

Abstract
The mediobasal segment (S7) of the right lung has been considered to correspond to the cardiac lobe generally seen in mammals. To investigate fetal development of the right mediobasal segmental bronchus (B7), we examined paraffin-embedded serial sections of 15 embrynic and fetal lungs at 7-8 weeks (serial sections) as well as semiserial sections of 8 fetuses at 15-18 weeks (semiserial sections). All of the smaller specimens did not contain B7, but 2 of the 8 larger specimens carried B7: one was found in the immediately anterior side of the inferior pulmonary vein, while in the other, the subdivisions (B7a, B7b) were overriding the vein. Although the incidence might be underestimated because of observations using semiserial sections, the B7 was most likely to develop secondarily during a period from 8 to 15 weeks. Fetal topographical changes (mainly, the descent) of the middle lobe and the inferior pulmonary vein might relate with the secondarily budding of B7. The present result does not reduce a clinical relevance of B7 as a segmental bronchus of the lung segment system.
Keywords: Lung segment, Right lower lobe, Mediobasal segment, Segmental bronchus VII, Human fetus
Introduction
The human lung segment system includes 2 small segments, namely the subsuperior and mediobasal segments [1]. The mediobasal or medial basal segment (segment VII, S7) is present in 84% of Japanese people [2, 3]. However, from a phylogenetic viewpoint, S7 seems to correspond to a stable structure termed cardiac lobe, which is a superomedial convex part of the lower lobe attaching to the pericardiac sac [4, 5, 6, 7, 8, 9]. Appleton [10], Boyden [11], and some Japanese surgeons [2, 3] consider S7 to be an independent lung segment, in contrast to Jackson and Huber [12], who consider the area to be a part of segment VIII (S8). In the right lung, the supplying bronchus or B7 (bronchus segmentalis basalis medialis or cardiacus) originates from the lower lobe of the bronchus at a point almost 5 mm inferior to the origin of the segmental brochus VI (B6). In contrast, in the left lung, B7 is frequently absent and the lung parenchyma corresponding to the right S7 is supplied by the adjacent subsegmental bronchi, that is, B6c, B6a, B9a, and/or B10a. However, it has been described in the left lung in a few cases (10% in Boyden [11]; 4.4% in Yamashita [2]).
It is not known whether B7 develops at the same time as the other major segmental bronchi or appears secondarily. We hypothesized that according to the phylogenetic background, B7 is likely to develop in the early prenatal period together with the adjacent segmental bronchi. However, to our knowledge, previous fetal studies have not described B7 or its associated vessels [13, 14, 15, 15, 16, 17, 18]. Descriptions of the fetal development of segmental bronchi are limited to those of Bucher and Reid [13], but they eliminated B7 in their excellent study. Fortunately, even in the small fetal lung, B7 in the right lung seems to be easily identified in the immediately anterior side of the right inferior pulmonary vein at the pulmonary hilum [2, 3]. B7 runs downward in the anterior side of the inferior pulmonary vein (type B7a: 53% in Yamashita [2]; 64% in Arai and Shiozawa [3]), or the bronchus divides into 2 subsegmental bronchi, B7a and B7b, and they sandwich the inferior pulmonary vein (type B7a,b: 30.5% in Yamashita [2]; 20% in Arai and Shiozawa [3]). In type B7a,b, B7a is anterior to the vein, whereas B7b is posterior to the vein. The other type, type BX7a,b, corresponds to the absence of S7 (16% in Arai and Shiozawa [3]; 16.5% in Yamashita [2]).
In the present study, the terminology for the bronchi is based on the Japanese classification system of lung segments [2], which was based on Jackson and Huber's system in the United States [12]. However, in the right lobe, the difference between the Japanese system and Boyden's is simple; it is limited to the numbering of S2 and S3. S3 in Boyden's classification corresponds to S2 in the Japanese system [11].
