

Abstract
Apigenin, a natural flavonoid found in a variety of vegetables and fruits, has been shown to possess many biological functions. The present study was undertaken to investigate the influence of apigenin on vascular smooth muscle contractility and to determine the mechanism involved. Denuded aortic rings from male rats were used and isometric contractions were recorded and combined with molecular experiments. Apigenin significantly relaxed fluoride-, thromboxane A2 mimetic- or phorbol ester-induced vascular contraction, which suggests that apigenin could be an anti-hypertensive that reduces agonist-induced vascular contraction regardless of endothelial nitric oxide synthesis. Furthermore, apigenin significantly inhibited fluoride-induced increases in pMYPT1 levels and phorbol ester-induced increases in pERK1/2 levels, which suggests the mechanism involving the inhibition of Rho-kinase and MEK activity and the subsequent phosphorylation of MYPT1 and ERK1/2. This study provides evidence regarding the mechanism underlying the relaxation effect of apigenin on agonist-induced vascular contraction regardless of endothelial function.
Keywords: Apigenin, ERK1/2, Fluoride, MYPT1, Phorbol ester, Rho-kinase
INTRODUCTION
Apigenin (4′,5,7-trihydroxyflavone, Fig. 1) is a natural flavonoid that is found abundantly in parsley, peppermint, lemon, perilla, berries, and fruits (Peterson and Dwyer, 1998). Apigenin has been reported to have beneficial effects which include anti-inflammatory (Nicholas et al., 2007) and anti-carcinogenic effects for skin, and free radical scavenging properties in many in vitro systems (Han et al., 2012). Studies on anti-carcinogenic effects have shown that apigenin possesses growth inhibitory properties against many human cancer cell lines, for examples, breast, colon (Wang et al., 2000), skin (Caltagirone et al., 2000), thyroid, leukemia cells, and solid malignant tumor cells (Fotsis et al., 1998). Although the influence of endothelial nitric oxide synthesis is well established, we investigated the possible influence and related mechanisms of the anti-inflammatory apigenin on vascular smooth muscle contractility to develop a better antihypertensive. Denuded aortic rings from male Sprague-Dawley rats were used and isometric contractions were recorded using a computerized data acquisition system and combined with molecular experiments.
Fig. 1.
The chemical structure of apigenin (4′,5,7-trihydroxyflavone).
Alterations in the arterial tone are frequently associated with cardiovascular diseases constituting an important cause of morbidity and mortality in humans, one of which is hypertension that is a multifactorial disorder that involves many mechanisms including endothelial dysfunction and leading to risk factors for cardiovascular diseases. Besides endothelial dysfunction, it is generally accepted that vascular smooth muscle contractility is predominantly controlled by Ca2+ signaling involving Ca2+ influx, release or sensitization and regulating a Ca2+-dependent increase in the phosphorylation of a 20 kDa myosin light chain (MLC20) (Somlyo and Somlyo, 1994). The extent of MLC20 phosphorylation or force of contraction induced by agonist stimulation is usually higher than that caused by an increase in the cytosolic Ca2+ concentration referred to as Ca2+ sensitization (Somlyo and Somlyo, 1994). Subsequent studies suggested that the inhibition of MLC phosphatase by Rho-kinase (Kitazawa et al., 1991; Uehata et al., 1997; Somlyo and Somlyo, 1998; Sakurada et al., 2003) or thin filament regulation including the activation of protein kinase C (PKC), mitogen-activated protein kinase kinases (MEK) and extracellular signal regulated kinase (ERK) 1/2, and phosphorylation of the actin binding protein caldesmon (Wier and Morgan, 2003) may be major components of the pathway that facilitates in Ca2+ sensitization.
