

Abstract
Terminalia is a genus of Combretaceous plants widely distributed in tropical and subtropical regions. Thus, the aim of this study was to quantify the majority compounds of the hydroalcoholic extract (7 : 3, v/v) of the leaves from T. catappa by HPLC-PDA, chemically characterize by hyphenated techniques (HPLC-ESI-IT-MSn) and NMR, and evaluate its mutagenic activity by the Salmonella/microsome assay on S. typhimurium strains TA98, TA97a, TA100, and TA102. The quantification of analytes was performed using an external calibration standard. Punicalagin is the most abundant polyphenol found in the leaves. The presence of this compound as a mixture of anomers was confirmed using HPLC-PDA and 1H and 13C NMR. Mutagenic activity was observed in strains TA100 and TA97a. As the extract is a complex mixture of punicalagin, its derivatives, and several other compounds, the observed mutagenicity may be explained in part by possible synergistic interaction between the compounds present in the extract. These studies show that mutagenic activity of T. catappa in the Ames test can only be observed when measured at high concentrations. However, considering the mutagenic effects observed for T. catappa, this plant should be used cautiously for medicinal purposes.
1. Introduction
As part of our efforts to identify the substances responsible for the pharmacological activities of plants utilized in Brazilian popular medicine (Biota-Fapesp Project number 09/52237-9), we have analyzed the hydroalcoholic extract of the leaves from Terminalia catappa L., which belongs to the family Combretaceae. The trivial names of these plants in Brazil include “amendoeira-da-praia,” “cuca,” and “chapéu de sol.”
In several Asian countries, physicians have used the leaves, bark, and fruit of T. catappa to treat dermatitis and pyresis [1–4]. Moreover, the antimicrobial and antifungal [5–8], antioxidative [9–11], anti-inflammatory [12, 13], hepatoprotective [3, 4], antidiabetic [14], carcinogenesis-preventing effects [15–17], antimalaric [18], and antinociceptive [19] of T. catappa could potentially provide benefits to human health.
Natural products have also been isolated from the plant T. catappa which contains triterpenoids (ursolic acid, asiatic acid), squalene but no caffeine, flavonoids (isovitexin, vitexin, and rutin), gallic acid, hydrolysed tannins such as punicalagin anomers as a major component, punicalin, terflavins A and B, tergallagin, tercatain, chebulagic acid, geranin, granato B, and corilagin [13, 20, 21].
Tannins have been reported to be responsible for decreases in food intake, growth rate, feeding efficiency, net metabolizable energy, and protein digestibility in experimental animals [21]. Incidences of certain cancers, such as esophageal cancer, have been reported to be related to consumption of tannin-rich foods such as betel nuts and herbal teas, suggesting that tannins might be carcinogenic [22]. However, other reports have indicated that the carcinogenic activity of tannins might be related to components that are associated with tannins, rather than the tannins themselves [23, 24]. Tannins have also been reported to exert other physiological effects, such as the acceleration of blood clotting, a reduction in blood pressure, a decrease in serum lipid levels, liver necrosis, and modulation of the immune response [21, 25].
Ellagitannins have been isolated along with gallotannins from various other plant families and show little chemotaxonomic significance. The tannins punicalagin and punicalin, both of which contain a gallagyl unit and were first isolated from the pericarps of pomegranate (Punica granatum, Punicaceae) [26], were a characteristic component in some Terminalia species. This supports the hypothesis that Punicaceae is chemotaxonomically proximate to Combretaceae as classified by Cronquist [27].
Despite its biological properties, the chemical composition may vary according to geographiclocation [28, 29], a fact impairing the standardization of raw material and commercial products for medicinal purposes [30].
Considering that there are no data in the literature on quantification of the majority secondary metabolites of the hydroalcoholic extract of the T. catappa by HPLC-PDA and the popular use of this plant, in this study the punicalagin anomers and ellagic acid were isolated and quantified.
Moreover, the mutagenic effects were evaluated using a Salmonella/microsome assay (Ames test) in the presence and in the absence of in vitro metabolizing systems in the concentrations 22.24, 16.68, 11.12, 5.56, and 2.78 mg/plate.
2. Experimental
2.1. Chemical and Reagents
HPLC-grade methanol (MeOH) and acetonitrile were purchased from J.T. Baker (Baker-Mallinckrodt, Phillipsburg, NJ, USA). Standards of ellagic acid were purchased from Sigma Chemical Co. (St. Louis, MO, USA) and had a purity of 95%. The anomers mixture of punicalagin α and β isolated of the hydroalcoholic extract from T. catappa (with 98% purity) were used as external standards. HPLC-grade water (18 MΩ·cm) was obtained using a direct Milli-Q purification system (Millipore Co., Bedford, MA, USA). Sep-Pak RP18 cartridges (500 mg/mL) for solid-phase extraction (SPE) were purchased from Phenomenex Co. (Torrance, CA, USA).
2.2. Apparatus
The HPLC system used was a JASCO 2010 HPLC (Jasco, Tokyo, Japan) equipped with PU-2089S Plus pump, a MD-2018 Plus Photodiode Array Detector (PDA), an AS-2055 Plus auto sampler, and column oven (CO-2065 plus). The software ChromNav (Workstation JASCO-ChromNav 1.18.03) was used to control the analytical system and data collection and processing. To purify the substances, a semipreparative JASCO HPLC equipped with two PU-2086 pumps, a high pressure mixer (Model MX-2080-32), a PDA detector (model MD-2018 Plus), and a Rheodyne 500 μL manual injector loop was used. The software used to operate the equipment and data processing was ChromNAV version 1 : 18 : 03.
The mass spectrometry experiments were performed on LCQ Fleet equipment (Thermo Scientific) equipped with a dispositive of direct insertion of the sample via flow injection analysis (FIA). The studied matrix was analyzed by electrospray ionization (ESI), and fragmentation in multiple stages (MS2, MS3, and MSn) was performed at an ion trap (IT) interface. The negative mode was selected for the generation and analysis of the mass spectra for the first order (MS) and for the remaining multistage experiments under the following conditions: capillary voltage, −25 V; voltage spray, −5 kV; and capillary temperature, 275°C. A carrier gas (N2) with a flow of 8 arbitrary units (A.U.) was used, and the collision gas was helium (He). The track acquisition was 100–2000 m/z. Xcalibur version 1.3 software (Thermo Finnigan) was used to acquire and process the data.
