

Abstract
Fungi native to agricultural soils that colonized commercially available biodegradable mulch (BDM) films were isolated and assessed for potential to degrade plastics. Typically, when formulations of plastics are known and a source of the feedstock is available, powdered plastic can be suspended in agar-based media and degradation determined by visualization of clearing zones. However, this approach poorly mimics in situ degradation of BDMs. First, BDMs are not dispersed as small particles throughout the soil matrix. Secondly, BDMs are not sold commercially as pure polymers, but rather as films containing additives (e.g. fillers, plasticizers and dyes) that may affect microbial growth. The procedures described herein were used for isolates acquired from soil-buried mulch films. Fungal isolates acquired from excavated BDMs were tested individually for growth on pieces of new, disinfested BDMs laid atop defined medium containing no carbon source except agar. Isolates that grew on BDMs were further tested in liquid medium where BDMs were the sole added carbon source. After approximately ten weeks, fungal colonization and BDM degradation were assessed by scanning electron microscopy. Isolates were identified via analysis of ribosomal RNA gene sequences. This report describes methods for fungal isolation, but bacteria also were isolated using these methods by substituting media appropriate for bacteria. Our methodology should prove useful for studies investigating breakdown of intact plastic films or products for which plastic feedstocks are either unknown or not available. However our approach does not provide a quantitative method for comparing rates of BDM degradation.
Keywords: Microbiology, Issue 75, Plant Biology, Environmental Sciences, Agricultural Sciences, Soil Science, Molecular Biology, Cellular Biology, Genetics, Mycology, Fungi, Bacteria, Microorganisms, Biodegradable plastic, biodegradable mulch, compostable plastic, compostable mulch, plastic degradation, composting, breakdown, soil, 18S ribosomal DNA, isolation, culture
Introduction
Degradation has historically been considered an undesirable attribute of plastic polymers, because breakdown shortens product life span and durability. Recently, awareness of the environmental problems presented by plastic waste in the natural environment1,2,3 has made biodegradable plastics an attractive alternative to conventional plastic materials. Degradation (defined as structural changes, fragmentation, and reduction in molecular weight, integrity, and strength4,5) occurs via a series of events, including both abiotic processes (thermal stress, photo-oxidation, hydrolysis, erosion and mechanical stress), and biological degradation6. While abiotic processes can change the fragment size and characteristics of plastics, microorganisms are required for their ultimate mineralization to water and carbon dioxide (in aerobic conditions) and/or methane (under anaerobic conditions).
A substantial niche for biodegradable plastics exists in agriculture, where plastic mulches are used to prevent weed growth, to retain soil moisture and to increase soil temperatures7,8 . Hundreds of thousands of acres in the United States alone are covered with plastic mulches9, including mulches composed of biodegradable plastic. Following a crop growing season, the options for disposing of biodegradable mulches (BDMs) include disposal in a landfill, incineration for energy recovery10, degradation via composting, or degradation in the soil after tillage11. Of these, the least labor-intensive fate is plowing BDMs into the soil, but without efficient degradation and mineralization during non-crop months (generally in the winter), plastic fragments could remain and interfere with agricultural equipment during spring tillage and planting, and persist in the environment where they significantly impact wildlife, plant life, and microbiota1,2,3,10.
Although many plastic products, including agricultural mulch films, bear the label "biodegradable" or "compostable", in practice, degradation and mineralization may be too inefficient and/or too incomplete for in-soil decomposition to be a viable alternative for disposal of these products. For example, oxo-biodegradable polyethylenes achieved only 12.4% mineralization after one year of weathering and three subsequent months in a 58 °C compost, and less than half that amount of mineralization occurred when the compost temperature was 25 °C12. In the winter, soil temperatures at most locations would be lower than either of these temperatures, presumably resulting in even lower microbial activity and consequently, less mineralization. In addition to slow degradation rates, misuse of the term "biodegradable" has led to distrust of these products by consumers13,14, including those in the agricultural industry. Biodegradation is the conversion of polymers to carbon dioxide (and/or methane) and water14 by naturally-occurring microorganisms4. Therefore, biodegradation must be measured chemically; the physical association of microorganisms with a substrate does not imply microbial degradation of that material.
As part of an effort to examine sustainable use of BDMs in agriculture, this study focused on discovering microorganisms native to agricultural soils that colonize and degrade commercially-available BDMs. Standard test methods have been published for chemically measuring the breakdown of biodegradable plastics by abiotic and biological means15,16,17. However, these methods do not address degradation of plastics by individual microbial species, or provide methods for their isolation. The methodology herein more closely resembles standard methods designed to evaluate plastics for resistance to microbial breakdown after inoculating specimens with fungal spores18,19.
When formulations of plastics are known and a source of the feedstock is available, powdered plastic can be suspended in agar-based media and degradation determined by visualization of clearing zones13. This method has been used previously to identify microorganisms that degrade polymers such as polyurethane20, poly-(butylene succinate-co-adipate)21, and poly(lactic acid)22. A similar method involves suspending pure powdered plastic in liquid medium where the plastic is the sole carbon source20,23. While these methods have the advantage of a defined system, they poorly mimic in situ degradation of BDMs. First, the surface area is distributed differently because BDMs are not dispersed in small particles throughout the soil matrix, but rather, sold and used as films. Second, the chemical makeup of BDMs is different from pure polymers. BDMs generally contain additives such as fillers, plasticizers, and colorants, and these additives may affect microbial growth and thereby, the rate of mineralization. For this reason, and because the composition of certain commercial films in this study were proprietary, plastic film in its field-ready form was utilized to isolate fungi and bacteria. For simplicity, the methods below are described only for fungi, with modifications noted where appropriate for bacterial isolations.
In a recent study24, three commercially-available BDMs and one experimental film were used at agricultural sites in three different regions of the United States for one growing season, and subsequently placed in mesh (250 micron) bags and buried for one winter in soil at the same sites. The 250 micron mesh openings allow fungal hyphae to penetrate while excluding roots and most soil fauna, and minimizing soil encroachment25,26. Nylon materials prevent bag degradation in soil. Following excavation, fungal isolates were recovered from BDM pieces and assessed for growth on minimal medium without a source of carbon except for the agar and a 5 cm x 5 cm surface-disinfested square of new, unused BDM film that was pre-disinfested. Most plastics used as films cannot be autoclaved without loss of integrity, so UV light was used to kill any microbial cells residing on the plastics. ISO 84619 recommends surface-disinfesting in 70% ethanol and subsequent drying, but if using this method, one must ensure that no component or additive of the film is adversely affected by the ethanol. Since BDMs presumably are manufactured to withstand sunlight, UV was chosen as a decontamination method.
