

Abstract
During a research project aimed at the study of the Culicinae fauna of Gabon and carried out in the National Park of La Lopé, we captured an unknown sandfly male specimen (genus Phlebotomus) by CDC miniature light trap belonging to a new species for Science. Furthermore, the originality of his genitalia does not allow us to include this species in one of the existing subgenus, thus in this paper we propose the creation of a new subgenus, as Phlebotomus (Legeromyia) multihamatus sp. nov., subg. nov. described from the National Park of La Lopé, through one male captured with CDC miniature light trap. A new species and a new subgenus of sandfly is characterised by a short style with three spines, a paramere wearing a basal hook as well as a basal pouch and the absence of basal lobe on the coxite. The originality of the genitalia of the male gives way to discussion about potential primary homologies between P. multihamatus sp. nov. and Phlebotomus (Abonnencius) fortunatarum, Phlebotomus (Anaphlebotomus) stantoni and Phlebotomus (Euphlebotomus) argentipes, which should be verified for future studies. The discovery of this new species in Gabon must encourage the study of sandflies in this country.
Keywords: sandfly, Phlebotomus, new species, Gabon
Despite the former mention of a native case of visceral leishmaniasis (Tournier 1920), Gabon does not actually constitute an endemic country for leishmaniasis (Alvar et al. 2012). As a result, the sandfly fauna in this country remains poorly documented and prospected. Unlike in the neighbouring countries where no less than 23 species in Cameroon, 30 in the Central African Republic, 28 in the Democratic Republic of the Congo and 42 in the Republic of the Congo were previously found (Niang et al. 2004); an exhaustive review of literature reports only four species in Gabon (Galliard & Nitzulescu 1931, Abonnenc 1972): Sergentomyia (Parrotomyia) africana africana (Newstead, 1912), Sergentomyia (Sergentomyia) antennata (Newstead, 1912), Sergentomyia (Sergentomyia) schwetzi (Adler, Theodor & Parrot, 1929) and Spelaeophlebotomus gigas (Parrot & Schwetz, 1937).
MATERIALS AND METHODS
The sandfly described here was collected in a secondary equatorial rainforest inside the National Park of La Lopé, Gabon at approximately 10 km away of the village of Mikongo (0º18'41.34"S 11º42'23.58"E) at an altitude of 303 m above mean sea level (Fig. 1). The insect collections and the access to the Park were authorised by the Ministry of Scientific Research and Technological Development of Gabon (AR0006/12/MENERSI/CENAREST/CG/CST/CSAR) and the National Agency of Natural Parks. The capture was performed in 2012 during the night of the 22 October, using a CDC miniature light trap placed 2 m above the ground level in an open forest undergrowth allowing a good visual attraction.
Fig. 1. location of the collection site in Gabon.
The sandfly was preserved in 100% ethanol then mounted whole in Euparal after successive different baths: 2 h in potassium hydroxide 10%, 2 h in distillated water, 10 h in a Marc-André solution (Abonnenc 1972), 10 h in distillated water, 20 min in ethanol 70%, 20 min in ethanol 90%, 20 min in ethanol 100% and 10 h in a beech wood solution.
Specimen was observed using a BX 50 microscope (Olympus, Japan). Measurements were collected using the Perfect Image software (ARIES Company, Chatillon, France) and a video camera connected to the microscope.
RESULTS
Phlebotomus multihamatus Rahola, Depaquit and Paupy, sp. nov.
(Figs 2-5)
Fig. 2. drawing of palp (A), third segment of palp showing five club-like Newstead spines (B), antennal segment III and IV + V with ascoids (C) and wing (D) of Phlebotomus multihamatus sp. nov.

Male - Holotype - Sandfly of small size (~1.5 mm) and rather light colour.
Head ( Figs 2A-C , 3A ): orientated laterally. Interocular suture, hardly visible with this orientation, seems to be complete. Cibarium with 13 discreet teeth arranged on a concave arc directed backwards and with rare denticles not well defined. Pigmented plate absent. Pharynx fine and short slightly shrunk backwards. Pharyngeal armature not well developed constituted with some wrinkles. Palpal formula: 1-2-4-3-5, third palpal article with five club-like Newstead spines. Ascoid formula: 2/III-IX, articles X-XVI are missing on both antennae. Ascoids long going over the next articulation. Sensillae present on article III-V. A III = 155 µm, A IV = 104 µm, A V = 104 µm, A III < A IV + A V; epipharynx = 143 µm. A III/E = 1.08.
Fig. 3. drawing of pharynx and cibarium (A), mesothoracic part showing mesanepisternal setae (B) and full view of male genitalia (C) of Phlebotomus multihamatus sp. nov.

