

Abstract
Seizures that are resistant to standard medications remain a major clinical problem. One underutilized option for patients with medication-resistant seizures is the high-fat, low-carbohydrate ketogenic diet. The diet received its name based on the observation that patients consuming this diet produce ketone bodies (e.g., acetoacetate, β-hydroxybutyrate, and acetone). Although the exact mechanisms of the diet are unknown, ketone bodies have been hypothesized to contribute to the anticonvulsant and antiepileptic effects. In this review, anticonvulsant properties of ketone bodies and the ketogenic diet are discussed (including GABAergic and glutamatergic effects). Because of the importance of ketone body metabolism in the early stages of life, the effects of ketone bodies on developing neurons in vitro also are discussed. Understanding how ketone bodies exert their effects will help optimize their use in treating epilepsy and other neurological disorders.
Keywords: seizure, epilepsy, ketogenic diet, acetoacetate, β-hydroxybutyrate, acetone
Introduction
Epilepsy is a devastating and widespread disease that affects around 1% of the U.S. population and at least 50 million people worldwide (Hauser & Kurland 1975, Hirtz et al. 2007, Kobau et al. 2008). While an array of antiepileptic drugs has been developed, they remain unable to control seizures in approximately 30% of patients with epilepsy (Kwan & Brodie 2000). Some of these patients are candidates for surgical resection of abnormal tissue (e.g., mesial temporal sclerosis) but options are limited for those without such lesions. To help these patients with nonsurgical intractable epilepsy, a high-fat, low-carbohydrate diet known as a ketogenic diet was developed in the 1920s to mimic the beneficial effects of fasting on seizure control (Hartman & Vining 2007). Many clinical studies have confirmed its beneficial effects, with a randomized study showing a 75% decrease in seizures in children on a ketogenic diet for three months (Neal et al. 2008). However, while many possibilities have been investigated, the mechanisms through which a ketogenic diet confers its anticonvulsant effects remain unknown. Unraveling these mechanisms will allow refinement of its clinical use as well as identification of new targets for future therapy development.
A ketogenic diet receives its name due to the high ketone levels found in the blood and urine of patients consuming the diet, which traditionally consists of either long- or medium-chain fatty acids (Hartman & Vining 2007). In the setting of elevated fatty acids and low dietary carbohydrate content while on a ketogenic diet, the liver produces ketone bodies by shunting excess acetyl-CoA to ketogenesis (Fig. 1). The two primary ketone bodies produced are acetoacetate and β-hydroxybutyrate. Acetone, the other major ketone elevated in patients on a ketogenic diet, is a metabolite of acetoacetate. During infancy, the brain is much more efficient at extracting and utilizing ketone bodies from the blood due to the higher levels of ketone metabolizing enzymes and monocarboxylic acid transporters produced during this period (Sokoloff 1973, Bilger & Nehlig 1992). Notably, however, the adult brain increases its levels of monocarboxylic acid transporters and ketone metabolizing enzymes rapidly during periods of stress such as ischemia, trauma, or low glucose (Prins 2008). In addition, there is evidence that astrocytes can produce ketone bodies under conditions of glucose deprivation (Blazquez et al. 1999). Thus, given the brain's ability to adapt to ketone utilization, the elevated levels of ketone bodies in patients on a ketogenic diet, and the robust anticonvulsant effects observed in these patients, ketone bodies are an attractive candidate to investigate in studies of a ketogenic diet's anticonvulsant mechanism.
Figure 1.
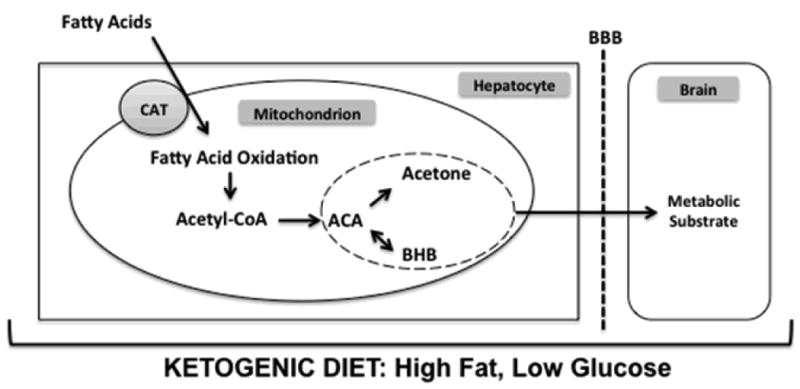
A high-fat, low-carbohydrate ketogenic diet causes a shift in the metabolic activity of hepatocytes. Under these conditions, the Krebs cycle can not utilize the high levels of acetyl-CoA generated from fat. Remaining acetyl-CoA is converted to the ketone body acetoacetate. The two additional ketone bodies, acetone and β-hydroxybutyrate, are derived from acetoacetate by spontaneous degradation and enzymatic conversion with β-hydroxybutyrate dehydrogenase, respectively. The three ketone bodies are then released from the hepatocyte, cross the blood brain barrier, and may exert their effects in the brain. Abbreviations: CAT, carnitine-acylcarnitine translocase; ACA, acetoacetate; BHB, β-hydroxybutyrate; BBB, blood brain barrier.
Ketone Bodies: Anticonvulsant Properties
In trying to understand a ketogenic diet's therapeutic mechanism, many laboratories have investigated whether ketone bodies at pharmacological doses have intrinsic anticonvulsant properties. The anticonvulsant properties of acetoacetate were first reported by Keith in 1935 where its administration in rabbits protected against thujone-induced seizures (Keith 1935). In agreement with this early report, Rho and colleagues found that intraperitoneally administered acetoacetate and acetone, but not β-hydroxybutyrate, protected against seizures in the Frings audiogenic seizure-susceptible juvenile mouse (Rho et al. 2002). More recently, acute acetone administration by intraperitoneal injection in rats decreased their seizure susceptibility in a broad range of seizure tests including the maximal electroshock test, the subcutaneous pentylenetetrazol test, the amygdala kindling test, and the AY-9944 test (Likhodii et al. 2003). Acutely administered acetone, but not its metabolites, protected juvenile mice from seizures induced by pentylenetetrazol and 4-aminopyridine (Gasior et al. 2007). However, in this study, the doses of acetone necessary to cause significant protection from seizures caused motor impairment in the mice, limiting its clinical pharmacological implementation. Collectively, the evidence is strong that acutely administered acetone and acetoacetate have intrinsic anticonvulsant properties in many standardized animal models of epilepsy. However, clinical translatability remains unclear. We are unaware of reports examining the anticonvulsant effects of chronic exogenously-administered ketone bodies in models similar to those discussed previously.
