

Abstract
Background
While considerable evidence implicates DA D1-receptor signaling in the nucleus accumbens in motivation for cocaine during early stages of addiction, less is known regarding its role following the development of addiction. Here, we examined its role in the development of an addicted phenotype in intact male and female rats, and in female rats that were either resistant or vulnerable to developing this phenotype.
Methods
Intact males, females, and ovariectomized (OVX) females with and without estradiol (vulnerable, OVX+E; resistant, OVX+Veh) were given either short access (ShA; 3 fixed-ratio 1 sessions, maximum of 20 infusions) or 24-hr extended access (ExA) to cocaine for 10 days (4 trials/hr). Motivation for cocaine was assessed following a 14-day abstinence period using a progressive-ratio schedule. Once responding stabilized, the effects of intra-accumbens infusion of the D1-receptor antagonist, SCH-23390 (0, 0.3, 1.0, 3.0 μg), were examined.
Results
Motivation for cocaine was markedly higher following abstinence from ExA versus ShA self-administration in intact males and females, indicating the development of an addicted phenotype in these groups. Motivation for cocaine was also higher than ShA controls in OVX+E, but not OVX+Veh females following ExA self-administration, confirming the categorization of these groups as vulnerable versus resistant. Following ExA self-administration, intact males and females and OVX+E, but not OVX+Veh females, were less sensitive to the effects of D1-receptor antagonism as compared to their ShA counterparts.
Conclusions
These results suggest that the role of D1-receptor signaling, though critical in “non-addicted” stages, becomes diminished once addiction has developed.
Keywords: cocaine, estradiol, extended access, nucleus accumbens, SCH-23390, self-administration
INTRODUCTION
Cocaine addiction begins with casual use but progresses to compulsive use coupled with a heightened motivation to obtain the drug. Considerable evidence from studies in male animals modeling early stages of addiction, such as drug use initiation and maintenance intake under short access (ShA) conditions (1–2 hr/day), implicates dopamine (DA) signaling in the nucleus accumbens (NAc; a critical region of the reward pathway) as a primary mechanism motivating cocaine use (1–5). Cocaine increases DA in the NAc (6–8), and inhibiting this signaling pathway, particularly via DA D1-receptors, disrupts cocaine self-administration and the development of a cocaine-conditioned place preference (9–15). ShA cocaine self-administration also enhances markers of D1-receptor signaling (16–18), and some of these markers have been associated with an enhanced vulnerability to initiation of cocaine self-administration (19–23).
However, it is important to note that the evidence for the role of D1-receptors has been predominantly collected following ShA conditions. As such these neurobiological data, although characteristic of early vulnerability, do not necessarily characterize the neurobiology of cocaine addiction. An accumulating body of research indicates that the neurobiological mechanisms that underlie early “non-addicted” stages versus later “addicted” stages are different (1–2). Although evidence suggests that the progression to cocaine addiction is mediated by DA-induced neuroplastic changes in the mesolimbic pathway, its role in motivating cocaine use once addiction has developed is less clear. For example, neurobiological adaptations caused by chronic exposure to cocaine lead to mesolimbic hypofunction (24–27), which may in turn promote cocaine use to reverse DA deficits following abstinence (28). It has also been suggested that the role of DA in motivating cocaine use may become less critical following chronic cocaine exposure (1), with evidence to suggest that other signaling pathways become increasingly recruited and drive the enhanced motivation for the drug (eg, glutamatergic signaling; 1,29). In part, these mechanisms remain unknown because few experiments have examined the neuroadaptations associated with the development of an addicted phenotype. This phenotype, which has been defined as an enhanced motivation to obtain cocaine, develops following protracted abstinence (7 days or more) from extended access (ExA; 6–24 hr/day; 7 days or more), but not ShA self-administration (30–34). Although ExA procedures can be technically challenging to complete due to patency issues and lengthy experimental timelines, the information gained from such studies are critical for determining not only factors that predict a vulnerability to developing addiction, but also the neurobiological mechanisms associated with an addicted phenotype.
