

This work presents a systematic analysis of nonessential plastid ribosomal proteins and reports that the combined deletion of two ribosomal protein genes (rps15 and rpl33) that individually can be deleted without causing a mutant phenotype results in synthetic lethality and loss of autotrophic growth.
Abstract
Consistent with their origin from cyanobacteria, plastids (chloroplasts) perform protein biosynthesis on bacterial-type 70S ribosomes. The plastid genomes of seed plants contain a conserved set of ribosomal protein genes. Three of these have proven to be nonessential for translation and, thus, for cellular viability: rps15, rpl33, and rpl36. To help define the minimum ribosome, here, we examined whether more than one of these nonessential plastid ribosomal proteins can be removed from the 70S ribosome. To that end, we constructed all possible double knockouts for the S15, L33, and L36 ribosomal proteins by stable transformation of the tobacco (Nicotiana tabacum) plastid genome. We find that, although S15 and L33 function in different ribosomal particles (30S and 50S, respectively), their combined deletion from the plastid genome results in synthetic lethality under autotrophic conditions. Interestingly, the lethality can be overcome by growth under elevated temperatures due to an improved efficiency of plastid ribosome biogenesis. Our results reveal functional interactions between protein and RNA components of the 70S ribosome and uncover the interdependence of the biogenesis of the two ribosomal subunits. In addition, our findings suggest that defining a minimal set of plastid genes may prove more complex than generally believed.
INTRODUCTION
Translation in bacteria occurs on 70S ribosomes that are composed of a large (50S) and a small (30S) subunit. The two DNA-containing organelles of eukaryotes (mitochondria and plastids) also possess 70S ribosomes that resemble bacterial ribosomes, in structure and molecular composition (Graack and Wittmann-Liebold, 1998; Yamaguchi et al., 2000; Yamaguchi and Subramanian, 2000; Manuell et al., 2007). Most of the components of the bacterial ribosome have remained remarkably well conserved over more than a billion years of evolution since the formation of eukaryotic cells through endosymbiosis. Consistent with their ancestry, mitochondrial ribosomes are most similar to ribosomes of proteobacteria, whereas plastid (chloroplast) ribosomes are most similar to cyanobacterial ribosomes (Graack and Wittmann-Liebold, 1998; Manuell et al., 2007).
The organellar genomes encode only a subset of the constituents of the 70S ribosomes of plastids and mitochondria. For example, the 30S subunit of the plastid ribosome (30S subunit) is composed of the 16S rRNA and 21 protein subunits (Rps proteins) that have orthologs in Escherichia coli. In seed plants, the plastid genome (plastome) encodes the 16S rRNA and 12 of the 21 ribosomal protein subunits, and the nuclear genome encodes the remaining nine subunits (Yamaguchi et al., 2000). The 50S subunit consists of three RNA species (23S rRNA, 5S rRNA, and 4.5S rRNA) and 31 ribosomal proteins (Rpl proteins) that have orthologs in E. coli. The plastome encodes the three rRNAs and nine of the 31 ribosomal proteins, and the nuclear genome encodes the remaining 22 ribosomal proteins (Yamaguchi and Subramanian, 2000). The nuclear encoding of organellar ribosomal proteins results from endosymbiotic gene transfer, an evolutionary process that gradually moved organellar genes into the nuclear genome and, in this way, resulted in a dramatic reduction of plastid and mitochondrial genome sizes (Gray, 1993; Timmis et al., 2004; Bock and Timmis, 2008). Chloroplast ribosomes also contain a few proteins that are not found in bacterial ribosomes (so-called plastid-specific ribosomal proteins) and may have been newly acquired during organelle evolution. Plastid-specific ribosomal proteins are encoded by nuclear genes and, while some of them appear to be bona fide ribosomal proteins that function in protein biosynthesis, others can be removed without causing obvious translational defects (Tiller et al., 2012).
In all plants, autotrophic growth requires plastid protein biosynthesis because the plastid genome encodes many of the components of the photosynthetic apparatus. In most plant species, heterotrophic growth also requires plastid protein biosynthesis (Ahlert et al., 2003; Rogalski et al., 2006; Alkatib et al., 2012a), presumably because a few plastid-encoded proteins affect metabolic functions that are essential for cell survival even in the presence of an exogenous carbon source (Kode et al., 2005).
Not all components of the 70S ribosome are essential for translation; in both bacteria and plastids, several ribosomal proteins have been identified as nonessential (Baba et al., 2006; Rogalski et al., 2008; Fleischmann et al., 2011). The genes for the Rps15 protein (S15) of the small ribosomal subunit and the Rpl33 (L33) and Rpl36 (L36) proteins of the large ribosomal subunit can be inactivated in E. coli (Maguire and Wild, 1997; Ikegami et al., 2005; Ikegami et al., 2005) and in plastids (Rogalski et al., 2008; Fleischmann et al., 2011) without losing viability. While the rpl36 gene knockout in tobacco (Nicotiana tabacum) plastids displays a severe mutant phenotype, rpl33 and rps15 knockout plants are nearly indistinguishable from the wild type (Rogalski et al., 2008; Fleischmann et al., 2011).
Nonessentiality of two genes does not necessarily imply that the combined knockout of both genes will also be viable. The phenomenon that the combined removal of two nonessential genes results in loss of viability is known as synthetic lethality. Synthetic lethality can occur if two genes are functionally redundant or have partially overlapping functions. Identifying and reliably predicting synthetic lethal interactions between nonessential genes represents one of the biggest challenges in top-down synthetic genomics, an area of synthetic biology that aims at designing, synthesizing, and booting up a functional minimum-size genome (Delaye and Moya, 2010; Klasson and Andersson, 2010; Cambray et al., 2011). Here, we have undertaken a systematic investigation of the 70S ribosome for synthetic lethality. Using stable chloroplast genome transformation in the model plant tobacco, we generated double knockout lines for the three nonessential ribosomal protein genes encoded in the plastid genome. We find that, although S15 and L33 are nonessential ribosomal proteins that function in distinct ribosomal subunits, their combined deletion from the plastid genome surprisingly results in synthetic lethality under autotrophic conditions. Interestingly, the lethality can be rescued by growth under elevated temperatures, which improves the efficiency of ribosome biogenesis in the double mutants.