While acetone has emerged as a broad-spectrum anticonvulsant in animal models, a ketogenic diet in the same tests has not consistently conferred a similar level of protection (specifically, against pentylenetetrazol-induced seizures in mice) suggesting that acetone and the ketogenic diet may have different anticonvulsant mechanisms (Uhlemann & Neims 1972, Samala et al. 2008). Additionally, plasma levels of acetone in rats fed a ketogenic diet were 10-fold lower than the levels found to be therapeutic (>2mM), even in the presence of an inhibitor of acetone metabolism (Nylen et al. 2006). In contrast, using proton magnetic resonance spectroscopy in epilepsy patients fully controlled by a ketogenic diet, brain acetone levels were approximately 0.7 mM (Seymour et al. 1999). These data suggest that humans and rats may produce acetone at different levels in response to a ketogenic diet, possibly explaining the different anticonvulsant profiles observed in animal seizure models using acetone versus a ketogenic diet (Rho et al. 2002). To better understand the observed inconsistency (and the possibility that acetone's volatility may lead to inaccurate estimates of actual brain exposure), studies comparing levels of acetone in the brain during a ketogenic diet versus those found with exogenously administered acetone are needed. At this time, acetone's role in the anticonvulsant mechanisms of the ketogenic diet remains unclarified.
Ketone Bodies: Possible Mechanisms of Action
The in vivo studies discussed previously suggest that ketone bodies have anticonvulsant properties. A number of theories have emerged to explain the decreased seizure threshold in these models. Glutamate and γ-aminobutyric acid (GABA), respectively, are the major excitatory and inhibitory neurotransmitters in the brain. How ketone bodies affect levels of these neurotransmitters and their receptors, their receptor activity, and their handling has been debated. New theories also are emerging surrounding the effects of ketone bodies on neuronal membrane potential, neuronal excitability, and reactive oxygen species. Presented here is an overview of the current data evaluating these evolving hypotheses.
GABA
GABA signaling has remained a key target of investigation since rodent models of epilepsy induced by GABA antagonists have a strong response to a ketogenic diet (Uhlemann & Neims 1972, Bough et al. 2002). The role of GABA signaling in the anticonvulsant mechanisms of ketone bodies and/or the ketogenic diet is highly debated. Supporting a role for GABA signaling, GABA levels in rat synaptosomes exposed to acetoacetate or β-hydroxybutyrate are increased (Erecinska et al. 1996). More recently, clinical studies of patients on a ketogenic diet show increased GABA levels in the cerebrospinal fluid, as well as with magnetic resonance spectroscopy (Wang et al. 2003, Dahlin et al. 2005). GABA is synthesized from the Krebs cycle intermediate α-ketoglutarate, which, via glutamate, is converted to GABA by glutamate decarboxylase. Aspartate is a known inhibitor of glutamate decarboxylase. Therefore, a decrease in aspartate could theoretically promote the synthesis of GABA. Exploring this further, a one-hour exposure to acetoacetate or β-hydroxybutyrate was found to decrease aspartate levels in cultured astrocytes (Yudkoff et al. 1997). A similar decrease in aspartate levels was detected in the forebrain and cerebellum of mice fed a ketogenic diet (Yudkoff et al. 2001). The body of data from Yudkoff's group suggest a model wherein the ketotic brain has increased conversion of glutamate to glutamine in astrocytes; glutamine is then taken up into neurons, where it is then converted eventually to GABA (preferentially over aspartate), thus increasing neuronal inhibition (Yudkoff et al. 2008). One unanswered question about the importance of GABA is why a ketogenic diet did not protect murine models of epilepsy induced by the GABA antagonist pentylenetatrazol in the same way it did rats (Uhlemann & Neims 1972). Furthermore, a number of studies have shown no changes in GABA levels from whole brain samples of rodents fed a ketogenic diet, seemingly inconsistent with an increase in GABA synthesis (Yudkoff et al. 2001, Yudkoff et al. 2005, Melo et al. 2006, Appleton & DeVivo 1974, Al-Mudallal et al. 1996). Additional studies are required to investigate whether there are local or regional changes in GABA levels, GABA shunt flow, or GABA responsiveness within the brain in the presence of ketone bodies.
VGLUTs
Ketone bodies may alter the behavior of vesicular glutamate transporters (VGLUTs) that are responsible for filling pre-synaptic vesicles with glutamate in a Cl--dependent manner. Using proteoliposomes as an in vitro model, Juge and colleagues demonstrated that Cl- is an allosteric activator of VGLUTs that is competitively inhibited by ketone bodies, acetoacetate more strongly than β-hydroxybutyrate (Juge et al. 2010). Strengthening their in vitro findings, they showed that acetoacetate reversibly inhibited glutamate release from both cultured rat neurons and mouse CA1 pyramidal neurons in hippocampal slices. Finally, acetoacetate was able to suppress in vivo glutamate release and seizures in rat brains exposed to 4-aminopyridine. However, the amount of acetoacetate that the authors used (10 mM) is likely supratherapeutic, given that hippocampal extracellular levels of β-hydroxybutyrate are 40-50 μM in mice fed a ketogenic diet for 2-3 weeks (Samala et al. 2011). As these two compounds have a stoichiometric relationship (Owen et al. 1967), it is expected that levels of acetoacetate would be significantly less than 10mM in these mice. In addition, children on a ketogenic diet have been found to have CSF levels of acetoacetate at 0.3mM, much lower than the concentrations utilized in the Juge et al. study (Nordli & De Vivo 1997). Therefore, the clinical relevance of higher levels used by Juge et al. needs to be further clarified.
