10 min), residual erythrocytes were removed by selective lysis with 0.83% ammonium chloride at 4°C for 10 min. The neutrophils, which were routinely of high purity (>90%) and viability (>95%), were resuspended to 1 × 107 cells mL−1 in PBS and held on ice until used.
2.3. Generation of Reactive Oxygen Species (ROS)
This was measured using a luminol (5-amino-2,3-dihydro-1,4-phthalazinedione)-enhanced chemiluminescence (LECL) procedure. Briefly, neutrophils (106 cells) were preincubated for 10 min at 37°C in 900 μL Hanks' balanced salt solution (HBSS, indicator-free; Highveld Biological (Pty) Ltd., Johannesburg, South Africa) containing luminol at a final concentration of 0.1 mM. Following preincubation, the test β2-agonists (1–1000 nM) or solvent control were added to the cells, followed after 20 s by fMLP (1 μM), and LECL responses recorded using a Lumac Biocounter (model 2010; Lumac Systems Inc., Titusville, FL, USA). The final volume in each tube was 1 mL, and the results, which are expressed in relative light units (rlu), are the peak values for fMLP-activated systems that were reached 40–50 s after addition of stimulant.
2.4. Oxygen Consumption
This was measured using a 3-channel oxygen electrode (Model DW1, Hansatech Ltd., King's Lynn, Norfolk, UK). Neutrophils (2 × 106) were incubated for 10 min at 37°C in HBSS after which the test β2-agonists, at a fixed, final concentration of 100 nM, were added to the cells, followed 20 s later by fMLP/CB (1 μM : 1 μM). PO2 was monitored over an 8 min period and O2 consumption calculated over the period when utilization was linear (1-2 min), which was approximately 1 min, and the results are expressed as nanomoles (nmols) O2 consumed min−1 by 2 × 106 cells.
2.5. Elastase
Neutrophil degranulation was measured according to the extent of release of the primary granule enzyme, elastase. Neutrophils were incubated at a concentration of 2 × 106 mL−1 in HBSS for 10 min at 37°C followed by sequential addition of the test β2-agonists (1–1000 nM) and, 20 s later, by fMLP/CB (1 μM : 1 μM), and the tubes then incubated for 10 min at 37°C. The tubes were then transferred to an ice bath, followed by centrifugation at 400 ×g for 5 min to pellet the cells, and the supernatants were decanted and assayed for elastase using a micromodification of a standard colorimetric procedure. Briefly, 125 μL of supernatant was added to the elastase substrate, N-succinyl-L-analyl-L-analyl-L-alanine-p-nitroanilide (3 mmol·L−1 in DMSO) in 0.05 M Tris-HCl (pH 8.0), and elastase activity was monitored spectrophotometrically at a wavelength of 405 nm. The results are expressed as milliunits enzyme per 2 × 106 cells.
2.6. CR3 Expression
Expression of CR3 on resting and fMLP/CB (1 μM : 1 μM)-activated neutrophils in the absence and presence of the test β2-agonists (added as above) was measured flow cytometrically. Neutrophils (1 × 106 mL−1 final) were incubated in HBSS for 10 min at 37°C followed by addition of the β2-agonists (100 nM, fixed, final concentration) and activator. The tubes were incubated for 1 min and the reactions stopped by dilution of the cells in ice-cold medium. Phycoerythrin-labelled monoclonal antibody to CD11b (Beckman Coulter, Miami, FL, USA) was added in a 10 μL volume to 1 mL of cell suspension and CR3 expression analyzed using a Gallios 10C Flow Cytometer (Beckman Coulter). The results were expressed as mean fluorescence intensity.
2.7. Measurement of Leukotriene B4 (LTB4) and cAMP
Competitive binding immunoassay procedures (Correlate-EIA; Assay Designs Inc., Ann Arbor, MI, USA) were used to measure LTB4 in the supernatants of fMLP (1 μM) or PAF (200 nM)-activated neutrophils, while cAMP was assayed in the extracts of unstimulated cells in the absence and presence of the test β2-agonists. In the case of LTB4, neutrophils (2 × 106 mL−1) were preincubated for 10 min at 37°C after which the test β2-agonists (10 and 100 nM, final) were added followed 20 s later by fMLP or PAF. Incubation was terminated 5 min later by the addition of an equal volume of ice-cold HBSS to the tubes, which were then held in an ice bath prior to pelleting the cells by centrifugation. The cell-free supernatants were then diluted (1 : 8) and assayed for LTB4, with the results expressed as pg LTB4 per 107 cells.
In the case of cAMP, neutrophils (2 × 106 mL−1) were preincubated for 10 min at 37°C after which the test β2-agonists (10 and 100 nM) were added and the reactions terminated 20 s later by the addition of an equal volume of ice-cold HBSS to the tubes. Following centrifugation, the supernatants were discarded and cAMP extracted from the cell pellets by addition of 0.1 M HCl for 30 min followed by centrifugation to remove cell debris and the supernatants decanted and assayed for cAMP. These results are expressed as pmol cAMP per 107 cells.