The 1H- and 13C-NMR experiments were performed using a 300 MHz (11.7 T) nuclear magnetic resonance spectrometer (Varian Inova). CD3OD was used as the solvent (Sigma Aldrich).
2.3. Plant Material
The leaves of T. catappa were collected from the beach-front city of Santos,SP, with the permission of the municipality, in April 2010 and were identified by botanist Paulo Salles Penteado of University Santa Cecilia and registered (M Tomaz 01) in the Herbarium of the University of Santa Cecilia (HUSC). Authorization was obtained from IBAMA (Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis) to the number 33152.
2.4. Extraction
The infusion and hydroalcoholic extract were prepared according to the experimental protocol established in Biota-Fapesp Project, which this work is linked (process number 09/52237-9).
2.4.1. Preparation of the Infusion of the Leaves from T. catappa
The leaves (1 g) of T. catappa, after drying, were prepared by infusion as their use in folk medicine [31].
2.4.2. Preparation of the Hydroalcoholic Extract of the Leaves from T. catappa
The dried and powdered leaves (524.6 g) of T. catappa were extracted by percolation at room temperature with a mixture of EtOH : H2O (7 : 3, v/v). The solvents were evaporated to dryness at low pressure to give 33.1 g of the crude hydroalcoholic extract (8.7%). The extract was analyzed by FIA-ESI-IT-MSn and HPLC-PDA.
2.4.3. Isolation of the Fraction Containing the Punicalagin Anomers by Semipreparative HPLC
A total of 14 g of hydroalcoholic extract (7 : 3, v/v) was dissolved in H2O : MeOH (8 : 2, v/v) and partitioned successively with hexane (500 mL) and ethyl acetate (500 mL), giving 0.68 g (4.8%) (hexane fraction), 1.93 g (13.78%) (ethyl acetate fraction), and the 10.15 g (72.5%) (hydromethanol fraction, 8 : 2 v/v).
2.5. Identification of the Punicalagin Anomers and Ellagic Acid in the Hydroalcoholic Extract
Identification of the punicalagin anomers was performed using FIA-ESI-IT-MSn and compared with the literature data [32], 1H and 13C NMR [33] and HPLC-PDA.
For the FIA-ESI-IT-MSn assay, 10 mg of the crude hydroalcoholic extract was dissolved in 1 mL of MeOH : H2O (1 : 1, v/v) after using an ultrasonic bath for 5 minutes. The samples were then filtered through a 0.22 μm PTFE filter, and aliquots of 20 μL were directly injected into the FIA-ESI-IT-MSn system.
For the HPLC-PDA a clean-up step was performed to remove any contaminants; the solution was purified by solid phase extraction (SPE) using Phenomenex Strata C18 cartridges (500 mg of stationary phase) that were previously activated with 5 mL of MeOH and equilibrated with 5 mL of MeOH : H2O (1 : 1, v/v). The compounds were eluted from cartridges using 1 mL of MeOH : H2O (1 : 1, v/v) with a final volume of 5 mL. The samples were then filtered through a 0.22 μm PTFE filter and dried. The dry extract was diluted to 10 mg/mL in HPLC solvent. Aliquots of 20 μL were injected directly into the HPLC-PDA with detection at 270 nm. The identification of the different compounds in the chromatographic profile of the hydroalcoholic extract was done by comparing their retention times (t r) and UV spectra with those obtained for the pure standards (ellagic acid, Sigma) and the compounds isolated from the hydroalcoholic extract of T. catappa (punicalagin anomers).
For the confirmation of the presence of ellagic acid experiments were done by coinjection. The hydroalcoholic extract (15 mg) was dissolved in 1 mL of MeOH : H2O (1 : 1, v/v) and subjected to a clean-up procedure using SPE cartridge RP18 (50 mg). The sample was then filtered through a PTFE filter (0.22 μm) and dried at room temperature. The resulting solid was redissolved in MeOH : H2O to a concentration of 10 mg/mL. Two milligrams of ellagic acid standard (Sigma) was then added to this solution, and the solution was centrifuged for 2 min and filtered through a 0.22 μm filter. A hydro column with a flow rate of 1 mL/min was used with a gradient of 5–60% MeOH for 60 minutes, and the absorbance at 270 nm was detected. The chromatograms of the hydroalcoholic extract were compared before and after the addition of the ellagic acid standard. From the coinjection of ellagic acid and the hydroalcoholic extract, we confirmed the presence of ellagic acid from the significant increase in the peak eluting at t r 47.30 min (peak 3).
2.6. Conditions for Quantification of Punicalagin (Anomers) and Ellagic Acid
The quantification of analytes was performed using an external calibration standard [34, 35]. The curves were constructed using standards of the commercial ellagic acid (Sigma) and the isolated punicalagin anomers. We evaluated the content of each punicalagin anomer and the total content of punicalagins in leaves of T. catappa. A stock solution of 1000 μg/mL ellagic acid was prepared, and dilutions to 500, 250, 125, 62.5, 15.6, 7.8, and 3.9 μg/mL were made. For punicalagin, a stock solution at concentrations of 4000 μg/mL of the α-anomer (peak 1) and 4000 μg/mL of the β-anomer (peak 2) was prepared. From the stock solution, serial dilutions were made to reduce the concentration in 2 : 1 increments (2000, 1000, 500, 250, 125, 62.50, and 31.25 μg/mL) for both the α- and β-anomers. Each concentration level was analyzed in triplicate. The mean areas of the chromatographic peaks obtained were interpolated as a function of concentration using linear regression and were used to generate the calibration curves. The linear coefficients (a) and angle (b) were obtained from the calibration curves. The correlation coefficient (r 2) and limits of detection (LOD) and quantitation (LOQ) for peaks 1 and 2 corresponding to the α- and β-anomers of punicalagin [36] were obtained. The accuracy of the HPLC method was estimated from the recovery tests. The recovery tests were performed by adding known concentrations (low, medium, and high) of the reference ellagic acid (15, 60, and 250 μg/mL) and isolated punicalagin anomer (250, 500, and 2000 μg/mL) standards. The average recovery value was calculated using the following formula: recovery % = [(C f/C nf + C pd)] × 100, where C f corresponds to fortified concentration, C nf corresponds to the unfortified concentration and C pd corresponds to the concentration of the added standard.