Materials and Methods
The study was performed in accordance with the provisions of the Declaration of Helsinki 1995 (as revised in Edinburgh 2000). We examined the paraffin-embedded histology of 25 embryos and fetuses at 7-18 weeks of estimated gestational age (21-155 mm crown-rump length [CRL]), that is, 17 embryos at 7-8 weeks (CRL, 21-38 mm) and 8 fetuses at 15-18 weeks (CRL, 118-155 mm). All the embryos and fetuses were part of the large collection kept at the Embryology Institute of the Universidad Complutense, Madrid, being the products of terminations, miscarriages, or ectopic pregnancies managed at the Department of Obstetrics of the University. The ethics committee of the university granted approval for the study. The donated embryos and fetuses were fixed with 10% v/v formalin solution for more than 3 months. After division into the head and neck, thorax, abdomen, and pelvis, and the four extremities, the thorax specimens from the 8 larger specimens (15-18 weeks) were decalcified by incubating them at room temperature using the Plank-Rychlo solution (AlCl2/6H2O, 7.0 w/v%; HCl, 3.6; HCOOH, 4.6) for 3-5 days. After the routine procedure for the paraffin-embedded histological examination, 7-µm-thick horizontal sections of the whole thorax were prepared (serially for 17 smaller specimens or at 50- to 100-µm intervals for 8 larger specimens). The sections were stained with hematoxylin and eosin staining or Azan staining.
Results
We searched B7 along the right lower lobe of the bronchus from the origin of B6 to the level in which B8, B9, and B10 were identified. However, the present photographic demonstration in figures was chosen especially sections including the inferior pulmonary vein. A candidate for B7 was absent in 17 specimens at 7-8 weeks (Figs. 1, 2) and in 6 of 8 specimens at 15-18 weeks (Fig. 3), although observations of the larger specimens were based on semiserial sections. B7 was present in 2 of 8 specimens at 15-18 weeks (Fig. 4); it was located in the anterior side of the inferior pulmonary vein in 1 specimen, while in another specimen, it divided into 2 subsegmental bronchi (i.e., B7a and B7b; see Introduction), both of which were overriding the vein (Fig. 4B). B7a (anterior subdivision) appeared to accompany a proper artery. In the 2 specimens with B7, a peninsula of S5 extended posteriorly to approach the inferior pulmonary vein. Without the peninsula, the area was occupied by lymphatic follicles. In the larger specimens without B7, a loose mesenchymal tissue in the immediately anterior side of the vein (a border area between S8 and the middle lobe) was occupied by candidate lymphatic vessels (Fig. 3) and the lung parenchyme anterior to the vein was supplied by branches of B8.
