

Abstract
In mammals, molecular clocks regulate transcription and glucose homeostasis. One way they do so is by controlling glucocorticoid-receptor signalling, which suggests that clocks are embedded in liver metabolism.
The idea that genes program behaviour dates back to the 1970s, when researchers discovered that heritable mutations in fruitflies alter the daily, or circadian, sleep–wake cycle. At a molecular level, the clock machinery, which is composed of an autoregulatory transcription feedback loop1, is present in almost all cell types and generates oscillations in the expression of at least 10% of the genes transcribed2,3. Such observations led to an intensive search for the clock’s physiological functions. Lamia et al.4 describe one such function on page 552 of this issue. They report that the clock protein cryptochrome acts as an on/off switch for the nuclear receptor of glucocorticoids — hormones that are involved in glucose and lipid metabolism5. The finding expands our understanding of how metabolism is coordinated with geophysical time.
The impact of circadian clocks on organismal energetics and survival has been investigated in photosynthetic organisms. In cyanobacteria6 and plants7, alignment of the period length with the environmental light cycle (a defining property of the clock) influences growth and reproduction. This suggests that ‘resonance’ of internal oscillators with the environment may provide a selective advantage during evolution. In these organisms, the clock machinery also separates oxygen- and nitrogen-fixation processes, averting accumulation of cytotoxic free radicals and preventing futile energetic cycles8.
In mammals, circadian rhythms operate at the behavioural, physiological and molecular levels. But studying the link between circadian cycles and metabolism in organisms with a nervous system is complicated. This is because the light response originates from the brain’s pacemaker neurons, whereas nutrient sensing occurs in other neurons and throughout many peripheral tissues. The discovery that fibroblasts maintained in culture exhibit self-sustaining circadian oscillations that show signature properties of a circadian clock9 was crucial because it hinted that peripheral clocks can function autonomously. Normally, however, brain and peripheral timepieces seem to communicate to give rise to coherent metabolic outputs. Indeed, genetic studies suggest that the brain and peripheral clocks participate in weight regulation and in glucose and lipid metabolism3.
Circadian oscillations in hormone levels are certainly crucial for metabolic health, and disturbances in these have been used in clinical diagnosis for decades. For instance, high blood levels of the glucocorticoid hormone cortisol at midnight are regarded as a sign of Cushing’s disease. Similarly, a ‘dawn phenomenon’ of elevated blood glucose levels arises in humans with diabetes owing to increased production of glucocorticoids in the early morning.
At a molecular level, expression of the nuclear receptors for glucocorticoids shows strong circadian oscillations10. In fact, certain nuclear hormone receptors are integral components of the clock machinery11. These receptors’ activity results in a complex pattern of gene activation and repression, affecting metabolic and haematopoietic tissues in various ways. The clock protein PER2 interacts with several nuclear hormone receptors12, but the physiological impact of the clock machinery on the receptors’ function has not been established.
Lamia and colleagues previously showed13 that the clock-repressor protein cryptochrome (Cry) is a sensor of cellular energy state. To further dissect the coupling between Cry and metabolic signals, some of the same authors now survey interactions of Cry with a broad range of nuclear hormone receptors in mice4.
The authors report a strong and specific interaction between Cry and glucocorticoid receptors that shifts the balance towards gene repression after activation of these receptors. In addition, Cry interacted with two nuclear hormone receptors that activate transcription of the clock-activator protein Bmal1. This suggests that Cry may contribute to the oscillator’s robustness by participating in a short feedback loop controlling Bmal1 expression. Intriguingly, Cry interaction with glucocorticoid receptors was enhanced by dexamethasone4 — a drug that acts as a glucocorticoid. Because circadian levels of Cry vary, this observation directly links glucocorticoid signalling to circadian-clock oscillations (Fig. 1). Lamia and collaborators also note that Cry modulates patterns of gene activation and repression stimulated by glucocorticoid receptors. In Cry-deficient cells, glucocorticoid signalling led to a higher number of activated targets, and a vast decrease in the number of genes normally repressed by this clock protein.
Figure 1. Biological clocks organize glucose metabolism.
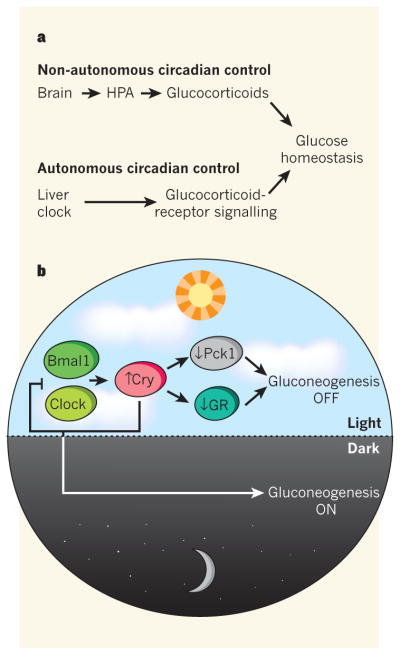
a, Glucose homeostasis is regulated by the interplay between non-autonomous circadian control — which affects glucocorticoid hormone levels through the brain and the hypothalamic–pituitary–adrenal (HPA) axis — and autonomous oscillatory regulation of these hormones’ signalling in peripheral tissues such as the liver. Lamia et al.4 report that the clock-repressor protein Cry plays a central part at both of these levels. b, In the liver, the clock-activator proteins Clock and Bmal1 increase Cry transcription, which downregulates gluconeogenesis both by interacting with — and so inhibiting — glucocorticoid receptors (GRs), and by reducing the expression of the Pck1 enzyme. As is typical of the clock machinery, high levels of Cry also downregulate Clock and Bmal1 transcription in a negative feedback loop, thus reducing its own levels and so promoting gluconeogenesis during the dark phase of the light–dark cycle.