Isolates that grew on BDM pieces better than on minimal medium alone were selected for further study. Agar, a polysaccharide produced by marine algae, is used to solidify microbial media because it is typically not utilized metabolically by agriculturally and medically notable microorganisms; however, agar-hydrolyzing enzymes have been isolated from marine bacteria27 and agar-hydrolyzing bacteria also have been isolated from soil28. BDM polymers and agar are both expected to be rare substrates for enzymes secreted by soil fungi, which have not evolved in environments that contain these polymers as potential nutrient sources, but both substrates are present in the plate bioassay described herein (Step 7). Fungi that use BDMs but not agar as a carbon source can be differentiated from fungi that use agar only, by comparing growth on agar-solidified medium containing i) no added carbon source except agar (negative control), ii) BDM films (experimental) and iii) glucose (positive control). Growth of all isolates is expected on minimal medium plus glucose; fungi not arising on glucose-containing plates may not be capable of growth on the particular minimal medium used in the experiment. Potential BDM degraders should grow on agar-solidified minimal medium + BDM film better than they grow on agar-solidified minimal medium alone. Fungi growing on minimal medium plates are agar-degraders or oligotrophs, and are also expected to grow on the agar associated with BDM films in bioassay plates, but not on the films themselves (unless they serendipitously also degrade BDM polymers).
To eliminate the possibility of seeing microbial growth due to utilization of agar and not of BDMs, we followed our initial assay for BDM colonization on agar plates with a bioassay in defined broth medium (Step 9). BDM pieces represented the only known carbon source in the bioassay tubes.
After the initial screening, and upon reviving glycerol stocks of the isolates, some formed scant but visible mycelia in liquid defined medium containing no known carbon source. These results suggested that some of the acquired isolates were oligotrophs - organisms that grow by scavenging very small amounts of carbon, nitrogen, and other nutrients dissolved either in the aqueous environment or existing as volatiles in the air29,30,31. Species identification via 18S ribosomal DNA analysis supported this view, as many of the isolates matched fungal genera previously reported to exhibit oligotrophy32. Oligotrophs, which are commonly saprophytes, require a broad range of metabolic capabilities for substrate utilization in a range of environments30. Thus, it is not surprising that the same fungi we isolated from BDMs (presumably requiring unusual enzymatic capabilities) demonstrated oligotrophic capacities, and were able to grow on trace contaminants such as skin oils from fingerprints, dust, or trace volatiles in the air. Due to the isolation of oligotrophs, we concluded that growth on a BDM surface alone could not be used to infer BDM breakdown. The methods described herein reflect our efforts to screen native BDM colonizers from agricultural soils for bona fide BDM breakdown.
Protocol
This procedure requires at least several months for incubation of BDM films in soil, and several more months for sequential bioassays both on agar plates and in agar-free, chemically defined broth to assess colonization and degradation. Individual methods are listed in the order they will be performed.
1. Incubation of BDM Films in Soil
Incorporate BDM films into soil under conditions mimicking those under which they will be expected to degrade. Acquire 400 g (dry weight equivalent) of resident soil and sandwich a 10 cm x 10 cm BDM with the soil inside a 13 x 13 cm nylon mesh (250 micron) bag. Close with nylon thread. Incubation times are at the experimenter's discretion. Monitor and/or modify parameters relevant to microbial activity (e.g. soil temperature, nutrients and moisture) at regular intervals throughout the incubation period as appropriate.
2. Preparation of Media and Reagents
To avoid introducing nutrient sources inadvertently into media and culture tubes, use reagent-grade chemicals, newly purchased culture tubes, Type I ultrapure (e.g. NanoPure) water, and stringently-washed glassware for mixing media. Avoid cross-contamination of reagents - it is best to use a dedicated set of reagents and glassware for this purpose. Note that it is possible that fungi may be isolated whose growth is inhibited by an ingredient in the reagents listed below. If that occurs, some optimization may be required.
Prior to making media, wash all necessary glassware with a laboratory detergent and then acid-wash (e.g soak overnight in 10% HCl). Rinse a minimum of twelve times in distilled water and twice in ultrapure water. Cover all glassware with clean aluminum foil to exclude dust. Wear gloves and avoid touching the inside and edges of vessels to exclude skin oils from fingerprints. To avoid any soap residues and scratches in glass which facilitate colonization, culture fungi in newly-purchased (unused), rinsed and autoclaved glass culture tubes and caps.
Potato dextrose agar (PDA) is used for the initial isolation of fungi from soil, and for establishment of single isolated colonies and generation of inoculum for long-term storage. Prepare the media according to the manufacturer's instructions and after autoclaving (when cooled to 50 °C), add chloramphenicol (from a 10 mg/ml stock solution in EtOH) to a final concentration of 30 μg/ml, to exclude bacteria. This medium is now referred to as PDA chl30.
Fungal minimal medium (FMM) is used as a solid base for BDMs inoculated with potential fungal plastic degraders. Aside from the agar (when used to solidify the medium), it is carbon-free. To approximately 800 ml of deionized H2O, add 6.0 g NaNO3, 0.52 g KCl, 0.52 g MgSO4·7H2O, 1.52 g KH2PO4, and 1 ml Hutner's trace elements stock solution (2.2 g ZnSO4·7H2O, 1.1 g H3BO3, 0.5 g MnCl2·4H2O, 0.5 g FeSO4·7H2O, 0.16 g CoCl2·5H2O, 0.16 g CuSO4·5H2O, 0.11 g (NH4)6Mo7O24·4H2O, and 5.0 g Na4 EDTA in 100 ml distilled H2O33,34. Adjust the pH to 6.5 using NaOH, and bring volume to 1 L. For the liquid bioassay, filter-sterilize FMM to avoid precipitation of salts. If solid medium is desired, add 16 g agar and autoclave. When semisolid agar is required, reduce agar to 8 g/L. When cooled to 50 °C, add chloramphenicol to a final concentration of 30 μg/ml. This medium is now referred to as FMM chl30. As a control to ensure that fungal isolates grow in this medium, FMM should be made as above but containing 10 g glucose per liter. This is called glucose minimal medium35 (GMM).