RESULTS
Construction of Transplastomic Double Knockout Plants for Three Nonessential Ribosomal Protein Genes
The development of technologies for the stable genetic transformation of plastid genomes (Boynton et al., 1988; Svab and Maliga, 1993) has facilitated the functional analysis of genes in the plastid genome by reverse genetics (Rochaix, 1997; Maliga, 2004). Taking advantage of the active homologous recombination system in plastids, genes can be knocked out by disrupting or replacing their coding region with the standard selectable marker gene for chloroplast transformation, the aadA gene, which confers spectinomycin resistance (e.g., Takahashi et al., 1991; Hager et al., 1999; Krech et al., 2012). The construction of double knockout lines that inactivate two unlinked genes remains more technically challenging because it requires either recycling of the aadA marker, cotransformation with two plasmids, or sequential transformation using two different selection markers (supertransformation). Although aadA is the only marker gene that is routinely used in plastid transformation experiments, a few alternative selectable marker genes have become available (Carrer et al., 1993; Huang et al., 2002; Li et al., 2011), enabling us to employ a supertransformation approach to analyze the effects of the combined inactivation of two nonessential plastid ribosomal protein genes. To this end, we used the aphA-6 marker gene, which confers kanamycin resistance (Huang et al., 2002) to disrupt a second ribosomal protein gene in the rps15, rpl33, and rpl36 knockout plants that had previously been generated with the aadA marker (Rogalski et al., 2008; Fleischmann et al., 2011). Supertransformation of rpl33 knockout plants with an aphA-6–based knockout construct for rps15 (Figures 1A and 1B) generated an rps15/rpl33 double knockout (subsequently referred to as Δrps15/Δrpl33 plants). Similarly, supertransformation of the rpl33 knockout with a knockout construct for rpl36 (Figures 1A and 1C) produced an rpl33/rpl36 double knockout (Δrpl33/Δrpl36 lines), and supertransformation of the rps15 knockout with a knockout construct for rpl36 (Figures 1B and 1C) resulted in an rps15/rpl36 double knockout (Δrps15/Δrpl36 plants). In this way, we constructed all three possible double knockout combinations of nonessential plastid-encoded ribosomal protein genes.
Figure 1.
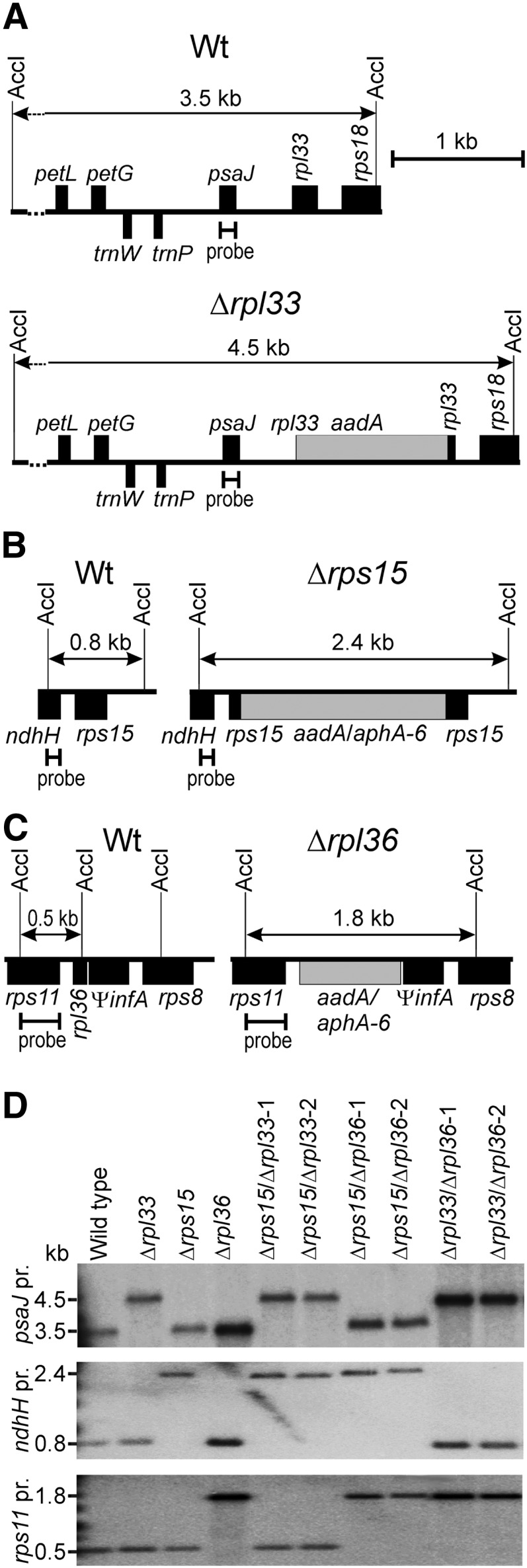
Construction of Transplastomic Double Knockout Plants for the Nonessential Ribosomal Protein Genes rps15, rpl33, and rpl36.
(A) Physical map of the region in the tobacco plastid genome (Shinozaki et al., 1986) containing the rpl33 gene and map of the transformed plastid genome (transplastome) in Δrpl33 mutant plants (Rogalski et al., 2008). Genes above the line are transcribed from the left to the right, and genes below the line are transcribed in the opposite direction. The AccI restriction sites employed for RFLP analyses are indicated, and the resulting fragment sizes are given for the wild type and the rpl33 knockout. The hybridization probe used for DNA gel blot analysis (derived from the psaJ coding region) is also indicated.
(B) Map of the genomic region containing rps15 and map of the transformed plastid genome in Δrps15 mutants (Fleischmann et al., 2011). In the Δrps15 single mutant and the Δrps15/Δrpl36 double knockout, the spectinomycin resistance marker aadA (Svab and Maliga, 1993) disrupts the rps15 gene, whereas in the Δrps15/Δrpl33 plants, rps15 is inactivated with the kanamycin resistance gene aphA-6 (Huang et al., 2002).
(C) Map of the genomic region containing rpl36 and map of the transformed plastid genome in Δrpl36 mutants (Fleischmann et al., 2011). In the Δrpl36 single mutant, the aadA marker disrupts the rpl36 gene, whereas in the Δrps15/Δrpl36 and the Δrpl33/Δrpl36 double knockout plants, rpl36 is inactivated with the aphA-6 marker.
(D) RFLP analysis of plastid transformants. The wild type, the three single mutants, and the three double knockouts (two independently generated transplastomic lines each) were analyzed by DNA gel blotting using the restriction enzyme AccI and specific radiolabeled probes (pr.) for each of the three knockout alleles (cf. panels [A] to [C]). Note that all transplastomic lines included in this blot are homoplasmic and show exclusively the bands diagnostic of the transgenic plastid genomes.
The knockout alleles were introduced into the tobacco chloroplast genome by biolistic transformation to replace the corresponding wild-type alleles by homologous recombination (Figure 1). Selection on kanamycin-containing regeneration medium produced multiple independent antibiotic-resistant lines for all three supertransformation constructs. Primary doubly transplastomic candidate lines were subjected to two to four additional rounds of regeneration and selection to enrich plastid genome copies containing the second (aphA-6-based) knockout allele and dilute out residual genome copies that harbor only the first (aadA-based) gene knockout. Successful transformation of the plastid genome was confirmed by restriction fragment length polymorphism (RFLP) analyses of DNA samples from doubly transplastomic candidate lines. In all cases, the double knockout lines displayed the expected hybridizing fragments (Figure 1D). Moreover, absence of a residual signal for the wild type–specific restriction fragment strongly suggested homoplasmy of the generated transplastomic lines.