2.8. Spectrofluorimetric Measurement of Cytosolic Ca2+
Fura-2 acetoxymethyl ester (fura-2AM) was used as the fluorescent, Ca2+-sensitive indicator for these experiments [17]. Neutrophils (1 × 107 mL−1) were incubated with fura-2AM (2 μM) for 30 min at 37°C in PBS, washed and resuspended in HBSS. The fura-2-loaded cells (2 × 106 mL−1) were then preincubated for 10 min at 37°C after which they were transferred to disposable reaction cuvettes which were maintained at 37°C in a Hitachi 650 10 S fluorescence spectrophotometer with excitation and emission wavelengths set at 340 and 500 nm, respectively. After a stable baseline was obtained (±1 min), test agents (1–100 nM) or solvent control were added to the relevant systems followed 20 s later by fMLP (1 μM) or PAF (200 nM) and alterations in fluorescence intensity were monitored over a 5–10 min time course. Alterations in cytosolic Ca2+ concentrations following activation of neutrophils with fMLP or PAF were calculated using procedures described in detail elsewhere, either as absolute concentrations [17] or by calculating the areas under the curve using the fura-2/AM tracings [16]. In the case of the latter, the tracings were manually copied, electronically scanned, and saved in jpg format. These images were then imported into the area calculator software program, SketchandCalc, and results are expressed as cm2.
A modification of this procedure, namely, Mn2+ quenching of fura-2 fluorescence, was used to monitor the effects of the test β2-agonists (10 and 100 nM) on store-operated uptake of extracellular Ca2+ by activated neutrophils. Briefly, cells (2 × 106 mL−1) loaded with fura-2/AM as described above were preincubated for 8 min at 37°C followed by successive addition of Mn2+ (600 μM) for a further 2 min period, then the test β2-agonists followed 20 s later by fMLP (1 μM) or PAF (200 nM). Ca2+ influx was then determined according to the rate and magnitude of quenching of fura-2-fluorescence, using an excitation wavelength of 360 nm, which is an isosbestic wavelength, and an excitation wavelength of 500 nm. These results are presented as the traces from 2 separate experiments, as well as by measurement of the rate of decline in fluorescence intensity at 1 and 4 min following addition of the chemoattractants.
2.9. Cell Viability
Neutrophils (1 × 106/mL) were treated with formoterol, indacaterol, or salbutamol, at a final concentration of 100 nM for 10–15 min at 37°C, followed by a 10 min exposure of the cells to propidium iodide (DNA prepstain, Beckman Coulter Miami, FL, USA, 50 μg/mL), at room temperature with flow cytometric detection of uptake of propidium iodide as a marker of cell membrane damage expressed as % viable cells.
2.10. Statistical Analysis
With the exception of the results of the oxygen consumption and fura-2/AM fluorescence experiments, some of which are presented as representative traces, the results of each series of experiments are presented as the mean ± SEM values, where n equals the number of different donors used in each series of experiments, with the number of replicates for each drug concentration and drug-free control system shown in the figure legends. Levels of statistical significance were determined by comparing the absolute values for each drug-treated system with the corresponding values for the relevant drug-free control systems for each assay using the Mann-Whitney test.
3. Results
3.1. Production of Reactive Oxygen Species
These results are shown in Figure 1. Treatment of neutrophils with either formoterol or indacaterol at concentrations of 1–1000 nM caused dose-related inhibition of the generation of ROS which achieved statistical significance at concentrations of ≥10 nM (P < 0.0188–0.004), thereafter gradually leveling off, possibly due to β2AR saturation, with the 2 agents being essentially equipotent. Salbutamol was less effective, only attaining statistically significant inhibition at 1000 nM (P < 0.04).
Figure 1.
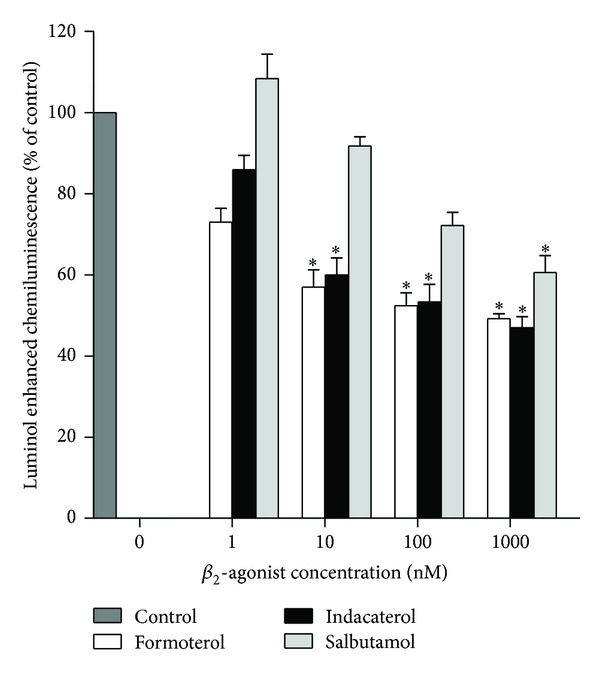
Effects of formoterol, indacaterol, and salbutamol (1–1000 nM) on the luminol-enhanced chemiluminescence responses of neutrophils activated by N-formyl-L-methionyl-L-leucyl-L-phenylalanine (fMLP, 1 μM). The results are expressed as the mean percentage of control ± SEM (n = 5) with duplicate values for each drug concentration and control system in each experiment. The absolute values for unstimulated neutrophils and for cells activated with fMLP in the absence of the drugs were 2136 ± 219 and 20394 ± 1447 relative light units respectively; *P < 0.04–0.004 for comparison with the fMLP-activated, drug-free control system.
3.2. Oxygen Consumption
As shown in Figure 2, which depicts the traces from 3 separate experiments using cells from 3 different donors, activation of neutrophils with fMLP/CB was accompanied by a marked, transient increase in O2 consumption by the cells which was linear for about 1-2 min, declining rapidly thereafter. Inclusion of the test β2-agonists at a fixed final concentration of 100 nM decreased the utilization of O2 by these cells. The mean values ± SEMs for the complete series of experiments (n = 3) for the fMLP/CB-activated control system and for systems containing formoterol, indacaterol, and salbutamol were 65.4 ± 3, 44.7 ± 3 (P < 0.0022 by comparison with the control system), 40.0 ± 4 (P < 0.0022), and 50.3 ± 2 (P < 0.0022) nmol O2 consumed by 2 × 106 cells min−1, respectively, over the 1 min period when the response was linear.