Other neurotransmitters, transporters, and receptors
Controversy remains over how ketone bodies affect neurotransmitter responsiveness. Available data suggest that a ketogenic diet does not alter levels of the neuronal glutamate transporter EAAC1 in neocortex, hippocampus, or cerebellum, measured by Western blot, electron microscopy and glutamate uptake in hippocampal proteoliposomes (Bough et al. 2007). In vitro patch clamp studies of post-synaptic GABAA receptors on rat hippocampal neurons suggest that there is no change in receptor activity in the presence of extracellular 2mM β-hydroxybutyrate or 1mM acetoacetate (Thio et al. 2000). In these studies, the ketone body containing solutions, which also contained glucose, were pre-incubated with the neurons for only one minute. Examining the behavior of the receptors following longer-term incubation with ketone bodies in the absence of glucose would be of interest as these conditions model more closely the clinical scenario in patients on a ketogenic diet. More recent work using Xenopus oocytes expressing human GABAA receptors shows that a 5-minute incubation with 10mM β-hydroxybutyrate or 50 mM acetone significantly increases GABAA receptor activity (Yang et al. 2007). While intriguing, Yang and colleagues did not find similar changes at lower concentrations of the ketone bodies that would be seen clinically. Good in vivo data to clarify what role, if any, that altered GABA receptor responsiveness plays in the anticonvulsant activity of ketone bodies, are lacking.
Other neurotransmitter systems not typically considered in epilepsy also appear to be necessary for the ketogenic diet's anticonvulsant effects. Data for an effect of norepinephrine includes the lack of a ketogenic diet's protective effects against the convulsant fluorothyl in dopamine beta-hydroxylase knockout mice that cannot synthesize norepinephrine (Szot et al. 2001). Evidence of the need for adenosine neurotransmission was shown when mice with adenosine A1 receptor mutations were not protected against recurrent seizures after ketogenic diet treatment (Masino et al. 2011). A ketogenic diet also decreased the adenosine metabolizing enzyme, adenosine kinase. Other neurotransmitters have not been investigated as thoroughly.
KATP channels
Another way ketone bodies may be anticonvulsant is through a direct effect on neuronal membrane potential. KATP channels are of particular interest given their close relationship with cellular metabolism (activated when cellular adenosine tri-phosphate (ATP) is low) and their hyperpolarizing effects. Additionally, they were found to play a key role in CA3 hippocampal neuron hyperpolarization when the neurons were cultured in low glucose media (Kawamura et al. 2010). Exploring this idea further, in cultured mouse hippocampal dentate gyrus neurons, both basal levels of inward-rectifying KATP channel opening and the amount of stimulus evoked KATP channel opening was increased in the presence of β-hydroxybutyrate (Tanner et al. 2011). Adding support to this in vitro work, live recordings of the hippocampal dentate gyrus in rats fed a ketogenic diet show decreased long-term potentiation, consistent with increased neuronal inhibition (Koranda et al. 2011). Previously, Ma and colleagues used GABAergic substantia nigra pars reticulata (SNr) neurons to study KATP channels given the hypothesis that very rapid firing of these neurons precede generalized seizure onset (Ma et al. 2007, Deransart et al. 2003). When GABAergic SNr neurons were cultured in the presence of acetoacetate or β-hydroxybutyrate with glucose, firing rates (measured by loose-patch cell-attached recordings) were decreased. Of note, this effect was abolished in a variety of settings, including (a) when the neurons were cultured from Kir6.2 knockout mice (no cell surface KATP channels) and (b) when the experiments were conducted in the presence of tolbutamide (an inhibitor of KATP channel opening) or a GABAB receptor blocker. These findings add further support to the hypothesis that the anticonvulsant activities of ketone bodies are, in part, mediated by their effects on KATP channels. The latter scenario suggests that GABAB signaling might be important to mediate these effects of ketone bodies. However, whether these results can shed light on mechanisms of the ketogenic diet needs to be considered carefully as rodent studies to date have shown that a ketogenic diet elevates or maintains normal cellular ATP levels, an otherwise negative regulator of KATP channels (DeVivo et al. 1978, Bough et al. 2006, Nylen et al. 2009). The notion has been advanced that a ketogenic diet enhances KATP channel activity through ATP compartmentation by decreasing glycolytic ATP levels, an hypothesis that awaits further experimental evidence (Yellen 2008).
Krebs cycle and the electron transport chain
One theory of the ketogenic diet's multifaceted effects is that it may optimize Krebs cycle function, with a subsequent effect on neuronal function. The logical starting point for this theory is that acetyl CoA (one downstream product of ketogenesis) is a primary substrate for the Krebs cycle (Fig. 1). Downstream of the Krebs cycle, β-hydroxybutyrate improved redox activity and decreased cell death in dopaminergic SH-SY5Y cells after exposure to the complex I toxin rotenone (Imamura et al. 2006). β-hydroxybutyrate appears to require an intact complex II to confer its beneficial effects in two different models of neurodegeneration, including a complex II toxin (3-nitropropionic acid) model and the SOD1-G93A mouse model of amyotrophic lateral sclerosis (Lou Gehrig's disease) (Tieu et al. 2003, Zhao et al. 2006). More recently, Kim et al. showed in hippocampal slices that a cocktail of β-hydroxybutyrate and acetoacetate preserved synaptic activity after exposure to rotenone or 3-nitropropionic acid, although there were some regional disparities (i.e., different responses even within the same slice) noted during ketone exposure to the latter (Kim do et al. 2010). The reason for differences between the two studies of 3-nitropropionic acid is unclear.