Phosphate-buffered saline36 (PBS) is used to suspend soil particles and microbial cells in the initial serial dilution of soil-exposed BDMs. To make PBS, add 8 g NaCl, 0.2 g KCl, 1.44 g Na2HPO4, and 0.24 g KH2PO4 to 800 ml of distilled water. Dissolve salts and adjust pH to 7.4 with HCl. Bring total volume to 1 L with ultrapure water. Fill clean culture tubes with 9.5 ml and 4.5 ml PBS per tube. Autoclave.
30% (v/v) glycerol is used to suspend cryopreserved bacterial and yeast cells, spores, and fungal mycelium during storage at -80 °C. Make this solution in ultrapure water. Autoclave.
0.01% (v/v) Triton X-100 is used to suspend fungal spores; the detergent acts as a non-toxic wetting agent for hydrophobic spores. Dissolve 100 μl Triton X-100 in 1 L of ultrapure water. Autoclave.
0.1 M sodium phosphate buffer (pH 7.2) is used as the solvent for glutaraldehyde during SEM fixation. Mix 68.4 ml of 1 M Na2HPO4 and 31.6 ml of 1 M NaH2PO4 and bring total volume to 1 L with ultrapure water36.
3. Preparation of Bioassay Materials: Surface Decontamination of BDM Films
Because remnants of contaminants like paper and tape on cutting utensils are potential carbon sources, use newly-purchased scissors or fresh razor blades, and an inorganic surface for cutting. Wear gloves to avoid contamination of BDM films with skin oils. Cut BDM films into small squares (4.25 x 4.25 cm). This size square fits into a standard 100 x 15 mm Petri plate.
Use a germicidal UV lamp (emission wavelength = 253.7 nm) to decontaminate BDM films. Such a lamp is often found in a standard biosafety cabinet. Ensure that when the UV light is operating, there is no flow of external air into the biosafety cabinet that might contaminate the BDM films.
Wet surfaces inside the hood with 70% EtOH and let blower run for 15 min to flush air. Then, close the sash and decontaminate the hood surface with UV for 2 hr.
Place the BDM films in the decontaminated hood, in rows. Allow the UV light to shine on films for 2 hr.
Use a clean, sterilized forceps to flip the BDM films. Start at the front row and work to the back row to minimize time that hands and arms are directly above the newly-decontaminated BDM surfaces. When flipping BDMs, place the newly-disinfested side downward on a clean area exposed to UV light but not previously in contact with the non-disinfested side of the film.
After 2 hr of UV treatment on each side, remove the BDM films from the biosafety cabinet with a sterile forceps, again working from front to back to avoid placing hands and arms over already-decontaminated films. Place the BDM films into sterile, dry, covered containers such as foil-covered beakers. Store at RT in darkness.
4. Plate Bioassay Setup
Perform all steps in a sterile transfer hood using aseptic technique.
Lay the UV-disinfested BDM pieces on the surface of agar plates. Set down one corner first, and gently let the remainder of the square roll into contact with the agar. Avoid wrinkles. Some films will form wrinkles as during storage; this is unavoidable. Particularly thin films may require the use of two forceps in order to lay the piece flat.
Place beads of semisolid FMM chl30 agar atop the plastic film to provide a water and nutrient source for initial colonization of hydrophobic plastics. Beads are not necessary for water-permeable films. Re-melt the agar in the microwave, and use a micropipettor to transfer four 10 μl drops of molten agar onto the mulch (one in each corner). Do not touch the tip of the pipette directly to the mulch as it may puncture. Do not exceed 10 μl per drop, because excess weight on the mulch can pull plastic off the agar surface when plates are inverted.
Let the plates sit right-side up with lids on overnight to solidify completely, and then store sealed sleeves agar-side up at 4 °C in darkness. When ready to perform the bioassay (Step 7), use i) one plate of FMM chl30 (negative control), ii) one bioassay plate containing four 10 μl drops of semisolid FMM chl30 on plastic film, and iii) one plate of GMM (positive control). In this way, four replicates of each isolate can be tested per plate of medium.
5. Liquid Bioassay Setup
Follow the instructions for BDM cutting in Step 3.1. Prior to cutting plastic films for the liquid bioassay, choose a size that fits into the culture tube. The area necessary to achieve a uniform mass among several plastic types will depend upon the film thickness. We found that pieces equivalent to 0.0175 g fit well in 15 x 150 mm culture tubes.
Surface-decontaminate films with UV light as described in Step 3 above.
For each replicate of a microbial isolate, prepare three tubes for inoculation: i) 5 ml FMM containing a BDM fragment of the predetermined size (experimental), ii) 5 ml FMM with no carbon source (negative control), and iii) 5 ml GMM (positive control). Additionally, prepare replicate tubes of FMM containing each BDM to be tested, but do not inoculate them. These control tubes will reveal any microbial growth resulting from contamination.
6. Isolation of Fungi
Following soil incubation and sample removal, fungi are isolated from the soil that adheres to the BDM films. If desired, bacteria can simultaneously be isolated with the same method using media appropriate for isolation of soil bacteria, such as 1/10X dilute tryptic soy yeast agar supplemented with 50 μg/ml cycloheximide to deter fungal growth37. When defined medium is required for bacterial isolations in Steps 5 and 7, M938 (plus cycloheximide) is a good choice.
Prior to sample processing, prepare PBS and PDA chl30. Allow PDA chl30 plates to dry (unbagged) at room temperature for approximately 24 hr to eliminate condensation on the lids and the agar surface. Pre-label PDA chl30 plates.
Excavate mesh bags containing BDM pieces from burial sites chosen in Step 1. Ship and store at 4 °C no longer than 48 hr after extracting from soil.
Using sterile spatula and forceps, gently remove soil until plastic is exposed but do not brush off soil that is clinging to BDM film surface. Cut BDM film into 1 cm2 pieces until 0.5 g of material is recovered for each of four total replications. Transfer 0.5 g BDM film and attached soil to 25 ml culture tubes containing 9.5 ml of PBS. If the mulch is completely degraded or a soil-only control is being processed, add 0.5 g of soil to the PBS; if possible, choose pieces that are still discolored from the residual mulch.
Vortex the culture tubes containing 9.5 ml PBS and 0.5 g mulch for 30 sec at high speed, sonicate in a sonicating waterbath for 10 min, and vortex again for 30 sec. The agitation is intended to break apart biofilms, physically remove cells embedded in or adhering to mulch pieces, and create a homogeneous suspension.