Figure 2.
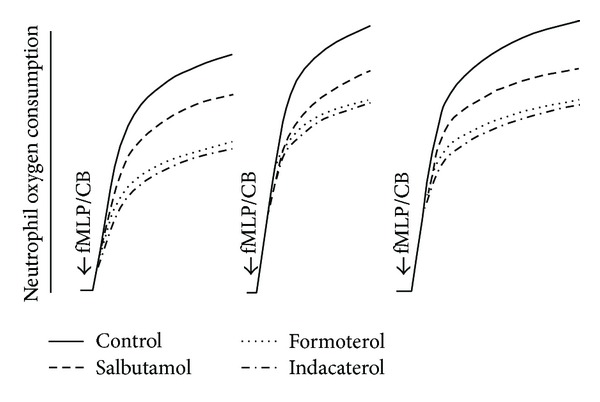
Typical traces from 3 separate experiments (n = 3) showing the effects of formoterol, indacaterol, and salbutamol at 100 nM on the magnitude of oxygen consumption by neutrophils activated with N-formyl-L-methionyl-L-leucyl-L-phenylalanine (1 μM)/cytochalasin B (1 μM) (fMLP/CB).
3.3. Release of Elastase
These results are shown in Figure 3. Treatment of neutrophils with either formoterol or indacaterol at concentrations of ≥ 1 nM caused significant (P < 0.0001) dose-related inhibition of the release of elastase, again leveling off at around 10 nM. Salbutamol was less effective, attaining statistically significant inhibition at concentrations of ≥ 10 nM (P < 0.0001).
Figure 3.
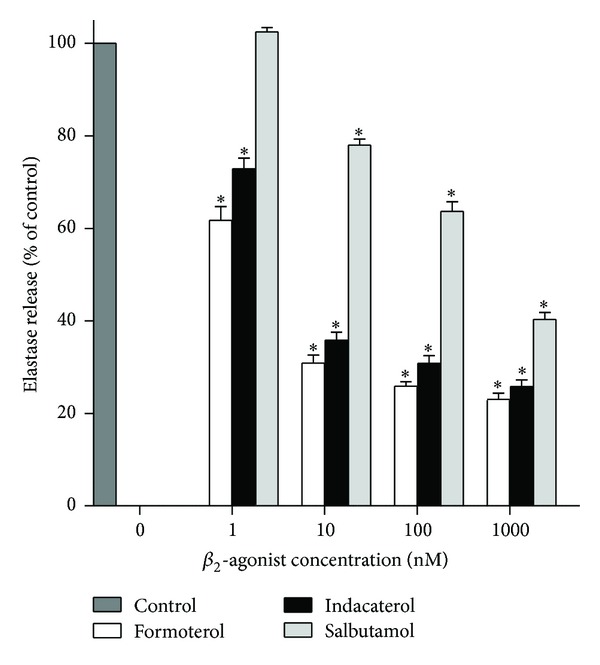
Effects of formoterol, indacaterol, and salbutamol (1–1000 nM) on the release of elastase from neutrophils activated with N-formyl-L-methionyl-L-leucyl-L-phenylalanine (1 μM)/cytochalasin B (1 μM) (fMLP/CB). The results of 3 separate experiments (n = 3) with 5 replicates for each drug concentration and control system in each experiment are expressed as the mean percentage of control ± SEM. The absolute values for the unstimulated control system and for cells activated with fMLP/CB were 47 ± 6 and 1551 ± 78 milliunits elastase per 107 cells. *P < 0.0102–0.0001 for comparison with the drug-free control systems.
3.4. CR3 Expression
Both formoterol and indacaterol, but not salbutamol, at the fixed concentration used (100 nM) caused statistically significant inhibition of expression of CR3 by fMLP/CB-activated neutrophils. The values for the unstimulated control system and for fMLP/CB-activated systems in the absence and presence of formoterol, indacaterol, or salbutamol were 58 ± 4, 121 ± 6, 97 ± 5 (P < 0.0045 by comparison with the control system), 99 ± 5 (P < 0.0071), and 110 ± 6 (NS) mean fluorescence intensity, respectively, (n = 7).
3.5. Leukotriene B4
The effects of the test β2-agonists on the production of LTB4 by neutrophils activated with fMLP or PAF are shown in Figure 4. At the concentrations tested (10 and 100 nM), both formoterol and indacaterol caused dose-related, essentially equivalent, statistically significant (P < 0.001) inhibition of LTB4 production activated by either chemoattractant, while the effects of salbutamol were evident at 100 nM only (P < 0.001).
Figure 4.