Analysis of Transplastomic Double Knockout Lines of Nonessential Ribosomal Protein Genes
Homoplasmy of the double knockout lines (Figure 1D) suggests their viability under heterotrophic growth conditions in vitro (i.e., on synthetic medium supplemented with Suc as an exogenous source of reduced carbon). This was further confirmed by raising regenerated plants on Suc-containing medium (Figure 2). However, despite provision of an exogenous carbon source, all three double knockouts grew slowly and displayed a mutant pigment-deficient phenotype (Figure 2B). This was particularly surprising in the case of the Δrps15/Δrpl33 plants because both single mutants have no visible phenotype in sterile culture or in soil (Figure 2A; Rogalski et al., 2008; Fleischmann et al., 2011).
Figure 2.

Phenotypes of Wild-Type Plants, Single Knockout Plants, and Double Knockout Plants of the Three Nonessential Plastid Ribosomal Protein Genes rps15, rpl33, and rpl36.
The plants were grown from stem cuttings on synthetic medium supplemented with Suc as carbon source.
(A) Comparison of the wild type with the single mutants ∆rps15, ∆rpl33, and ∆rpl36. While the ∆rps15 and ∆rpl33 plants display a wild-type-like phenotype, the ∆rpl36 knockout plants are pale and show strongly delayed greening (Rogalski et al., 2008; Fleischmann et al., 2011).
(B) Phenotypes of the ∆rps15/∆rpl33, ∆rps15/∆rpl36, and ∆rpl33/∆rpl36 double knockout plants. Note the strong pigment deficiency (particularly pronounced in the older leaves) in all three double mutants.
To investigate possible molecular causes of the mutant phenotypes, we determined the ribosome contents in all single and double mutants. Using a microfluidics-based platform for sizing and quantification of rRNA species, we measured the accumulation of the rRNAs in the large and small subunits of the plastid and cytosolic ribosomes (which serve as a proxy for ribosomal subunit accumulation; Walter et al., 2010). By calculating ratios of cytosolic to plastid rRNA species, changes in the abundance of plastid ribosomal subunits become apparent (Figures 3A and 3B). In all three double mutants, plastid ribosome content was reduced more strongly than in the single mutant with the strongest phenotype, Δrpl36. While this was not unexpected for the double mutants involving the knockout of rpl36 (Δrps15/Δrpl36 and Δrpl33/Δrpl36), it was surprising in the case of the Δrps15/Δrpl33 double knockout plants. Especially the accumulation of the 50S subunit (as evidenced by the ratio 23S rRNA:18S rRNA) was drastically reduced in the Δrps15/Δrpl33 double mutant compared with the two single mutants, which, consistent with their wild-type-like phenotype, had similar 50S levels as wild-type plants (Figure 3B).
Figure 3.

Accumulation and Processing of rRNAs in Ribosomal Protein Mutants Grown Heterotrophically on Synthetic Medium.
Wild-type plants, single mutants of rps15, rpl33, and rpl36, and the three double mutants were analyzed.
(A) Accumulation of the 16S rRNA as a proxy for the accumulation of the 30S ribosomal subunit. The values give the ratio of the plastid 16S rRNA to the cytosolic 18S rRNA and represent the means of three biological replicates. The error bars indicate the sd. Statistically significant differences (determined by one-way ANOVA and Fisher’s LSD test, P < 0.05) are indicated by the letters above the bars.
(B) Accumulation of the 23S rRNA as a proxy for the abundance of 50S ribosomal subunits.
(C) Physical map and transcript processing pattern of the plastid rRNA operon. The 7.3-kb primary transcript, the different processing intermediates, and the mature forms of the 16S and 23S rRNAs are shown. The 23S rRNA is cut into three pieces, a phenomenon known as “hidden break” processing (Delp and Kössel, 1991). The positions of the hybridization probes for the 16S and 23S rRNAs are also indicated.
(D) Accumulation and processing of the plastid 16S rRNA determined by RNA gel blot hybridization. As loading control, the ethidium bromide–stained gel region containing the cytosolic 18S rRNA is shown.
(E) Analysis of the accumulation and processing pattern of the 23S rRNA. Note quantitative differences in the efficiency of hidden break processing, which is known to be influenced by developmental cues (Rosner et al., 1974).
We next analyzed plastid rRNA accumulation and processing of the 16S rRNA and 23S rRNA by RNA gel blot analyses (Figures 3C to 3E). All three double mutants showed strongly reduced levels of both 16S and 23S rRNA. Moreover, the mutants accumulated significant amounts of unprocessed or partially processed precursor RNAs (Figures 3D and 3E), suggesting that ribosome biogenesis is less efficient than in the wild type.
Synthetic Lethality of the Δrps15/Δrpl33 Double Mutant
The Δrpl36 mutant has a severe mutant phenotype and grows very poorly under autotrophic conditions in soil (Fleischmann et al., 2011). Consistent with their even lower accumulation of plastid ribosomes, the two double mutants involving knockout of rpl36 (Δrps15/Δrpl36 and Δrpl33/Δrpl36) grew even slower and were nearly impossible to maintain under autotrophic conditions. Plants raised heterotrophically and then transferred to soil survived for some time but did not reach the reproductive stage; therefore, no seeds could be obtained.
Δrps15/Δrpl33 double knockout plants raised heterotrophically and then transferred to the greenhouse died quickly under standard growth conditions (22 to 25°C). However, when incubated under high temperatures (35°C) plants continued to grow and ultimately flowered and produced seeds. This allowed us to test the viability of this double mutant during the entire life cycle. To this end, we germinated seeds in soil and compared the seedling phenotype of the double mutant with that of the wild type and the two single mutants (Δrps15 and Δrpl33; Figure 4A). Interestingly, while the two single mutants had wild-type-like phenotypes (Rogalski et al., 2008; Fleischmann et al., 2011), the double mutant turned out to be seedling lethal (Figure 4A). The seedlings were white and died early after germination, presumably after using up the energy reserves stored in the endosperm. The seedling phenotype was similarly severe on synthetic medium supplemented with Suc (Figure 4B), indicating that the level of protein biosynthesis in the plastid is very low in the Δrps15/Δrpl33 double knockout (Figure 4B).
Figure 4.

Seedling Lethality of the ∆rps15/∆rpl33 Mutants and Rescue by Elevated Growth Temperatures.
Wild-type, ∆rps15, ∆rpl33, and ∆rps15/∆rpl33 plants were germinated either on soil ([A] and [C]) or on synthetic Suc-containing medium ([B] and [D]) and grown either at 22°C ([A] and [B]) or at 35°C ([C] and [D]).
(A) Plants germinated and grow on soil at 22°C. The photos were taken 29 d after sowing.
(B) Plants germinated and grown on synthetic medium at 22°C in the absence of antibiotics (w/o sel.) or in the presence of 500 µg/mL spectinomycin (Spec) or 50 µg/mL kanamycin (Kan). The photos were taken 22 d after sowing.
(C) Plants germinated and grown on soil at 35°C.
(D) Plants grown as in (B) but at 35°C. The plants grown without selection and the plants grown under spectinomycin selection were photographed 15 d after sowing, and the plants grown under kanamycin selection were photographed 41 d after sowing. Due to the low stability of kanamycin at elevated temperatures, the kanamycin-containing medium was replaced every 14 d.