Activation of ERK1/2 cannot only regulate vascular contractility but also is connected with pathologic hypertrophy, hyperplasia, hypertension and atherosclerosis (Xu et al., 1996; Touyz et al., 1999). ERK1/2 is activated by threonine and tyrosine phosphorylation by the specific kinase MEK activated by Raf. In various smooth muscles, fluoride, phorbol ester or thromboxane A2 mimetic has been shown to induce contractions, which may be due to primarily enhanced Ca2+ sensitivity or partially increased Ca2+ concentration only in thromboxane A2 mimetic. ERK1/2 activation was induced by the phorbol ester, phorbol 12,13-dibutyrate (PDBu). The stimulus PDBu triggers ERK1/2 dependent cytoskeletal remodeling and formation of podosomes inducing ERK1/2 activation (Gu et al., 2007). On the other hand, it is possible that the contractions induced by fluoride or thromboxane A2 mimetic involve the RhoA/Rho-kinase pathway (Jeon et al., 2006). However, it has not been reported as to whether this pathway is inhibited during apigenin-induced vascular smooth muscle relaxation in aortic rings precontracted with Rho-kinase activator fluoride or MEK activator phorbol ester. Therefore, the aim of the present study was to investigate the possible roles of Rho-kinase or MEK inhibition on Ca2+ desensitization during the apigenin-induced relaxation of isolated rat aortas by using RhoA/Rhokinase activators fluoride or thromboxane A2 mimetic or a MEK activator phorbol ester excluding endothelial nitric oxide synthesis.
MATERIALS AND METHODS
Tissue preparation
Male Sprague-Dawley rats weighing 200–300 g were anesthetized with sodium pentobarbital (50 mg/kg i.p.) as subjected to cervical dislocation, in accord with the procedures approved by the Institutional Animal Care and Use Committee at our institutions. Thoracic aortas were quickly removed and immersed in oxygenated (95% O2/5% CO2) physiological saline solution composed of (mM): 115.0 NaCl, 4.7 KCl, 2.5 CaCl2, 1.2 MgCl2, 25.0 NaHCO3, 1.2 KH2PO4, and 10.0 dextrose (pH 7.4). They were then freed of adherent connective tissue, and aortic endothelia were removed by gentle abrasion using a cell scraper.
Contraction measurements
Two stainless-steel triangles were inserted through each vessel ring and each aortic ring was then suspended in a water-jacketed organ bath (10 ml) maintained at 37°C and aerated with a mixture of 95% O2 and 5% CO2. One triangle was anchored to a stationary support, and the other was connected to an isometric force transducer (Grass FT03C, Quincy, Mass., USA). The rings were stretched passively by applying an optimal resting tension of 2.0 g, which was maintained throughout the experiment. Each ring was equilibrated in the organ bath solution for 60 min before contractile responses to 50 mM KCl were measured. Isometric contractions were recorded using a computerized data acquisition system (PowerLab/8SP, AD Instruments, Castle Hill, NSW, Australia).
The direct effect of apigenin was determined by addition of it after KCl (50 mM), thromboxane A2 mimetic (U46619, 0.1 μM), phorbol ester (1 μM) or fluoride (6 mM) induced contractions had plateaued in normal Krebs' solution.
Western blot analysis
Muscle strips were quick-frozen by immersion in a dry ice/acetone slurry containing 10% trichloroacetic acid (TCA) and 10 mM dithiothreitol (DTT). Muscles were stored at −80°C until use. Tissues were brought up to room temperature in a dry ice/acetone/TCA/DTT mixture and then homogenized in a buffer containing 20 mM MOPS, 4% SDS, 10% glycerol, 10 mM DTT, 20 mM β-glycerophosphate, 5.5 μM leupeptin, 5.5 μM pepstatin, 20 kIU aprotinin, 2 mM Na3VO4, 1 mM NaF, 100 μM ZnCl2, 20 μM 4-(2-aminoethyl) benzenesulphonyl fluoride (AEBSF) and 5 mM EGTA. Protein-matched samples (modified Lowry protein assay, DC Protein Assay Kit, Bio-Rad) were electrophoresed on sodium dodecyl sulfate polyacrylamide gel electrophoresis SDS-PAGE (Protogel, National Diagnostics), transferred to polyvinylidene fluoride PVDF membranes, and subjected to immunostaining and densitometry using appropriate antibodies. The success of protein matching was confirmed by Naphthol Blue Black staining of the membrane and by densitometry of the actin band. Lane loading variations were corrected by normalization versus β-actin.