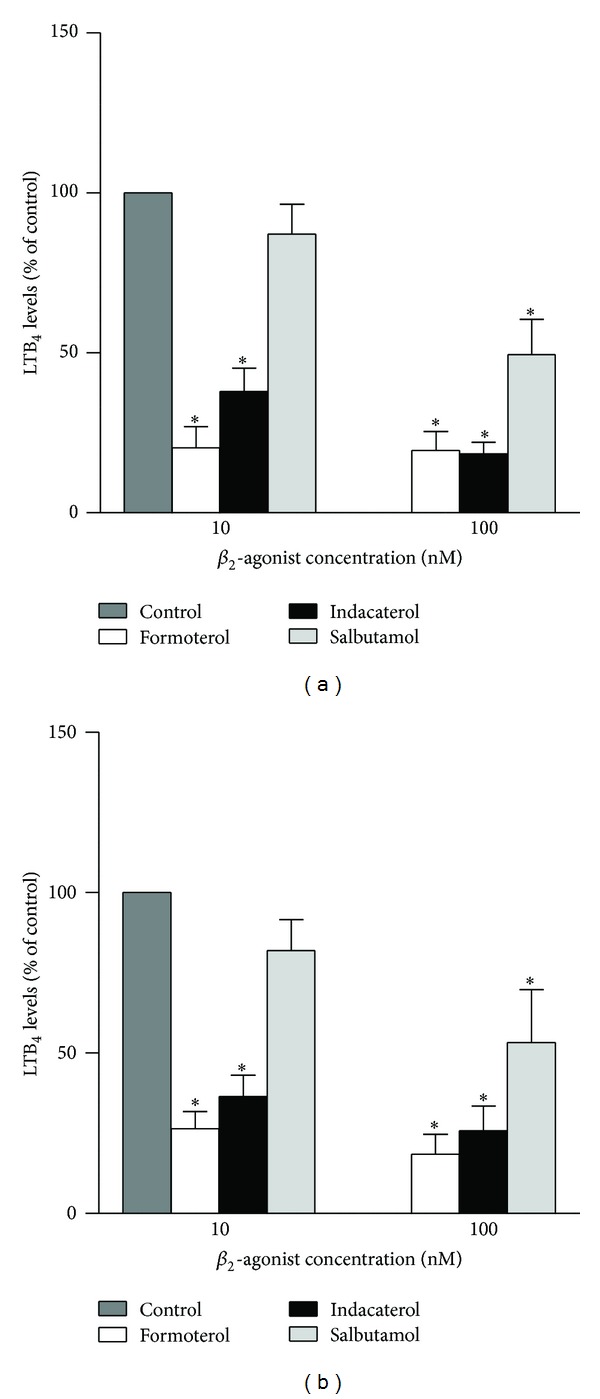
Effects of formoterol, indacaterol, and salbutamol (10 and 100 nM) on the production of LTB4 by fMLP- (a) and PAF- (b) activated neutrophils. The results of 5 separate experiments (n = 5) with 2 replicates for each drug concentration and control system in each experiment are expressed as the mean percentage of control ± SEM. The absolute values for the unstimulated control system and for cells activated with either fMLP or PAF were 33 ± 7, 1095.4 ± 362.2, and 1546 ± 1108.5 pg mL−1. *P < 0.001 for comparison with the drug-free control systems.
3.6. Cyclic AMP
These results are shown in Figure 5. Treatment of neutrophils with formoterol and indacaterol for 20 s at both concentrations used (10 and 100 nM) resulted in significant (P < 0.0001) and comparable elevations in intracellular cAMP, while salbutamol was apparently ineffective at these concentrations. No clear dose-response relationship was evident with formoterol and indacaterol consistent with β2AR saturation at 10 nM of these agents.
Figure 5.
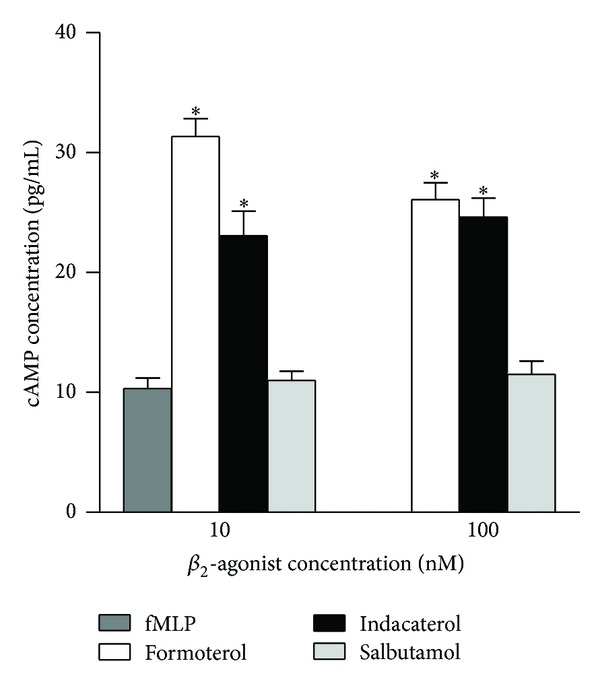
Effects of formoterol, indacaterol, and salbutamol (10 and 100 nM) on cAMP levels in neutrophils. The results of 5 separate experiments (n = 5) with 2 replicates for each drug concentration and control system in each experiment are expressed as the mean ± SEM. The absolute value for the control system was 10.4 ± 0.9 pg mL−1. *P < 0.001 for comparison with the drug-free control systems.
3.7. Cytosolic Ca2+
The results shown in Figure 6 are traces from 2 representative experiments showing the effects of formoterol, indacaterol, and salbutamol (10 and 100 nM) on the alterations in cytosolic Ca2+ following activation of the cells with either fMLP or PAF, while data from the full series of experiments are summarized in Table 1. In the case of fMLP, addition of the chemoattractant to neutrophils resulted in the abrupt, characteristic increase in cytosolic Ca2+ (fura-2 fluorescence) originating predominantly via mobilization of the cation from intracellular stores, rising from a basal concentration of 89 ± 7 nM to 364 ± 31 nM Ca2+. This was followed by a rapid decline over an approximately 2 min period due to clearance of cytosolic Ca2+ and a slowing thereafter due to store-operated influx of the cation, respectively [18]. Treatment of the cells with the β2-agonists did not affect the magnitude of the initial peak increase in cytosolic Ca2+ (not shown). However, as shown in Figure 6 and Table 1, all 3 agents accelerated the rate of decline in fluorescence intensity attaining statistical significance with formoterol and indacaterol at concentrations of 10 nM, but not in the case of salbutamol. These findings are compatible with β2-agonist-mediated enhancement of the clearance of Ca2+ from the cytosol of fMLP-activated neutrophils.