We recently showed that after 14 days of abstinence from ExA self-administration, both male and female rats developed an addicted state, with 9 of 11 males and 8 of 10 females showing a greater than 15% increase in motivation for cocaine (as measured under a progressive-ratio (PR) schedule) as compared to ShA controls (35). We also showed that in females ovarian hormones influence vulnerability to developing an addicted phenotype (35). Specifically, following abstinence from ExA self-administration, females with estradiol (eg, ovariectomized, OVX, females treated with estradiol, E) showed a markedly enhanced motivation for cocaine as compared to ShA controls (46.5% increase); whereas, those without estradiol (eg, OVX rats treated with vehicle, Veh), did not display this phenotype (1.7% difference; 35). These findings add to a growing body of work demonstrating that estradiol enhances vulnerability in females at each of the different phases of the addiction process (35–41), potentially through interactions with DA (42–44), and indicates that estradiol may be necessary for the development of addiction in females. Importantly, these distinct behavioral phenotypes were generated in groups of rats that all had ExA cocaine exposure, and as such, they provide a unique tool for assessing the neurobiology associated with the development of an addicted phenotype. Thus, the goal of the current study was to determine the potential mechanism for the development of an addicted phenotype in intact males and females tested following ShA versus ExA self-administration as well as groups of OVX females that were resistant (OVX+Veh) versus vulnerable (OVX+E) to developing this phenotype. We focused on the role of DA D1-receptor signaling in the NAc given the key role that this signaling pathway plays at other stages of the addiction process. Specifically, we evaluated the effect of site-specific infusion of the D1-receptor antagonist, SCH-23390, in the NAc on motivation for cocaine following abstinence from ShA versus ExA self-administration using the same access conditions that we previously tested (35). Based on the idea that the role of DA in motivating cocaine use may become less prominent with the development of addiction, we predicted that the effects of SCH-23390 on motivation for cocaine would be less pronounced following ExA versus ShA self-administration and in females vulnerable (OVX+E) versus resistant (OVX+Veh) to developing an addicted phenotype.
METHODS and MATERIALS
Subjects
Sexually mature intact male (N=20), female (N=25), and OVX female (N=51) Sprague-Dawley rats were used in this study. OVX rats were randomly assigned to a vehicle (OVX+Veh, resistant; 0.1 ml/day, 5 days/week) or estradiol-treated (OVX+E, vulnerable; 5.0 μg/ml/day, 5 days/week) group using conditions that have been shown previously to generate a vulnerable versus a resistant phenotype when responding was assessed following abstinence from ExA self-administration (35). Further details on the daily procedures for estradiol/vehicle treatment, as well as vaginal swabbing procedures to confirm hormonal status are provided in Supplementary Materials.
Procedures
The surgical methods used for jugular catheterization and brain cannulation targeting the core region of the NAc, and the procedures used for cocaine self-administration were conducted as previously described (19,35,45) and as described in more detail in Supplementary Materials. Briefly, rats were trained to self-administer cocaine infusions (1.5 mg/kg) under a fixed-ratio 1 schedule. Following acquisition, rats assigned to the ShA group were maintained under the same fixed-ratio conditions for three additional sessions, and rats assigned to the ExA group were given 24-hr/day access to cocaine (1.5 mg/kg/infusion) under a discrete trial procedure (10 min trials, 4 trials/hr) for a total of 10 days. Motivation for cocaine (0.5 mg/kg/infusion) was assessed under a PR schedule beginning following a 14-day abstinence period.
The effect of intra-NAc infusion of SCH-23390 (0.0, 0.3, 1.0, 3.0 μg/side) on PR responding for cocaine began once baseline responding (defined as three consecutive sessions with no increasing or decreasing trend in the number of infusions obtained) was established using a within-subject design. Infusions (0.5 ul/side) were administered immediately prior to the PR session and a minimum of 3 stable sessions separated each test session. Due to experimental circumstances associated with this lengthy protocol, each animal received between 2 and 4 doses, with a minimum of 6 animals tested under each of the dose conditions. Justification for the doses selected, and further details on the procedures used for SCH-23390 infusion, including placement verification (Figure S1), are provided in the Supplementary Materials. The effect of SCH-23390 was also examined on PR responding for sucrose in separate groups of intact male (N=5) and female (N=5) and OVX+E (N=8) and OVX+Veh (N=8) rats using procedures similar to those used for ShA cocaine self-administering controls (see Supplementary Materials for further details). Data were analyzed using repeated measures ANOVA with post-hoc comparisons to control/vehicle conditions made using the Dunnett t-test, and with comparisons within each day/dose made using the Bonferroni corrected t-test. Separate analyses were conducted within the intact groups of males and females and within the OVX groups of females and within the cocaine and sucrose groups. Further details are provided in Supplementary Materials.
