

Abstract
Behavioural responses can help species persist in habitats modified by humans. Roads and traffic greatly affect animals' mortality not only through habitat structure modifications but also through direct mortality owing to collisions. Although species are known to differ in their sensitivity to the risk of collision, whether individuals can change their behaviour in response to this is still unknown. Here, we tested whether common European birds changed their flight initiation distances (FIDs) in response to vehicles according to road speed limit (a known factor affecting killing rates on roads) and vehicle speed. We found that FID increased with speed limit, although vehicle speed had no effect. This suggests that birds adjust their flight distance to speed limit, which may reduce collision risks and decrease mortality maximizing the time allocated to foraging behaviours. Mobility and territory size are likely to affect an individuals' ability to respond adaptively to local speed limits.
Keywords: flight initiation distance, phylogenetic mixed model, road kill
1. Introduction
Human development can affect animal population persistence. Species differentially respond to these changes; some avoid anthropogenic habitats while others benefit from exploiting them. For example, specialist species tend to disappear in urban areas, whereas some generalists (human-adapted species) increase in such areas [1]. Some species are known to change their behaviour in adaptive ways in response to human-induced habitat changes. For instance, in urban areas, songbirds sing at higher frequencies as a response to noise pollution [2] or change their singing activity to avoid noisy periods [3]. Similarly, within species, flight distance from humans decreases in urban areas as compared with rural areas [4]. Responses to stressors such as predation or human disturbance can be costly. Individuals must trade the benefits acquired during current activity against the costs of escaping from a potential threat. The exposure to frequently occurring stressors, for example humans in urban areas, may thus select for a reduction in sensitivity to predator exposure, decreasing the flight distance of urban birds as a response to predation risk [4,5].
Roads and traffic are a substantial cause of biodiversity loss [6] that affect animals by causing habitat loss, isolation of populations, creating barriers and increasing mortality [7–10]. Several field and comparative studies have investigated factors explaining interspecific variation in the probability of being killed on a road. Species and habitat characteristics are main factors affecting this probability that also increases with bird mobility [11]. Road speed limits or surrounding landscape features can also impact the probability of being killed [12,13]. We may also expect individuals living near roads to be habituated to traffic, exhibiting potentially adaptive behaviours, for example fear responses, to limit mortality via roadways.
Bird species have highly heterogeneous fear responses. Flight initiation distance (FID), the distance at which prey flees from an approaching predator, is affected by individual state and species characteristics [14] but also by external factors like availability of resources, microhabitat characteristics, distance to a refuge and predator behaviour [15,16]. Anti-predator behaviour can allow species to respond adaptively to different predators or sources of disturbance.
As road traffic directly causes mortality in birds, we expect birds to respond to road traffic in a similar way as they would respond to predation. As road mortality probably increases with road speed limits [13], we would expect individuals to adjust their anti-predator behaviour to vehicle speed and/or to the speed limit. Species with longer FID are known to have a smaller risk of getting killed by cars [10], suggesting that adjusting FID might be an adaptive way to respond to road traffic. Measuring the FID of common European birds from cars, we derive two non-mutually exclusive hypotheses to explain FID in response to traffic. Individuals may directly assess risk and adjust their FID for each given vehicle speed. Alternatively, road speed limit alone could determine FID, suggesting that road speed limit is considered as a habitat characteristic.
2. Material and methods
(a). Study site
The study took place in western France mostly on our way home from the laboratory (CEBC-CNRS; 46°10′ N; 0°28′ W; from November 2006 to November 2007). Croplands, forests and small villages dominate the landscape. We assessed FID on road sections with speed limits of 20, 50, 90 and 110 km h−1. In order to disentangle the effects of speed limit and car speed, three speed regimes were randomly used: the car (a compact car: a white Peugeot 205) was driven either at the speed limit, over, or under the speed limit.
(b). Flight initiation distance
Birds standing on the road or on road edges were sighted while driving and the timer started when the bird took off. We recorded the elapsed time (tenths of seconds precision) to cover the distance to the flight initiation location at a constant speed. The FID was calculated (in meters) as the product of vehicle speed by elapsed time. Other variables such as bird position (on the road or on the approx. 4 m edges), bird species and road speed limit were also collected. We divided the year into two distinct seasons (autumn and winter; spring and summer) to account for different cohorts (with more naive juveniles in autumn than in spring). Different road sections were used (see the electronic supplementary material, table S1). The same observer (P.L.) recorded all observations.