Repeatability of intra- and interday was done to determine the accuracy of the developed method. The relative standard deviation (RSD) was taken as a measure of accuracy. Repeatability of intra- and interday was determined in six replicates within the range found in the extract in a day and on three consecutive days, respectively.
2.7. Salmonella/Microsome Assay
Mutagenic activity was evaluated using the Salmonella/microsome assay with the Salmonella typhimurium tester strains TA98, TA100, TA97a, and TA102, which were kindly provided by Dr. B. N. Ames (Berkeley, CA, USA), with (+S9) and without (−S9) metabolization using the preincubation method [37]. The strains were grown overnight from frozen cultures for 12–14 h in Oxoid Nutrient Broth number 2. The metabolic activation mixture (S9 fraction), prepared from the livers of Sprague-Dawley rats treated with the polychlorinated biphenyl mixture Aroclor 1254 (500 mg/kg), was purchased from Molecular Toxicology Inc. (Boone, NC, USA) and freshly prepared before each test. The metabolic activation system consisted of 4% S9 fraction, 1% 0.4 M MgCl2, 1% 1.65 M KCl, 0.5% 1 M D-glucose-6-phosphate disodium, 4% 0.1 M NADP, 50% 0.2 M phosphate buffer, and 39.5% sterile distilled water. For the determination of the mutagenic activity, five different concentrations of extract (1.56 to 22.24 mg/plate), diluted in DMSO, were assayed. The concentrations of the sample were selected on the basis of a preliminary toxicity test. In all subsequent assays, the upper limit of the dose range tested was either the highest nontoxic dose or the lowest toxic dose determined in the preliminary assay. Toxicity was detected either as a reduction in the number of histidine revertants (His+) or as a thinning of the auxotrophic background (i.e., background lawn).
The various concentrations of extract to be tested were added to either 0.5 mL of 0.2 M phosphate buffer or 0.5 mL of 4% S9 mixture with 0.1 mL of bacterial culture and then incubated at 37°C for 20–30 min. Next, 2 mL of top agar was added, and the mixture was poured onto a plate containing minimal agar.
The plates were incubated at 37°C for 48 h, and the His+ revertant colonies were counted manually. All experiments were analyzed in triplicate. The results were analyzed with the statistical software package Salanal 1.0 (U.S. Environmental Protection Agency, Monitoring Systems Laboratory, Las Vegas, NV, from the Research Triangle Institute, RTP, NC, USA), adopting the Bernstein et al. [38] model. The data (revertants/plate) were assessed by analysis of variance (ANOVA) followed by linear regression. The mutagenic index (MI), which is the average number of revertants per plate with the test compound divided by the average number of revertants per plate with the negative (solvent) control, was also calculated for each concentration tested. A sample was considered mutagenic when a dose-response relationship was detected and a twofold increase in the number of mutants (MI ≥ 2) was observed for at least one concentration [39]. The standard mutagens used as positive controls in experiments without the S9 mix were 4-nitro-o-phenylenediamine (10.0 μg/plate, TA98), sodium azide (1.25 μg/plate, TA100), mitomycin (0.5 μg/plate, TA102) and 2-anthramine (1.25 μg/plate, TA98, TA100), and 2-aminofluorene (10.0 μg/plate, TA102) in the presence of S9. DMSO served as the negative (solvent) control.
3. Results and Discussion
The analysis of mass spectra of the hydroalcoholic extract (Figure 1) showed the precursor ions [M-H]− m/z 1083 (punicalagin), m/z 781 (punicalin), m/z 601 (gallagic acid), and m/z 301 (ellagic acid) [32]. Second-order fragmentation (MS2) confirmed the presence of the metabolites shown in the scheme of Figure 2 for substances 1–4 (Table 1). Seeram et al. [32] studying the commercial juice industry of the pomegranate husk identified the same fragmentation of these same compounds.
Figure 1.
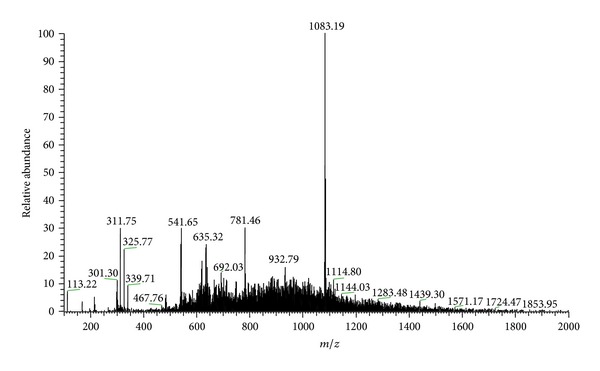
First-order mass spectrum of the hydroalcoholic extract from the leaves of T. catappa in the negative mode. Range of ions with m/z 100–2000 Da.
Figure 2.
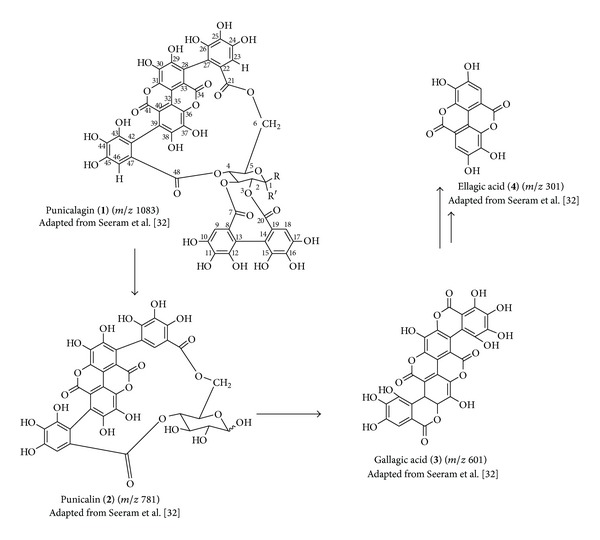
Punicalagin (1), punicalin (2), gallagic acid (3), and ellagic acid (4). Adapted from Seeram et al. [32].
Table 1.