MLC20 Phosphorylation
Muscle strips were quick-frozen by immersion in acetone containing 10% TCA and 10 mM DTT precooled to −80°C. Tissues were brought to room temperature in acetone/TCA/DTT, then ground with glass pestles, and washed with ether to remove TCA. Tissues were extracted in a urea sample buffer as previously described and run on 10% polyacrylamide gels. Proteins were transferred to PVDF membranes and subjected to immunoblot with a specific MLC20 antibody (1:1500, Sigma). Anti-mouse IgG (Goat) conjugated with horseradish peroxidase was used as a secondary antibody (1:2000, Upstate). Bands were detected with enhanced chemiluminescence (ECL) visualized on films and then analyzed by a molecular imager.
Chemicals and antibodies
Drugs and chemicals were obtained from the following sources. Sodium fluoride, KCl, acetylcholine, apigenin, U46619 and phorbol 12,13-dibutyrate were purchased from Sigma (St. Louis, MO, USA). DTT, TCA and acetone were obtained from Fisher Scientific (Hampton, NH, USA). Enhanced chemiluminescence (ECL) kits were from Pierce (Rockford, IL, USA). Antibodies against phospho-myosin phosphatase targeting subunit protein 1 (phospho-MYPT1) at Thr855 (1:5,000), MYPT1, ERK or phosphoERK at Thr202/Tyr204 were purchased from Cell Signaling Technology (Danvers, MA, USA) or Upstate Biotechnology (Lake Placid, NY, USA) to determine levels of RhoA/Rho-kinase activity (Wilson et al., 2005; Wooldridge et al., 2004) or MEK activity. Anti-mouse IgM (goat) and anti-rabbit IgG (goat), conjugated with horseradish peroxidase, were used as secondary antibodies (1:2,000 and 1:2,000, respectively, Upstate, Lake Placid, NY). Apigenin was prepared in dimethyl sulfoxide (DMSO) as a 100 mM stock solution and frozen at −20°C for later use. DMSO alone had no observable effect at concentrations used (data not shown).
Statistics
The data were expressed as mean ± standard error of the mean (SEM). The student’s unpaired t test or ANOVA was used to determine the statistical significance of the means between two groups using SPSS 12.0 (SPSS Inc., Chicago, Illinois, USA). p values <0.05 were regarded as statistically significant.
RESULTS
Effect of apigenin on contractions of endothelium-denuded aortas induced by a full RhoA/Rho-kinase activator fluoride or thromboxane A2 mimetic
Endothelium was removed by gentle abrasion with a cell scraper to identify the direct effect of apigenin on vascular smooth muscle. The absence of endothelium was confirmed by a lack of relaxation after treating precontracted ring segments with acetylcholine (1 μM). Apigenin showed no significant effect on basal tension (data not shown), and significantly inhibited the contraction induced by a Rho-kinase activator fluoride regardless of endothelial nitric oxide synthesis (Fig. 2). This suggests that the relaxation mechanism of apigenin might involve the inhibition of Rho-kinase activity in addition to endothelial nitric oxide synthesis and the subsequent activation of guanylyl cyclase. On the other hand, apigenin at the same concentration significantly inhibited thromboxane A2 mimetic U46619-induced contraction in denuded muscles (Fig. 3) suggesting that thromboxane A2 mimetic acts similarly with fluoride where Rho-kinase activation was the main pathway.
Fig. 2.

Effect of apigenin on fluoride-induced vascular contraction in denuded muscles. Each ring was equilibrated in the organ bath solution for 30–60 min before relaxation responses to apigenin were measured. Data are expressed as the means of 3–5 experime nts with vertical lines representing SEMs. *p<0.05, **p<0.01, presence versus absence of apigenin.
Fig. 3.

Effect of apigenin on thromboxane A2 mimetic-induced vascular contraction in denuded muscles. Each ring was equilibrated in the organ bath solution for 30–60 min before relaxation responses to apigenin were measured. Data are expressed as the means of 3–5 experiments with vertical lines representing SEMs. *p<0.05, **p<0.01, presence versus absence of apigenin.