Figure 6.
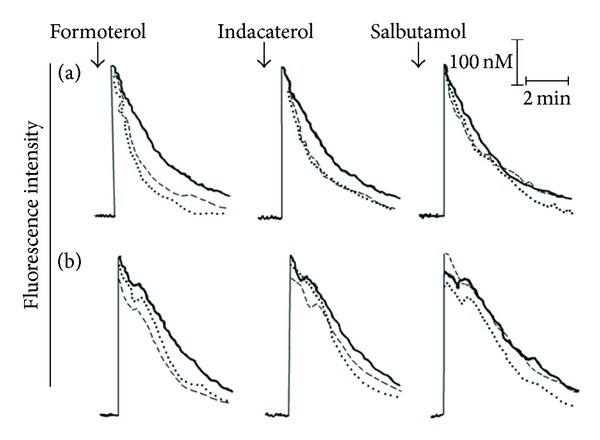
Fura-2 fluorescence traces from 2 representative experiments (n = 4–7 in the series) showing the effects of formoterol, indacaterol, and salbutamol (10 and 100 nM) on the alterations in cytosolic Ca2+ concentrations following activation of the cells with either 1 μM fMLP (a) or 200 nM PAF (b). The 3 lines in each trace correspond to the control system (—) and systems treated with either 10 nM (_ _) or 100 nM (●●●●●) of each drug; ↓ denotes the addition of either fMLP or PAF.
Table 1.
Effects of formoterol, indacaterol, and salbutamol (1, 10, 100, and 1000 nM) on the cytosolic Ca2+ concentrations (area under curve) of FMLP- and PAF-activated neutrophils.
| Agent | FMLP Mean ± SEM (cm2) |
PAF Mean ± SEM (cm2) |
|---|---|---|
| Control (DMSO) | 8.6 ± 0.64 | 13.8 ± 0.53 |
| Formoterol 1 nM | ND | 10.9 ± 0.90 |
| Indacaterol 1 nM | ND | 12.2 ± 1.34 |
| Salbutamol 1 nM | ND | 14.3 ± 1.20 |
| Formoterol 10 nM | 4.8 ± 0.64* | 10.9 ± 0.74* |
| Indacaterol 10 nM | 5.6 ± 0.56* | 10.9 ± 1.08* |
| Salbutamol 10 nM | 7.3 ± 0.50 | 13.6 ± 1.25 |
| Formoterol 100 nM | 4.5 ± 0.28* | 9.8 ± 0.90* |
| Indacaterol 100 nM | 5.1 ± 0.57* | 10.7 ± 0.90* |
| Salbutamol 100 nM | 7.7 ± 0.74 | 12.8 ± 1.40 |
| Formoterol 1000 nM | ND | 8.0 ± 0.74* |
| Indacaterol 1000 nM | ND | 8.4 ± 0.90* |
| Salbutamol 1000 nM | ND | 10.3 ± 1.90 |
ND: not done.
*P = 0.02–0.0003.
Activation of neutrophils with PAF also resulted in an abrupt increase in the concentration of cytosolic Ca2+ which persisted for about 1 min due to rapid influx of the cation, which, relative to fMLP, was followed by a more gradual clearance of the cation as reported previously [19]. The effects of the 3 β2-agonists on PAF-activated alterations in neutrophil cytosolic Ca2+ were comparable to those of fMLP-activated cells, namely, no effects on immediate peak responses, followed by accelerated clearance of the cation, with formoterol and indacaterol being most effective (Figure 6, Table 1).
The effects of the 3 β2-agonists on store-operated influx of extracellular Ca2+ following activation of neutrophils with either fMLP or PAF are shown in Figure 7 which are the traces from 2 representative experiments; these are summarized in Table 2. At the concentrations tested (10 and 100 nM), formoterol and indacaterol effectively inhibited the influx of Ca2+ by cells activated with either fMLP or PAF, while the effects of salbutamol were evident at a concentration of 1000 nM.
Figure 7.

Mn2+-quenching of fura-2 fluorescence traces from 2 representative experiments (n = 3 in the series) showing the effects of formoterol, indacaterol (100 nM), and salbutamol (100 and 1000 nM) on the alterations in cytosolic Ca2+ concentrations following activation of the cells with either 1 μM fMLP (a) or 200 nM PAF (b). The 3 lines in each trace correspond to the control system (—) and systems treated with 100 nM of each drug (●●●●●) or 1000 nM salbutamol (xxxx); ↓ denotes the addition of either fMLP or PAF.
Table 2.
Effects of formoterol, indacaterol, and salbutamol on Mn2+ quenching of fura-2 fluorescence in FMLP- and PAF-activated neutrophils.
| Agent | FMLP-activated neutrophils | PAF-activated neutrophils | ||
|---|---|---|---|---|
| Magnitude of decrement at 1 minute (cm) Mean ± SEM |
Magnitude of decrement at 4 minutes (cm) Mean ± SEM |
Magnitude of decrement at 1 minute (cm) Mean ± SEM |
Magnitude of decrement at 4 minutes (cm) Mean ± SEM |
|
| Control (DMSO) | 1.40 ± 0.09 | 5.40 ± 0.30 | 2.7 ± 0.15 | 7.4 ± 0.67 |
| Formoterol 100 nM | 1.24 ± 0.09 | 3.70 ± 0.13* | 2.5 ± 0.16 | 5.1 ± 0.55* |
| Indacaterol 100 nM | 1.30 ± 0.07 | 3.74 ± 0.19* | 2.4 ± 0.17 | 5.2 ± 0.50 |
| Salbutamol 100 nM | 1.60 ± 0.09 | 5.30 ± 0.22 | 2.7 ± 0.17 | 6.8 ± 0.73 |
*P = 0.04–0.009.