Identification of substances in T. catappa by FIA-ESI-IT-MSn.
| [M–H]− | MSn ions | Identification |
|---|---|---|
| 1083 | 781 [M-152-152-H]−
601 [M-152-152-180-H]− |
Punicalagin (1) |
| 781 | 601 [M-180-H]− | Punicalin (2) |
| 601 | 409 [M-191-H]− | Gallagic acid (3) |
| 301 | 257 [M-44-H]−
229 [M-44-28-H]− |
Ellagic acid (4) |
The chromatographic profile by HPLC-PDA of the hydroalcoholic extract and the infusion are shown in Figures 3 and 4. In the chromatogram, three major peaks (1–3) eluting at t r 14.75 min, t r 18.80 min, and t r 47.30 min are observed. Figures 3 and 4 show the UV region of the spectrum obtained using a PDA detector for the peaks eluting at t r 14.75 min (peak 1) and t r 18.80 min (peak 2). The data confirm the presence of both punicalagin anomers with absorption maxima at λ max 218, 260, and 379 nm. The compound eluting at t r 47.30 min (peak 3) corresponds to ellagic acid with λ max values of 250, 306, and 368 nm, as confirmed by coinjection experiments with a standard analyte (Figures 3 and 4). The data obtained for UV anomers punicalagin and ellagic acid are consistent with those found in the literature [40] and are characteristic of gallagyl chromophore [41].
Figure 3.
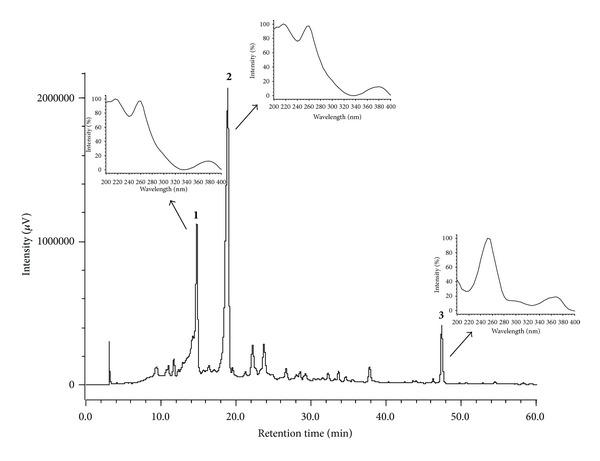
Chromatogram of hydroalcoholic extract of the leaves of T. catappa dissolved in MeOH : H2O (1 : 1 v/v), at 10 mg/mL. The method utilized a hydro column at a flow rate of 1 mL/min with a gradient of 5–60% MeOH for 60 min HPLC-PDA (Jasco), 270 nm. The UV spectra from t r 14.75 (peak 1) min and at t r 18.86 min (peak 2), characteristic of the α- and β-anomers of punicalagin (1, 2) and ellagic acid (3).
Figure 4.
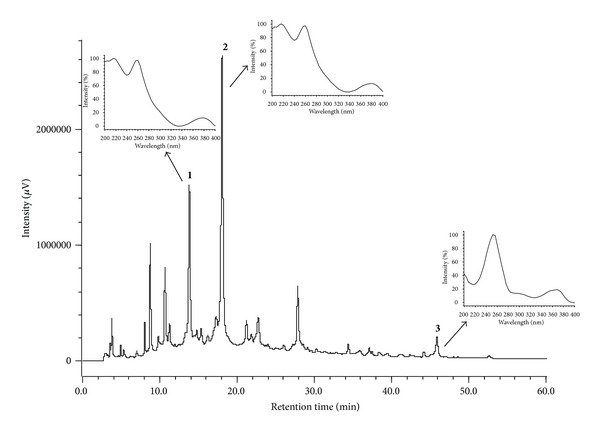
Chromatogram of infusion of the leaves of T. catappa. Gradient 5–60% MeOH, 60 min, 270 nm. Peaks 1 and 2 = punicalagins, 3 = ellagic Acid. The UV spectra of peak 1, eluting at t r 14.70 min and peak 2 at t r 18.70 min, characteristic of the α- and β-punicalagin anomers. Peak 3, at t r 47.30 min.
Peaks 1 and 2 (Figure 3) were partitioned (item 2.4.3), collected separately, and analyzed by HPLC-PDA (Figure 5). The analysis of the peaks by HPLC-PDA with t r 14.75 min and 18.80 min, respectively, display an interesting behavior, as they resolve into two peaks with distinct retention times when analyzed under the same conditions to obtain peaks at similar retention times with the same UV spectra. This indicates that the compounds are the mixture of anomers (α- and β-punicalagin). These compounds have absorption maxim at λ max 218, 260, and 379 nm and are characteristic of the chromophore gallagyl [41]. For peak 1, the HPLC-PDA analysis shows the presence of two peaks (1a and 1b) with retention times of 15.29 min and 21.07 min, respectively (Figure 5).
Figure 5.
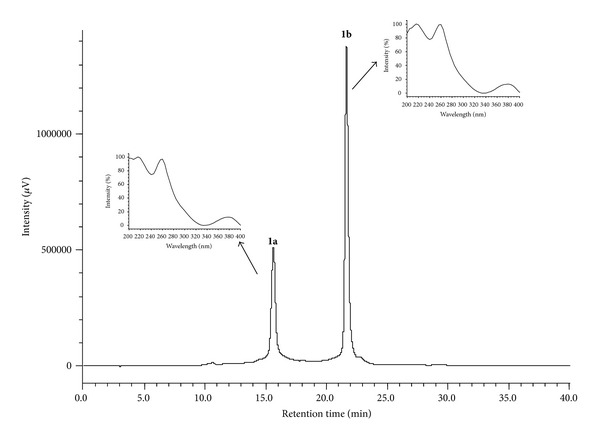
Chromatogram peak 1a (α-punicalagin) and peak 1b (β-punicalagin) collected from H2O : MeOH (8 : 2 v/v) at 10 mg/mL of the leaves of T. catappa, eluting at t r 15.29 min (1a) and t r 21.07 min (1b). The flow rate was 1 mL/min with a gradient 5–30% MeOH for 30 minutes using a hydro column. HPLC-PDA (Jasco), 270 nm. The UV spectra of peak 1, eluting at t r 15.29 min (1a) and at t r 21.07 (1b) min, characteristic of the α- and β-punicalagin anomers.