RESULTS
ShA versus ExA Cocaine Self-Administration and Subsequent Motivation for Cocaine in Intact Males and Females
In contrast to ShA self-administration where cocaine intake was limited to a total of 20 infusions/day (30 mg/kg/day), high levels of intake were observed under ExA conditions where the average number of infusions over the 10-day period was 67.3±5.5 (approximately 100 mg/kg/day). As expected, females self-administered significantly more cocaine infusions than did males under ExA conditions (averaged over 10-days: 71.2±6.1 and 61.6±8.3, respectively; t10=4.52, p<0.05). Also as expected, ExA rats maintained markedly higher levels of PR responding for cocaine during the first three stable sessions following abstinence as compared to ShA controls (Figure 1; group effect, [F(1,31)=13.14, p=0.001]), with a significant group difference for all three sessions (Day 1, t33=3.33, p<0.01; Day 2, t33=3.85, p<0.001; Day 3, t33=3.54, p<0.001). Subsequent PR responding was similar in males and females with results from the overall analysis, as well as subsequent analyses within the ShA and ExA groups, revealing non-significant effects of sex (p's>0.05). Thus, females and males showed a similar level of motivation for cocaine following both ShA and ExA self-administration, and both showed a similar increase in motivation for cocaine following ExA self-administration. These results confirm the development of an addicted phenotype in both sexes following abstinence from ExA self-administration.
Figure 1. ShA versus ExA self-administration on subsequent motivation for cocaine in intact males and females.
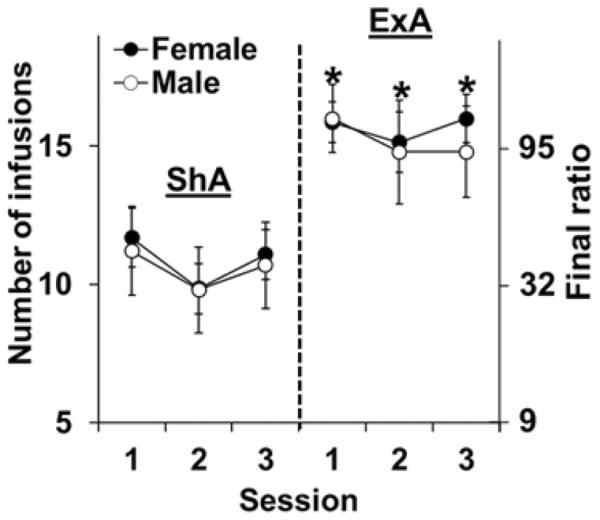
Mean (± SEM) number of cocaine infusions obtained and the corresponding final ratios completed during the first three stable PR sessions in males (open circles) and females (filled circles) following either ShA (N=10 males, N=13 females) or ExA (N=5 males, N=7 females) self-administration and a 14-day abstinence period. *Significant group difference (p<0.05). ShA=short access, ExA=extended access.