3.8. Cell Viability
Exposure of the neutrophils to formoterol, indacaterol, or salbutamol at a final concentration of 100 nM had no effect on cell viability according to propidium iodide exclusion, the respective values being 95.9 ± 0.5%, 95.9 ± 0.5% and 96.0 ± 0.5% viability relative to a control value of 96.4 ± 0.6% (data from eight determinations).
4. Discussion
The findings of the current study demonstrate that formoterol and indacaterol effectively suppress a spectrum of proinflammatory activities of activated human neutrophils in vitro, while salbutamol is much less effective. The different magnitudes of anti-inflammatory activity observed with formoterol, indacaterol, and salbutamol are most likely due to variability in the intrinsic efficacy of these agents. Formoterol, a full agonist [5] and indacaterol, a near full agonist [20], have higher intrinsic efficacy than the partial agonist, salbutamol [5]. With respect to formoterol and indacaterol, all of the neutrophil functions tested were suppressed at low nanomolar (1–10 nM) concentrations of these agents including the generation of ROS and LTB4, release of the primary granule protease, elastase, and expression of the β2-integrin, CR3. These neutrophil-derived mediators of inflammation are intimately involved in the immunopathogenesis not only of COPD, but also other acute and chronic inflammatory diseases of the airways such as severe persistent bronchial asthma, COPD, cystic fibrosis, bronchiolitis obliterans, and acute respiratory distress syndrome, resulting in pulmonary abnormalities including bronchial hyperresponsiveness and obstruction, mucus hypersecretion, and airway remodeling [10, 21–27].
From a mechanistic perspective, exposure of neutrophils to formoterol and indacaterol, at concentrations equivalent to those which suppressed neutrophil reactivity, resulted in β2AR-mediated increases in intracellular cAMP. This, in turn, apparently underpins the increased efficacy of restoration of Ca2+ homeostasis to neutrophils activated with either fMLP or PAF, with consequent attenuation of Ca2+-dependent proinflammatory activity. Several mechanisms have been described, all involving cAMP-dependent protein kinase (PKA), whereby cAMP promotes accelerated clearance of Ca2+ from the cytosol of activated neutrophils and other cell types (reviewed in 5). However, PKA-mediated enhancement of the efficacy of the Ca2+ resequestering endomembrane Ca2+-ATPase, which diverts mobilized cytosolic Ca2+ back into storage compartments [28], is likely to be the predominant mechanism operative in the setting of the observed β2-agonist/neutrophil interactions. This contention is based on the following observations: (i) the lack of effect of the test agents on the immediate peak cytosolic Ca2+ response following exposure of the cells to fMLP or PAF, excluding interference with phospholipase C and generation of Ca2+-mobilizing inositol triphosphate, and (ii) decreased store-operated influx of extracellular Ca2+ which is probably secondary to efficient utilization of cytosolic Ca2+ for store refilling [28].
The apparent disconnect between the failure to detect elevated levels of intracellular cAMP in the setting of moderate inhibition of oxygen consumption, generation of ROS and LTB4, and release of elastase following treatment of neutrophils with salbutamol at a concentration of 100 nM may be attributed to several reasons. Firstly, as a consequence of both the partial agonist activity and comparatively low partition coefficient of salbutamol, sustained elevations in cAMP may be difficult to detect due to rapid hydrolysis of the cyclic nucleotide by the cyclic AMP-specific phosphodiesterase, PDE4. This contention is substantiated by the findings of a previous study demonstrating that the elevating actions of salbutamol on neutrophil cAMP are dependent on the inclusion of an inhibitor, either selective or nonselective, of PDEs [16]. Alternatively, salbutamol may suppress the proinflammatory activities of neutrophils by mechanisms unrelated to β2AR-mediated activation of adenylyl cyclase [29]. Importantly, the anti-inflammatory effects of formoterol and indacaterol could not be attributed to cellular cytotoxicity.
Despite a considerable body of evidence demonstrating potent, cAMP-dependent, suppressive effects of β2-agonists on cells of the innate and adaptive immune systems in vitro [3, 5, 28], as well as anti-inflammatory activity in animal models of experimental airways inflammation [30], compelling evidence for the existence of these activities in the clinical setting has not been forthcoming [6, 7]. This may be attributable to several possible causes. These are (i) aspects of molecular structure, as mentioned above, such as those in the case of formoterol and indacaterol which confer full agonist activity, good partition coefficients, and long duration of action, as opposed to salbutamol [5]; (ii) failure to attain adequate concentrations in the airways, although inhalation of 24 μg formoterol has been estimated to result in a local concentration of 11 nM [16], which would be effective for this agent, but ineffective in the case of salbutamol; and (iii) lower numbers of β2ARs on inflammatory cells relative to airway smooth muscle cells [8, 11], possibly resulting in an increased propensity to undergo receptor desensitization [8]. Of these, β2AR desensitization is likely to be the major obstacle to harnessing the anti-inflammatory potential of formoterol and indacaterol in the clinical setting. In addition to augmenting β2AR expression by combining β-agonists with inhaled corticosteroids [31, 32], potential strategies to overcome receptor desensitization in the clinical setting include adjunctive therapy using selective and nonselective inhibitors of cyclic AMP PDEs and possibly anticholinergics such as the long-acting antimuscarinic, tiotropium [5].