To exclude the possibility that the seedling lethal phenotype results from photooxidative stress due to the delayed buildup of the photosynthetic apparatus, we germinated seeds of the Δrps15/Δrpl33 mutants under different light conditions, including low-light conditions (50 µE m−2 s−1). However, the mutant seedlings died in soil under all of the tested conditions, suggesting that the seedling lethality of the Δrps15/Δrpl33 double knockout is not primarily caused by light stress–induced oxidative damage.
Expression of the selectable marker genes aadA and aphA-6 does not cause mutant phenotypes (Klaus et al., 2003; Petersen et al., 2011; Rott et al., 2011; Krech et al., 2013). Moreover, extensive characterization of plant growth and photosynthetic parameters revealed no detectable difference between wild-type plants and transplastomic plants expressing the aadA marker from different neutral insertion sites in the plastid genome (e.g., Rott et al., 2011; Petersen et al., 2011; Krech et al., 2013), making it unlikely that expression of the antibiotic resistance genes affects the phenotypes observed here.
Rescue of the Synthetic Lethality of the Δrps15/Δrpl33 Double Mutant at Elevated Growth Temperatures
In bacteria, the severity of the growth phenotypes of ribosomal mutants can depend strongly on the growth conditions, especially the growth temperature (Akanuma et al., 2012). Therefore, we tested the Δrps15/Δrpl33 double knockout for survival under different temperatures. Interestingly, growth at 35°C resulted in survival of the mutant seedlings and continued growth under autotrophic conditions (Figure 4C). When kept at 35°C for at least 3 months, the Δrps15/Δrpl33 plants completed their entire life cycle, ultimately flowered, and produced seeds.
To preliminarily assess whether growth at 35°C improves plastid translation, we assayed the expression of the two plastid-encoded antibiotic resistances by germinating seeds on synthetic medium (Figure 4D). Indeed, Δrps15/Δrpl33 seedlings displayed significant resistance to both spectinomycin and kanamycin, suggesting that heat rescues the double knockout at the level of protein biosynthesis in the plastid (Figures 4B and 4D).
To provide direct evidence for the impact of elevated growth temperatures on the biogenesis of plastid ribosomes in Δrps15/Δrpl33 plants, ribosome accumulation was quantified (Figures 5A and 5B) and the accumulation and processing patterns of the 16S and 23S rRNAs were comparatively investigated at 22 and 35°C (Figures 5C and 5D). These analyses revealed that, indeed, the double mutant accumulated significantly more plastid ribosomes (relative to cytosolic ribosomes) at 35°C compared with standard growth conditions. Accumulation of the small ribosomal subunit was almost 3 times higher upon growth at 35°C than at 22°C (Figure 5A). Likewise, growth at 35°C nearly doubled the relative amount of large ribosomal subunits (Figure 5B). This result was confirmed by direct analysis of plastid rRNAs. While the double mutant had strongly reduced levels of both 16S and 23S rRNAs at 22°C, we observed a significant rescue of rRNA accumulation at 35°C (Figures 5C and 5D). Interestingly, growth under elevated temperatures also improved the efficiency of rRNA processing (Figure 5D).
Figure 5.

Growth at Elevated Temperatures Alleviates the Molecular Phenotype of ∆rps15/∆rpl33 Mutants.
(A) and (B) Accumulation of rRNAs as a proxy for the corresponding ribosomal subunits in seedlings of the wild type and the ribosomal protein mutants ∆rps15, ∆rpl33, and ∆rps15/∆rpl33. The plants were germinated and grown for 29 d on synthetic medium at either 22 or 35°C. The mean of three biological replicates is shown. The error bars indicate the sd. Statistically significant differences (two-way ANOVA and Scheffé test; P < 0.05) are indicated by the letters above the bars.
(A) Ratio of the plastid 16S rRNA to the cytosolic 18S rRNA at 22°C (dark-gray bars) and 35°C (light-gray bars).
(B) Ratio of the plastid 23S rRNA to the cytosolic 18S rRNA at 22 and 35°C.
(C) and (D) Accumulation and processing patterns of plastid rRNAs at 22 and 35°C. The ethidium bromide–stained cytosolic 18S rRNA band of the agarose gel prior to blotting is shown below each blot as a loading control. For a schematic representation of rRNA processing patterns, see Figure 3C.
(C) Comparison of 16S rRNA accumulation and processing at 22°C versus 35°C.
(D) Comparison of 23S rRNA accumulation and processing at 22°C versus 35°C.
To provide further proof of ribosome biogenesis proceeding more efficiently at elevated temperatures and to exclude the alternative possibility that higher temperatures simply make the translation process more efficient in the Δrps15/Δrpl33 plants, we sought to demonstrate that the missing ribosomal proteins are not necessary for translation once the ribosomes are assembled. To this end, we isolated and analyzed polysomes (complexes of mRNAs covered with translating ribosomes) from plants raised at 35°C and then either shifted to 22°C or kept at 35°C. Polysomes can be separated from free ribosomes and untranslated mRNAs by density gradient centrifugation. The distribution profile of an mRNA across fractionated Suc gradients correlates with its loading with ribosomes and thus provides a proxy of the intensity of its translation (Barkan, 1988, 1998; Kahlau and Bock, 2008). When we compared the polysome profiles of wild-type plants and Δrps15/Δrpl33 plants at 35°C with the profiles of plants shifted for 24 h to 22°C, no differences were detectable (Figure 6), strongly suggesting that plastid translation is equally efficient at 22 and 35°C. Moreover, the polysome profiles of the wild type and the double mutant were very similar, providing additional evidence for the mutant phenotype being caused by ribosome deficiency rather than inefficiently functioning plastid ribosomes. Thus, the L33 and S15 proteins are not required for efficient translation but rather for efficient ribosome biogenesis at ambient growth temperatures.
Figure 6.

Comparison of Translational Activities at 22 and 35°C by Polysome Analysis in Wild-Type and ∆rps15/∆rpl33 Mutant Plants.
Polysome profiles of the polycistronic psaA/psaB/rps14 transcript are shown. Polysomal complexes were separated in Suc gradients and each gradient was fractionated into six fractions (numbered from the top to the bottom, as indicated above each panel). The extracted RNAs were separated by gel electrophoresis, blotted, and hybridized to a radiolabeled psaA probe. The gradient fractions containing the bulk of the transcripts are indicated by brackets. As a control, polysomes were isolated in the presence of the polysome-dissociating antibiotic puromycin. The ethidium bromide–stained gel region containing the cytosolic 26S and 18S rRNAs is shown below each blot.
(A) Polysome loading of the psaA/psaB/rps14 transcript in wild-type plants at 35°C and 24 h after shifting the growth temperature to 22°C.
(B) Polysome loading in ∆rps15/∆rpl33 mutant plants at 35°C and 24 h after shifting the growth temperature to 22°C.
(C) Dissociation of polysomal complexes by puromycin treatment in wild-type and mutant plants.