(c). Statistical analyses
FID was modelled by fitting generalized linear mixed models with Markov chain Monte Carlo (MCMC) techniques using the R package MCMCglmm [17]. Speed limit, car speed, season, bird position and bird mass were included as fixed effects. For car speed, we used both the absolute car speed and the difference between car speed and road speed limit. To account for phylogeny, we used trees provided by http://birdtree.org/. Details of the procedure are given in the electronic supplementary material. To test whether the phylogeny affected the FID, we compared the average deviance information criterion (DIC) of phylogenetic models with the DIC of a model with the same fixed effects, but including species as a random effect instead of phylogeny. Best models were then selected using backward selection, eliminating non-significant fixed effects step by step.
3. Results
A total of 134 FID was measured for 21 different bird species (see the electronic supplementary material, table S1). As models with phylogeny had a higher DIC (ΔDIC = 3.3), we include only the results of the model with species as a random effect but without phylogeny. Note however that both types of models yielded similar results (see the electronic supplementary material, table S2 for models with phylogeny). Birds had significantly higher FID on road sections with higher speed limits (table 1 and figure 1). By contrast, car speed had no significant effect on FID, both when considering the absolute car speed (posterior mean (pm) = 0.007; CI = (−0.003; 0.019); pMCMC = 0.219) or the difference between car speed and speed limit (pm = 0.006; CI = (−0.006; 0.017); pMCMC = 0.271; figure 2). The interaction between car speed and speed limit was not significant (pm = 0.001; CI = (−0.001; 0.001); pMCMC = 0.070). Bird species with higher body mass initiated flight at a greater distance (table 1), and FID was longer for birds on the road as compared with those on road edges (table 1). Finally, season significantly affected FID, and FID was longer in autumn and winter (table 1). Meteorological variables had no effect on FID (p = 0.742).
Table 1.
Best model explaining bird FID according to speed limit and covariates (see text for details). (Birds standing on the road or on the edge was defined as location. Season is a binomial variable, with spring–summer used as reference season. CI, confidence interval; pm, posterior mean. Species was included as a random effect.)
| variable | pm | CI | pMCMC |
|---|---|---|---|
| intercept | 0.454 | (−0.678; 1.398) | 0.324 |
| location | 0.790 | (0.440; 1.174) | <0.001 |
| speed limit | 0.016 | (0.007; 0.024) | <0.001 |
| body mass | 0.002 | (0.000; 0.003) | 0.030 |
| season | 0.591 | (0.229; 0.999) | 0.004 |
Figure 1.
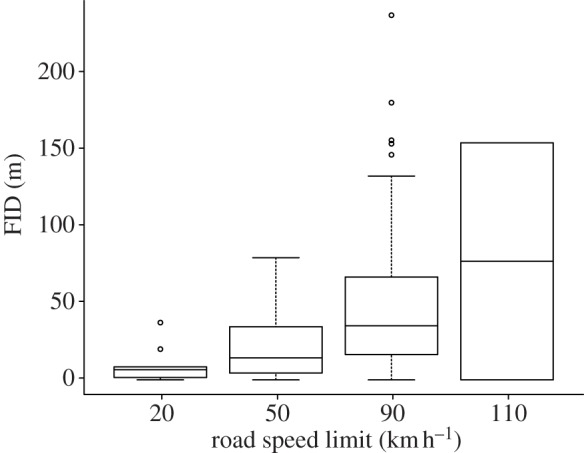
Boxplot of FID in common European birds for different road speed limits. Medians, first and third quartile, and outliers are presented for each speed limit. Whiskers represent 5 and 95 percentiles.
Figure 2.
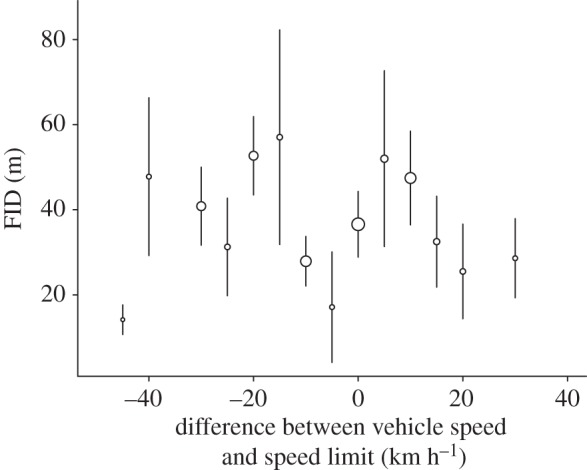
Illustration of the absence of relationship between FID and difference between vehicle speed and speed limit in common European birds. Mean FID ± s.e. is represented. Dot size is proportional to the number of observations.