A ketogenic diet also has been shown to induce transcription of some electron transport chain subunit mRNAs (Bough et al. 2006). An increased number of electron transport chain subunits might increase neuronal “energy reserves” and, in fact, ketogenic diets increase brain ATP levels in some (but not all) rodent studies (DeVivo et al. 1978, Bough et al. 2006, Nylen et al. 2009, Kim do et al. 2010). A final hypothesis is that increased ATP levels may stabilize neuronal membrane potential by enhancing the function of Na+-K+-ATPases (Bough & Rho 2007). Although teleologically satisfying, the events outlined here have not been completely linked experimentally in either normal or epilepsy animal models. Nonetheless, individual experiments point to the potential importance of the Krebs cycle and electron transport chain function in the effects of ketone bodies.
Immature and developing neurons
Many studies have looked at the effects of ketone bodies on immature neurons as a possible physiological model of the ketogenic diet. In suckling rodents, circulating levels of ketone bodies are high due to the high ketogenic ratio of rodent milk (Nehlig 1999). During this time, the brain relies most heavily on ketone bodies as a source of metabolic fuel (Nehlig & Pereira de Vasconcelos 1993). Also during this time, GABA is excitatory, an action found to be dependent on the higher intracellular chloride levels in immature neurons. As a neuron matures the Na+-K+-2Cl- co-transporter (NKCC1) is replaced with the K+-Cl- co-transporter (KCC2) shifting GABA to an inhibitory neurotransmitter (Aronica et al. 2011). Patch clamp work on neocortical pyramidal neurons from mice demonstrated that β-hydroxybutyrate incubation at supra-physiologic levels decreased both the resting membrane potential and the amount of GABA-mediated excitation in a dose-dependent manner (Rheims et al. 2009). This group also showed that ketone bodies at similarly high levels cause the same inhibition of GABA-mediated excitation in murine hippocampal CA3 neurons (Holmgren et al. 2010), another important brain region in epilepsy development. These data are supported by recent in vivo work examining physiological levels of ketone bodies in neonatal rats (Minlebaev & Khazipov 2011). Minlebaev and Khazipov used 16-channel extracellular field potential recordings from somatosensory barrel cortex in postnatal day 5-9 awake rat pups to study epileptiform activity induced by flurothyl when physiological ketosis was inhibited. They found that blocking ketogenesis did not affect a single flurothyl-evoked seizure, however consecutively evoked seizures were more severe in these animals. In contrast to the preceding studies, a number of groups have found that GABA-induced depolarization in neonatal rodent cortical neurons is unaffected by the presence of ketone bodies at both supraphysiologic and physiologic levels (Thio et al. 2000, Kirmse et al. 2010, Tyzio et al. 2011). Possible reasons for the discrepancies among the studies are outlined in Tyzio et al. 2011. Thus, while an effect of ketone bodies on neonatal cortex excitability remains an intriguing possibility that could shed light on possible therapeutic mechanisms of a ketogenic diet, its existence remains controversial. These data also should be interpreted with caution when drawing parallels to a ketogenic diet as all these results were found with solutions containing both ketone bodies and high levels of glucose (5-10 mM). Basal glucose concentrations in murine hippocampal extracellular fluid as determined by in vivo microdialysis were approximately 2 mM, and only ∼1.3 mM when the mice were fed a ketogenic diet (Samala et al. 2011).
Antioxidant effects
Ketone bodies also may have antioxidant effects. Reactive oxygen species production resulting from glutamate exposure is inhibited by ketone bodies in primary cultures of rat neocortical neurons (Maalouf et al. 2007). The mechanism appears to involve NADH oxidation, rather than a glutathione-mediated antioxidant effect. Conversely, the ketogenic diet (rather than ketone bodies) may alter glutathione metabolism via the transcription factor Nrf2 (Milder et al. 2010). Ketone bodies also protect against cell death after neocortical slices are exposed to hydrogen peroxide (Kim do et al. 2007). Additionally, ketone bodies prevent hydrogen peroxide-induced impairment of hippocampal long-term potentiation (Maalouf & Rho 2008). Therefore, in addition to effects on neurotransmission, these data suggest ketone bodies may be neuroprotective against reactive oxygen species produced in neuropathological states.
Conclusions
A growing amount of in vivo and in vitro data show that ketone bodies can alter neuronal activity and protect against seizures. The mechanisms through which ketone bodies may confer an anticonvulsant effect are still debated. The potential mechanisms discussed in this review center around neurotransmitters and the resting neuronal membrane potential (Fig. 2). Moving forward, work will need to be completed exploring how physiological levels of ketone bodies affect these systems. The exact targets of ketone bodies still are unknown and may be direct (i.e., binding to a specific neuronal target, as demonstrated for acetoacetate in VGLUT), or indirect (i.e., supplying carbon equivalents to the Krebs cycle or an antioxidant effect). Given both the success of AC-1202 (an oral ketogenic compound that rapidly elevates β-hydroxybutyrate levels) in improving cognitive outcomes in Alzheimer disease patients (Henderson et al. 2009) and the success of a ketogenic diet in treating children with medically intractable epilepsy, there is significant interest in pursuing these types of compounds for epilepsy therapies. However, the contribution of ketone bodies to the anticonvulsant effects of the ketogenic diet remains elusive. Because of study design issues and the amount of overlap between values in different patient groups, the relationship between seizure control and serum levels of β-hydroxybutyrate remains unclear (Gilbert et al. 2000). Many other studies have failed to show that the levels of serum or urine ketosis correlate with seizure protection in animal models (Likhodii et al. 2000, Bough et al. 2000) or patients (Ross et al. 1985, Fraser et al. 2003). Breath acetone levels correlate with plasma levels of all three ketone bodies but not with seizure control (Musa-Veloso et al. 2006). Some of these studies highlight the potential difference between administration of ketone bodies versus a ketogenic diet (understanding the former as a product of the latter). Together, some of the data discussed here suggest the ketogenic diet may have anticonvulsant effects via mechanisms other than elevated ketone body levels, even though ketone bodies have numerous potential beneficial effects. Therefore, based on current data, ketone bodies may represent a unique, additional therapeutic modality to optimize for patients with epilepsy and possibly other neurological disorders. Further studies are required to understand how using these compounds may help these populations.
Figure 2.