Effect of SCH-23390 on Motivation for Cocaine following ShA versus ExA Self-Administration in Intact Males and Females
Bilateral infusion of SCH-23390 into the NAc dose-dependently decreased PR responding for cocaine following ShA but not ExA self-administration. The effects of SCH-23390 were similar in males and females under all conditions (p's>0.05), and thus the data are presented collapsed across sex (Figure 2). A repeated measures ANOVA comparing the number of infusions obtained at baseline to the day of SCH-23390 treatment revealed significant effects of access condition [F(1,77)=9.76, p<0.01], day [F(1,77)=15.81, p<0.001], day-by-dose [F(3,77)=4.47, p<0.01], day-by-access condition [F(1,77)=4.20, p<0.05], and day-by-dose-by-access condition [F(3,77)=3.90, p<0.05]. As observed for PR responding during the first three days of access, PR responding averaged across each of the pretreatment baselines was significantly higher in the ExA versus ShA group (t33=3.38, p=0.05). Within the ShA group (Figure 2A), there were significant effects of day [F(1,53)=26.56, p<0.001], and day-by-dose [F(3,53)=10.01, p<0.001], with post-hoc comparison within each dose revealing significant decreases from baseline following the 1.0 (p<0.01) and 3.0 μg doses (p<0.001). There were no significant effects of SCH-23390 within the ExA group (Figure 2B). Similar results were observed when the data were analyzed as percent change from baseline (Figure 2C), with repeated measures ANOVA revealing significant effects of dose [F(3,77)=4.72, p<0.01], access condition [F(1,77)=4.13, p<0.05], and dose-by-access condition [F(3,77)=4.6, p<0.01]. Analysis within the ShA group revealed a significant effect of dose [F(3,53)=11.3, p<0.001], with post-hoc comparison revealing significant differences from vehicle for the 1.0 (p<0.01) and 3.0 μg doses (p<0.001). As with the analysis of the absolute number of infusions, no significant effects of SCH-23390 were observed on percent change from baseline in the ExA group. Thus, while D1-receptor antagonism dose-dependently reduced motivation for cocaine following ShA self-administration, it was without effect following ExA self-administration.
Figure 2. Effect of SCH-23390 on motivation for cocaine following ShA versus ExA self-administration in intact males and females.
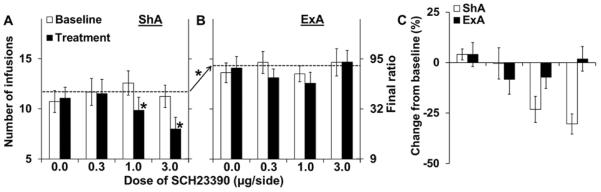
Mean (± SEM) number of cocaine infusions obtained and the corresponding final ratios completed in (A) ShA and (B) ExA rats at baseline (open bars) and on the day of SCH-23390 treatment (filled bars; 0, 0.3, 1.0, and 3.0 μg). (C) Mean (± SEM) percent change from baseline in the number of infusions obtained on the day of treatment (ShA, open bars; ExA, filled bars). Dashed line indicates average pre-treatment baseline (A–B). * indicates significant difference from pre-treatment baseline (A) or from vehicle (C) for each individual dose. Arrow with asterisk indicates significant difference in number of infusions obtained averaged across the pre-treatment baseline (A–B). ShA, N= 13–17; ExA: N=7–10 for each dose of SCH-23390.
ShA versus ExA Cocaine Self-Administration and Subsequent Motivation for Cocaine in the Vulnerable (OVX+E) versus the Resistant (OVX+Veh) Group
As expected, the vulnerable group self-administered significantly more cocaine infusions as compared to the resistant group during the 10-day ExA period (74.2±2.4 versus 67.3±2.1, respectively; t15=2.3, p<0.05). Also as predicted, the vulnerable, but not the resistant group, showed markedly higher levels of responding for cocaine during the first three stable PR sessions following abstinence from ExA self-administration as compared to ShA controls (Figure 3; group [F(1,31)=4.33, p<0.05]; access condition [F(1,31)=5.54, p<0.05]). Comparison within the vulnerable group revealed significant differences between ExA versus ShA groups for each sessions (p's<0.05). A significant difference was also observed within the two ExA groups [F(1,16)=9.87, p<0.001], with significant differences observed between the vulnerable versus resistant groups for each of the three session (p's<0.05). Thus, the vulnerable group (OVX+E rats) self-administered more cocaine under ExA conditions and following abstinence from ExA self-administration, showed a greater level of motivation to obtain cocaine as compared to the resistant group (OVX+Veh rats). These results confirm the development of an addicted phenotype in the vulnerable, but not the resistant group, following ExA self-administration.
Figure 3. Effect of ShA versus ExA self-administration on motivation for cocaine in the vulnerable versus the resistant group.