In solution, punicalagin rapidly interconverts between the α- and β-anomers (Figure 2, compound 1). The 13C NMR (Table 2) spectrum reveals the presence of two anomeric carbons at δ 89.87 and δ 93.77, corresponding to punicalagins α (present in lesser proportion in peak 1) and β (present in a greater proportion in peak 2), respectively. The stereochemistry at the anomeric position can be easily determined by measuring the coupling constant between the anomeric proton H-1 and the adjacent proton H-2 in the 1H NMR spectrum. A large coupling constant (J H-1,H-2 > 5 Hz) generally indicates a β-glycosidic configuration (substituent in the equatorial orientation), whereas a small coupling constant (J H-1,H-2 = 0–5, Hz) indicates an α-glycosidic configuration (substituent in the axial orientation). The chemical shift and the coupling constant corresponding to the α-anomer is δ 5.08 (J = 3.5 Hz) and for the β-anomer is δ 4.66 (J = 8.0 Hz). NMR data of the isolated anomers (α- and β-punicalagin) are shown in the Table 2 (enumeration of carbons and hydrogens in Figure 2) compared with data found in the literature [33]. The deprotonated molecule generated in the FIA-ESI-IT-MS [M-H]− at m/z 1083 (1084 MW) (Table 1) and the UV spectra exhibited by α- and β-punicalagin confirmed this identification (Figure 5).
Table 2.
Chemical shifts (1H and 13C) of the α- and β-punicalagins in CD3OD isolated from T. catappa.
| Position | α-punicalagin | β-punicalagin | ||||
|---|---|---|---|---|---|---|
| δ (1H) | Multiplicity (Hz) | δ (13C) | δ (1H) | Multiplicity (Hz) | δ (13C) | |
| 1 | 5.08 | d(3.5) | 89.87 | 4.66 | d(8.0) | 93.77 |
| 2 | 4.87 | dd(3.0, 8.0) | 73.53 | 4.62 | dd(3.0, 8.1) | 75.48 |
| 3 | 5.20 | t(9.3) | 76.08 | 4.92 | t(10.0) | 78.78 |
| 4 | 4.78 | t(9.0) | 70.23 | 4.85 | t(10.0) | 74.88 |
| 5 | 3.23 | t(9.0) | 66.33 | 2.57 | dd(9.9, 11.0) | 71.58 |
| 6a | 2.10 | d(9.9) | 63.63 | 4.05 | d(9.9) | 63.03 |
| 6b | 4.09 | dd(7.0, 7.0) | 63.63 | 4.10 | d(7.8) | 63.03 |
| 9 | 6.52 | s | 105.46 | 6.54 | s | 106.21 |
| 18 | 6.66 | s | 107.41 | 6.68 | s | 106.21 |
| 23 | 6.73 | s | 108.76 | 6.77 | s | 109.51 |
| 46 | 6.94 | s | 110.11 | 6.97 | s | 110.71 |
The linear regression analysis of the peak area for α- and β-punicalagin, the total punicalagins, and ellagic acid are showed in the Table 3. Thus, it becomes necessary to use sufficient number of standard solutions to adequately define the relationship between concentration and response [42]. These results demonstrate the linearity of the method over the analyzed concentration range.
Table 3.
Parameters for quantification of the isolated compounds.
| Linear coefficient (a) | Slope (b) | Correlation coefficient | LOD µg/mL |
LOQ µg/mL |
|
|---|---|---|---|---|---|
| Ellagic acid | 21753 | 71514 | 0.998 | 1.00 | 3.04 |
| α-punicalagin | 37326 | 11861 | 0.997 | 10.38 | 31.46 |
| β-punicalagin | 44760 | 16830 | 0.999 | 8.77 | 26.59 |
| Total punicalagins | 82086 | 28691 | 0.998 | 9.44 | 28.61 |
The recovery percentages for ellagic acid and total punicalagins were found to be 87% and 86%, respectively (Table 4). The results were presented as mean and standard deviation of the percentages recovered. The results showed that the accuracy of the method is very good for all analytes (RSD < 2%). The relative standard deviations also showed that the process of sample preparation is reproducible [43].
Table 4.
Determining the accuracy of the method for the determination of each of the substances quantified.
| Substances investigated | Repetition (%) | Recovery mean | DP | RSD (%) | ||
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | ||||
| Ellagic acid | 90.4 | 87.75 | 84.52 | 87.56 | 2.94 | 0.03 |
| Total punicalagins | 86.9 | 86.26 | 85.86 | 86.34 | 0.52 | 0.01 |
RSD (%) = RSD: relative standard deviation (100 × SD/mean).
The relative standard deviations (RSDs) for repeatability testing intra- and interday analyses are shown in Table 5. The result is presented as relative standard deviation from the average. The method showed excellent repeatability, with all RSD values lower than 1% [44].
Table 5.
Repeatability test results for each substance quantified.
| Substances investigated | Intraday (n = 6) | Interday (n = 6) | ||||
|---|---|---|---|---|---|---|
| Mean (%) | DP | RSD (%) | Mean (%) | DP | RSD (%) | |
| Ellagic acid | 87.44 | 0.12 | 0.001 | 87.53 | 0.19 | 0.002 |
| Total Punicalagins | 86.34 | 0.19 | 0.002 | 86.19 | 0.17 | 0.002 |
RSD (%) = RSD: relative standard deviation (100 × SD/mean).
Table 6 shows the mean number of revertants/plate (M), the standard deviation (SD), and the mutagenic index (MI) after the treatments with the T. catappa hydroalcoholic extract, observed in S. typhimurium strains TA98, TA100, TA102, and TA97a, in the presence (+S9) and absence (−S9) of metabolic activation.
Table 6.