Numerous clinical trials have confirmed the clinical efficacy of indacaterol for the treatment of patients with moderate and severe COPD [33]. In this clinical setting indacaterol provides extended relief of airflow obstruction and dyspnoea and reduces the frequency of exacerbations, resulting in symptomatic relief and a decreased requirement for rescue medications, thereby improving patient quality of life and adherence to therapy [10, 33, 34]. Indacaterol also appears to have a favourable safety profile. In the case of the cardiovascular system for example, 14 days of therapy did not induce any prolongation of the QT interval [35]. This is important as many patients with COPD are elderly and may have associated comorbid cardiovascular disorders. A mild and transient cough is a common adverse effect described by patients treated with indacaterol [34].
In conclusion, indacaterol, like formoterol, effectively targets the potentially harmful activities of stimulated neutrophils in vitro. These anti-inflammatory, albeit secondary, actions of both agents may contribute to their therapeutic efficacy in COPD.
Acknowledgments
This study was supported by research grants awarded to Ronald Anderson and Charles Feldman by the Medical Research Council and National Research Foundation of South Africa, respectively.
Conflict of Interests
Charles Feldman has acted on the Advisory Boards and/or received honoraria for lectures and/or received assistance for congress travel from Aspen-GSK, Boehringer Ingelheim, Cipla Medpro, and Novartis. The other authors have no conflict of interests to declare.
References
- 1.Cazzola M, Page CP, Rogliani P, Matera MG. β-agonist therapy in lung disease. American Journal of Respiratory and Critical Care Medicine. 2013;187(7):690–696. doi: 10.1164/rccm.201209-1739PP. [DOI] [PubMed] [Google Scholar]
- 2.Sears MR. The FDA-mandated trial of safety of long-acting beta-agonists in asthma: finality or futility? Thorax. 2013;68(2):195–198. doi: 10.1136/thoraxjnl-2012-202414. [DOI] [PubMed] [Google Scholar]
- 3.Johnson M. Effects of β2-agonists on resident and infiltrating inflammatory cells. Journal of Allergy and Clinical Immunology. 2002;110(6):S282–S290. doi: 10.1067/mai.2002.129430. [DOI] [PubMed] [Google Scholar]
- 4.Serezani CH, Ballinger MN, Aronoff DM, Peters-Golden M. Cyclic AMP: master regulator of innate immune cell function. American Journal of Respiratory Cell and Molecular Biology. 2008;39(2):127–132. doi: 10.1165/rcmb.2008-0091TR. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Theron AJ, Steel HC, Tintinger GR, Feldman C, Anderson R. Can the anti-inflammatory activities of β2-agonists be harnessed in the clinical setting? Journal of Drug Design, Development and Therapy. 2013;2013(7):1387–11398. doi: 10.2147/DDDT.S50995.S50995 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Matthay MA, Brower RG, Carson S, et al. Randomized, placebo-controlled clinical trial of an aerosolized β2-agonist for treatment of acute lung injury. American Journal of Respiratory and Critical Care Medicine. 2011;184(5):561–568. doi: 10.1164/rccm.201012-2090OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Smith FG, Perkins GD, Gates S, et al. Effect of intravenous β-2 agonist treatment on clinical outcomes in acute respiratory distress syndrome (BALTI-2): a multicentre, randomised controlled trial. The Lancet. 2012;379(9812):229–235. doi: 10.1016/S0140-6736(11)61623-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Barnes PJ. Effect of β-agonists on inflammatory cells. Journal of Allergy and Clinical Immunology. 1999;104(2, part 2):S10–S17. doi: 10.1016/s0091-6749(99)70269-1. [DOI] [PubMed] [Google Scholar]
- 9.Ray SM, McMillen JC, Treadway SA, Helmer RS, Franks AS. Indacaterol: a novel long-acting β(2)-agonist. Pharmacotherapy. 2012;32(5):456–474. doi: 10.1002/j.1875-9114.2012.01025.x. [DOI] [PubMed] [Google Scholar]
- 10.Han J, Dai L, Zhong N. Indacaterol on dyspnea in chronic obstructive pulmonary disease: a systematic review and meta-analysis of randomized placebo-controlled trials. BMC Pulmonary Medicine. 2013;13, article 26 doi: 10.1186/1471-2466-13-26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.De Coupade C, Gear RW, Dazin PF, Sroussi HY, Green PG, Levine JD. β2-adrenergic receptor regulation of human neutrophil function is sexually dimorphic. British Journal of Pharmacology. 2004;143(8):1033–1041. doi: 10.1038/sj.bjp.0705972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Hoenderkos K, Condliffe A. The neutrophil in chronic obstructive pulmonary disease. American Journal of Respiratory Cell and Molecular Biology. 2013;48(5):531–539. doi: 10.1165/rcmb.2012-0492TR. [DOI] [PubMed] [Google Scholar]
- 13.Barnes PJ. New molecular targets for the treatment of neutrophilic diseases. Journal of Allergy and Clinical Immunology. 2007;119(5):1055–1062. doi: 10.1016/j.jaci.2007.01.015. [DOI] [PubMed] [Google Scholar]
- 14.Tannheimer SL, Wright CD, Salmon M. Combination of roflumilast with a beta-2 adrenergic receptor agonist inhibits proinflammatory and profibrotic mediator release from human lung fibroblasts. Respiratory Research. 2012;13, article 28 doi: 10.1186/1465-9921-13-28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Scola A-M, Loxham M, Charlton SJ, Peachell PT. The long-acting β-adrenoceptor agonist, indacaterol, inhibits IgE-dependent responses of human lung mast cells. British Journal of Pharmacology. 2009;158(1):267–276. doi: 10.1111/j.1476-5381.2009.00178.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Gravett CM, Theron AJ, Steel HC, et al. Interactive inhibitory effects of formoterol and montelukast on activated human neutrophils. The European Respiratory Journal. 2010;36(6):1417–1424. doi: 10.1183/09031936.00157409. [DOI] [PubMed] [Google Scholar]