Prepare to serially dilute the PBS/soil solution to obtain individual colony forming units from platings. For each sample, place in a laminar flow hood one culture tube filled with 4.5 ml sterile PBS. For each sample, fill three wells of a deep-well 96-well plate filled with 450 μl PBS.
The original plastic/soil suspension (0.5 g mulch plus 9.5 ml of PBS solution) represents a 5.0 x 10-2 dilution of microbial material adhering to the plastic. Add 0.5 ml of suspension from this original tube to 4.5 ml of sterile PBS in a culture tube to obtain a 5.0 x 10-3 dilution. Vortex for 30 sec at high speed. Add 50 μl to 450 μl of PBS in the 96-well plate. Repeat for all samples/replicates. Use a multichannel pipettor to dilute samples en masse. Mix each new dilution by pipetting up and down ten times. Change tips between dilutions. Make dilutions up to 5.0 x 10-6.
(Optional) Bacterial isolates were more abundant than fungal isolates in samples described herein; thus if simultaneously isolating bacteria, then perform serial dilutions up to 5 x 10-8 in Step 6.6 above.
Evenly spread 100 μl from each of the 5.0 x 10-3 through the 5.0 x 10-6 dilutions across the agar surface of PDA chl30 plates with flame-sterilized bent glass rods. Store plates upright for 30 min to let liquid soak in, then seal plates with Parafilm to avoid escape of spores from unidentified isolates. Incubate inverted plates at 20 °C in darkness for 5 days to allow both quick and slow-growing fungi to form colonies.
7. Initial Selection of Plastic-degrading Fungi
Important: All cultures from this point forward should be opened only in a biosafety cabinet (not a laminar flow hood) to avoid contamination of the environment with spores of unknown identity. Some soil fungi and bacteria are potential human pathogens.
Examine dilution plates containing well-isolated colonies. Select a colony that is not touching another colony.
Using a single sterile toothpick, gently touch a colony and then touch the agar of FMM (made in Step 2.3), FMM + plastic (made in Step 4), and GMM (made in Step 2.3) plates, in that order. To inoculate agar beads atop plastic films, rub the toothpick gently on the surface of the agar dot and then swipe across the plastic film. Repeat the inoculation from the same source colony until there are four replicate streaks on each plate.
Repeat Steps 7.1 through 7.2 until several replicates of every visually-unique colony type are selected from every BDM sample.
Seal plates with Parafilm, invert, and incubate at 20 °C in the dark for 5 days.
Identify isolates that grow on BDMs better than they grow on FMM. If growth is not obvious, view the specimen under a dissecting microscope to verify colonization or lack thereof (Figure 1). Keep the lid on; if the lid has condensation, replace with a new lid but do so in the biosafety cabinet.
Once potential BDM degraders are selected, use a sterile toothpick to scrape inoculum from the fungus colonizing the plastic itself, and streak for isolation of single colony-forming-units on PDA chl30 plates.
Seal plates with Parafilm and incubate PDA chl30 plates inoculated with plastic-degrading isolates at 20 °C for 5 days in the dark.
Now that single isolated colonies are purified, ensure that they are truly able to colonize the plastic films being tested. Collect inoculum (spores, yeast cells, or hyphae) from a single colony-forming unit with a sterile toothpick. Repeat Steps 7.2 through 7.5. After 5 days of incubation at 20 °C, examine the bioassay plates for fungal growth. If the isolate grows on a BDM film better than it grows on FMM in both the first (7.5) and second (7.8) trials, then store the isolate as described below in Step 8.
8. Long-term Storage of Plastic Degraders
Overall, plastic-degrading isolates will be tested in the plate bioassay (7.5), re-purified to single isolated colony-forming units (7.6), tested in the plate bioassay again (7.8), and finally, tested in the liquid bioassay (9.1 - 9.5). During testing, isolates should be transferred to fresh media every month and working stock plates stored at 4 °C as soon as sufficient growth is visible. Isolates also should be stored as glycerol stocks at -80 °C, and/or as dried spores on sterile filter disks at 4 °C. Both storage methods are described below.
For each isolate that grew on a BDM film better than it grew on FMM in both the first (7.5) and second (7.8) plate bioassays, prepare two PDA chl30 plates with two or three sterilized filter paper disks, approximately 1 cm in diameter, on the surface. Inoculate these two plates from the same single colony chosen in Step 7.8 above. Make a lawn by spreading inoculum uniformly over the agar to maximize growth. Seal plates with Parafilm and incubate at 20 °C for 5 days. This step can be initiated simultaneously with Step 7.8 for efficiency.
The filter disk on the agar surface should be covered with mycelium/spores. Remove the filter disks from the agar surface with sterilized forceps, and place into a sterile Eppendorf tube. Air dry the filter disks overnight inside the Eppendorf tubes (with the cap off) in a biosafety cabinet or other dry, sterile environment with minimal air movement to avoid cross-contamination. Then seal with Parafilm, and store at 4 °C.
Use a sterile cotton swab or toothpick to harvest spores and mycelia from the lawn plates. Suspend spores and hyphae in 30% glycerol in cryovials, flash-freeze in liquid nitrogen, and store at -80 °C.
9. Stringent Confirmation of Plastic Utilization via Liquid Bioassay
Generate fresh spores and/or cells with which to inoculate liquid cultures: inoculate PDA chl30 plates with spores from potential plastic degrading isolates. Seal the plates with Parafilm, and incubate at 20 °C until fungal spores are visible.
In a biosafety cabinet, harvest spores by washing the plate with 0.01% Triton X-100 (5 ml per 15 x 100 mm Petri plate works well) and scrubbing the spores (or yeast cells) from the surface with a sterile bent-glass rod. Aspirate spore (or cell) suspension and transfer into a sterile container.
Count spores or cells with a hemacytometer and determine the appropriate volume to add to 5 ml of FMM broth for a total of 1 x 106 spores/tube. The volume should be less than 10 μl. Dilution of concentrated spore suspensions in 0.01% Triton X-100 may be necessary.
In a sterile transfer hood, inoculate culture tubes prepared in Step 5 with 1 x 106 spores (or yeast cells). Seal the caps with Parafilm to avoid any escape of spores, particularly for unidentified isolates. Incubate in darkness at 20 °C and observe samples weekly for growth.
Mycelial growth of fungi on plastic films may be visible within the first week after inoculation, especially at the edges of the plastic films. For planktonic yeast (as evidenced by cloudy media), growth can be monitored by optical density readings at 600 nm. Because fungi are likely to attach directly to the plastic film, growth may be assessed by eye, and can be confirmed by light microscopy and/or scanning electron microscopy (SEM).