As three species accounted for more than half the data (Corvus corone: n = 27; Passer domesticus: n = 21; Turdus merula: n = 27), the models were run again considering only these three species. The results were very similar, FID increasing with speed limit (pMCMC < 0.001) without any effect of car speed (pMCMC = 0.151). Similarly, when excluding these three species, the results remained similar; FID increasing with speed limit (pMCMC = 0.024) but not with car speed (p = 0.798).
4. Discussion
Our results show that birds change their FID according to speed limit rather than car speed. This strongly suggests that birds are able to associate road sections with speed limits as a way to assess collision risk. The results are consistent when testing for potential confounding variables such as season, phylogeny, bird mass and location of the bird on or off the road. The absence of car speed effect on FID is consistent with a study in galahs Cacatua roseicapilla [18], where variation in approach speed (from human walk) to these Australian parrots did not affect their FID. Consistent with several other studies, FID increased with body mass [19]. Smaller species are more agile and thus more efficient at initiating flight, allowing them to have shorter FID [19]. FID was also longer in spring and summer than in autumn and winter, which may be explained by the presence of naive juveniles in spring and summer. Alternatively, adults may take more risks during the reproductive season, increasing the amount of time spent foraging to feed their offspring. Birds standing on the road had greater FID than birds standing on road edges, highlighting predation risk perception by birds. Finally, inclusion of phylogeny or not yielded similar relationships between FID and speed limit, suggesting that our results could not be attributed to phylogenetic similarities. The fact that including mass, species and phylogeny in the models did not affect the results provides additional support for the robustness of our conclusions. In addition, although our data were unbalanced, the fact that we obtained similar results when considering only species for which we had more records with data homogeneously distributed among different speed limits also strengthens our results' robustness. As road sections with speed limits of 50 km h−1 are within inhabited villages, the differences in FID between these sections and sections with higher speed limits could simply be explained by differences in human presence and landscape structure (rural versus urban areas [4,6]). However, FID for road sections with limits of 110 km h−1 was also higher than at 90 km h−1 (figure 1), even though these two speed limits had similar surrounding landscapes. This probably indicates that speed limit is a real factor determining FID. Our results thus probably reflect an optimal strategy to maximize the time allocated to foraging while minimizing collision risk.
Several different mechanisms are expected to explain variation in FID. First, individuals with FID adapted to local conditions could have increased in frequency through natural selection. The effects of natural selection on bird FID were investigated by Møller [5], who showed that urban birds were selected for shorter FID. However, natural selection probably does not drive FID variation in this study, because similar patterns were found whatever the species considered (whether urban, rural, migratory or sedentary; results not shown), and also because the same bird can easily be exposed to different road sections with different speed limits.
Alternatively, mechanisms of risk allocation and habituation may also explain variation in FID [12]. According to the risk allocation hypothesis [20], animals should decrease their allocation of anti-predator effort with increasingly frequent high-risk situations. This latter hypothesis is not likely to be involved here because car speed itself had no effect on FID (no direct risk assessment). Habituation and associative learning mechanisms could also explain our results; individuals may associate road sections with different collision risks according to average vehicle speed (i.e. speed limit). More mobile species and/or species inhabiting larger territories, which probably have a less precise knowledge of their territory, might then be less efficient in adapting their FID to speed limits. More mobile bird species have a higher probability of being killed by cars [11], which could indeed be explained by their lower ability to habituate to local speed limits because of a lesser knowledge of their territory. Traffic density could also strongly affect individuals' ability to adjust their behaviour to vehicle speed, as individuals would have a better knowledge of vehicle speed on high-density road sections. In addition, parameters such as traffic density and traffic noise are likely to be associated with speed limit, so that the detected effect of speed limit might in fact be owing to combined effects of average car speed, traffic density and noise intensity. A better understanding of the mechanisms responsible for our results would require either experiments in more controlled conditions [21] or the consideration of a large number of species with known territory size and mobility. Response to humans and associated mortality risk is considered a major issue for bird conservation. FIDs from car traffic could easily become a standard metric to assess how bird species assess their risky environment and potentially enhance bird conservation facing important human development.
Acknowledgements
We thank Daniel Blumstein, Lisa Jacquin, Kristen Peck, Clément Chevallier, Marine Danek-Gontard and two anonymous referees for helpful comments on the manuscript. Aurélie Mélot greatly helped during fieldwork.
Funding statement
S.D. was supported by a Fyssen fellowship.