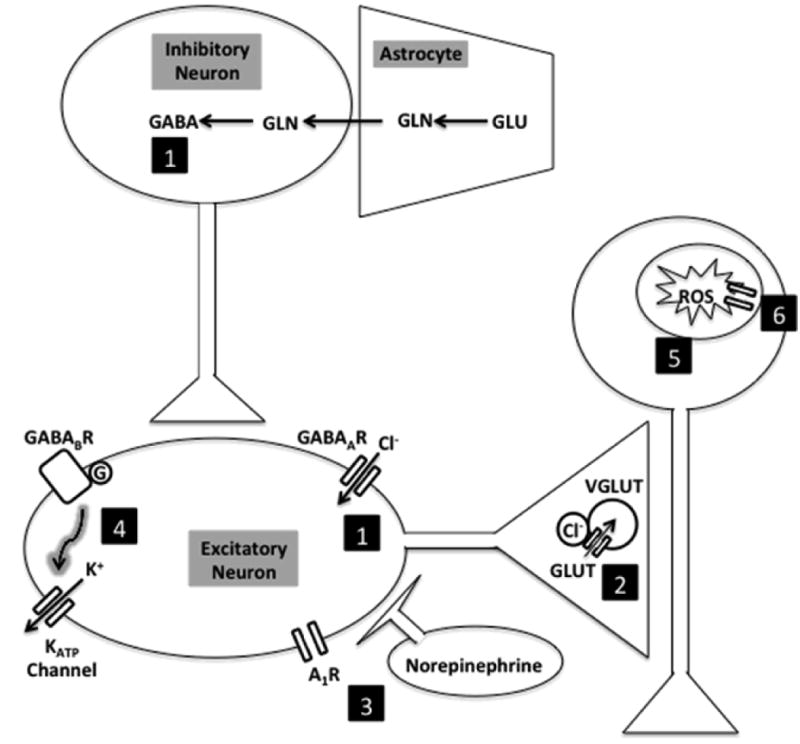
Possible anticonvulsant effects of ketone bodies on the brain. (1) Increased GABA synthesis through alteration of glutamate cycling in glutamate-glutamine cycle or altered neuronal responsiveness to GABA at GABAA receptors. (2) Decreased glutamate release by competitive inhibition of vesicular glutamate transporters. (3) Other neurotransmitters, including norepinephrine and adenosine. (4) Increased membrane potential hyperpolarization via KATP channels possibly mediated by GABAB receptor signaling. (5) Decreased reactive oxygen species production from glutamate exposure. (6) Electron transport chain subunit transcription. Abbreviations: A1R, adenosine receptor; Cl, chloride; GLN, glutamine; GLU, glutamate; GABA, γ-aminobutyric acid ; GABABR, γ-aminobutyric acid beta receptor; GABAAR, γ-aminobutyric acid alpha receptor; VGLUT, vesicular glutamate transporter; ROS, reactive oxygen species.
Acknowledgments
Supported by the Howard Hughes Medical Institute Medical Research Fellows Program (MAM), K12NS001696, K08NS070931, and a Clinician Scientist Award (Johns Hopkins University School of Medicine) (ALH). The authors have no conflicts of interest to disclose.
References
- Al-Mudallal AS, LaManna JC, Lust WD, Harik SI. Diet-induced ketosis does not cause cerebral acidosis. Epilepsia. 1996;37:258–261. doi: 10.1111/j.1528-1157.1996.tb00022.x. [DOI] [PubMed] [Google Scholar]
- Appleton DB, DeVivo DC. An animal model for the ketogenic diet. Epilepsia. 1974;15:211–227. doi: 10.1111/j.1528-1157.1974.tb04943.x. [DOI] [PubMed] [Google Scholar]
- Aronica E, Iyer A, Zurolo E, Gorter JA. Ontogenetic modifications of neuronal excitability during brain maturation: developmental changes of neurotransmitter receptors. Epilepsia. 2011;52(Suppl 8):3–5. doi: 10.1111/j.1528-1167.2011.03222.x. [DOI] [PubMed] [Google Scholar]
- Bilger A, Nehlig A. Quantitative histochemical changes in enzymes involved in energy metabolism in the rat brain during postnatal development. II. Glucose-6-phosphate dehydrogenase and beta-hydroxybutyrate dehydrogenase. Int J Dev Neurosci. 1992;10:143–152. doi: 10.1016/0736-5748(92)90042-x. [DOI] [PubMed] [Google Scholar]
- Blazquez C, Woods A, de Ceballos ML, Carling D, Guzman M. The AMP-activated protein kinase is involved in the regulation of ketone body production by astrocytes. J Neurochem. 1999;73:1674–1682. doi: 10.1046/j.1471-4159.1999.731674.x. [DOI] [PubMed] [Google Scholar]
- Bough KJ, Gudi K, Han FT, Rathod AH, Eagles DA. An anticonvulsant profile of the ketogenic diet in the rat. Epilepsy Res. 2002;50:313–325. doi: 10.1016/s0920-1211(02)00086-4. [DOI] [PubMed] [Google Scholar]
- Bough KJ, Paquet M, Pare JF, Hassel B, Smith Y, Hall RA, Dingledine R. Evidence against enhanced glutamate transport in the anticonvulsant mechanism of the ketogenic diet. Epilepsy Res. 2007;74:232–236. doi: 10.1016/j.eplepsyres.2007.03.002. [DOI] [PubMed] [Google Scholar]
- Bough KJ, Rho JM. Anticonvulsant mechanisms of the ketogenic diet. Epilepsia. 2007;48:43–58. doi: 10.1111/j.1528-1167.2007.00915.x. [DOI] [PubMed] [Google Scholar]
- Bough KJ, Wetherington J, Hassel B, et al. Mitochondrial biogenesis in the anticonvulsant mechanism of the ketogenic diet. Ann Neurol. 2006;60:223–235. doi: 10.1002/ana.20899. [DOI] [PubMed] [Google Scholar]
- Bough KJ, Yao SG, Eagles DA. Higher ketogenic diet ratios confer protection from seizures without neurotoxicity. Epilepsy Res. 2000;38:15–25. doi: 10.1016/s0920-1211(99)00077-7. [DOI] [PubMed] [Google Scholar]
- Dahlin M, Elfving A, Ungerstedt U, Amark P. The ketogenic diet influences the levels of excitatory and inhibitory amino acids in the CSF in children with refractory epilepsy. Epilepsy Res. 2005;64:115–125. doi: 10.1016/j.eplepsyres.2005.03.008. [DOI] [PubMed] [Google Scholar]
- Deransart C, Hellwig B, Heupel-Reuter M, Leger JF, Heck D, Lucking CH. Single-unit analysis of substantia nigra pars reticulata neurons in freely behaving rats with genetic absence epilepsy. Epilepsia. 2003;44:1513–1520. doi: 10.1111/j.0013-9580.2003.26603.x. [DOI] [PubMed] [Google Scholar]
- DeVivo DC, Leckie MP, Ferrendelli JS, McDougal DB., Jr Chronic ketosis and cerebral metabolism. Ann Neurol. 1978;3:331–337. doi: 10.1002/ana.410030410. [DOI] [PubMed] [Google Scholar]
- Erecinska M, Nelson D, Daikhin Y, Yudkoff M. Regulation of GABA level in rat brain synaptosomes: fluxes through enzymes of the GABA shunt and effects of glutamate, calcium, and ketone bodies. J Neurochem. 1996;67:2325–2334. doi: 10.1046/j.1471-4159.1996.67062325.x. [DOI] [PubMed] [Google Scholar]