In FMM-only controls, look for miniscule white flecks that are too small to register a change in optical density using spectrophotometric methods. These flecks can be aspirated with a Pasteur pipette and observed using differential interference (DIC) microscopy. Flecks are often unidentifiable precipitate, but in some cases they may be clumps of the original inoculant that had germinated but not grown substantially.
Use visual and microscopic observations to compare colonization of experimental samples and controls, and assign ratings to the growth on each sample of BDM film. Because many BDMs are dark in color and permit minimal observation via microscopy except at the edges, and film thickness prohibits observation under an oil-immersion lens, SEM is useful to estimate i) the presence of microbial growth, e.g. via the rating system described in Table 1, and ii) the extent of BDM degradation.
10. SEM Sample Preparation
Use clean, sterile scissors to cut small pieces of BDM from replicate samples. Fix BDMs by immersing in 2.5% glutaraldehyde in 0.1 M sodium phosphate buffer (pH 7.2) for 24-48 hr.
While fixation is occurring, dehydrate pure 100% ethanol by adding 5-10% volume 3 Å molecular sieve (activated in a drying oven at 175-260 °C and cooled in a desiccator), and allow to sit at room temperature overnight to ensure total dehydration.
Immerse BDMs in 0.1 M sodium phosphate buffer (pH 7.2) for four washes of 15 min each.
Immerse BDMs in ultrapure water for three washes of 5 min each.
Dehydrate BDM samples in an ethanol series, immersing for 20 min at each concentration: 50%, 70%, 80%, 90%, 95%, and 100%. Rinse BDMs twice more in 100% ethanol for 20 min per rinse.
Subject samples to critical point drying and sputter coating.
Representative Results
In a recent study24, four replicates each of three commercially-available BDMs labeled "biodegradable", plus an experimental film and a conventional plastic control, were placed over soil as mulch for tomato production in the spring of 2010 at Mount Vernon, WA, Knoxville, TN, and Lubbock, TX. In the fall of 2010, BDM film squares were cut from each weathered mulch in four replicate plots, and native soil was removed from directly beneath the area where the mulch sample had been excised. Each weathered BDM square was sandwiched in soil inside a nylon mesh bag as described in Step 1. A soil-only control was also included. Bags were buried under 7.5 cm of soil in the same plots and rows previously covered by the mulch, and left in place until spring 2011 tillage, seven months later. Bags were then lifted from the plots with a shovel, placed into Ziplock plastic bags, and shipped to the laboratory on ice packs for next-day delivery.
Here we report representative findings from our study. Native soil fungi were isolated from each BDM at all three sites, for a total of 54 isolates. We describe isolates from two of the commercial products used at one site (Mount Vernon). One of the products was a biodegradable plastic called BioAgri. This black film is labeled as "starch-based" (BioAgri, Palm Harbor, FL) and made from Mater-Bi bioplastic by Novamont (Terni, Italy). Mater-Bi is a blend of "corn starch, and biodegradable polymers obtained both from renewable raw materials and fossil raw materials" (http://www.novamont.com/default.asp?id=505). The other product was not plastic but rather a brown "paper-like material made from specially-engineered cellulose fiber" (WeedGuard Plus, SunShine Paper Co, Aurora, CO); (http://www.weedguardplus.com/faqs.php). This product originally was included as a control and degradation of its cellulosic components was expected.
Using the plate bioassay described herein, three potential BDM-degrading isolates from WeedGuard Plus and three from BioAgri were recovered. These isolates were subjected to the liquid bioassay (Steps 9.1 - 9.7 above) in triplicate. Although some fungal growth was visible along the edges of BDMs after one week in the liquid bioassay, samples were incubated for 68 days before harvesting and final observation. Using SEM, fungal hyphae were observed on all samples except XX and YY, where we observed rod-shaped cells (Table 2). Because XX and YY grow in yeast form on PDA chl30, this observation was expected.
For the WeedGuard Plus isolates, no growth was observed in the FMM-only controls. The most BDM degradation was observed in the samples colonized by Isolate SS. In the uninoculated controls for the WeedGuard Plus mulch (Figure 2A), residual tracheary elements of the plant-derived fibers are only detectable as slight pock-marking seen in rows on some of the fibers. Though fungal hyphae were observed in all three inoculated samples, tracheary elements were plainly visible only in the SS samples (Figure 2B), suggesting that digestion of cellulosic material revealed the lignified tracheary elements beneath. Isolate SS was tentatively identified using 18S ribosomal DNA as a sordariomycete within the order Sordariales.
For the BioAgri isolates, no significant growth was observed in the FMM-only controls. Single white specks visible upon swirling the culture tubes of isolates VV and ZZ were observed using DIC microscopy. For Isolate VV, the speck was a spore mass (not shown), presumably residual from the initial inoculation. For Isolate ZZ, the mass was composed of loose mesh of hyphae approximately 0.2 mm diameter, suggesting that the original spore inoculum had germinated, but had not grown beyond the mass pictured in Figure 3A.
In the uninoculated controls for the BioAgri mulch (Figure 3B), a bumpy texture is observed. The identity of the white features is unknown, but they are absent in a film colonized by Isolate VV (Figure 3C). Both the white particles and bumps are gone in a film colonized by Isolate ZZ (Figure 3D). In addition, the latter sample showed the uniform occurrence of fissures across the BDM surface. Isolates VV and ZZ were tentatively identified using 18S ribosomal DNA as Penicillium sp. (VV) and a sordariomycete within the order Hypocreales (ZZ), respectively.
| Score | Appearance of microbial growth |
| 0 | No growth apparent by eye or by microscopy |
| 1 | Germinated spores but no apparent subsequent growth |
| 2 | Growth covering ≤25% of test surface or medium |
| 3 | Growth covering ≤ 50% of test surface or medium |
| 4 | Growth covering ≤75% of test surface or medium |
| 5 | Growth covering >75% of test surface or medium |
Table 1. Scoring system for rating microbial growth on test samples of plastic films.