Effect of apigenin on the contractions of denuded aortas induced by a MEK activator phorbol ester
Phorbol esters used have been proved to be MEK activators and partial RhoA/Rho-kinase activators (data not shown). Interestingly, phorbol 12,13-dibutyrate-induced contraction was significantly inhibited by apigenin at a low concentration regardless of endothelial nitric oxide synthesis (Fig. 4), which suggested that thin or actin filament regulation including MEK/ERK activation were significantly inhibited.
Fig. 4.

Effect of apigenin on phorbol ester-induced vascular contraction in denuded muscles. Each ring was equilibrated in the organ bath solution for 30–60 min before relaxation responses to apigenin were measured. Data are expressed as the means of 3–5 experiments with vertical lines representing SEMs. *p<0.05, **p<0.01, presence versus absence of apigenin.
Effect of apigenin on levels of ERK1/2 phosphorylation at Thr-202/Tyr-204
To confirm the role of apigenin on thin filament regulation of smooth muscle contractility, we measured levels of ERK1/2 and phospho-ERK1/2 in muscles quick frozen after 60 minutes of exposure to apigenin for the equilibration. Each relaxing ring was precontracted with 1 μM phorbol ester (phorbol 12,13-dibutyrate). As compared with vehicle-treated rat aortas, a significant decrease in ERK 1/2 phosphorylation at Thr202/Tyr204 was led by apigenin in these apigenin (0.1 mM)-treated rat aortas in the absence of endothelium (Fig. 5) showing full vasorelaxation (Fig. 4) and thin filament regulation. These findings show that thin or actin filament regulation including ERK1/2 phosphorylation via MEK activation might be of importance in the decreased contractility induced by apigenin.
Fig. 5.
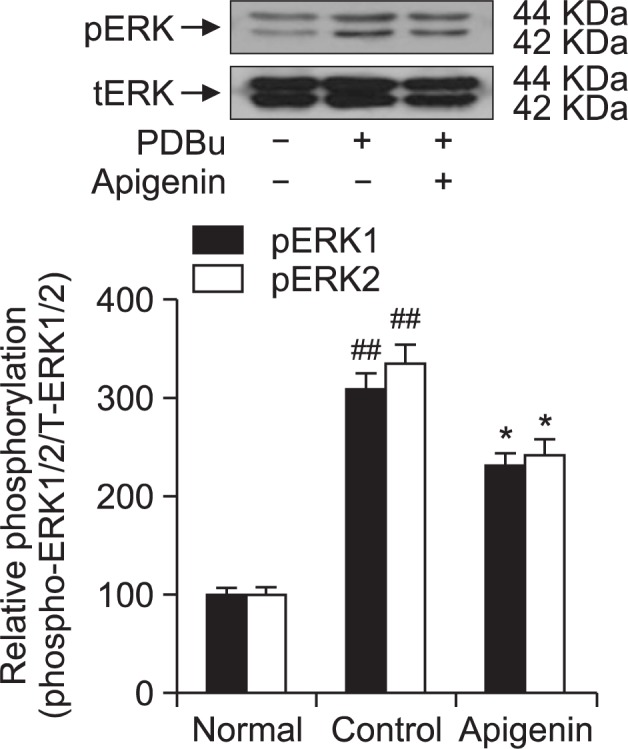
Effect of apigenin on phorbol ester-induced increases in phospho-ERK1/2 levels. Phospho-ERK1/2 protein levels were decreased in quick frozen apigenin-treated rat aortas in the absence of endothelium compared to vehicle-treated rat aortas precontracted with phorbol ester. The upper panel shows a typical blot and the lower panel shows average densitometry results for relative levels of phospho-ERK1/2. Data are expressed as the means of 3–5 experiments with vertical lines representing SEMs. *p<0.05, ##p<0.01, versus control or normal group respectively. Apigenin: 0.1 mM apigenin; PDBu: 1 μM phorbol 12,13-dibutyrate.