Thorax ( Figs 2D , 3B ): presence of three antero-inferior mesanepisternal setae. Wing length = 1,205 µm, width = 412 µm, length/width = 2.9º, α = 241 µmº, β = 140 µm, δ = 35, ɣ = 270 µm, wing width/ɣ = 1.5º ad π = 59 µm.
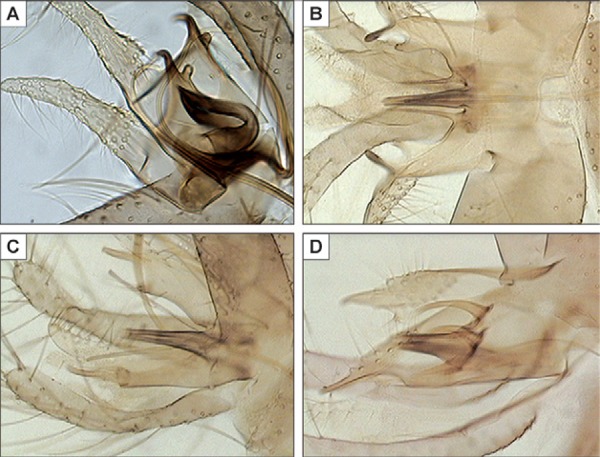
Genitalia ( Figs 3C , 4A-C ): coxite length: 148 µm bearing a small number of scattered setae in its internal face. Style extremely short (60 µm), globular, pear-shaped, with three spines: one terminal and two median. Paramere is very complex measuring 134 µm in its whole. The superior lobe is measuring 84 µm [measurement sensu (Leng & Lewis 1987)]. It bears a very high number of setae. The inferior lobe is subdivided in two parts: (i) a short pointed hook without any setae curved towards the top and pointing backwards and (ii) a small rounded bump bearing three to four setae. At the base of the paramere, near the implantation of the penis, a remarkable process fits in a huge pigmented hook measuring 78 µm long, directed backwards and supplied with a little dewclaw in subapical position. The set composed by this hook and the penis is covered on its external face by a membrane constituting a basal pouch. Surstyle: 168 µm. Penis: 51 µm, wide at its base and contracting regularly to its apex. Apex translucent in its inferior part. Genital pump heavily sclerotised and well developed measuring 114 µm.
Fig. 4. drawing of style (A), paramere with penis and genital filaments (B) and paramere showing pouch surrounding penis (C) of Phlebotomus multihamatus sp. nov.

Female - Unknown.
Type locality - National Park of La Lopé, Gabon.
Type data and depository - Holotype: male: Gabon, National Park of La Lopé 0º18'41.34"S 11º42'23.58"E, 22 October, 2012, CDC miniature light trap, N Rahola, J Depaquit and C Paupy leg. Deposited in the Institut de Recherche pour le Développement, Montpellier, France.
Etymology - The subgenus Legeromyia is dedicated to our colleague Nicole Léger which has greatly contributed to the systematics of sandflies. The species name multihamatus refers to the paramere's hooks that characterise this species.
DISCUSSION
To date, seven genera of sandflies are recorded in the Old World according Rispail and Léger (1998b): Phlebotomus Rondani, 1843, Sergentomyia França & Parrot, 1921, Spelaeophlebotomus Theodor 1948, Spelaeomyia Theodor, 1948, Idiophlebotomus Quate & Fairchild, 1961, Australophlebotomus Theodor, 1948 and Chinius Leng, 1987. Moreover, these authors considered the taxa Parvidens Theodor & Mesghali, 1964 as incertae sedis. The transmission of Leishmania parasites in the Old World is mainly ensured by species belonging to the genus Phlebotomus (Killick-Kendrick 1990). However, it was recently found that species from other genera could also play a role in leishmaniases transmission in the Old World. It is the case of the genus Sergentomyia (Sergentomyia darlingi) involved in the transmission of Leishmania major in Mali (Berdjane-Brouk et al. 2012). The genus Phlebotomus is currently subdivided in nine subgenera according Rispail and Léger (1998b): (Phlebotomus) Rondani, 1843, (Paraphlebotomus) Theodor, 1948, (Synphlebotomus) Theodor, 1948, (Larroussius) Theodor, 1948, (Adlerius) Nitzulescu, 1931, (Transphlebotomus) Artemiev, 1984, (Euphlebotomus) Theodor, 1948, (Anaphlebotomus) Theodor, 1948, and (Kasaulius) Lewis, 1982. However, Rispail and Léger (1998a) thought the subgenus (Anaphlebotomus) is artificial, because it includes two lineages. New species from Madagascar belonging to this group were recently described (Depaquit et al. 2002, 2004, Randrianambinintsoa et al. 2013), suggesting this subgenus needs to be revised. Until it, we think that the subgenus (Abonnencius) Ontiveros et al. (1982), considered as a non-valid taxa (Lane & Alexander 1988), has to be retained.