Fig. 1.

Five right lung specimens at 7-8 weeks. Panels are shown in order from the smaller lung to the larger one. Note that a specimen with 39 mm crown-rump length (CRL) carries a lung smaller than those in specimens with 37 mm CRL. Panels (A), (D), and (E) include the middle and lower lobes, while panels (B) and (C) the lower lobe only. Star indicates an area in which the segmental bronchus VII is likely to present in the immediately anterior side of the inferior pulmonary vein (IPV). Asterisk indicates a cavity through the oval window between the right atrium (RA) and left atrium (LA). AO, aorta; B6, B8, B9, B10, segmental bronchi VI, VIII, IX, and X; B9+10, a common trunk bronchus with the segmental bronchi IX and X; ES, esophagus. All panels are prepared at the same magnification. Scale bar in panel (A)=1 mm (A-E).
Fig. 2.
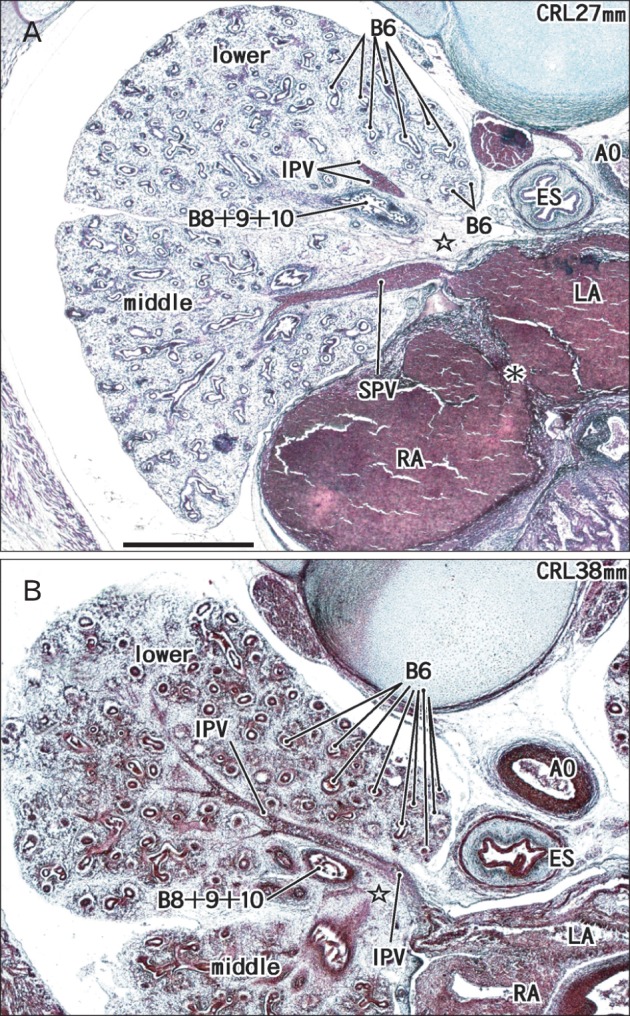
Two right lung specimens at 7-8 weeks. Note that a specimen with 27 mm crown-rump length (CRL) (A) carries a lung larger than those in specimens with 37 or 39 mm CRL shown in Fig. 1. Both panels include the middle and lower lobes. Star indicates an area in which the segmental bronchus VII is likely to present in the immediately anterior side of the inferior pulmonary vein (IPV). Asterisk indicates a cavity through the oval window between the right atrium (RA) and left atrium (LA). AO, aorta; B6, segmental bronchus VI; B8+9+10, a common trunk bronchus with the segmental bronchi VIII, IX, and X; ES, esophagus; SPV, superior pulmonary vein. The two panels are prepared at the same magnification. Scale bar in panel (A)=1 mm (A, B).
Fig. 3.
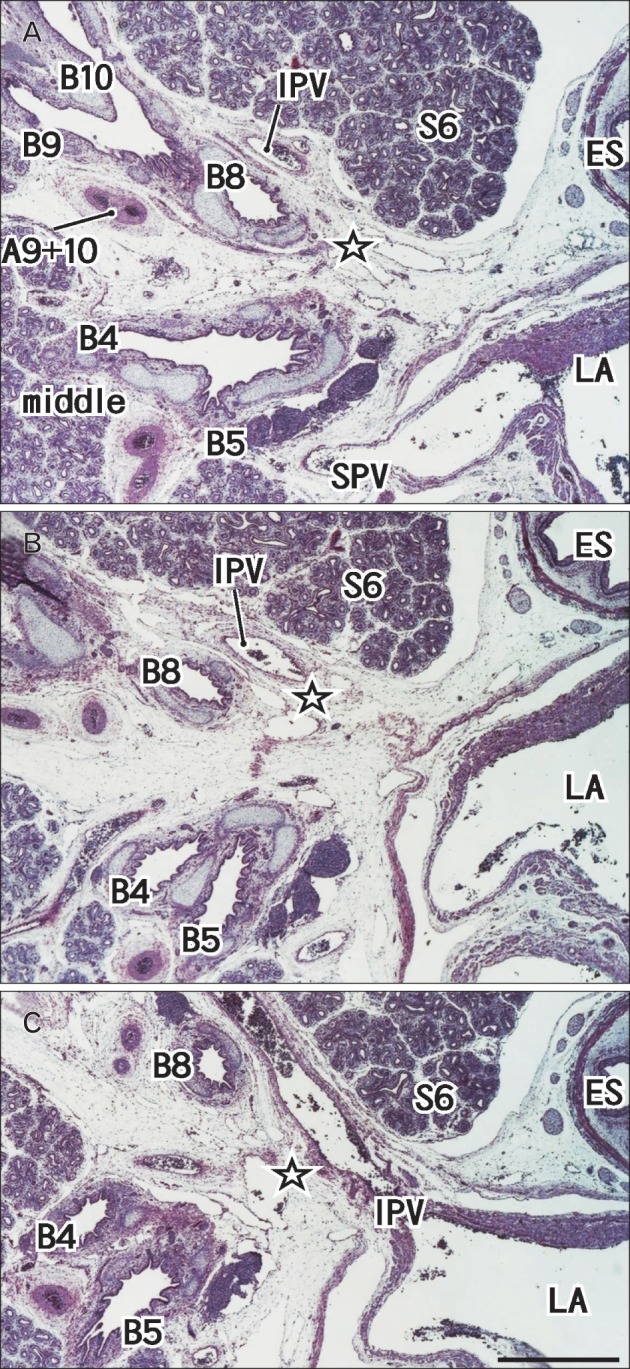
A 16-week specimen with segmental bronchus VII. Panel (A) (or C) is the most superior (or inferior) in the figure. Intervals between panels are 0.3 mm (A-B, B-C). Star indicates an area in which the segmental bronchus VII is likely to present in the immediately anterior side of the inferior pulmonary vein (IPV). A9+10, a common trunk artery of the segmental arteries IX and X; B4, B5, B6, B8, B9, and B10, segmental bronchi IV, V, VI, VIII, IX, and X; ES, esophagus; LA, left atrium; S6, segment VI; SPV, superior pulmonary vein. All panels are prepared at the same magnification and include the middle and lower lobes. Scale bar in panel (C)=1 mm (A-C).