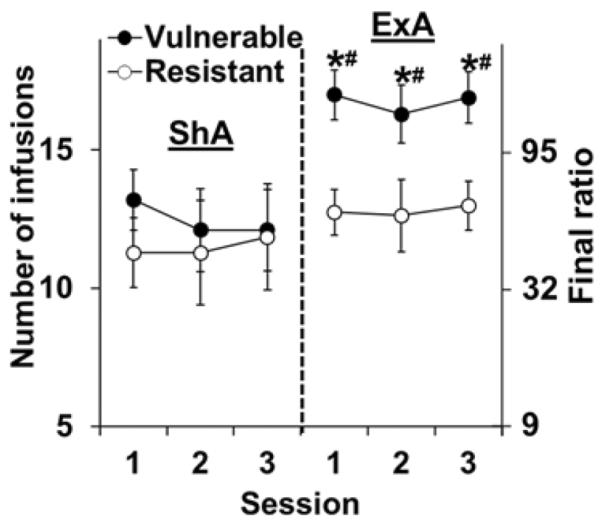
Mean (± SEM) number of cocaine infusions obtained and the corresponding final ratios completed during the first three stable PR sessions for the vulnerable (OVX+E, filled circles) and the resistant (OVX+Veh, open circles) groups tested following either ShA (vulnerable, N=10; resistant, N=7) or ExA (vulnerable, N=10; resistant, N=8) self-administration and a 14-day abstinence period. * represents a significant difference from the ExA resistant group; # represents a significant difference from ShA controls.
Effect of SCH-23390 on Motivation for Cocaine following ShA versus ExA Self-Administration in the Vulnerable versus the Resistant Group
Bilateral infusion of SCH-23390 into the NAc decreased PR responding for cocaine in both of the OVX-ShA control groups, as well as in the ExA resistant group; however, its effects were diminished in the ExA vulnerable group (Figure 4). Since the effects of SCH-23390 were similar in both OVX ShA control groups under all conditions (p's>0.05), the data are presented collapsed across the groups (ShA: OVX controls; but see Supplement: Figure S2 which illustrates the similarity in the effects of SCH-23390 on PR responding for cocaine between the vulnerable and resistant ShA control groups). A repeated measures ANOVA comparing the number of infusions obtained at baseline to the day of SCH-23390 treatment across all groups of OVX rats revealed significant effects of day [F(1,83)=67.09, p<0.001], hormone group [F(1,83)=7.67, p<0.01], access condition [F(1,83)=7.26, p<0.01], day-by-hormone group [F(1,83)=6.00, p<0.05], day-by-dose [F(3,83)=12.99, p<0.001], day-by-hormone group-by-access condition [F(1,83)=4.00, p<0.05], and day-by-dose-by-access condition [F(3,83)=3.16, p<0.05]. Within the ShA condition, repeated measures ANOVA revealed significant effects of day [F(1,33)=41.45, p<0.001], and day-by-dose [F(3,33)=8.22, p<0.001], with post-hoc comparison revealing a significant decrease from baseline for each dose of SCH-23390 (0.3, p<0.05; 1.0, p<0.001; 3.0, p<0.001). In contrast, within the ExA condition we observed significant effects of hormone group [F(1,50)=12.37, p=0.001], and day-by-hormone group [F(1,50)=10.94, p<0.01]. We also observed significant effects of day [F(1,50)=30.02, p<0.001], and day-by-dose [F(3,50)=9.43, p<0.001]. Subsequent analysis within the ExA resistant group (OVX+Veh) revealed significant effects of day [F(1,26)=75.40, p<0.001] and day-by-dose [F(3,26)=12.18, p<0.001] with post-hoc comparison revealing that each dose of SCH-23390 decreased responding as compared to baseline (0.3, p<0.01; 1.0, p<0.001; 3.0, p=0.001). No significant effects of SCH-23390 were observed within the ExA vulnerable group although there was a trend for an effect of day-by-dose (p=0.061). This trend appears to be accounted for by the effects of the highest dose of SCH-23390 (3.0 μg) where responding tended to be decreased from baseline. In fact, comparison of the ExA resistant and vulnerable groups showed that while the effects of the 0.3 (p<0.05) and 1.0 μg (p=0.01) doses were different between the two groups, the effects of the 3.0 μg dose did not differ indicating that this dose produced similar effects in both groups. A comparison of the ShA controls to the ExA vulnerable group revealed significant effects of access condition [F(1,57)=5.73, p<0.05], and day-by-dose-by-access condition [F(3,57)=3.55, p<0.05] with post-hoc comparison within each dose revealing a significant difference for the 1.0 dose (p<0.05), and a trend for an effect at the 3.0 dose (p=0.08). Notably, no significant differences for the effects of SCH-23390 were observed between the ShA controls and the ExA resistant group (p's>0.05).