Structural Basis for the Synthetic Lethality of rps15 and rpl33
The synthetic lethality of Δrps15/Δrpl33 seedlings raised the interesting question why the combined knockout of rps15 and rpl33 is incompatible with autotrophic growth under normal conditions. In view of the absence of a mutant phenotype of the two single mutants and considering the fact that Rps15 and Rpl33 are components of different subunits of the ribosome that accumulate largely independently of each other (Tiller et al., 2012), the synthetic lethality of the double mutant was unexpected. To determine the possible molecular cause of the synthetic lethality, we examined the molecular interactions of the two proteins (and, additionally, the Rpl36 protein knocked out in the two other double mutants created in this study) in the 70S ribosome (Figure 7). In particular, we considered the possibility that one of the ribosomal proteins plays a role in both the 50S and the 30S subunit of the ribosome. While Rpl33 and Rpl36 are deeply embedded in the 50S subunit and thus are far away from the 30S subunit, the Rps15 protein is located close to the interface between the 30S and the 50S subunit (Figure 7A). Interestingly, the protein touches a loop of the 23S rRNA (Figure 7B; Culver et al., 1999). This observation raises the intriguing possibility that, in the absence of Rpl33, the biogenesis of the 50S ribosomal subunit is critically dependent on a molecular interaction between the Rps15 protein and the 23S rRNA.
Figure 7.

Position of the Ribosomal Proteins S15, L33, and L36 in the 70S Ribosome.
Shown are the 16S rRNA (pink ribbon) and the S15 protein (dark gray and yellow) of the small ribosomal subunit (30S) as well as the 23S rRNA (gray ribbon), the L33 (light blue and yellow), and the L36 (brown) proteins of the large ribosomal subunit (50S).
(A) Localization of S15, L33, and L36 in relation to the 16S and 23S rRNAs.
(B) Close-up view revealing the interaction of S15 with the 23S rRNA via an RNA loop that protrudes into the 30S subunit.
DISCUSSION
In this work, we generated all possible combinations of double mutants for the three nonessential plastid genome–encoded proteins of the 70S ribosome: S15, L33, and L36 (Rogalski et al., 2008; Fleischmann et al., 2011). The initial objective of this study was to obtain a minimum-size ribosome that is sufficient for plant survival and consists of the smallest possible set of plastid ribosomal proteins. Unexpectedly, the combined knockout of S15 and L33, the two nonessential ribosomal proteins that can be removed individually without causing mutant phenotypes under standard conditions, turned out to be incompatible with seedling survival and autotrophic growth at standard temperatures.
In view of the strong mutant phenotype of the rpl36 single knockout (Fleischmann et al., 2011), the severe phenotypes of the two double mutants involving rpl36 were expected (Figure 2) and are readily explained by additive effects (Figure 3). By contrast, the synthetic lethality of the Δrps15/Δrpl33 seedlings was surprising because Rps15 is a component of the 30S ribosomal subunit, whereas Rpl33 is a component of the 50S ribosomal subunit. The two ribosomal subunits accumulate largely independently of each other and are thought to not require each other during subunit assembly.
The significant reduction in ribosome content seen in the rps15 single knockout (Figure 3; Fleischmann et al., 2011) indicates that plants tolerate a moderate reduction in their plastid translational capacity relatively well. The synthetic lethal phenotype of the Δrps15/Δrpl33 double mutant suggests that, in these plants, plastid ribosome levels fall below a critical threshold that is required for autotrophic growth. Previous work has established that a high activity of plastid protein biosynthesis is particularly important at the young seedling stage and in young developing leaves (Fleischmann et al., 2011; Alkatib et al., 2012b; Tiller et al., 2012). This is because in these tissues, the entire photosynthetic apparatus needs to be built up, a process that requires high translational capacity in the plastid compartment. By contrast, in mature (fully expanded) leaves, lower levels of plastid translation appear to be sufficient for maintenance and repair of the photosynthetic apparatus. This may explain why Δrps15/Δrpl33 seedlings are white both on Suc-containing medium and in soil (Figures 4A and 4B), whereas the leaves of older plants raised on Suc are green (Figure 2B).
In the assembly of the 30S ribosomal subunit, ribosomal protein S15 binds early to the 16S rRNA and is possibly involved in triggering conformational changes that facilitate subsequent assembly steps (Talkington et al., 2005; Kaczanowska and Rydén-Aulin, 2007; Woodson, 2008; Sykes et al., 2010). Analysis of the three-dimensional structure of the 70S ribosome revealed that S15 also interacts with a loop of the 23S rRNA (Figure 7), providing a possible functional link between S15 and the 50S ribosomal subunit (Culver et al., 1999). However, this link becomes apparent only in the absence of L33. While biogenesis of 30S and 50S subunits proceeds efficiently when only S15 is missing, the biogenesis of both ribosomal subunits is strongly impaired when S15 and L33 are missing.
Elevated growth temperatures rescue the synthetic lethality of the Δrps15/Δrpl33 double mutant. While wild-type-like growth is not fully restored, ribosome biogenesis is improved to an extent sufficient to enable autotrophic growth (Figures 3 and 4). In view of the RNA binding properties of ribosomal proteins and, especially, the early binding of S15 to the 16S rRNA (Talkington et al., 2005), it seems conceivable that elevated temperatures facilitate proper folding of rRNAs in the absence of S15 and L33. This could occur, for example, by melting of incorrectly folded secondary structures that, under ambient temperatures, would require the action of rRNA binding ribosomal proteins.
Interestingly, a number of previously isolated mutants display temperature-dependent ribosome deficiencies. The erythromycin-resistant Chlamydomonas reinhardtii mutant ery-M2 (Hanson and Bogorad, 1978), the barley (Hordeum vulgare) mutant tigrina-o34 (Høyer-Hansen and Casadoro, 1982), and the maize (Zea mays) virescent mutants v16 and hcf7 (Hopkins and Elfman, 1984; Barkan, 1993) all show more severe ribosome deficiency upon growth at lower temperatures. Together with the chilling-sensitive phenotype of previously characterized ribosomal protein mutants in tobacco (Rogalski et al., 2008; Fleischmann et al., 2011) and the rescue of the synthetic lethality of the Δrps15/Δrpl33 mutant by heat reported here, this indicates that, upon genetic disturbances, the biogenesis of plastid ribosomes can become highly sensitive to temperature.
The synthetic lethality of the Δrps15/Δrpl33 double mutant also brings to light an important lesson in functional genomics and synthetic biology. The reduction of a genome to its minimum size and coding capacity is far from straightforward. Even if nonessential genes act in different macromolecular complexes that are believed to accumulate independently of each other (such as the 30S and 50S ribosomal particles in the case of rps15 and rpl33), their combined deletion can be lethal. Thus, to predict synthetic lethality or develop experimental approaches that allow the systematic detection of synthetically lethal interactions between genes represents a major challenge in synthetic genomics.