Effect of apigenin on the level of MYPT1 phosphorylation at Thr-855
To confirm the role of apigenin on the thick filament regulation of smooth muscle contractility, we measured levels of myosin phosphatase targeting subunit 1 (MYPT1) and phospho-MYPT1 in muscles quick frozen after 60 min exposure to apigenin for the equilibration. Each relaxing ring was precontracted with 6 mM fluoride. This work was done using quick frozen apigenin (0.1 mM)-treated rat aortas in the absence of endothelium and the levels were compared to those of vehicle-treated rat aortas (Fig. 6). Interestingly, a significant decrease in fluoride-induced MYPT1 phosphorylation at Thr855 was found to be led by apigenin (Fig. 6). Thus, thick or myosin filament regulation including myosin phosphatase activation via RhoA/Rho-kinase inactivation might be involved in the reduced contractility of apigenin-treated rat aorta.
Fig. 6.
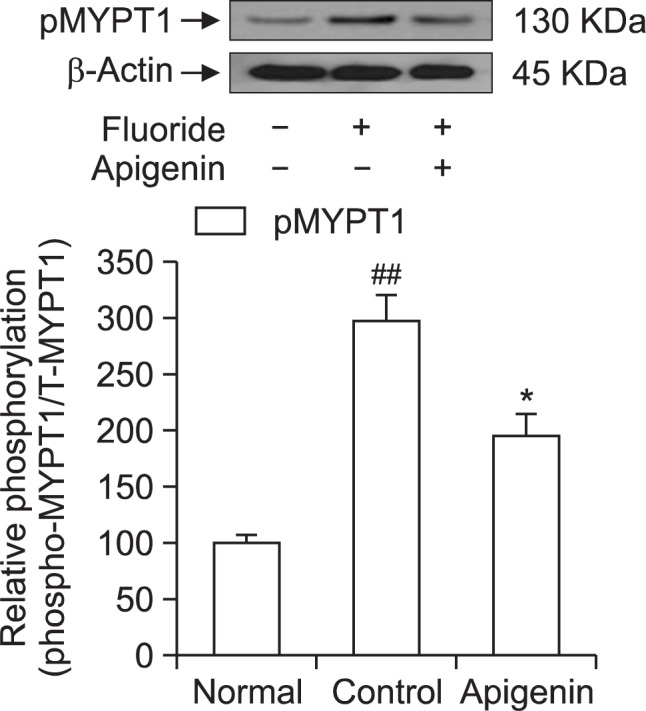
Effect of apigenin on fluoride-induced increases in phospho-MYPT1 levels. Phospho-MYPT1 protein levels were significantly decreased in quick frozen apigenin-treated rat aorta in the absence of endothelium compared to vehicle-treated rat aorta precontracted with fluoride. The upper panel shows a typical blot and the lower panel shows average densitometry results for relative levels of phospho-MYPT1. Data are expressed as the means of 3–5 experiments with vertical lines representing SEMs. *p<0.05, ##p<0.01, versus control or normal group respectively. Apigenin: 0.1 mM apigenin; Fluoride: 6 mM sodium fluoride.
Effect of apigenin on the level of MLC20 phosphorylation
In order to investigate whether apigenin affect fluoride-induced MLC20 phosphorylation, aortic rings were treated with apigenin after the addition of fluoride. Each relaxing ring was precontracted with 6 mM sodium fluoride. This work was done using quick frozen apigenin (0.1 mM)-treated rat aortas in the absence of endothelium and the levels were compared to those of vehicle-treated rat aortas (Fig. 7). Interestingly, a significant decrease in fluoride-induced LC20 phosphorylation was found to be led by apigenin (Fig. 7). Thus, thick or myosin filament regulation including calcium immobilization and MLCK inactivation might be involved in the reduced contractility of apigenin-treated rat aorta.
Fig. 7.