References
- 1.Devictor V, Julliard R, Couvet D, Lee A, Jiguet F. 2007. Functional homogenization effect of urbanization on bird communities. Conserv. Biol. 21, 741–751 (doi:10.1111/j.1523-1739.2007.00671.x) [DOI] [PubMed] [Google Scholar]
- 2.Brumm H, Zollinger SA. 2011. The evolution of the Lombard effect: 100 years of psychoacoustic research. Behaviour 148, 1173–1198 (doi:10.1163/000579511x605759) [Google Scholar]
- 3.Fuller RA, Warren PH, Gaston KJ. 2007. Daytime noise predicts nocturnal singing in urban robins. Biol. Lett. 3, 368–370 (doi:10.1098/rsbl.2007.0134) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Møller AP. 2008. Flight distance of urban birds, predation, and selection for urban life. Behav. Ecol. Soc. 63, 63–75 (doi:10.1007/s00265-008-0636-y) [Google Scholar]
- 5.Møller AP. 2010. Interspecific variation in fear responses predicts urbanization in birds. Behav. Ecol. 21, 365–371 (doi:10.1093/beheco/arp199) [Google Scholar]
- 6.Benítez-López A, Alkemade R, Verweij PA. 2010. The impacts of roads and other infrastructure on mammal and bird populations: a meta-analysis. Biol. Cons. 143, 1307–1316 (doi:10.1016/j.biocon.2010.02.009) [Google Scholar]
- 7.Fahrig L, Rytwinski T. 2009. Effects of roads on animal abundance: an empirical review and synthesis. Ecol. Soc. 14, 21 [Google Scholar]
- 8.Forman RT, Alexander LE. 1998. Roads and their major ecological effects. Annu Rev. Ecol. Syst. 29, 207–231 (doi:10.1146/annurev.ecolsys.29.1.207) [Google Scholar]
- 9.Spellerberg I. 1998. Ecological effects of roads and traffic: a literature review. Glob. Ecol. Biogeogr. 7, 317–333 (doi:10.1046/j.1466-822x.1998.00308.x) [Google Scholar]
- 10.Møller AP, Erritzøe H, Erritzøe J. 2011. A behavioural ecology approach to traffic accidents: interspecific variation in causes of traffic casualties among birds. Zool. Res. 32, 115–127 [DOI] [PubMed] [Google Scholar]
- 11.Rytwinski T, Fahrig L. 2012. Do species life history traits explain population responses to roads? A meta-analysis. Biol. Conserv. 147, 87 (doi:10.1016/j.biocon.2011.11.023) [Google Scholar]
- 12.Rodriguez-Prieto I, Fernández-Juricic E, Martín J, Regis Y. 2009. Antipredator behaviour in blackbirds: habituation complements risk allocation. Behav. Ecol. 20, 371–377 (doi:10.1093/beheco/arn151) [Google Scholar]
- 13.Farmer RG, Brooks RJ. 2012. Integrated risk factors for vertebrate roadkill in southern Ontario. J. Wildl. Manag. 76, 1215–1224 (doi:10.1002/jwmg.358) [Google Scholar]
- 14.Carrete M, Tella JL. 2010. Individual consistency in flight initiation distances in burrowing owls: a new hypothesis on disturbance-induced habitat selection. Biol. Lett. 6, 167–170 (doi:10.1098/rsbl.2009.0739) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Lima SL, Dill LM. 1990. Behavioural decisions made under the risk of predation: a review and prospectus. Can. J. Zool. 68, 619–640 (doi:10.1139/z90-092) [Google Scholar]
- 16.Stankowich T, Blumstein DT. 2005. Fear in animals: a meta-analysis and review of risk assessment. Proc. R. Soc. B 272, 2627–2634 (doi:10.1098/rspb.2005.3251) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Hadfield JD. 2010. MCMC methods for multi-response generalized linear mixed models: the MCMCglmm R package. J. Stat. Softw. 33, 1–2220808728 [Google Scholar]
- 18.Cárdenas YL, Shen B, Zung L, Blumstein DT. 2005. Evaluating temporal and spatial margins of safety in galahs. Anim. Behav. 70, 1395–1399 (doi:10.1016/j.anbehav.2005.03.022) [Google Scholar]
- 19.Blumstein DT. 2006. Developing an evolutionary ecology of fear: how life history and natural history traits affect disturbance tolerance in birds. Anim. Behav. 71, 389–399 (doi:10.1016/j.anbehav.2005.05.010) [Google Scholar]
- 20.Lima SL, Bednekoff PA. 1999. Temporal variation in danger drives antipredator behaviour: the predation risk allocation hypothesis. Am. Nat. 153, 649–659 (doi:10.1086/303202) [DOI] [PubMed] [Google Scholar]
- 21.Zimmer C, Boos M, Poulin N, Gosler A, Petit O, Robin J-P. 2011. Evidence of the trade-off between starvation and predation risks in ducks. PLoS ONE 6, e22352 (doi:10.1371/journal.pone.0022352) [DOI] [PMC free article] [PubMed] [Google Scholar]