- Fraser DD, Whiting S, Andrew RD, Macdonald EA, Musa-Veloso K, Cunnane SC. Elevated polyunsaturated fatty acids in blood serum obtained from children on the ketogenic diet. Neurology. 2003;60:1026–1029. doi: 10.1212/01.wnl.0000049974.74242.c6. [DOI] [PubMed] [Google Scholar]
- Gasior M, French A, Joy MT, Tang RS, Hartman AL, Rogawski MA. The anticonvulsant activity of acetone, the major ketone body in the ketogenic diet, is not dependent on its metabolites acetol, 1,2-propanediol, methylglyoxal, or pyruvic acid. Epilepsia. 2007;48:793–800. doi: 10.1111/j.1528-1167.2007.01026.x. [DOI] [PubMed] [Google Scholar]
- Gilbert DL, Pyzik PL, Freeman JM. The ketogenic diet: seizure control correlates better with serum beta-hydroxybutyrate than with urine ketones. J Child Neurol. 2000;15:787–790. doi: 10.1177/088307380001501203. [DOI] [PubMed] [Google Scholar]
- Hartman AL, Vining EP. Clinical aspects of the ketogenic diet. Epilepsia. 2007;48:31–42. doi: 10.1111/j.1528-1167.2007.00914.x. [DOI] [PubMed] [Google Scholar]
- Hauser WA, Kurland LT. The epidemiology of epilepsy in Rochester, Minnesota, 1935 through 1967. Epilepsia. 1975;16:1–66. doi: 10.1111/j.1528-1157.1975.tb04721.x. [DOI] [PubMed] [Google Scholar]
- Henderson ST, Vogel JL, Barr LJ, Garvin F, Jones JJ, Costantini LC. Study of the ketogenic agent AC-1202 in mild to moderate Alzheimer's disease: a randomized, double-blind, placebo-controlled, multicenter trial. Nutr Metab (Lond) 2009;6:31. doi: 10.1186/1743-7075-6-31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hirtz D, Thurman DJ, Gwinn-Hardy K, Mohamed M, Chaudhuri AR, Zalutsky R. How common are the “common” neurologic disorders? Neurology. 2007;68:326–337. doi: 10.1212/01.wnl.0000252807.38124.a3. [DOI] [PubMed] [Google Scholar]
- Holmgren CD, Mukhtarov M, Malkov AE, Popova IY, Bregestovski P, Zilberter Y. Energy substrate availability as a determinant of neuronal resting potential, GABA signaling and spontaneous network activity in the neonatal cortex in vitro. J Neurochem. 2010;112:900–912. doi: 10.1111/j.1471-4159.2009.06506.x. [DOI] [PubMed] [Google Scholar]
- Imamura K, Takeshima T, Kashiwaya Y, Nakaso K, Nakashima K. D-beta-hydroxybutyrate protects dopaminergic SH-SY5Y cells in a rotenone model of Parkinson's disease. J Neurosci Res. 2006;84:1376–1384. doi: 10.1002/jnr.21021. [DOI] [PubMed] [Google Scholar]
- Juge N, Gray JA, Omote H, et al. Metabolic control of vesicular glutamate transport and release. Neuron. 2010;68:99–112. doi: 10.1016/j.neuron.2010.09.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kawamura M, Jr, Ruskin DN, Masino SA. Metabolic autocrine regulation of neurons involves cooperation among pannexin hemichannels, adenosine receptors, and KATP channels. J Neurosci. 2010;30:3886–3895. doi: 10.1523/JNEUROSCI.0055-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keith H. Experimental convulsions induced by administration of thujone. Arch Neurol Psychiatry. 1935;34:1022–1040. [Google Scholar]
- Kim do Y, Davis LM, Sullivan PG, Maalouf M, Simeone TA, van Brederode J, Rho JM. Ketone bodies are protective against oxidative stress in neocortical neurons. J Neurochem. 2007;101:1316–1326. doi: 10.1111/j.1471-4159.2007.04483.x. [DOI] [PubMed] [Google Scholar]
- Kim do Y, Vallejo J, Rho JM. Ketones prevent synaptic dysfunction induced by mitochondrial respiratory complex inhibitors. J Neurochem. 2010;114:130–141. doi: 10.1111/j.1471-4159.2010.06728.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kirmse K, Witte OW, Holthoff K. GABA depolarizes immature neocortical neurons in the presence of the ketone body ss-hydroxybutyrate. J Neurosci. 2010;30:16002–16007. doi: 10.1523/JNEUROSCI.2534-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kobau R, Zahran H, Thurman DJ, Zack MM, Henry TR, Schachter SC, Price PH. Epilepsy surveillance among adults--19 States, Behavioral Risk Factor Surveillance System, 2005. MMWR Surveill Summ. 2008;57:1–20. [PubMed] [Google Scholar]
- Koranda JL, Ruskin DN, Masino SA, Blaise JH. A ketogenic diet reduces long-term potentiation in the dentate gyrus of freely behaving rats. J Neurophysiol. 2011;106:662–666. doi: 10.1152/jn.00001.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kwan P, Brodie MJ. Epilepsy after the first drug fails: substitution or add-on? Seizure. 2000;9:464–468. doi: 10.1053/seiz.2000.0442. [DOI] [PubMed] [Google Scholar]