METHODS
Plant Material and Growth Conditions
To generate leaf material for chloroplast transformation experiments, tobacco plants (Nicotiana tabacum cv Petit Havana; Δrps15, Δrpl33, and Δrpl36; Rogalski et al., 2008; Fleischmann et al., 2011) were grown under aseptic conditions on agar-solidified Murashige and Skoog medium supplemented with 30 g/L Suc (Murashige and Skoog, 1962). Transplastomic double knockout lines were rooted and propagated on the same medium in the presence of kanamycin (20 mg/L). Plant lines were maintained in sterile culture under low-light conditions (3.5 µE m−2 s−1). To obtain seeds from the Δrps15/Δrpl33 lines, plants were grown in soil under elevated temperature (35°C) in a controlled environment chamber. Inheritance patterns and seedling phenotypes were analyzed by germination of surface-sterilized seeds on Petri dishes containing Murashige and Skoog medium supplemented with spectinomycin (500 mg/L) or kanamycin (20 mg/L).
Growth tests under different temperatures were performed by raising wild-type and mutant plants from seeds in soil at 22 or 35°C under low-light conditions (50 to 100 μE m−2 s−1) in a 16-h-light/8-h-dark regime.
Construction of Plastid Transformation Vectors
The construction of transformation vectors for the targeted inactivation of rps15, rpl33, and rpl36 was described previously (Rogalski et al., 2008; Fleischmann et al., 2011). For supertransformation, a modified Δrps15 vector was built that contained the kanamycin resistance gene aphA-6 instead of the spectinomycin resistance gene aadA (Bateman and Purton, 2000; Huang et al., 2002; Fleischmann et al., 2011). The aphA-6 expression cassette was driven by the psbA promoter from Chlamydomonas reinhardtii and the rbcL 3′ untranslated region from C. reinhardtii (Zou et al., 2003). The cassette was excised with SmaI and inserted into a unique HincII restriction site in the rps15 coding region (Fleischmann et al., 2011; Figure 1). A vector for targeted disruption of rpl36 with the aphA-6 gene was assembled from cloned PCR products. The aphA-6 gene was amplified with primers 36-aphfor and 36-2aphrev (Supplemental Table 1) using plasmid DNA as template. The sequence flanking rpl36 was amplified with the primers 36-2inffor and 36-2rps8rev using tobacco genomic DNA as template. The two PCR products were fused by amplification with primers 36-aphfor and 36-2rps8rev. The resulting fragment was digested with XhoI and BsrGI and inserted into the similarly cut plasmid pSA8 (Fleischmann et al., 2011), generating the final plastid transformation vector for rpl36 disruption with the aphA-6 cassette (Figure 1).
Plastid Transformation and Selection of Transplastomic Lines
Transplastomic Δrpl33/Δrpl36 lines were produced by supertransformation of Δrpl33 mutants (Rogalski et al., 2008) with the aphA-6–containing knockout allele for rpl36 (Figures 1A and 1C). Similarly, Δrps15/Δrpl33 lines were obtained by supertransformation of Δrpl33 with the kanamycin-resistant rps15 knockout allele (Figure 1B). Δrps15/Δrpl36 double mutants were generated by supertransformation of Δrps15 mutants (Fleischmann et al., 2011) with the aphA-6 knockout allele for rpl36 (Figure 1C). For plastid transformation, young leaves from aseptically grown tobacco plants of the individual recipient lines were bombarded with plasmid-coated 0.6-μm gold particles using a helium-driven biolistic gun (PDS1000He; Bio-Rad). Primary transplastomic lines were selected on plant regeneration medium (Svab and Maliga, 1993) containing 25 mg/L kanamycin. Several independent transplastomic lines were generated for each construct and subjected to two to three additional rounds of regeneration on kanamycin-containing plant regeneration medium to select for stable homoplasmy.
Isolation of Nucleic Acids and Hybridization Procedures
Total plant DNA was extracted from plants grown under kanamycin selection in vitro by a rapid cetyltrimethylammoniumbromide-based method (Doyle and Doyle, 1990). For RFLP analysis, DNA samples were digested with restriction enzymes, separated in 1.0% agarose gels, and blotted onto Hybond N nylon membranes (GE Healthcare). Total plant RNA was isolated with the peqGOLD TriFast reagent (Peqlab) following the protocol of the manufacturer. RNA samples were denatured, separated in denaturing formaldehyde-containing 1.2% agarose gels, and blotted onto Hybond N nylon membranes (GE Healthcare). For hybridization, [α-32P]dCTP-labeled probes were produced by random priming (Multiprime DNA labeling kit; GE Healthcare). Amplified PCR products were used as hybridization probes (for primer sequences, see Supplemental Table 1). Hybridizations were performed at 65°C using standard protocols.
Quantitative Analysis of rRNA Accumulation
For relative quantitation of plastid and cytosolic rRNA species, total RNA samples were analyzed in an Agilent 2100 Bioanalyzer using the RNA 6000 nano kit according to the instructions of the manufacturer (Agilent Technologies). rRNA ratios were determined as described previously (Walter et al., 2010). Three biological replicates were measured for all plant lines and growth conditions. The mitochondrial rRNAs, although of different size than the plastid rRNAs (mitochondrial 26S rRNA, 3406 nucleotides; mitochondrial 18S rRNA, 1902 nucleotides; NC_006581), are not detected in these assays due to their much lower abundance.
Polysome Analyses
For isolation of polysomes, plants were germinated in soil and grown at 35°C and 100 µE m−2 s−1 until they reached the four-leaf stage. For temperature-shift experiments, plants were transferred to 22°C for 24 h. To avoid artifacts from differences in leaf age or development, whole plants (aboveground parts) were harvested and several plants were pooled prior to polysome extraction. Polysome analysis and puromycin treatment were performed essentially as described previously (Barkan, 1998; Rogalski et al., 2008) with the only exception that RNA pellets were dissolved in 20 μL of 10 mM Tris HCl, pH 8.1. Aliquots of 5 μL per fraction were heat-denatured and loaded onto 1% denaturing formaldehyde-containing agarose gels. Following electrophoretic separation, the gels were blotted and hybridized to a radiolabeled psaA-specific probe generated by PCR amplification with primer pair P5′psaA/P3′psaA (Supplemental Table 1).
Analysis of Ribosome Structure
The structure of the Escherichia coli ribosome (Seidelt et al., 2009) was obtained from the Molecular Modeling Database MMDB (Madej et al., 2012; PDB ID: 2WWL and 2WWQ). To visualize protein–protein interactions and RNA–protein interactions, three-dimensional structures were depicted with the Cn3D viewer (Wang et al., 2000).
Accession Numbers
Sequence data from this article can be found in the GenBank/EMBL databases under accession number Z00044.2 (tobacco chloroplast genome).
Supplemental Data
The following materials are available in the online version of this article.
Supplemental Table 1. List of Oligonucleotides Used in This Study.
Supplementary Material
Acknowledgments
We thank Dirk Walther (Max-Planck-Institut für Molekulare Pflanzenphysiologie) for help with depicting three-dimensional ribosome structures and Sandra Stegemann (Max-Planck-Institut für Molekulare Pflanzenphysiologie) for technical assistance with the polysome analyses. This work was supported by the Max Planck Society and by a grant from the Deutsche Forschungsgemeinschaft to R.B. (FOR 804; BO 1482/15-2).