- 17.Grynkiewicz G, Poenie M, Tsien RY. A new generation of Ca2+ indicators with greatly improved fluorescence properties. The Journal of Biological Chemistry. 1985;260(6):3440–3450. [PubMed] [Google Scholar]
- 18.Anderson R, Mahomed AG. Calcium efflux and influx in f-met-leu-phe (fMLP)-activated human neutrophils are chronologically distinct events. Clinical and Experimental Immunology. 1997;110(1):132–138. doi: 10.1046/j.1365-2249.1997.5051403.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Steel HC, Anderson R. Dissociation of the PAF-receptor from NADPH oxidase and adenylate cyclase in human neutrophils results in accelerated influx and delayed clearance of cytosolic calcium. British Journal of Pharmacology. 2002;136(1):81–89. doi: 10.1038/sj.bjp.0704685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Cazzola M, Matera MG. Emerging inhaled bronchodilators: an update. European Respiratory Journal. 2009;34(3):757–769. doi: 10.1183/09031936.00013109. [DOI] [PubMed] [Google Scholar]
- 21.Mak JCW, Chan-Yeung MMW. Reactive oxidant species in asthma. Current Opinion in Pulmonary Medicine. 2006;12(1):7–11. doi: 10.1097/01.mcp.0000198067.50457.71. [DOI] [PubMed] [Google Scholar]
- 22.Lee K-Y, Ho S-C, Lin H-C, et al. Neutrophil-derived elastase induces TGF-β1 secretion in human airway smooth muscle via NF-κB pathway. American Journal of Respiratory Cell and Molecular Biology. 2006;35(4):407–414. doi: 10.1165/rcmb.2006-0012OC. [DOI] [PubMed] [Google Scholar]
- 23.Li Q, Zhou X, Yu H, Nie X, Xu X. Roles of ROS/TACE in neutrophil elastase-induced mucus hypersecretion in NCI-H292 airway epithelial cells. European Cytokine Network. 2010;21(3):177–185. doi: 10.1684/ecn.2010.0195. [DOI] [PubMed] [Google Scholar]
- 24.Ohnishi H, Miyahara N, Gelfand EW. The role of leukotriene B4 in allergic diseases. Allergology International. 2008;57(4):291–298. doi: 10.2332/allergolint.08-RAI-0019. [DOI] [PubMed] [Google Scholar]
- 25.Lee IT, Yang CM. Role of NADH oxidase/ROS in pro-inflammatory mediators-induced airway and pulmonary diseases. Biochemical Pharmacology. 2012;84(5):581–590. doi: 10.1016/j.bcp.2012.05.005. [DOI] [PubMed] [Google Scholar]
- 26.Gaggar A, Hector A, Bratcher PE, Mall MA, Griese M, Hartl D. Series “matrix metalloproteinases in lung health and disease”: the role of matrix metalloproteinases in cystic fibrosis lung disease. The European Respiratory Journal. 2011;38(3):721–727. doi: 10.1183/09031936.00173210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Sandhaus RA, Torino G. Neutrophil elastase-mediated lung disease. COPD. 2013;10, supplement 1:60–63. doi: 10.3109/15412555.2013.764403. [DOI] [PubMed] [Google Scholar]
- 28.Anderson R, Goolam Mahomed A, Theron AJ, Ramafi G, Feldman C. Effect of rolipram and dibutyryl cyclic AMP on resequestration of cytosolic calcium in FMLP-activated human neutrophils. British Journal of Pharmacology. 1998;124(3):547–555. doi: 10.1038/sj.bjp.0701849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Brunskole Hummel I, Reinartz MT, Kälble S, et al. Dissociations in the effects of β2-adrenergic receptor agonists on cAMP formation and superoxide production in human neutrophils: support for the concept of functional selectivity. PLoS ONE. 2013;8(5) doi: 10.1371/journal.pone.0064556.e64556 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Bosmann M, Grailer JJ, Zhu K, et al. Anti-inflammatory effects of β2 adrenergic receptor agonists in experimental acute lung injury. The FASEB Journal. 2012;26(5):2137–2144. doi: 10.1096/fj.11-201640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Black JL, Oliver BGG, Roth M. Molecular mechanisms of combination therapy with inhaled corticosteroids and long-acting β-agonists. Chest. 2009;136(4):1095–1100. doi: 10.1378/chest.09-0354. [DOI] [PubMed] [Google Scholar]
- 32.Chung KF, Caramori G, Adcock IM. Inhaled corticosteroids as combination therapy with β-adrenergic agonists in airways disease: present and future. European Journal of Clinical Pharmacology. 2009;65(9):853–871. doi: 10.1007/s00228-009-0682-z. [DOI] [PubMed] [Google Scholar]
- 33.Ridolo E, Montagni M, Olivieri E, Riario-Sforza GG, Incorvaia C. Role of indacaterol and the newer very long-acting β2-agonists in patients with stable COPD: a review. International Journal of Chronic Obstructive Pulmonary Disease. 2013;2013(8):425–432. doi: 10.2147/COPD.S49179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Rossi A, Polese G. Indacaterol: a comprehensive review. International Journal of Chronic Obstructive Pulmonary Disease. 2013;2013(8):353–363. doi: 10.2147/COPD.S21625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Khindri S, Sabo R, Harris S, Woessner R, Jennings S, Drollmann AF. Cardiac safety of indacaterol in healthy subjects: a randomized, multidose, placebo- and positive-controlled, parallel-group thorough QT study. BMC Pulmonary Medicine. 2011;11, article 31 doi: 10.1186/1471-2466-11-31. [DOI] [PMC free article] [PubMed] [Google Scholar]