Figure 4. Effect of SCH-23390 on motivation for cocaine following ShA versus ExA self-administration in the vulnerable versus the resistant group.
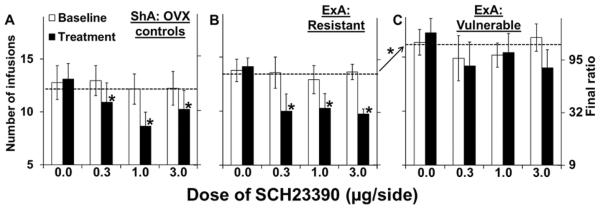
Mean (± SEM) number of infusions obtained in (A) ShA controls, (B) ExA Resistant, and (C) ExA Vulnerable rats on the day of treatment with SCH-23390 (0, 0.3, 1.0, and 3.0 μg; filled bars) as compared to the pre-treatment baseline (open bars) for each dose. Dashed line indicates average pre-treatment baseline. Arrow with asterisk indicates significant difference in number of infusions obtained averaged across the pre-treament baseline. *Significant difference from pre-treatment baseline. ShA, N=9–11; ExA resistant, N=7–8; ExA vulnerable, N=6–8 for each dose of SCH-23390.
Similar results were observed when the data were analyzed as percent change from baseline (Figure 5). This analysis revealed significant effects of dose [F(3,87)=12.82, p<0.001], and access-by-hormone condition [F(2,87)=6.85, p<0.01]. Within ShA controls, there was a significant effect of dose [F(3,37)=5.70, p<0.01], with post-hoc comparison showing a significant difference for each of the three doses (p<0.05) compared to vehicle. Within the ExA resistant group there was a significant overall effect of dose [F(3,26)=14.26, p<0.001], with significant effects for all three doses of SCH-23390 as compared to vehicle (p's<0.001). Within the ExA vulnerable group, although the 3.0 dose tended to reduce PR responding, the overall effect of dose did not reach statistical significance (p=0.091). Between groups comparison within each dose revealed a significant effect of group for the 0.3 and 1.0 doses (p's<0.01), but not for the 3.0 dose (p>0.05). Thus, while D1-receptor antagonism reduced motivation for cocaine similarly in the OVX ShA controls groups as compared to the ExA resistant group (OVX+Veh), its effects were diminished in the ExA vulnerable group (OVX+E).
Figure 5. Effect of SCH-23390 on motivation for cocaine in the ShA controls as compared to the ExA vulnerable and resistant groups.
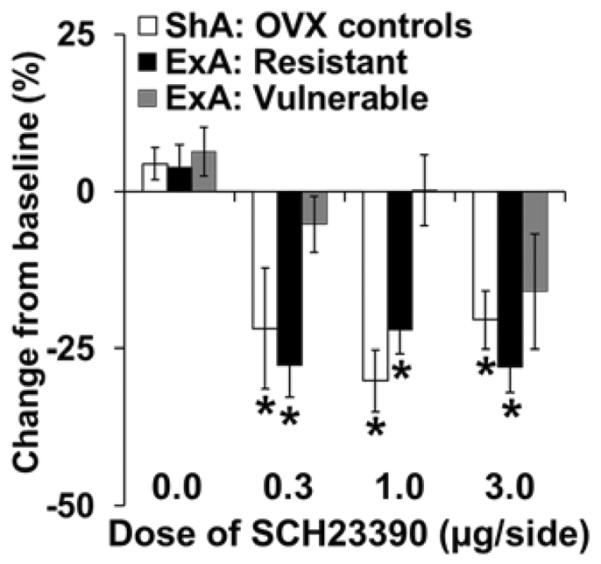
Mean (± SEM) percent change from baseline in the number of infusions obtained on the day of treatment with 0, 0.3, 1.0, or 3.0 μg of SCH23990. ShA controls, open bars; ExA Resistant, black bars; ExA Vulnerable, gray bars. *Significant difference from vehicle. ShA, N=9–11; ExA resistant, N=7–8; ExA vulnerable, N=6–8 for each dose of SCH-23390.