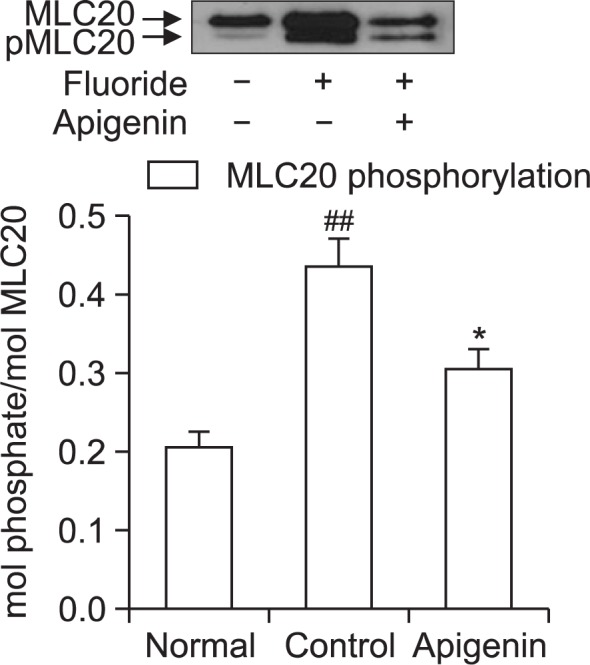
Effect of apigenin on fluoride-induced increases in phospho-MLC20 levels. Phospho-MLC20 levels expressed as a percentage to the total MLC20 were significantly decreased in quick frozen apigenin-treated rat aorta in the absence of endothelium compared to vehicle-treated rat aorta precontracted with fluoride. Data are expressed as the means of 3–5 experiments with vertical lines representing SEMs. *p<0.05, ##p<0.01, versus control or normal group respectively. Apigenin: 0.1 mM apigenin; Fluoride: 6 mM sodium fluoride.
DISCUSSION
The present study demonstrates that apigenin can modulate the vascular contractility in an agonist-dependent manner. Interestingly, the mechanism involved seems to be not only endothelium-dependent but also to involve the nonspecific inhibition of MEK and Rho-kinase activity. Apigenin has been previously recognized for its anti-inflammatory, antioxidant activity or endothelial vasorelaxation (Zhang et al., 2000). Therefore, we investigated whether the inhibition of RhoA/Rho-kinase or MEK activity contributes to apigenin-induced vascular relaxation in rat aortas denuded and precontracted by a RhoA/Rho-kinase activator fluoride or by a MEK activator phorbol ester.
The mechanism by which phorbol ester activates MEK/ERK has been established (Kordowska et al., 2006; Gu et al., 2007). On the other hand, previous studies that examined the mechanisms underlying arterial contractions induced by fluoride or thromboxane A2 mimetic have reported variable findings with regard to the contraction triggered by calcium entry and Rho-kinase activation (Wilson et al., 2005; Tsai and Jiang, 2006). These findings are consistent with the notion that apigenin can decrease phorbol ester or fluoride-induced contraction by inhibiting MEK or Rho-kinase activity.
The mechanisms by which MEK activation causes vascular contraction is an area of intense study, and several possibilities exist. The phosphorylation of caldesmon by MEK/ERK appears to regulate smooth muscle contractility (Kordowska et al., 2006). In this process MEK/ERK is activated by PKC which in turn can be stimulated by phorbol esters or GPCR receptor agonists.
The present study demonstrates that apigenin ameliorates the maximal or submaximal contraction induced by vasoconstrictor fluoride or phorbol ester endothelium-independently (Fig. 2, 4), and that this ameliorative mechanism involves the MEK/ERK and RhoA/Rho-kinase pathway. Previously, most vasodilation was believed to be caused by endothelial nitric oxide synthesis and the subsequent activation of guanylyl cyclase (Zhang et al., 2000). Zhang et al. reported apigenin-induced endothelium-dependent relaxation and related pathway involving cGMP production. In the present study, apigenin at a low concentration significantly inhibited phorbol ester-or fluoride-induced contraction regardless of endothelial function (Fig. 2, 4). Furthermore, apigenin decreased phosphorylation of extracellular signal-regulated kinase (ERK) 1/2 induced by phorbol ester (Fig. 5) and significantly decreased the phosphorylation of MYPT1 at Thr855 induced by fluoride (Fig. 6) with full relaxation (Fig. 2) suggesting the inhibition of Rhokinase or MEK activity as a major mechanism.
MLC20 is known to be phosphorylated by both MLCK and Rho-kinase (Somlyo and Somlyo, 1994). The activation of Rho-kinase by U46619 or fluoride inhibits the activity of myosin light chain phosphatase through the phosphorylation of MYPT1, leading to an increased MLC20 phosphorylation as well as contractions (Sakurada et al., 2003; Wilson et al., 2005). Coincidentally, fluoride phosphorylated MLC20, which was inhibited by apigenin (Fig. 7).