The main species involved in the Leishmania transmission are: Phlebotomus (Phlebotomus) papatasi (Scopoli, 1786), Phlebotomus (Paraphlebotomus) sergenti Parrot, 1917, Phlebotomus (Euphlebotomus) argentipes Annandale & Brunetti, 1908, as well as many species of the subgenus Larroussius for the transmission of L. major, Leishmania tropica, Leishmania donovani and Leishmania infantum, respectively. The creation of all these subgenera was gradually made initially without phylogenetic arguments, but has become customary among specialists. P. multihamatus sp. nov. possesses antero-inferior mesanepisternal setae. Therefore its inclusion in the genus Phlebotomus is obvious. In addition, its geographical localisation is also in adequacy with this systematic position. However, in sight of the morphology of its genitalia, this species cannot be included in any of the existing subgenus of Phlebotomus. The presence of three spines on the style is a criteria shared with genus Australophlebotomus confined to the Australian region. However, in P. multihamatus sp. nov., the paramere is complex and the genital filaments are long whereas the paramere is simple and the genital filaments are short and even nonexistent in Australophlebotomus (Lewis & Dyce 1982, Léger & Pesson 1993).
We cannot include it either in the subgenera with simple paramere such as (Larroussius), (Adlerius), (Transphlebotomus), (Paraphlebotomus), (Synphlebotomus) and (Kasaulius) The absence of a basal lobe on the coxite prevents its inclusion in the subgenus (Phlebotomus). Possibly, it could be related to the subgenus Euphlebotomus, Anaphlebotomus and Abonnencius, the validity of the latter being obvious to us despite the curious position without real scientific foundation adopted by Lane and Alexander (1988).
The systematics of these groups deserves a revision based on phylogenetic arguments. Meanwhile, we note down the existence of points of convergence in the structure of the male genitalia of these three subgenera with that of P. multihamatus sp. nov.: genital pump well developed and strongly pigmented, complex parameres and short styles. We also note a point of convergence between the hook implanted at the base of the paramere in P. multihamatus sp. nov. and in Phlebotomus fortunatarum.
Moreover, we also observe a certain degree of homology between these structures and the accessory sticks observed in some (Anaphlebotomus) and (Euphlebotomus) like their respective type-species: Phlebotomus stantoni and P. argentipes (Fig. 5). The hypothetical establishment of primary homologies between these two structures will certainly act upon an attempt of cladistics morphological analysis in these groups. On the other hand, the membrane forming a pouch on the paramere seems very original to us among sandflies.
Fig. 5. parameres including organs and accessory sticks from penis of Phlebotomus multihamatus sp. nov. (A), Phlebotomus argentipes (B), Phlebotomus stantoni (C) and Phlebotomus fortunatarum (D).
On the basis of this argumentation, we propose the creation of a new subgenus: (Legeromyia) subg. nov. defined as follows: style with three spines, paramere wearing a basal hook as well as a basal pouch, absence of basal lobe on the coxite. Type-species of the subgenus (Legeromyia) subg. nov: P. multihamatus sp. nov.
The discovery of this new species of sandfly must encourage prospecting in this direction in Gabon. Even if only one case of leishmaniasis has been reported in this country (Tournier 1920), the role of sandflies in the transmission of such parasites or other infectious agents, such as virus or bacteria (Depaquit et al. 2010) to wildlife and domestic animals or to human and their trophic preferences, should be evaluated.
ACKNOWLEDGEMENTS
To Donald Ndongo and Patrice Mikelet, for their technical help during the field work, to the Station d'Etude des Gorilles et Chimpanzés, for the logistic support, to the Agence Nationale des Parcs Naturels and the Centre National de la Recherche Scientifique et Technologique of Gabon, that authorised this study and facilitated the access to the National Park of La Lopé.