Effect of SCH-23390 on Motivation for Sucrose
In order to control for potential non-specific effects of SCH-23390, we also measured its effect on PR responding for sucrose pellets in intact male and female rats as well as OVX+E and OVX+Veh rats (Supplement: Figure S3). PR responding for sucrose did not differ between males and females (data not shown; p>0.05), or between OVX+E and OVX+Veh rats (data not shown; p>0.05). No significant effects of SCH-23390 were observed in any of the groups (p's>0.05). Thus, SCH-23390 did not affect general motivational behaviors.
DISCUSSION
The goal of this study was to examine the role of D1-receptor signaling in the NAc in the development of an addicted phenotype using intact males and females tested following ShA versus ExA self-administration as well as groups of OVX females that were resistant (OVX+Veh) versus vulnerable (OVX+E) to developing this phenotype. Consistent with our previous findings (35), motivation for cocaine was markedly higher following abstinence from ExA versus ShA self-administration in intact males and females, confirming the development of an addicted phenotype in these groups. In addition, following ExA self-administration, OVX+E females, but not OVX+Veh females, displayed a higher motivation for cocaine as compared to ShA controls, thus confirming the categorization of these groups as vulnerable versus resistant. Consistent with our hypothesis, we demonstrated that following ExA self-administration, intact males and females and OVX+E vulnerable females were less sensitive to the effects of D1-receptor antagonism as compared to ShA controls. Notably, in the group of rats resistant to developing an addicted phenotype (OVX+Veh), the effects of D1-receptor antagonism did not differ from ShA controls. SCH-23390 had no impact on motivation for sucrose pellets indicating that these effects were specific to cocaine. These results indicate that the role of D1-receptor signaling in motivating cocaine use, though critical in “non-addicted” stages, becomes diminished once addiction has developed.
Our findings in ShA controls and in the ExA resistant group (OVX+Veh) are consistent with a large body of work indicating a key role for D1-receptor signaling in the NAc under ShA conditions and during “non-addicted” stages (9–15). In contrast, following ExA self-administration the effects of D1-receptor antagonism were diminished in intact males and females and in the OVX+E vulnerable group suggesting that the role of D1-receptor signaling in the NAc becomes diminished with the development of an addicted phenotype. Further support for this idea is provided by previous results showing that chronic cocaine exposure and ExA cocaine self-administration produce a hypo-DAergic state in the NAc and lead to a decreased sensitivity of DA terminals in this brain region to cocaine (28,46–48). Results have also shown that during ExA self-administration rats maintain higher steady-state levels of DA in the NAc as compared to ShA rats (49), a finding hypothesized to be a compensatory response to counteract a decrease in DA receptor function (49). In contrast to these findings, D1-receptor antagonism has been reported to effectively decrease PR responding for cocaine following both ShA and ExA self-administration (11), and an increased sensitivity to systemic treatment with a non-selective D1/D2-receptor antagonist has been reported in rats given ExA versus ShA to cocaine (50). However, in both of these previous studies, responding was assessed following ExA self-administration without an intervening abstinence period, and thus, it is unlikely that the animals tested had developed an addicted phenotype. Collectively, these results suggest that increases in D1-receptor sensitivity may occur during ShA exposure, as well as during escalation of intake without abstinence, but that following ExA self-administration and the development of an addicted phenotype, there is a shift in the role of D1-receptor signaling.
Although these findings indicate a diminished role for D1-receptor signaling with the development of an addicted phenotype, it is still possible that signaling via the D2-receptor remains a prominent mechanism. D2-receptor signaling is also known to play a critical role in motivating cocaine use with results showing that manipulation of either the D1- or the D2-receptor similarly affect cocaine self-administration under ShA conditions (14). Thus, one possibility is that D1- and D2-receptors play a similar role under ShA conditions, but following chronic exposure and with the development of an addicted phenotype, their roles change. There is evidence to suggest that following chronic cocaine exposure there is a dysregulation of the balance between D1- and D2-receptor signaling (23), and evidence showing that changes in D2-receptors following ExA self-administration persist longer than changes in D1-receptors (46). Further research is needed to examine the role of D2-receptors in motivating cocaine use following the development of an addicted phenotype.