| Isolate | BDM | FMM-only control (score) | FMM containing 0.0175 g BDM(score and/or comments) | ||
| Visual assessment | Light microscopy | SEM | |||
| SS | WG | 0 | Hyphae and BDM fibers indistinguishable | Hyphae and BDM fibers indistinguishable | 2;Fungal hyphae visible |
| TT | WG | 0 | Hyphae and BDM fibers indistinguishable | Hyphae and BDM fibers indistinguishable | 2; Fungal hyphae visible |
| XX | WG | 0 | Broth cloudy | Few cells observed in broth | 2; Rod-shaped cells on surface |
| VV | BA | 1 | Fine mycelia visible at edge | Mycelia and spore chains observed at edge | 4; Prolific hyphae and conidiophores |
| YY | BA | 0 | Broth cloudy | Few cells observed in broth | 2; Rod-shaped cells on surface |
| ZZ | BA | 1 | Luxurious mycelial growth on entire surface | Hyphae visible at edge | 5; Prolific hyphae |
Table 2. Observations and growth ratings for fungal isolates obtained from weathered BDM films buried in field plots from fall 2010 until spring 2011 (7 months) at Mount Vernon, later inoculated onto unweathered BDM material and incubated for 68 days at 20 °C in darkness. WG = WeedGuard Plus; BA = BioAgri. See Table 1 for growth rating scheme.
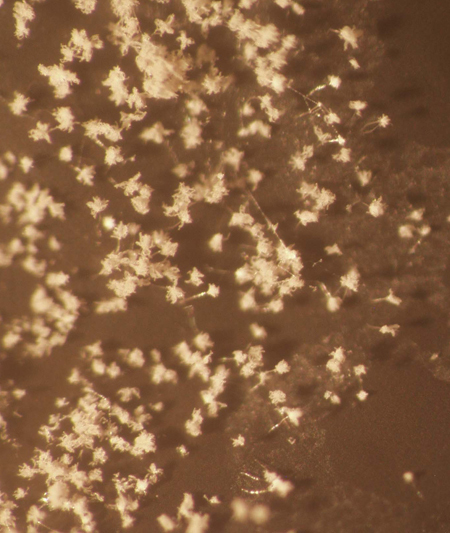 Figure 1. Colonization of BioAgri mulch in the plate bioassay described in Step 7, as viewed under a dissecting microscope at 20X magnification.
Figure 1. Colonization of BioAgri mulch in the plate bioassay described in Step 7, as viewed under a dissecting microscope at 20X magnification.
 Figure 2. Appearance of WeedGuard Plus mulch after 68 days in FMM but not inoculated (A), or inoculated with isolate ZZ, tentatively identified as Chaetomium sp. (B).Note the tracheary elements (arrows) that are barely distinguishable in A, but clearly seen in B. No growth was visible in the FMM-only control for the Chaetomium isolate. Samples were coated with Au/Pd using a Quorum SC7640 sputter coater, and imaged at an accelerating voltage of 15 kV on a Tescan VEGA 5136 MM SEM.
Figure 2. Appearance of WeedGuard Plus mulch after 68 days in FMM but not inoculated (A), or inoculated with isolate ZZ, tentatively identified as Chaetomium sp. (B).Note the tracheary elements (arrows) that are barely distinguishable in A, but clearly seen in B. No growth was visible in the FMM-only control for the Chaetomium isolate. Samples were coated with Au/Pd using a Quorum SC7640 sputter coater, and imaged at an accelerating voltage of 15 kV on a Tescan VEGA 5136 MM SEM.
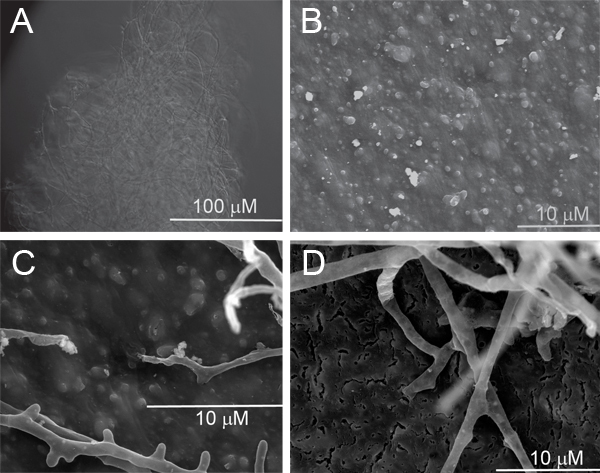 Figure 3. Appearance of BioAgri mulch after 68 days in the liquid bioassay. A single clump of germinated spores was observed in the FMM-only control for the ZZ isolate, tentatively identified as a Sordariomycete
(A). BioAgri mulch incubated in uninoculated FMM (B), BioAgri mulch inoculated with isolate VV, tentatively identified as Penicillium sp. (C), and BioAgri mulch incubated with isolate ZZ (D). Samples were coated with Au/Pd using a Quorum SC7640 sputter coater, and imaged at an accelerating voltage of 15 kV on a Tescan VEGA 5136 MM SEM.
Figure 3. Appearance of BioAgri mulch after 68 days in the liquid bioassay. A single clump of germinated spores was observed in the FMM-only control for the ZZ isolate, tentatively identified as a Sordariomycete
(A). BioAgri mulch incubated in uninoculated FMM (B), BioAgri mulch inoculated with isolate VV, tentatively identified as Penicillium sp. (C), and BioAgri mulch incubated with isolate ZZ (D). Samples were coated with Au/Pd using a Quorum SC7640 sputter coater, and imaged at an accelerating voltage of 15 kV on a Tescan VEGA 5136 MM SEM.
Discussion
The procedure described herein represents a first-pass technique for isolating potential BDM degraders from soil, and was successfully used to isolate fungi from BDMs buried in soil for seven months. Fungi grew when reinoculated onto fresh BDM material of the same type, indicating that the isolated fungi were indeed colonizers, and that the films were not inhibitory to fungal growth. Isolation of plastic-degrading fungi and bacteria potentially could lead to their use, individually or in combinations, for amendments to soil or compost where plastics need to be degraded.
In their native soil environment, microbial species do not exist in isolation. Plastic degraders and even oligotrophic organisms growing on BDMs, but using other carbon and energy sources, could be keystone species in the colonization of these recalcitrant nutrient sources. In an effort to evaluate BDM degradation in vitro but better mimic a real soil microbial community, the methodology described herein could be altered for testing of mixtures, rather than pure isolates, of fungal and bacterial inocula.
Of note, isolates TT, XX, VV, and YY achieved minimal degradation of the BDMs, but each showed visible BDM colonization in both plate and liquid bioassays. Thus, visible colonization did not necessarily signify BDM polymer degradation. Isolates TT, XX, VV, and YY are likely to represent oligotrophs that obtained nutrients from the environment as well as from the BDM29,30,31.