Mutagenic activity expressed as the mean and standard deviation of the number of revertants/plate and mutagenic index (in parenthesis) in Salmonella typhimurium TA98, TA100, TA97a, and TA102 strains treated with hydroalcoholic extract of T. catappa at various doses, with (+S9) or without (−S9) metabolic activation.
| Treatments | Number of revertants (M ± SD)/ plate and MI | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| TA98 | TA100 | TA102 | TA97a | |||||||
| mg/plate | −S9 | +S9 | mg/plate | −S9 | +S9 | −S9 | +S9 | −S9 | +S9 | |
| Terminalia catappa | 0.0a | 16 ± 3 | 68 ± 1 | 0.0a | 93 ± 6 | 87 ± 4 | 352 ± 30 | 435 ± 43 | 172 ± 2 | 99 ± 1 |
| 2.78 | 17 ± 6 (1.1) | 75 ± 7 (1.1) | 1.56 | 91 ± 2 (1.0) | 88 ± 9 (1.0) | 461 ± 11* (1.3) | 481 ± 23 (1.1) | 223 ± 3* (1.3) | 127 ± 3 (1.3) | |
| 5.56 | 21 ± 3 (1.3) | 75 ± 7 (1.1) | 3.12 | 188 ± 19** (2.0) | 90 ± 6 (1.0) | 514 ± 16* (1.5) | 668 ± 75* (1.5) | 252 ± 12* (1.5) | 102 ± 7 (1.0) | |
| 11.12 | 18 ± 5 (1.1) | 57 ± 1 (0.8) | 6.25 | 210 ± 24** (2.3) | 91 ± 6 (1.0) | 653 ± 22* (1.9) | 689 ± 57* (1.6) | 287 ± 14* (1.7) | 95 ± 12 (1.0) | |
| 16.68 | 14 ± 3 (0.9) | 75 ± 4 (1.1) | 9.38 | 40 ± 4 (0.4) | 95 ± 1 (1.1) | 563 ± 32* (1.6) | 728 ± 28* (1.7) | 337 ± 19** (2.0) | 93 ± 10 (0.9) | |
| 22.24 | 2 ± 1 (0.1) | 68 ± 10 (1.0) | 12.5 | Nd | 88 ± 4 (1.0) | 648 ± 32* (1.8) | 621 ± 23* (1.4) | 397 ± 19** (2.3) | 83 ± 2 (0.8) | |
| Ctrol+ | 720 ± 63 | 500 ± 57 | Ctrol+ | 1225 ± 75 | 700 ± 80 | 1143 ± 53 | 1309 ± 38 | 1426 ± 67 | 700 ± 45 | |
*P < 0.05 (ANOVA); **P < 0.01 (ANOVA), 0a: Negative control: dimethyl sulfoxide (DMSO—75 μL/plate); Positive Control: 4-nitro-o-phenylenediamine (10.0 μg/plate—TA98 and TA97a); sodium azide (1.25 μg/plate—TA100); mitomycin (0.5 μg/plate—TA102), in the absence of S9, and 2-anthramine (1.25 μg/ plate—TA98, TA100, TA97a); 2-aminofluorene (10.0 μg/plate—TA102), in the presence of S9. Nd: not determined.
In the TA98 strain, the T. catappa hydroalcoholic extract did not induce any increase in the number of revertant colonies under the conditions used in this study. The other strains were more sensitive to the toxic effects of extract, and it was thus necessary to decrease the doses.
Only in the absence of the external metabolizing system, S9 mix, in the S. typhimurium strains TA100 and TA97a, the T. catappa hydroalcoholic extract induced an increase in the number of revertant colonies relative to the negative control, with a MI higher than 2.0 at the concentration of 3.12 mg/plate in strain TA100 and 9.38 mg/plate in strain TA97a, indicating the direct mutagenic activity for these strains.
In the TA102 strain (+S9 and –S9), the T. catappa hydroalcoholic extract shows signs of mutagenicity, with MI values approximately 2.
The S. typhimurium test strain TA97a detects frameshift mutations in C-C-C-C-C-C; +1 cytosine and TA98 frameshift in DNA target-C-G-C-G-C-G-C-G, the S. typhimurium tester strain TA100 is capable of revealing base-pair substitution point mutations and the S. typhimurium tester strain TA102 is normally used to detect cross-linking agents and base-pair substitution mutations [39]. Thus, according to the strains involved, the T. catappa hydroalcoholic extract mainly induces substitution of base pairs (TA100) and frameshift mutations (TA97a).
Ko et al. [17] evaluated the toxicity and mutagenicity of supercritical carbon dioxide (SC-CO2) extracts of T. catappa leaves at a dose of 0.5 mg/plate in the strain TA100 (detect base-pair substitution) and observed absence of mutagenic effect. The absence of mutagenic effect was also observed for TA102 and TA98 when the ethanolic extract was evaluated.
These studies show that mutagenic activity of T. catappa in the Ames test can only be observed when measured at high concentrations. The results of this study are consistent with a review of the literature indicating that high concentrations of tannins (15 and 60 mM) can cause DNA damage. These data corroborate the observations of Gupta et al. [45], which indicated that all tannins result in polyphenol toxicity at high concentrations [46].
4. Conclusions
In the hydroalcoholic extract of T. catappa, tannins were the most abundant compounds observed, and the anomers α- and β-punicalagin were the major compounds. Moreover, considering that the extract is a complex mixture of several unknown organic compounds, the mutagenicity observed may be explained in part by a synergy between compounds present in the extract.
The dosage and type of tannin involved are critical for these effects. Thus, the results obtained in this study are useful for better understanding the pharmacological activities of T. catappa. However, considering the mutagenic effects observed in this study, this plant should be used cautiously for medicinal purposes.
Based on the obtained results, it can be concluded that the developed method (HPLC-PDA) is suitable for its purpose, namely, the determination of anomers punicalagin and ellagic acid in extract of T. catappa.
Acknowledgments
The authors are grateful for Grant 09/52237-9, São Paulo Research Foundation (FAPESP) for providing financial aid to W. Vilegas and L. C. Santos. The authors also thank Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for grants to L. C. Santos, E. A. Varanda and W. Vilegas.
Conflict of Interests
The authors declare that they have no conflict of interests.