In summary, apigenin significantly attenuates the contractions induced by a MEK activator phorbol ester regardless of endothelial function. Furthermore, a Rho-kinase activator fluoride-induced contraction was significantly inhibited by apigenin at this low concentration. Thus, the mechanism underlying the relaxation induced by apigenin in phorbol ester or fluoride-induced contractions involves the nonspecific inhibition of MEK activity and Rho-kinase activity. Interestingly, during fluoride-induced contraction, the inhibition of Rho-kinase activity and subsequent MYPT1 phosphorylation induced by apigenin suggest that Rho-kinase inactivation is required for relaxation. In conclusion, in addition to endothelial nitric oxide synthesis (Zhang et al., 2000), both MEK and Rho-kinase inhibition make a major contribution to the mechanism responsible for apigenin-induced vasorelaxation in the denuded muscle and the decrease of intracellular calcium level which should be elucidated directly may be involved as well (Rotondo et al., 2009).
REFERENCES
- Caltagirone S, Rossi C, Poggi A, Ranelletti FO, Natali PG, Brunetti M, Aiello FB, Piantelli M. Flavonoids apigenin and quercetin inhibit melanoma growth and metastatic potential. Int J Cancer. 2000;87:595–600. doi: 10.1002/1097-0215(20000815)87:4<595::aid-ijc21>3.0.co;2-5. [DOI] [PubMed] [Google Scholar]
- Fotsis T, Pepper MS, Montesano R, Aktas E, Breit S, Schweigerer L, Rasku S, Wahala K, Adlercreutz H. Phytoestrogens and inhibition of angiogenesis. Baillieres Clin Endocrinol Metab. 1998;12:649–666. doi: 10.1016/s0950-351x(98)80009-8. [DOI] [PubMed] [Google Scholar]
- Gu Z, Kordowska J, Williams GL, Wang CL, Hai CM. Erk1/2 MAPK and caldesmon differentially regulate podosome dynamics in A7r5 vascular smooth muscle cells. Exp Cell Res. 2007;313:849–866. doi: 10.1016/j.yexcr.2006.12.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han JY, Ahn SY, Kim CS, Yoo SK, Kim SK, Kim HC, Hong JT, Oh KW. Protection of apigenin against kainate-induced excitotoxicity by anti-oxidative effects. Biol Pharm Bull. 2012;35:1440–1446. doi: 10.1248/bpb.b110686. [DOI] [PubMed] [Google Scholar]
- Jeon SB, Jin F, Kim JI, Kim SH, Suk K, Chae SC, Jun JE, Park WH, Kim IK. A role for Rho kinase in vascular contraction evoked by sodium fluoride. Biochem Biophys Res Commun. 2006;343:27–33. doi: 10.1016/j.bbrc.2006.02.120. [DOI] [PubMed] [Google Scholar]
- Kitazawa T, Masuo M, Somlyo AP. G-protein-mediated inhibition of myosin light-chain phosphatase in vascular smooth muscle. Proc Natl Acad Sci USA. 1991;88:9307–9310. doi: 10.1073/pnas.88.20.9307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kordowska J, Huang R, Wang CL. Phosphorylation of caldesmon during smooth muscle contraction and cell migration or proliferation. J Biomed Sci. 2006;13:159–172. doi: 10.1007/s11373-005-9060-8. [DOI] [PubMed] [Google Scholar]
- Nicholas C, Batra S, Vargo MA, Voss OH, Gavrilin MA, Wewers MD, Guttridge DC, Grotewold E, Doseff AI. Apigenin blocks lipopolysaccharide-induced lethality in vivo and proinflammatory cytokines expression by inactivating NF-κB through the suppression of p65 phosphorylation. J Immunol. 2007;179:7121–7127. doi: 10.4049/jimmunol.179.10.7121. [DOI] [PubMed] [Google Scholar]
- Peterson J, Dwyer J. Flavonoids: dietary occurrence and biochemical activity. Nutr Res. 1988;18:1995–2018. [Google Scholar]