Definitive evidence that fungi are using a particular BDM as a carbon source requires measurement of degradation via chemical methods. Carbon dioxide evolution during incubation of plastics with microogranisms indicates respiration, and can be used as an indirect measure of polymer breakdown15,16,39. Various methods are used to gain information about the size and structure of polymers, including high performance liquid chromatography, size exclusion chromatography, differential scanning calorimetry, thermal gravimetric analysis, nuclear magnetic resonance spectroscopy, X-ray diffraction, and Fourier transform infrared spectroscopy. Studies to assess CO2 evolution and polymer breakdown would represent a logical progression from the screening procedure described herein. However, chemical methods for measuring polymer sizes cannot differentiate enzymatic degradation from hydrolysis by organic acids secreted during growth by fungi and bacteria. Implication of an enzymatic mechanism in plastic breakdown would require isolation of a pure enzyme(s) capable of degrading polymers in vitro.
The procedures described herein should be broadly applicable for any plastic that is used in a film conformation, such as food packaging material or garbage bags. These procedures have several advantages. First, the plastic films used in the bioassay are in their commercial form, as available for purchase in the marketplace and eventual degradation in the environment. A caveat is that used plastics, like the BDMs harvested after the 2010 growing season in this study, may be more weathered than new plastic films used in the in vitro bioassays. Prior microbial attack, UV, wind, abrasion by soil particulates, chemical hydrolysis via inorganic soil components, and soil fauna all contribute to oxidation and fragmentation of polymers, altering the efficiency of enzymatic action on BDM films11. A second advantage is that this method can be employed whether or not the individual constituents of proprietary plastic products are known and/or available in pure form for testing. However, a limitation of using plastics with multiple or unknown constituents is that visible degradation cannot be definitely assigned to a single constituent. For example, the degradation in Figure 3D could represent starch breakdown without degradation of the more recalcitrant polymers in the "starch-based" BioAgri film. In a previous study on in-soil degradation of starch-amended plastic films, carbon dioxide evolution rates reflected the amounts of starch and dye added to films, while unchanged polymer molecular weights supported the hypothesis that additives, not polymers, were being utilized by microorganisms39. Finally, this procedure is focused on in vitro degradation, thus providing a more defined system than those currently in standard use15,16. It is expected that samples degrading in chemically defined medium will prove easier to assess via chemical methods than samples degrading in soil or compost. Such assessments are the current focus of this work.
This procedure was successful for isolation of BDM-degrading fungi and bacteria from four types of BDM films at three locations within the United States (TN, WA, and TX), with different weather, soil types, and microbial community structures. This procedure takes a reductionist view, focusing on individual isolates with the capacity for growth upon, and breakdown of BDMs. However, the results emphasize the importance of microbial community analysis to describe the separate and linked roles of oligotrophic colonizers, bona fide BDM depolymerizers, and other community members, all of which are expected to be important in the ultimate goal of complete degradation of plastic films to biomass, carbon dioxide and/or methane, and water.
Disclosures
The authors declare that they have no competing financial interests.
Acknowledgments
Dr. Stephen Alderman, Dr. David Leaf, and Erin Macri are gratefully acknowledged for help with microscopy. This research was funded through a grant from the NIFA Specialty Crops Research Initiative, USDA SCRI-SREP Grant Award No. 2009-02484. Briana Kinash, Kevin Kinloch, Megan Leonhard Joseph McCollum, Maria McSharry and Nicole Sallee provided excellent technical assistance and thoughtful discussions.
References
- Gregory MR. Environmental implications of plastic debris in marine settings - entanglement, ingestion, smothering, hangers-on, hitch-hiking and alien invasions. Philosophical Transactions of the Royal Society. 2009;364:2013–2025. doi: 10.1098/rstb.2008.0265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Teuten EL, Saquing JM, et al. Transport and release of chemicals from plastics to the environment and to wildlife. Philosophical Transactions of the Royal Society. 2009;364:2027–2045. doi: 10.1098/rstb.2008.0284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson RC, Moore CJ, vom Saal FS, Swan SH. Plastics the environment and human health: current consensus and future trends. Philosophical Transactions of the Royal Society. 2009;364:2153–2166. doi: 10.1098/rstb.2009.0053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ASTM D 883. Standard terminology relating to plastics. West Conshohocken, PA: American Society for Testing and Materials International; 1991. [Google Scholar]
- SO 472. Plastics - vocabulary, amendment 3. General terms and terms relating to degradable plastics. Geneva, Switzerland: International Organization for Standardization; 1993. [Google Scholar]
- Krzan A, Hemjinda S, Miertus S, Corti A, Chiellini E. Standardization and certification in the area of environmentally degradable plastics. Polymer Degradation and Stability. 2006;91:2819–2833. [Google Scholar]
- Shogren RL. Biodegradable mulches from renewable resources. Journal of Sustainable Agriculture. 2000;16:33–47. [Google Scholar]
- Takakura T, Fang W. Climate under cover. Kluwer Academic Publishers; 2001. pp. 1–10. [Google Scholar]
- Miles C, Hayes D, Brodhagen M, Lee J, Wszelaki A, Moore-Kucera J, Wallace R, Marsh T, Inglis D. Plastic mulches, biodegradable alternatives, China and US. In: van Steenbergen F, Tuinhof A, Knoop L, editors. Transforming Landscapes, Transforming Lives: The Business of Sustainable Water Buffer Management. Wageningen, The Netherlands: 3R Water Secretariat; 2011. [Google Scholar]
- Song JH, Murphy RJ, Narayan R, Davies GBH. Biodegradable and compostable alternatives to conventional plastics. Transactions of the Royal Society B. 2009;364:2127–2139. doi: 10.1098/rstb.2008.0289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayes DG, Dharmalingam S, Wadsworth LC, Leonas KK, Miles C, Inglis DA. Biodegradable agricultural mulches derived from biopolymers. In: Khemani KC, Scholz C, editors. Degradable polymers and materials, principles and practice. ACS Symposium Series. 2nd Ed. Vol. 1114. Washington, D.C: American Chemical Society Press; 2012. pp. 201–223. [Google Scholar]
- Ojeda TFM, Dalmolin E, Forte MMC, Jacques RJS, Bento FM, Camargo FAO. Abiotic and biotic degradation of oxo-biodegradable polyethylenes. Polymer Degradation and Stability 94. 2009. pp. 965–970.