References
- 1.Yeh CB, Hsieh MJ, Chien MH, Lin PY, Chiou HL, Yang SF. Terminalia catappa exerts antimetastatic effects on hepatocellular carcinoma through transcriptional inhibition of matrix metalloproteinase-9 by modulating NF-κB and AP-1 activity. Evidence Based and Complementary Alternative Medicine. 2012;2012:11 pages. doi: 10.1155/2012/595292.595292 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Morioka T, Suzui M, Nabandith V, et al. Modifying effects of Terminalia catappa on azoxymethane-induced colon carcinogenesis in male F344 rats. European Journal of Cancer Prevention. 2005;14(2):101–105. doi: 10.1097/00008469-200504000-00005. [DOI] [PubMed] [Google Scholar]
- 3.Tang X, Gao J, Wang Y, et al. Effective protection of Terminalia catappa L. leaves from damage induced by carbon tetrachloride in liver mitochondria. Journal of Nutritional Biochemistry. 2006;17(3):177–182. doi: 10.1016/j.jnutbio.2005.06.008. [DOI] [PubMed] [Google Scholar]
- 4.Tang X-H, Gao L, Gao J, et al. Mechanisms of hepatoprotection of Terminalia catappa L. extract on D-galactosamine-induced liver damage. American Journal of Chinese Medicine. 2004;32(4):509–519. doi: 10.1142/S0192415X04002156. [DOI] [PubMed] [Google Scholar]
- 5.Fyhrquist P, Mwasumbi L, Hæggström C-A, Vuorela H, Hiltunen R, Vuorela P. Ethnobotanical and antimicrobial investigation on some species of Terminalia and Combretum (Combretaceae) growing in Tanzania. Journal of Ethnopharmacology. 2002;79(2):169–177. doi: 10.1016/s0378-8741(01)00375-0. [DOI] [PubMed] [Google Scholar]
- 6.Nair R, Chanda S. Antimicrobial activity of Terminalia catappa, Manilkara zapota and Piper betel leaf extract. Indian Journal of Pharmaceutical Sciences. 2008;70(3):390–393. doi: 10.4103/0250-474X.43012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Manzur A, Raju A, Rahman S. Antimicrobial activity of Terminalia catappa extracts against some pathogenic microbial strains. Pharmacology & Pharmacy. 2011;2:299–305. [Google Scholar]
- 8.Chanda S, Rakholiya K, Nair R. Antimicrobial activity of Terminalia catappa L. leaf extracts against some clinically important pathogenic microbial strains. Chinese Medicine. 2011;2:171–177. [Google Scholar]
- 9.Chen P-S, Li J-H. Chemopreventive effect of punicalagin, a novel tannin component isolated from Terminalia catappa, on H-ras-transformed NIH3T3 cells. Toxicology Letters. 2006;163(1):44–53. doi: 10.1016/j.toxlet.2005.09.026. [DOI] [PubMed] [Google Scholar]
- 10.Chyau C-C, Ko P-T, Mau J-L. Antioxidant properties of aqueous extracts from Terminalia catappa leaves. LWT. 2006;39(10):1099–1108. [Google Scholar]
- 11.Masuda T, Yonemori S, Oyama Y, et al. Evaluation of the antioxidant activity of environmental plants: activity of the leaf extracts from seashore plants. Journal of Agricultural and Food Chemistry. 1999;47(4):1749–1754. doi: 10.1021/jf980864s. [DOI] [PubMed] [Google Scholar]
- 12.Lin C-C, Hsu Y-F, Lin T-C. Effects of punicalagin and punicalin on carrageenan-induced inflammation in rats. American Journal of Chinese Medicine. 1999;27(3-4):371–376. doi: 10.1142/S0192415X99000422. [DOI] [PubMed] [Google Scholar]
- 13.Fan YM, Xu LZ, Gao J, et al. Phytochemical and antiinflammatory studies on Terminalia catappa . Fitoterapia. 2004;75(3-4):253–260. doi: 10.1016/j.fitote.2003.11.007. [DOI] [PubMed] [Google Scholar]
- 14.Nagappa AN, Thakurdesai PA, Rao NV, Singh J. Antidiabetic activity of Terminalia catappa Linn fruits. Journal of Ethnopharmacology. 2003;88(1):45–50. doi: 10.1016/s0378-8741(03)00208-3. [DOI] [PubMed] [Google Scholar]
- 15.Chu S-C, Yang S-F, Liu S-J, Kuo W-H, Chang Y-Z, Hsieh Y-S. In vitro and in vivo antimetastatic effects of Terminalia catappa L. leaves on lung cancer cells. Food and Chemical Toxicology. 2007;45(7):1194–1201. doi: 10.1016/j.fct.2006.12.028. [DOI] [PubMed] [Google Scholar]
- 16.Pandya NB, Tigari P, Dupadahalli K, Kamurthy H, Nadendla RR. Antitumor and antioxidant status of Terminalia catappa against Ehrlich ascites carcinoma in Swiss albino mice. Indian Journal of Pharmacology. 45(5):464–469. doi: 10.4103/0253-7613.117754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ko T-F, Weng Y-M, Lin S-B, Chiou RY-Y. Antimutagenicity of supercritical CO2 extracts of Terminalia catappa leaves and cytotoxicity of the extracts to human hepatoma cells. Journal of Agricultural and Food Chemistry. 2003;51(12):3564–3567. doi: 10.1021/jf034102v. [DOI] [PubMed] [Google Scholar]
- 18.Mudi SY, Muhammad A. Antimalaria activity of ethanolic extracts of leaves of Terminalia catappa L. Combretaceae [indian almond] Bayero Journal of Pure and Applied Sciences. 2010;2:14–18. [Google Scholar]
- 19.Ratnasooriya WD, Dharmasiri MG, Rajapakse RAS, et al. Tender leaf extract of Terminalia catappa antinociceptive activity in rats. Pharmaceutical Biology. 2002;40(1):60–66. [Google Scholar]
- 20.Lin Y-L, Kuo Y-H, Shiao M-S, Chen C-C, Ou J-C. Flavonoid glycosides from Terminalia catappa L. Journal of the Chinese Chemical Society. 2000;47(1):253–256. [Google Scholar]
- 21.Chung K-T, Wong TY, Wei C-I, Huang Y-W, Lin Y. Tannins and human health: a review. Critical Reviews in Food Science and Nutrition. 1998;38(6):421–464. doi: 10.1080/10408699891274273. [DOI] [PubMed] [Google Scholar]
- 22.Orr I. Oral cancer in betel nut chewersin travancore, its etiology, pathology and treatment. The Lancet. 1933;222(5741):575–580. [Google Scholar]
- 23.Ranadive KJ, Ranadive SN, Shivapurkar NM, Gothoskar SV. Betel quid chewing and oral cancer: experimental studies on hamsters. International Journal of Cancer. 1979;24(6):835–843. doi: 10.1002/ijc.2910240623. [DOI] [PubMed] [Google Scholar]