Fig. 4.

An 18-week specimen without segmental bronchus VII. Panel (A) (or C) is the most superior (or inferior) in the figure. Intervals between panels are 0.5 mm (A-B, B-C). The topographical orientation (posterior, the right-hand side of each panel) is different from Figs. 1, 2, 3. Segmental bronchus VII (B7) is originated in the 0.5 mm superior side of panel (A), runs inferiorly (A), divides into two subsegmental bronchi (B7a and B8b) in the immediately superior side of the inferior pulmonary vein (IPV in panel B). The B7a (anterior) runs further inferiorly, while the B7b (posterior) runs posterolaterally along the inferolateral margin of the segment VI (S6 in panel C). AO, aorta; B4, B5, B6, B8, B9, and B10, segmental bronchi IV, V, VI, VIII, IX, and X; ES, esophagus; LA, left atrium. All panels are prepared at the same magnification and include the middle and lower lobes. Scale bar in panel (A)=1 mm (A-C).
The long longitudinal course of the inferior pulmonary vein was consistently seen in a horizontal plane of the right lung. In the plane and different from that in adults, the middle lobe was as large as the lower lobe. In the smaller specimens, the inferior pulmonary vein ran laterally at the level of the dividing site of "B8 and B9+10" or "B8, B9, and B10" (Figs. 1, 2), but the vein ran in the plane inferior to the lower segmental bronchial division(s) in the larger specimens (Figs. 3, 4). Likewise, at the level of the vein, more peripheral divisions of B6 were seen in the larger specimens. Thus, the extent of the descent of the vein appeared to depend on the stage. Conversely, the division of the middle lobe segmental bronchi appeared in the plane of the vein in the larger specimens in contrast to the smaller specimens. The middle lobe appeared to descend further than the vein and became as large as the lower lobe from 8 to 15 weeks. In addition, the size of the lung was not always related to growth stage (CRL); the smaller specimens sometimes had a larger lung containing paradoxically numerous peripheral bronchi (e.g., Fig. 2A). The present results are summarized in Fig. 5.
Fig. 5.

Schematic representation of the present results. At 7 weeks, except for B7, the bronchial tree in the right lower lobe has already developed at the subsegmental level: B6a, B6b, B6c, B8a, B8b, B9a, B9b, B10a, B10b, B10c. However, the inferior pulmonary vein (IPV) is thin and difficult to identify in the segmental level. At 8 weeks, the vein reaches each of the segments. At 15 weeks, in some specimens, B7 is seen crossing the anterior side of the vein.
Discussion
The present study revealed that despite the phylogenetic background, the minor segment S7 was most likely to develop secondarily to occupy a space between S8 and the middle lobe under some backgrounds (further details in the succeeding paragraphs). With regard to the frequency of the presence of B7 in the larger specimens, there appeared to be underestimation in 2 (25%) of the 8 specimens because of observations using semiserial sections of 50-100 µm. However, in the smaller specimens at 7-8 weeks, observations of serial sections prevented the possibility of identifying early budding of B7 at and before this stage.
Before the budding of B7, loose mesenchymal tissue occupied an area in the immediately anterior side of the inferior pulmonary vein (bordering the area between S8 and the middle lobe). This area of tissue seemed to be unstable and liable to change its form because of the suggested descent of the middle lobe and the inferior pulmonary vein during a period from 8 to 15 weeks, and in the larger specimens without B7, it was occupied by lymphatic vessels or follicles. A posterior peninsula of S5 might, if present, inhibit the lymphatic development and accelerate the budding of B7. According to Mawatari et al. [19], the independent posterior lobe of the right lung (an isolated S6) consistently accompanied well developed B7 and A7 (a proper segmental artery to S7). There seems to be a factor (or factors) favorable for the secondary budding of B7. On occasion, the segmental arterial supply does not take the same pattern as the bronchial ramification [20]. Because they are much thinner than the bronchi, the developing segmental arteries of the lung seemed not to influence the budding of B7. The so-called tracheal bronchi arising from the trachea may also have a secondary developmental pattern. Finally, we should state that the present results and discussion do not reduce the clinical relevance of B7 as a segmental bronchus in the lung segment system.