- van der Zee M. Analytical methods for monitoring biodegradation processes of environmentally degradable polymers. In: Lendlein A, Sisson A, editors. Handbook of Biodegradable Polymers. Weinheim, Germany: Wiley-VCH; 2011. pp. 263–281. [Google Scholar]
- Narayan R. Misleading claims and misuse of standards continues to proliferate in the nascent bioplastics industry space. BioPlastics. 2010;01/10 [Google Scholar]
- ASTM D 5338-98. Annual Book of ASTM Standards. West Conshohocken, PA: American Society for Testing and Materials International; 1998. Standard test method for determining aerobic biodegradation of plastic materials under controlled composting conditions; pp. 504–509. [Google Scholar]
- ASTM D 5988-03. Annual Book of ASTM Standards. West Conshohocken, PA: American Society for Testing and Materials International; 2003. Standard test method for determining aerobic biodegradation in soil of plastic materials or residual plastic materials after composting; pp. 354–358. [Google Scholar]
- ASTM D 6954-04. Annual Book of ASTM Standards. West Conshohocken, PA: American Society for Testing and Materials International; 2004. Standard guide for exposing and testing plastics that degrade in the environment by a combination of oxidation and biodegradation; pp. 748–753. [Google Scholar]
- ASTM G21-96. Annual Book of ASTM Standards. West Conshohocken, PA: American Society for Testing and Materials International; 2002. Standard practice for determining resistance of synthetic polymeric materials to fungi; pp. 433–437. [Google Scholar]
- ISO 846. Plastics - evaluation of the action of microorganisms. Geneva, Switzerland: International Organization for Standardization; 1997. pp. 1–22. [Google Scholar]
- Russell JR, Huang J, et al. Biodegradation of polyester polyurethane by endophytic fungi. Applied and Environmental Microbiology. 2011;77:6076–6084. doi: 10.1128/AEM.00521-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maeda H, Yamagata Y, Abe K, Hasegawa F, Machida M, Ishioka R, Gomi K, Nakajima T. Purification and characterization of a biodegradable plastic-degrading enzyme from Aspergillus oryzae. Applied Microbiology and Biotechnology. 2005;67:778–788. doi: 10.1007/s00253-004-1853-6. [DOI] [PubMed] [Google Scholar]
- Tokiwa Y, Calabia BP. Biodegradability and biodegradation of poly(lactide. Applied Microbiology and Biotechnology. 2006;72:244–251. doi: 10.1007/s00253-006-0488-1. [DOI] [PubMed] [Google Scholar]
- Karjomaa S, Suortti T, Lempiäinen R, Selin J-F, Itävaara M. Microbial degradation of poly-(L-lactic acid) oligomers. Polymer Degradation and Stability. 1998;59:333–336. [Google Scholar]
- Miles C, Wallace R, et al. Deterioration of potentially biodegradable alternatives to black plastic mulch in three tomato production regions. HortScience. 2012;47(9):1270–1277. [Google Scholar]
- Hedh J, Wallander H, Erland S. Ectomycorrhizal mycelial species composition in apatite amended and non-amended mesh bags buried in a phosphorus-poor spruce forest. Mycological Research. 2008;112:681–688. doi: 10.1016/j.mycres.2007.11.008. [DOI] [PubMed] [Google Scholar]
- Wallander H, Hagerberg D. Do ectomycorrhizal fungi have a significant role inweathering of minerals in forest soil?. 4th International Symbiosis Congress; Halifax, Canada. 2003. [Google Scholar]
- Hehemann J-H, Correc G, et al. Biochemical and structural characterization of the complex agarolytic enzyme system from the marine bacterium Zobellia galactanivorans. Journal of Biological Chemistry. 2012;287:30571–30584. doi: 10.1074/jbc.M112.377184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stanier RY. Studies on marine agar-digesting bacteria. Journal of Bacteriology. 1941;42(4):527–559. doi: 10.1128/jb.42.4.527-559.1941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hirsch P. Microbial life at extremely low nutrient levels. Advances in Space Research. 1986;6:287–298. doi: 10.1016/0273-1177(86)90097-9. [DOI] [PubMed] [Google Scholar]
- Wainwright M, Adam T, Barakah F. A review of the role of oligotrophic micro-organisms in biodeterioration. International Biodeterioration and Biodegradation. 1993;31:1–13. [Google Scholar]
- Wainwright M, Barakah R, Al-Turk I, Ali TA. Oligotrophic micro-organisms in industry, medicine, and the environment. Science Progress. 1991;75:313–322. [PubMed] [Google Scholar]
- Parkinson SM, Wainwright M, Killham K. Observations on oligotrophic growth of fungi on silica gel. Mycological Research. 1989;93(4):529–534. [Google Scholar]
- Hill T, Kafer E. Improved protocols for Aspergillus minimal medium: trace element and minimal medium salt stock solutions. Fungal Genetics Newsletter. 2001;48:20–21. [Google Scholar]
- Hutner SH, Provasoli L, Schatz A, Haskins CP. Some approaches to the study of the role of metals in the metabolism of microorganisms. Proceedings of the American Philosophical Society. 1950;94:152–170. [Google Scholar]
- Affeldt KJ, Brodhagen M, Keller NP. Aspergillus oxylipin signaling and quorum sensing pathways depend on G protein-coupled receptors. Toxins. 2012;4:695–6171. doi: 10.3390/toxins4090695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sambrook J, Russell DW. Molecular cloning: a laboratory manual. 3rd edition. Cold Spring Harbor, New York, NY: Cold Spring Harbor Laboratories Press; 2001. [Google Scholar]
- Marzluf GA. Physiology, metabolism, and molecular aspects of filamentous fungi. In: Reddy CA, Beveridge TJ, Breznak JA, Marzluf GA, Schmidt TM, Snyder LR, editors. Methods for General and Molecular Microbiology. Washington, D.C: ASM Press; 2007. pp. 952–964. [Google Scholar]
- Peters JE. Gene transfer in Gram-negative bacteria. In: Reddy CA, Beveridge TJ, Breznak JA, Marzluf GA, Schmidt TM, Snyder LR, editors. Methods for General and Molecular Microbiology. Washington, D.C: ASM Press; 2007. pp. 735–755. [Google Scholar]
- Yabannavar AV, Bartha R. Methods for assessment of biodegradability of plastic films in soil. Applied and Environmental Microbiology. 1994;60(10):3608–3614. doi: 10.1128/aem.60.10.3608-3614.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]