- Likhodii SS, Musa K, Mendonca A, Dell C, Burnham WM, Cunnane SC. Dietary fat, ketosis, and seizure resistance in rats on the ketogenic diet. Epilepsia. 2000;41:1400–1410. doi: 10.1111/j.1528-1157.2000.tb00115.x. [DOI] [PubMed] [Google Scholar]
- Likhodii SS, Serbanescu I, Cortez MA, Murphy P, Snead OC, 3rd, Burnham WM. Anticonvulsant properties of acetone, a brain ketone elevated by the ketogenic diet. Ann Neurol. 2003;54:219–226. doi: 10.1002/ana.10634. [DOI] [PubMed] [Google Scholar]
- Ma W, Berg J, Yellen G. Ketogenic diet metabolites reduce firing in central neurons by opening K(ATP) channels. J Neurosci. 2007;27:3618–3625. doi: 10.1523/JNEUROSCI.0132-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maalouf M, Rho JM. Oxidative impairment of hippocampal long-term potentiation involves activation of protein phosphatase 2A and is prevented by ketone bodies. J Neurosci Res. 2008;86:3322–3330. doi: 10.1002/jnr.21782. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maalouf M, Sullivan PG, Davis L, Kim DY, Rho JM. Ketones inhibit mitochondrial production of reactive oxygen species production following glutamate excitotoxicity by increasing NADH oxidation. Neuroscience. 2007;145:256–264. doi: 10.1016/j.neuroscience.2006.11.065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Masino SA, Li T, Theofilas P, Sandau US, Ruskin DN, Fredholm BB, Geiger JD, Aronica E, Boison D. A ketogenic diet suppresses seizures in mice through adenosine A receptors. J Clin Invest. 2011;121:2679–2683. doi: 10.1172/JCI57813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Melo TM, Nehlig A, Sonnewald U. Neuronal-glial interactions in rats fed a ketogenic diet. Neurochem Int. 2006;48:498–507. doi: 10.1016/j.neuint.2005.12.037. [DOI] [PubMed] [Google Scholar]
- Milder JB, Liang LP, Patel M. Acute oxidative stress and systemic Nrf2 activation by the ketogenic diet. Neurobiol Dis. 2010;40:238–244. doi: 10.1016/j.nbd.2010.05.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Minlebaev M, Khazipov R. Antiepileptic effects of endogenous beta-hydroxybutyrate in suckling infant rats. Epilepsy Res. 2011;95:100–109. doi: 10.1016/j.eplepsyres.2011.03.003. [DOI] [PubMed] [Google Scholar]
- Musa-Veloso K, Likhodii SS, Rarama E, et al. Breath acetone predicts plasma ketone bodies in children with epilepsy on a ketogenic diet. Nutrition. 2006;22:1–8. doi: 10.1016/j.nut.2005.04.008. [DOI] [PubMed] [Google Scholar]
- Neal EG, Chaffe H, Schwartz RH, Lawson MS, Edwards N, Fitzsimmons G, Whitney A, Cross JH. The ketogenic diet for the treatment of childhood epilepsy: a randomised controlled trial. Lancet Neurol. 2008;7:500–506. doi: 10.1016/S1474-4422(08)70092-9. [DOI] [PubMed] [Google Scholar]
- Nehlig A. Age-dependent pathways of brain energy metabolism: the suckling rat, a natural model of the ketogenic diet. Epilepsy Res. 1999;37:211–221. doi: 10.1016/s0920-1211(99)00073-x. [DOI] [PubMed] [Google Scholar]
- Nehlig A, Pereira de Vasconcelos A. Glucose and ketone body utilization by the brain of neonatal rats. Prog Neurobiol. 1993;40:163–221. doi: 10.1016/0301-0082(93)90022-k. [DOI] [PubMed] [Google Scholar]
- Nordli DR, Jr, De Vivo DC. The ketogenic diet revisited: back to the future. Epilepsia. 1997;38:743–749. doi: 10.1111/j.1528-1157.1997.tb01460.x. [DOI] [PubMed] [Google Scholar]
- Nylen K, Likhodii SS, Hum KM, Burnham WM. A ketogenic diet and diallyl sulfide do not elevate afterdischarge thresholds in adult kindled rats. Epilepsy Res. 2006;71:23–31. doi: 10.1016/j.eplepsyres.2006.05.008. [DOI] [PubMed] [Google Scholar]
- Nylen K, Velazquez JL, Sayed V, Gibson KM, Burnham WM, Snead OC., 3rd The effects of a ketogenic diet on ATP concentrations and the number of hippocampal mitochondria in Aldh5a1(-/-) mice. Biochim Biophys Acta. 2009;1790:208–212. doi: 10.1016/j.bbagen.2008.12.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Owen OE, Morgan AP, Kemp HG, Sullivan JM, Herrera MG, Cahill GF., Jr Brain metabolism during fasting. J Clin Invest. 1967;46:1589–1595. doi: 10.1172/JCI105650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prins ML. Cerebral metabolic adaptation and ketone metabolism after brain injury. J Cereb Blood Flow Metab. 2008;28:1–16. doi: 10.1038/sj.jcbfm.9600543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rheims S, Holmgren CD, Chazal G, Mulder J, Harkany T, Zilberter T, Zilberter Y. GABA action in immature neocortical neurons directly depends on the availability of ketone bodies. J Neurochem. 2009;110:1330–1338. doi: 10.1111/j.1471-4159.2009.06230.x. [DOI] [PubMed] [Google Scholar]