Because the main outcome of glucocorticoid-receptor activity is glucose generation in the liver by the process of gluconeogenesis, and because Cry-deficient animals display impaired glucose tolerance14, Lamia et al.4 focused on Cry function in the liver. The authors found that Cry directly represses the expression of Pck1 — a gene that encodes the rate-limiting enzyme in gluconeogenesis (Fig. 1). In Cry-deficient cells, dexamethasone increased Pck1 expression, but did not affect repression of genes mediating the inflammatory NF-κB signalling pathway. This observation indicates a separation of Cry functions in the gluconeogenic and inflammatory pathways.
Cry deficiency also increased the sensitivity of mice to glucocorticoid-induced diabetes4. This finding has therapeutic implications given the widespread use of steroids — of which glucocorticoids are an example — for the management of inflammation associated with cancer and conditions related to the immune system. Increasing Cry levels may enhance the anti-inflammatory effects of glucocorticoids by blocking the adverse effects of these hormones on glucose metabolism.
Whereas previous work14 implicated Cry in regulating the signalling of glucagon (another hormone that mediates glucose homeostasis) in the cytoplasm, this paper pinpoints another role for Cry in the control of gluconeogenesis at the nuclear level. The modulation of glucocorticoid signalling by Cry to control glucose homeostasis may be relevant for understanding glucose overproduction by the liver in human diabetes, not least because a genomic-association study15 indicated that CRY2 gene variants are associated with glucose homeostasis in humans.
At the organismal level, glucocorticoid regulation in animals is controlled through a classical neuroendocrine feedback loop involving communication between the hypothalamus, pituitary and adrenal tissues (Fig. 1). Lamia et al. find that, in Cry-deficient mice, blood glucocorticoid levels remain high at all times of the day–night cycle, with the hormone’s overproduction arising at the level of the hypothalamus. How Cry deficiency might impair feedback inhibition of steroidogenesis in the hypothalamus is a fascinating question.
Lamia and colleagues’ findings may also be relevant to the spike seen in metabolic disorders among individuals subjected to shift work, sleep curtailment and other forms of circadian disruption. A feature of these disorders may be misalignment of oscillations in blood levels of glucocorticoids with oscillations in both the interaction between Cry and glucocorticoid receptors, and their signalling in the liver. This study provides an additional example of the extensive molecular integration and relatedness of temporal behavioural programmes with cell metabolism.
References
- 1.Rosbash M, et al. Cold Spring Harb Symp Quant Biol. 2007;72:75–83. doi: 10.1101/sqb.2007.72.062. [DOI] [PubMed] [Google Scholar]
- 2.Panda S, et al. Cell. 2002;109:307–320. doi: 10.1016/s0092-8674(02)00722-5. [DOI] [PubMed] [Google Scholar]
- 3.Ramsey KM, Bass J. Cold Spring Harb Symp Quant Biol. 2011 http://dx.doi.org/10.1101/sqb.2011.76.010546.
- 4.Lamia KA, et al. Nature. 2011;480:552–556. doi: 10.1038/nature10700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Hollenberg SM, et al. Nature. 1985;318:635–641. doi: 10.1038/318635a0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ouyang Y, Andersson CR, Kondo T, Golden SS, Johnson CH. Proc Natl Acad Sci USA. 1998;95:8660–8664. doi: 10.1073/pnas.95.15.8660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Dodd AN, et al. Science. 2005;309:630–633. doi: 10.1126/science.1115581. [DOI] [PubMed] [Google Scholar]
- 8.Rutter J, Reick M, McKnight SL. Annu Rev Biochem. 2002;71:307–331. doi: 10.1146/annurev.biochem.71.090501.142857. [DOI] [PubMed] [Google Scholar]
- 9.Balsalobre A, Damiola F, Schibler U. Cell. 1998;93:929–937. doi: 10.1016/s0092-8674(00)81199-x. [DOI] [PubMed] [Google Scholar]
- 10.Yang X, et al. Cell. 2006;126:801–810. doi: 10.1016/j.cell.2006.06.050. [DOI] [PubMed] [Google Scholar]
- 11.Alenghat T, et al. Nature. 2008;456:997–1000. doi: 10.1038/nature07541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Schmutz I, Ripperger JA, Baeriswyl-Aebischer S, Albrecht U. Genes Dev. 2010;24:345–357. doi: 10.1101/gad.564110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Lamia KA, et al. Science. 2009;326:437–440. doi: 10.1126/science.1172156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Zhang EE, et al. Nature Med. 2010;16:1152–1156. doi: 10.1038/nm.2214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Dupuis J, et al. Nature Genet. 2010;42:105–116. doi: 10.1038/ng.520. [DOI] [PMC free article] [PubMed] [Google Scholar]