AUTHOR CONTRIBUTIONS
M.E., L.B.S., and T.T.F. designed and performed research and analyzed data. C.H. performed research. S.R. designed research and analyzed data. R.B. designed research, analyzed data, and wrote the article.
Glossary
- RFLP
restriction fragment length polymorphism
Footnotes
Online version contains Web-only data.
References
- Ahlert D., Ruf S., Bock R. (2003). Plastid protein synthesis is required for plant development in tobacco. Proc. Natl. Acad. Sci. USA 100: 15730–15735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Akanuma G., Nanamiya H., Natori Y., Yano K., Suzuki S., Omata S., Ishizuka M., Sekine Y., Kawamura F. (2012). Inactivation of ribosomal protein genes in Bacillus subtilis reveals importance of each ribosomal protein for cell proliferation and cell differentiation. J. Bacteriol. 194: 6282–6291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alkatib S., Fleischmann T.T., Scharff L.B., Bock R. (2012a). Evolutionary constraints on the plastid tRNA set decoding methionine and isoleucine. Nucleic Acids Res. 40: 6713–6724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alkatib S., Scharff L.B., Rogalski M., Fleischmann T.T., Matthes A., Seeger S., Schöttler M.A., Ruf S., Bock R. (2012b). The contributions of wobbling and superwobbling to the reading of the genetic code. PLoS Genet. 8: e1003076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baba T., Ara T., Hasegawa M., Takai Y., Okumura Y., Baba M., Datsenko K.A., Tomita M., Wanner B.L., Mori H. (2006). Construction of Escherichia coli K-12 in-frame, single-gene knockout mutants: The Keio collection. Mol. Syst. Biol. 2: 0008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barkan A. (1988). Proteins encoded by a complex chloroplast transcription unit are each translated from both monocistronic and polycistronic mRNAs. EMBO J. 7: 2637–2644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barkan A. (1993). Nuclear mutants of maize with defects in chloroplast polysome assembly have altered chloroplast RNA metabolism. Plant Cell 5: 389–402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barkan A. (1998). Approaches to investigating nuclear genes that function in chloroplast biogenesis in land plants. Methods Enzymol. 297: 38–57. [Google Scholar]
- Bateman J.M., Purton S. (2000). Tools for chloroplast transformation in Chlamydomonas: Expression vectors and a new dominant selectable marker. Mol. Gen. Genet. 263: 404–410. [DOI] [PubMed] [Google Scholar]
- Bock R., Timmis J.N. (2008). Reconstructing evolution: gene transfer from plastids to the nucleus. Bioessays 30: 556–566. [DOI] [PubMed] [Google Scholar]
- Boynton J.E., Gillham N.W., Harris E.H., Hosler J.P., Johnson A.M., Jones A.R., Randolph-Anderson B.L., Robertson D., Klein T.M., Shark K.B., Sanford J.C. (1988). Chloroplast transformation in Chlamydomonas with high velocity microprojectiles. Science 240: 1534–1538. [DOI] [PubMed] [Google Scholar]
- Cambray G., Mutalik V.K., Arkin A.P. (2011). Toward rational design of bacterial genomes. Curr. Opin. Microbiol. 14: 624–630. [DOI] [PubMed] [Google Scholar]
- Carrer H., Hockenberry T.N., Svab Z., Maliga P. (1993). Kanamycin resistance as a selectable marker for plastid transformation in tobacco. Mol. Gen. Genet. 241: 49–56. [DOI] [PubMed] [Google Scholar]
- Culver G.M., Cate J.H., Yusupova G.Z., Yusupov M.M., Noller H.F. (1999). Identification of an RNA-protein bridge spanning the ribosomal subunit interface. Science 285: 2133–2136. [DOI] [PubMed] [Google Scholar]
- Delaye L., Moya A. (2010). Evolution of reduced prokaryotic genomes and the minimal cell concept: Variations on a theme. Bioessays 32: 281–287. [DOI] [PubMed] [Google Scholar]
- Delp, G., and Kössel, H. (1991). rRNAs and rRNA genes of plastids. In Cell Culture and Somatic Cell Genetics of Plants, L. Bogorad and I.K. Vasil, eds (New York: Academic Press), pp. 139–167. [Google Scholar]
- Doyle J.J., Doyle J.L. (1990). Isolation of plant DNA from fresh tissue. Focus 12: 13–15. [Google Scholar]
- Fleischmann T.T., Scharff L.B., Alkatib S., Hasdorf S., Schöttler M.A., Bock R. (2011). Nonessential plastid-encoded ribosomal proteins in tobacco: A developmental role for plastid translation and implications for reductive genome evolution. Plant Cell 23: 3137–3155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graack H.-R., Wittmann-Liebold B. (1998). Mitochondrial ribosomal proteins (MRPs) of yeast. Biochem. J. 329: 433–448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gray M.W. (1993). Origin and evolution of organelle genomes. Curr. Opin. Genet. Dev. 3: 884–890. [DOI] [PubMed] [Google Scholar]
- Hager M., Biehler K., Illerhaus J., Ruf S., Bock R. (1999). Targeted inactivation of the smallest plastid genome-encoded open reading frame reveals a novel and essential subunit of the cytochrome b(6)f complex. EMBO J. 18: 5834–5842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanson M.R., Bogorad L. (1978). The ery-M2 group of Chlamydomonas reinhardii: Cold-sensitive, erythromycin-resistant mutants deficient in chloroplast ribosomes. J. Gen. Microbiol. 105: 253–262. [Google Scholar]
- Hopkins W.G., Elfman B. (1984). Temperature-induced chloroplast ribosome deficiency in virescent maize. J. Hered. 75: 207–211. [Google Scholar]
- Huang F.-C., Klaus S.M.J., Herz S., Zou Z., Koop H.-U., Golds T.J. (2002). Efficient plastid transformation in tobacco using the aphA-6 gene and kanamycin selection. Mol. Genet. Genomics 268: 19–27. [DOI] [PubMed] [Google Scholar]
- Høyer-Hansen G., Casadoro G. (1982). Unstable chloroplast ribosomes in the cold-sensitive barley mutant tigrina-o34. Carlsberg Res. Commun. 47: 103–118. [Google Scholar]
- Ikegami A., Nishiyama K.-i., Matsuyama S.-i., Tokuda H. (2005). Disruption of rpmJ encoding ribosomal protein L36 decreases the expression of secY upstream of the spc operon and inhibits protein translocation in Escherichia coli. Biosci. Biotechnol. Biochem. 69: 1595–1602. [DOI] [PubMed] [Google Scholar]
- Kaczanowska M., Rydén-Aulin M. (2007). Ribosome biogenesis and the translation process in Escherichia coli. Microbiol. Mol. Biol. Rev. 71: 477–494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kahlau S., Bock R. (2008). Plastid transcriptomics and translatomics of tomato fruit development and chloroplast-to-chromoplast differentiation: Chromoplast gene expression largely serves the production of a single protein. Plant Cell 20: 856–874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klasson L., Andersson S.G.E. (2010). Research on small genomes: Implications for synthetic biology. Bioessays 32: 288–295. [DOI] [PubMed] [Google Scholar]