- Rho JM, Anderson GD, Donevan SD, White HS. Acetoacetate, acetone, and dibenzylamine (a contaminant in l-(+)-beta-hydroxybutyrate) exhibit direct anticonvulsant actions in vivo. Epilepsia. 2002;43:358–361. doi: 10.1046/j.1528-1157.2002.47901.x. [DOI] [PubMed] [Google Scholar]
- Ross DL, Swaiman KF, Torres F, Hansen J. Early biochemical and EEG correlates of the ketogenic diet in children with atypical absence epilepsy. Pediatr Neurol. 1985;1:104–108. doi: 10.1016/0887-8994(85)90045-1. [DOI] [PubMed] [Google Scholar]
- Samala R, Klein J, Borges K. The ketogenic diet changes metabolite levels in hippocampal extracellular fluid. Neurochem Int. 2011;58:5–8. doi: 10.1016/j.neuint.2010.10.011. [DOI] [PubMed] [Google Scholar]
- Samala R, Willis S, Borges K. Anticonvulsant profile of a balanced ketogenic diet in acute mouse seizure models. Epilepsy Res. 2008;81:119–127. doi: 10.1016/j.eplepsyres.2008.05.001. [DOI] [PubMed] [Google Scholar]
- Seymour KJ, Bluml S, Sutherling J, Sutherling W, Ross BD. Identification of cerebral acetone by 1H-MRS in patients with epilepsy controlled by ketogenic diet. MAGMA. 1999;8:33–42. doi: 10.1007/BF02590633. [DOI] [PubMed] [Google Scholar]
- Sokoloff L. Metabolism of ketone bodies by the brain. Annu Rev Med. 1973;24:271–280. doi: 10.1146/annurev.me.24.020173.001415. [DOI] [PubMed] [Google Scholar]
- Szot P, Weinshenker D, Rho JM, Storey TW, Schwartzkroin PA. Norepinephrine is required for the anticonvulsant effect of the ketogenic diet. Brain Res Dev Brain Res. 2001;129:211–214. doi: 10.1016/s0165-3806(01)00213-9. [DOI] [PubMed] [Google Scholar]
- Tanner GR, Lutas A, Martinez-Francois JR, Yellen G. Single K ATP channel opening in response to action potential firing in mouse dentate granule neurons. J Neurosci. 2011;31:8689–8696. doi: 10.1523/JNEUROSCI.5951-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thio LL, Wong M, Yamada KA. Ketone bodies do not directly alter excitatory or inhibitory hippocampal synaptic transmission. Neurology. 2000;54:325–331. doi: 10.1212/wnl.54.2.325. [DOI] [PubMed] [Google Scholar]
- Tieu K, Perier C, Caspersen C, et al. D-beta-hydroxybutyrate rescues mitochondrial respiration and mitigates features of Parkinson disease. J Clin Invest. 2003;112:892–901. doi: 10.1172/JCI18797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tyzio R, Allene C, Nardou R, et al. Depolarizing actions of GABA in immature neurons depend neither on ketone bodies nor on pyruvate. J Neurosci. 2011;31:34–45. doi: 10.1523/JNEUROSCI.3314-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Uhlemann ER, Neims AH. Anticonvulsant properties of the ketogenic diet in mice. The Journal of pharmacology and experimental therapeutics. 1972;180:231–238. [PubMed] [Google Scholar]
- Wang ZJ, Bergqvist C, Hunter JV, Jin D, Wang DJ, Wehrli S, Zimmerman RA. In vivo measurement of brain metabolites using two-dimensional double-quantum MR spectroscopy--exploration of GABA levels in a ketogenic diet. Magn Reson Med. 2003;49:615–619. doi: 10.1002/mrm.10429. [DOI] [PubMed] [Google Scholar]
- Yang L, Zhao J, Milutinovic PS, Brosnan RJ, Eger EI, 2nd, Sonner JM. Anesthetic properties of the ketone bodies beta-hydroxybutyric acid and acetone. Anesth Analg. 2007;105:673–679. doi: 10.1213/01.ane.0000278127.68312.dc. [DOI] [PubMed] [Google Scholar]
- Yellen G. Ketone bodies, glycolysis, and KATP channels in the mechanism of the ketogenic diet. Epilepsia. 2008;49(Suppl 8):80–82. doi: 10.1111/j.1528-1167.2008.01843.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yudkoff M, Daikhin Y, Horyn O, Nissim I. Ketosis and brain handling of glutamate, glutamine, and GABA. Epilepsia. 2008;49(Suppl 8):73–75. doi: 10.1111/j.1528-1167.2008.01841.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yudkoff M, Daikhin Y, Nissim I, Grunstein R. Effects of ketone bodies on astrocyte amino acid metabolism. J Neurochem. 1997;69:682–692. doi: 10.1046/j.1471-4159.1997.69020682.x. [DOI] [PubMed] [Google Scholar]
- Yudkoff M, Daikhin Y, Nissim I, Horyn O, Lazarow A, Luhovyy B, Wehrli S. Response of brain amino acid metabolism to ketosis. Neurochem Int. 2005;47:119–128. doi: 10.1016/j.neuint.2005.04.014. [DOI] [PubMed] [Google Scholar]
- Yudkoff M, Daikhin Y, Nissim I, Lazarow A. Brain amino acid metabolism and ketosis. J Neurosci Res. 2001;66:272–281. doi: 10.1002/jnr.1221. [DOI] [PubMed] [Google Scholar]
- Zhao Z, Lange DJ, Voustianiouk A, et al. A ketogenic diet as a potential novel therapeutic intervention in amyotrophic lateral sclerosis. BMC Neurosci. 2006;7:29. doi: 10.1186/1471-2202-7-29. [DOI] [PMC free article] [PubMed] [Google Scholar]