REFERENCES
- Abonnenc E. Les phlébotomes de la Région Ethiopienne (Diptera, Psychodidae) Memóires ORSTOM 55; Paris: 1972. 289 (Fre). [Google Scholar]
- Alvar J, Vélez ID, Bern C, Herrero M, Desjeux P, Cano J, Jannin J, den Boer M. WHO Leishmaniasis Control Team 2012. Leishmaniasis worldwide and global estimates of its incidence. PLoS ONE. 7: doi: 10.1371/journal.pone.0035671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berdjane-Brouk Z, Koné AK, Djimdé AA, Charrel RN, Ravel C, Delaunay P, del Giudice P, Diarra AZ, Doumbo S, Goita S, Thera MA, Depaquit J, Marty P, Doumbo OK, Izri A. First detection of Leishmania major DNA in Sergentomyia (Spelaeomyia) darlingi from cutaneous leishmaniasis foci in Mali. PLoS ONE. 2012;7: doi: 10.1371/journal.pone.0028266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Depaquit J, Grandadam M, Fouque F, Andry PE, Peyrefitte C. Arthropod-borne viruses transmitted by phlebotomine sand flies in Europe: a review. Euro Surveill. 2010;15:19507–19507. [PubMed] [Google Scholar]
- Depaquit J, Léger N, Robert V. Première mention de Phlebotomus à Madagascar (Diptera: Psychodidae). Description de Phlebotomus (Anaphlebotomus) fertei n. sp. et de Phlebotomus (Anaphlebotomus) huberti n. sp. Parasite. 2002;9:325–331. doi: 10.1051/parasite/2002094325. (Fre). [DOI] [PubMed] [Google Scholar]
- Depaquit J, Léger N, Robert V. Les phlébotomes de Madagascar. III - Description de Phlebotomus (Anaphlebotomus) fontenillei n. sp. Parasite. 2004;11:261–265. doi: 10.1051/parasite/2004113261. (Fre). [DOI] [PubMed] [Google Scholar]
- Galliard H, Nitzulescu V. Contribution à l'étude des phlébotomes du Gabon. Phlebotomus sanneri n. sp. Ann Parasitol Hum Comp. 1931;9:233–246. (Fre). [Google Scholar]
- ICZN - International Commission on Zoological Nomenclature 2000 . International Code of Zoological Nomenclature. 4th ed. The International Trust for Zoological Nomenclature 1999/The Natural History Museum; London: 124 [Google Scholar]
- Killick-Kendrick R. Phlebotomine vectors of the leishmaniases: a review. Med Vet Entomol. 1990;4:1–24. doi: 10.1111/j.1365-2915.1990.tb00255.x. [DOI] [PubMed] [Google Scholar]
- Lane RP, Alexander B. Sand flies (Diptera: Phlebotominae) of the Canary Islands. J Nat Hist. 1988;22:313–319. [Google Scholar]
- Léger N, Pesson B. Première mention de phlébotomes en Nouvelle-Calédonie et description de deux espèces nouvelles (Diptera, Psychodidae, Phlebotominae) Mémoires du Muséum National d'Histoire Naturelle. 1993;157:123–129. (Fre). [Google Scholar]
- Leng YJ, Lewis DJ. The subgenus Euphlebotomus (Diptera: Psychodidae) in China, with description of a new species, Phlebotomus yunshengensis. Ann Trop Med Parasitol. 1987;81:305–309. doi: 10.1080/00034983.1987.11812124. [DOI] [PubMed] [Google Scholar]
- Lewis D, Dyce A. The subgenus Australophlebotomus Theodor of Phlebotomus Rondani and Berté (Diptera: Psychodidae) Aust J Entomol Soc. 1982;21:37–54. [Google Scholar]
- Niang AA, Geoffroy B, Davidson I, Depaquit J, Hervy J-P, Boussès P, Leger N, Trouillet J, Killick-Kendrick R, Killick-Kendrick M, Rioux J-A, Brunhes J. Sand flies of the Afrotropical Region: identification and training program. Collection Didactiques, IRD Editions; Paris: 2004. [Google Scholar]
- Ontiveros JMU, Marquez FM, Benitez DCG, Roman RL, Barrios CC. Flebotomos de las Islas Canarias (España) Rev Iber Parasitol. 1982;42:197–206. [Google Scholar]
- Randrianambinintsoa FJ, Léger N, Robert V, Depaquit J. Phlebotomine sand flies from Madagascar (Diptera: Psychodidae). VII. An identification key for Phlebotomus with the description of Phlebotomus (Anaphlebotomus) vaomalalae n. sp. Parasite. 2013;20:6–6. doi: 10.1051/parasite/2013005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rispail P, Léger N. Numerical taxonomy of Old World Phleboto-minae (Diptera: Psychodidae). 1. Considerations of morphological characters in the genus Phlebotomus Rondani & Berté 1840. Mem Inst Oswaldo Cruz. 1998;93:773–785. doi: 10.1590/s0074-02761998000600015. [DOI] [PubMed] [Google Scholar]
- Rispail P, Léger N. Numerical taxonomy of Old World Phlebotominae (Diptera: Psychodidae). 2. Restatement of classification upon subgeneric morphological characters. Mem Inst Oswaldo Cruz. 1998;93:787–793. doi: 10.1590/s0074-02761998000600016. [DOI] [PubMed] [Google Scholar]
- Theodor O. Classification of the Old World species of the subfamily Phlebotominae (Diptera: Psychodidae) Bull Entomol Res. 1948;39:85–118. doi: 10.1017/s0007485300024305. [DOI] [PubMed] [Google Scholar]
- Tournier E. Note sur un cas de Kala-azar infantile observé au Gabon. Bull Soc Pathol Exot. 1920;13:175–176. (Fre). [Google Scholar]