- Klaus S.M.J., Huang F.-C., Eibl C., Koop H.-U., Golds T.J. (2003). Rapid and proven production of transplastomic tobacco plants by restoration of pigmentation and photosynthesis. Plant J. 35: 811–821. [DOI] [PubMed] [Google Scholar]
- Kode V., Mudd E.A., Iamtham S., Day A. (2005). The tobacco plastid accD gene is essential and is required for leaf development. Plant J. 44: 237–244. [DOI] [PubMed] [Google Scholar]
- Krech K., Fu H.-Y., Thiele W., Ruf S., Schöttler M.A., Bock R. (2013). Reverse genetics in complex multigene operons by co-transformation of the plastid genome and its application to the open reading frame previously designated psbN. Plant J. 75: 1062–1074. [DOI] [PubMed] [Google Scholar]
- Krech K., Ruf S., Masduki F.F., Thiele W., Bednarczyk D., Albus C.A., Tiller N., Hasse C., Schöttler M.A., Bock R. (2012). The plastid genome-encoded Ycf4 protein functions as a nonessential assembly factor for photosystem I in higher plants. Plant Physiol. 159: 579–591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li W., Ruf S., Bock R. (2011). Chloramphenicol acetyltransferase as selectable marker for plastid transformation. Plant Mol. Biol. 76: 443–451. [DOI] [PubMed] [Google Scholar]
- Madej T., et al. (2012). MMDB: 3D structures and macromolecular interactions. Nucleic Acids Res. 40: D461–D464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maguire B.A., Wild D.G. (1997). The roles of proteins L28 and L33 in the assembly and function of Escherichia coli ribosomes in vivo. Mol. Microbiol. 23: 237–245. [DOI] [PubMed] [Google Scholar]
- Maliga P. (2004). Plastid transformation in higher plants. Annu. Rev. Plant Biol. 55: 289–313. [DOI] [PubMed] [Google Scholar]
- Manuell A.L., Quispe J., Mayfield S.P. (2007). Structure of the chloroplast ribosome: Novel domains for translation regulation. PLoS Biol. 5: e209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murashige T., Skoog F. (1962). A revised medium for rapid growth and bio assays with tobacco tissue culture. Physiol. Plant. 15: 473–497. [Google Scholar]
- Petersen K., Schöttler M.A., Karcher D., Thiele W., Bock R. (2011). Elimination of a group II intron from a plastid gene causes a mutant phenotype. Nucleic Acids Res. 39: 5181–5192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rochaix J.-D. (1997). Chloroplast reverse genetics: New insights into the function of plastid genes. Trends Plant Sci. 2: 419–425. [Google Scholar]
- Rogalski M., Ruf S., Bock R. (2006). Tobacco plastid ribosomal protein S18 is essential for cell survival. Nucleic Acids Res. 34: 4537–4545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rogalski M., Schöttler M.A., Thiele W., Schulze W.X., Bock R. (2008). Rpl33, a nonessential plastid-encoded ribosomal protein in tobacco, is required under cold stress conditions. Plant Cell 20: 2221–2237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosner A., Porath D., Gressel J. (1974). The distribution of plastid ribosomes and the integrity of plastid ribosomal RNA during the greening and maturation of Spirodela fronds. Plant Cell Physiol. 15: 891–902. [Google Scholar]
- Rott M., Martins N.F., Thiele W., Lein W., Bock R., Kramer D.M., Schöttler M.A. (2011). ATP synthase repression in tobacco restricts photosynthetic electron transport, CO2 assimilation, and plant growth by overacidification of the thylakoid lumen. Plant Cell 23: 304–321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seidelt B., Innis C.A., Wilson D.N., Gartmann M., Armache J.-P., Villa E., Trabuco L.G., Becker T., Mielke T., Schulten K., Steitz T.A., Beckmann R. (2009). Structural insight into nascent polypeptide chain-mediated translational stalling. Science 326: 1412–1415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shinozaki K., et al. (1986). The complete nucleotide sequence of the tobacco chloroplast genome: Its gene organization and expression. EMBO J. 5: 2043–2049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Svab Z., Maliga P. (1993). High-frequency plastid transformation in tobacco by selection for a chimeric aadA gene. Proc. Natl. Acad. Sci. USA 90: 913–917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sykes M.T., Shajani Z., Sperling E., Beck A.H., Williamson J.R. (2010). Quantitative proteomic analysis of ribosome assembly and turnover in vivo. J. Mol. Biol. 403: 331–345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takahashi Y., Goldschmidt-Clermont M., Soen S.-Y., Franzén L.G., Rochaix J.-D. (1991). Directed chloroplast transformation in Chlamydomonas reinhardtii: Insertional inactivation of the psaC gene encoding the iron sulfur protein destabilizes photosystem I. EMBO J. 10: 2033–2040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Talkington M.W.T., Siuzdak G., Williamson J.R. (2005). An assembly landscape for the 30S ribosomal subunit. Nature 438: 628–632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tiller N., Weingartner M., Thiele W., Maximova E., Schöttler M.A., Bock R. (2012). The plastid-specific ribosomal proteins of Arabidopsis thaliana can be divided into non-essential proteins and genuine ribosomal proteins. Plant J. 69: 302–316. [DOI] [PubMed] [Google Scholar]
- Timmis J.N., Ayliffe M.A., Huang C.Y., Martin W. (2004). Endosymbiotic gene transfer: Organelle genomes forge eukaryotic chromosomes. Nat. Rev. Genet. 5: 123–135. [DOI] [PubMed] [Google Scholar]
- Walter M., Piepenburg K., Schöttler M.A., Petersen K., Kahlau S., Tiller N., Drechsel O., Weingartner M., Kudla J., Bock R. (2010). Knockout of the plastid RNase E leads to defective RNA processing and chloroplast ribosome deficiency. Plant J. 64: 851–863. [DOI] [PubMed] [Google Scholar]
- Wang Y., Geer L.Y., Chappey C., Kans J.A., Bryant S.H. (2000). Cn3D: Sequence and structure views for Entrez. Trends Biochem. Sci. 25: 300–302. [DOI] [PubMed] [Google Scholar]
- Woodson S.A. (2008). RNA folding and ribosome assembly. Curr. Opin. Chem. Biol. 12: 667–673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamaguchi K., Subramanian A.R. (2000). The plastid ribosomal proteins. Identification of all the proteins in the 50 S subunit of an organelle ribosome (chloroplast). J. Biol. Chem. 275: 28466–28482. [DOI] [PubMed] [Google Scholar]
- Yamaguchi K., von Knoblauch K., Subramanian A.R. (2000). The plastid ribosomal proteins. Identification of all the proteins in the 30 S subunit of an organelle ribosome (chloroplast). J. Biol. Chem. 275: 28455–28465. [DOI] [PubMed] [Google Scholar]
- Zou Z., Eibl C., Koop H.-U. (2003). The stem-loop region of the tobacco psbA 5’UTR is an important determinant of mRNA stability and translation efficiency. Mol. Genet. Genomics 269: 340–349. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.