- 24.Butler LG, Rogler JC. Biochemicalmechanisms of the antinutritional effectsof tannins. In: Ho C-T, Lee CY, Huang M-T, editors. Phenolic Compounds in Food and Their Effects on Health. I. chapter 23. Vol. 506. Washington, DC, USA: American Chemical Society; 1992. p. p. 298. (ACS Symposium Series). [Google Scholar]
- 25.Marzo F, Tosar A, Santidrian S. Effect of tannic acid on the immune response of growing chickens. Journal of Animal Science. 1990;68(10):3306–3312. doi: 10.2527/1990.68103306x. [DOI] [PubMed] [Google Scholar]
- 26.Tanaka T, Nonaka G-I, Nishioka I. Tannins and related compounds. XL. Revision of the structures of punicalin and punicalagin, and isolation and characterization of 2-O-galloylpunicalin from the bark of Punica granatum L. Chemical and Pharmaceutical Bulletin. 1986;34(2):650–655. [Google Scholar]
- 27.Cronquist A. An Integrated System of Classification of Flowering Plants. New York, NY, USA: Columbia University Press; 1988. [Google Scholar]
- 28.Resende FA, Alves JM, Munari CC, et al. Inhibition of doxorubicin-induced mutagenicity by Baccharis dracunculifolia . Mutation Research. 2007;634(1-2):112–118. doi: 10.1016/j.mrgentox.2007.06.008. [DOI] [PubMed] [Google Scholar]
- 29.Sforcin JM, Orsi RO, Bankova V. Effect of propolis, some isolated compounds and its source plant on antibody production. Journal of Ethnopharmacology. 2005;98(3):301–305. doi: 10.1016/j.jep.2005.01.042. [DOI] [PubMed] [Google Scholar]
- 30.Leitão DPDS, Da Silva Filho AA, Polizello ACM, Bastos JK, Spadaro ACC. Comparative evaluation of in-vitro effects of Brazilian green propolis and Baccharis dracunculifolia extracts on cariogenic factors of Streptococcus mutans . Biological & Pharmaceutical Bulletin. 2004;27(11):1834–1839. doi: 10.1248/bpb.27.1834. [DOI] [PubMed] [Google Scholar]
- 31.Chen P-S, Li J-H, Liu T-Y, Lin T-C. Folk medicine Terminalia catappa and its major tannin component, punicalagin, are effective against bleomycin-induced genotoxicity in Chinese hamster ovary cells. Cancer Letters. 2000;152(2):115–122. doi: 10.1016/s0304-3835(99)00395-x. [DOI] [PubMed] [Google Scholar]
- 32.Seeram N, Lee R, Hardy M, Heber D. Rapid large scale purification of ellagitannins from pomegranate husk, a by-product of the commercial juice industry. Separation and Purification Technology. 2005;41(1):49–55. [Google Scholar]
- 33.Kraszni M, Marosi A, Larive CK. NMR assignments and the acid-base characterization of the pomegranate ellagitannin punicalagin in the acidic pH-range. Analytical and Bioanalytical Chemistry. 2013;405(17):5807–5816. doi: 10.1007/s00216-013-6987-x. [DOI] [PubMed] [Google Scholar]
- 34.Robards K. Strategies for the determination of bioactive phenols in plants, fruit and vegetables. Journal of Chromatography A. 2003;1000(1-2):657–691. doi: 10.1016/s0021-9673(03)00058-x. [DOI] [PubMed] [Google Scholar]
- 35.Ribani M, Bottoli CBG, Collins CH, Jardim ICS, Melo LCF. Validação em métodos cromatográficos e eletroforéticos. Química Nova. 2004;27(5):771–780. [Google Scholar]
- 36.Qu W, Breksa AP, III, Pan Z, Ma H. Quantitative determination of major polyphenol constituents in pomegranate products. Food Chemistry. 2012;132(3):1585–1591. doi: 10.1016/j.foodchem.2011.11.106. [DOI] [PubMed] [Google Scholar]
- 37.Maron DM, Ames BN. Revised methods for the Salmonella mutagenicity test. Mutation Research. 1983;113(3-4):173–215. doi: 10.1016/0165-1161(83)90010-9. [DOI] [PubMed] [Google Scholar]
- 38.Bernstein L, Kaldor J, McCann J, Pike MC. An empirical approach to the statistical analysis of mutagenesis data from the Salmonella test. Mutation Research. 1982;97(4):267–281. doi: 10.1016/0165-1161(82)90026-7. [DOI] [PubMed] [Google Scholar]
- 39.Mortelmans K, Zeiger E. The Ames Salmonella/microsome mutagenicity assay. Mutation Research. 2000;455(1-2):29–60. doi: 10.1016/s0027-5107(00)00064-6. [DOI] [PubMed] [Google Scholar]
- 40.Machado TDB, Leal ICR, Amaral ACF, Dos Santos KRN, Da Silva MG, Kuster RM. Antimicrobial ellagitannin of Punica granatum fruits. Journal of the Brazilian Chemical Society. 2002;13(5):606–610. [Google Scholar]
- 41.Doig AJ, Williams DH, Oelrichs PB, Baczynskyj L. Isolation and structure elucidation of punicalagin, a toxic hydrolysable tannin, from Terminalia oblongata . Journal of the Chemical Society, Perkin Transactions 1. 1990;(8):2317–2321. [Google Scholar]
- 42.Bressolle F, Bromet-Petit M, Audran M. Validation of liquid chromatographic and gas chromatographic methods. Applications to pharmacokinetics. Journal of Chromatography B. 1996;686(1):3–10. doi: 10.1016/s0378-4347(96)00088-6. [DOI] [PubMed] [Google Scholar]
- 43.Ardrey RE. Liquid Chromatography-Mass Spectrometry: An Introduction. San Francisco, Calif, USA: John Wiley & Sons; 2003. [Google Scholar]
- 44.Snyder LR, Kirkland JJ, Glajch JL. Practical HPLC Method Development. 2nd edition. New York, NY, USA: John Wiley & Sons; 1997. [Google Scholar]
- 45.Gupta S, Saha B, Giri AK. Comparative antimutagenic and anticlastogenic effects of green tea and black tea: a review. Mutation Research. 2002;512(1):37–65. doi: 10.1016/s1383-5742(02)00024-8. [DOI] [PubMed] [Google Scholar]
- 46.Labieniec M, Gabryelak T. Response of DNA, proteins and membrane bilayer in the digestive gland cells of freshwater mussel Unio tumidus to tannins exposure. Toxicology in Vitro. 2004;18(6):773–781. doi: 10.1016/j.tiv.2004.03.005. [DOI] [PubMed] [Google Scholar]