- Rotondo A, Serio R, Mulè F. Gastric relaxation induced by apigenin and quercetin: analysis of the mechanism of action. Life Sci. 2009;85:85–90. doi: 10.1016/j.lfs.2009.04.022. [DOI] [PubMed] [Google Scholar]
- Sakurada S, Takuwa N, Sugimoto N, Wang Y, Seto M, Sasaki Y, Takuwa Y. Ca2+-dependent activation of Rho and Rho-kinase in membrane depolarization-induced and receptor stimulation-induced vascular smooth muscle contraction. Circ Res. 2003;93:548–556. doi: 10.1161/01.RES.0000090998.08629.60. [DOI] [PubMed] [Google Scholar]
- Somlyo AP, Somlyo AV. Signal transduction and regulation in smooth muscle. Nature. 1994;372:231–236. doi: 10.1038/372231a0. [DOI] [PubMed] [Google Scholar]
- Somlyo AP, Somlyo AV. From pharmacomechanical coupling to G-proteins and myosin phosphatase. Acta Physiol Scand. 1998;164:437–448. doi: 10.1046/j.1365-201X.1998.00454.x. [DOI] [PubMed] [Google Scholar]
- Touyz RM, Deng LY, He G, Wu XH, Schiffrin EL. Angiotensin II stimulates DNA and protein synthesis in vascular smooth muscle cells from human arteries: role of extracellular signal-regulated kinases. J Hypertens. 1999;17:907–916. doi: 10.1097/00004872-199917070-00006. [DOI] [PubMed] [Google Scholar]
- Tsai MH, Jiang MJ. Rho-kinase-mediated regulation of receptor-agonist-stimulated smooth muscle contraction. Pflugers Arch. 2006;453:223–232. doi: 10.1007/s00424-006-0133-y. [DOI] [PubMed] [Google Scholar]
- Uehata M, Ishizaki T, Satoh H, Ono T, Kawahara T, Morishita T, Tamakawa H, Yamagami K, Inui J, Maekawa M, Narumiya S. Calcium sensitization of smooth muscle mediated by a Rho-associated protein kinase in hypertension. Nature. 1997;389:990–994. doi: 10.1038/40187. [DOI] [PubMed] [Google Scholar]
- Wang W, Heideman L, Chung CS, Pelling JC, Koehler KJ, Birt DF. Cell-cycle arrest at G2/M and growth inhibition by apigenin in human colon carcinoma cell lines. Mol Carcinog. 2000;28:102–110. [PubMed] [Google Scholar]
- Wier WG, Morgan KG. α1-Adrenergic signaling mechanisms in contraction of resistance arteries. Rev Physiol Biochem Pharmacol. 2003;150:91–139. doi: 10.1007/s10254-003-0019-8. [DOI] [PubMed] [Google Scholar]
- Wilson DP, Susnjar M, Kiss E, Sutherland C, Walsh MP. Thromboxane A2-induced contraction of rat caudal arterial smooth muscle involves activation of Ca2+ entry and Ca2+ sensitization: Rho-associated kinase-mediated phosphorylation of MYPT1 at Thr-855, but not Thr-697. Biochem J. 2005;389:763–774. doi: 10.1042/BJ20050237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wooldridge AA, MacDonald JA, Erdodi F, Ma C, Borman MA, Hartshorne DJ, Haystead TA. Smooth muscle phosphatase is regulated in vivo by exclusion of phosphorylation of threonine 696 of MYPT1 by phosphorylation of Serine 695 in response to cyclic nucleotides. J Biol Chem. 2004;279:34496–34504. doi: 10.1074/jbc.M405957200. [DOI] [PubMed] [Google Scholar]
- Xu Q, Liu Y, Gorospe M, Udelsman R, Holbrook NJ. Acute hypertension activates mitogen-activated protein kinases in arterial wall. J Clin Invest. 1996;97:508–514. doi: 10.1172/JCI118442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang YH, Park YS, Kim TJ, Fang LH, Ahn HY, Hong JT, Kim Y, Lee CK, Yun YP. Endothelium-dependent vasorelaxant and antiproliferative effects of apigenin. Gen Pharmacol. 2000;35:341–347. doi: 10.1016/s0306-3623(02)00113-1. [DOI] [PubMed] [Google Scholar]