References
- 1.Williams PL. Gray's anatomy. 38th ed. Edinburgh: Churchill Livingstone; 1995. [Google Scholar]
- 2.Yamashita H. Roentgenologic anatomy of the lung. Tokyo: Igaku-Shoin; 1978. [Google Scholar]
- 3.Arai T, Shiozawa M. Pulmonary resections: regional anatomy and surgical procedures. 2nd ed. Tokyo: Asakura Publishing; 1992. [Google Scholar]
- 4.Huntington GS. A critique of the theories of pulmonary evolution in the mammalia. Am J Anat. 1920;27:99–201. [Google Scholar]
- 5.Iino T. Cast anatomy of the bronchial tree and ramification patterns of the pulmonary artery and vein in raccoon dogs. Ann Dep Anat Jikei Univ Sch Med. 1962;24:21–27. [Google Scholar]
- 6.Iwasaki M, Matsunaga M. Cast anatomy of the bronchial tree and ramification patterns of the pulmonary artery and vein in cats. Ann Dep Anat Jikei Univ Sch Med. 1962;24:1–15. [Google Scholar]
- 7.Kijima T. Cast anatomy of the bronchial tree and ramification patterns of the pulmonary artery and vein in raccoon dogs. Ann Dep Anat Jikei Univ Sch Med. 1962;24:128–142. [Google Scholar]
- 8.Moriya M. Cast anatomy of the bronchial tree and ramification patterns of the pulmonary artery and vein in goats. Ann Dep Anat Jikei Univ Sch Med. 1962;24:31–64. [Google Scholar]
- 9.Nakakuki S. Comparative anatomical studies on the mammalian lung. Bull Fac Agric Tokyo Univ Agric Technol. 1980;21:1–74. [Google Scholar]
- 10.Appleton AB. Segments and blood-vessels of the lungs. Lancet. 1944;244:592–594. [Google Scholar]
- 11.Boyden EA. Segmental anatomy of the lung. New York: McGraw-Hill; 1955. [Google Scholar]
- 12.Jackson CL, Huber JF. Correlated applied anatomy of the bronchial tree and lungs with a system of nomenclature. Dis Chest. 1943;9:319–326. [Google Scholar]
- 13.Bucher U, Reid L. Development of the intrasegmental bronchial tree: the pattern of branching and development of cartilage at various stages of intra-uterine life. Thorax. 1961;16:207–218. doi: 10.1136/thx.16.3.207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Burri PH. Fetal and postnatal development of the lung. Annu Rev Physiol. 1984;46:617–628. doi: 10.1146/annurev.ph.46.030184.003153. [DOI] [PubMed] [Google Scholar]
- 15.DiFiore JW, Wilson JM. Lung development. Semin Pediatr Surg. 1994;3:221–232. [PubMed] [Google Scholar]
- 16.Jeffrey PK. The development of large and small airways. Am J Respir Crit Care Med. 1998;157(5 Pt 2):S174–S180. doi: 10.1164/ajrccm.157.5.rsaa-1. [DOI] [PubMed] [Google Scholar]
- 17.Laudy JA, Wladimiroff JW. The fetal lung. 1: developmental aspects. Ultrasound Obstet Gynecol. 2000;16:284–290. doi: 10.1046/j.1469-0705.2000.00228.x. [DOI] [PubMed] [Google Scholar]
- 18.Szpinda M. The normal growth of the pulmonary trunk in human foetuses. Folia Morphol (Warsz) 2007;66:126–130. [PubMed] [Google Scholar]
- 19.Mawatari T, Murakami G, Koshino T, Morishita K, Abe T. Posterior pulmonary lobe: segmental and vascular anatomy in human specimens. Clin Anat. 2000;13:257–262. doi: 10.1002/1098-2353(2000)13:4<257::AID-CA5>3.0.CO;2-5. [DOI] [PubMed] [Google Scholar]
- 20.Koshino T, Murakami G, Sato TJ, Tsugane MH, Fujisawa Y, Mawatari T, Abe T. Configurations of the segmental and subsegmental bronchi and arteries in the right upper lobe of the human lung with special reference to their concomitant relations and double subsegmental arterial supply. Anat Sci Int. 2002;77:64–73. doi: 10.1046/j.0022-7722.2002.00010.x. [DOI] [PubMed] [Google Scholar]