The two groups of OVX rats, resistant and vulnerable, provide an important tool to elucidate the mechanisms of the addicted phenotype. Although both groups self-administered a high level of cocaine under ExA conditions, only the vulnerable group (OVX+E) developed the addicted phenotype (35), and showed a reduced sensitivity to the effects of D1-receptor antagonism. Although OVX+E rats self-administered higher levels of cocaine as compared to the OVX+Veh rats under ExA conditions, levels of cocaine prior to abstinence were comparable between the two groups (ie, we built a control into the study where following ExA self-administration each animal was limited to 20 infusions/day for 2 sessions). Additionally, levels of intake under ExA conditions were comparable in the OVX+Veh group as compared to the intact groups of males and females (average infusions: 67.3±2.1 versus 67.3±5.5, respectively; also see 35), and these intact groups did develop an addicted phenotype and show a reduced sensitivity to the effects of D1-receptor antagonism. Taken together, these results suggest that these differential effects of D1-receptor antagonism are not a result of chronic exposure per se, but rather a marker for the development of addiction.
It is unlikely that the differences we found in sensitivity to D1-receptor antagonism between our ExA vulnerable and resistant groups are simply due to estradiol treatment. Although estradiol treatment does appear to be essential for the development of an addicted phenotype in female rats, we argue that it is the development of this phenotype, whether caused by estradiol treatment or not, that underlies this change in sensitivity. Support for this idea is provided by our findings showing that although females are more vulnerable than males to developing an addicted phenotype (34), under optimized conditions, like the ones used here, both males and females display a similarly enhanced level of motivation for cocaine (35), and as shown here, both groups show a similar reduction in sensitivity to the effects of D1-receptor antagonism. There is also a significant body of work showing that estradiol treatment enhances rather than attenuates DA signaling under acute and ShA conditions (42–43,52). In fact, the predominant theory to explain an enhanced vulnerability in females versus males to the reinforcing effects of cocaine (38, 51–56) is through estradiol augmentation of DA signaling, particularly D1-receptor signaling (42–43,57–59). Estradiol has been shown to increase both basal and pyschostimulant-induced DA release in the reward pathway (42), including the NAc (57), and markers of D1-receptor signaling are increased by estradiol treatment (60–61). Interestingly, whereas intact ShA rats in this study exhibited a typical dose-dependent decrease in PR responding for cocaine following SCH-23390 treatment (11), the ShA OVX females showed robust and equivalent decreases in PR responding to each of the three doses of SCH-23390. Although not yet examined, this increased sensitivity to D1-receptor antagonism in the OVX females may be due to altered estradiol/DA interactions resulting from the loss of normal ovarian hormone function. Further research is necessary to address this possibility. Taken together, these results, in addition to informing on the underlying mechanism of addiction, also have important implications for mechanisms of sex and hormonal influences in addiction. Specifically, these findings indicate that in contrast to early “non-addicted” stages, following the development of addiction, estradiol does not appear to mediate enhanced vulnerability in females via D1-receptor signaling.
The results from studies conducted under ShA conditions have revealed DAergic signaling as a major modulator of cocaine-taking behavior (1–5,9–15,62–63), and these findings have served as a basis for the development of pharmacotherapies for treating cocaine addiction in humans. Our current findings suggest a diminished role of D1-receptor signaling with the development of an addicted phenotype which could account for the disappointing results of DA-based pharmacotherapies for cocaine addiction. However, it is still possible that the role of D2-receptor signaling remains a prominent mechanism underlying motivation for cocaine, and future research is needed to address this possibility. Additionally, a growing body of work suggests that with the development of addiction, other signaling pathways may become increasingly recruited (glutamatergic signaling; for review see 1–2,29) and drive the enhanced motivation to obtain cocaine (64–65). Further research using ExA conditions that induce an addicted phenotype are needed to uncover the role of other signaling pathways as potential mechanisms of cocaine addiction.
Supplementary Material
Acknowledgements
This work was supported by NIDA grant R01DA024716 (WJL). We thank Catherine Moore, Diana Naim, Daniel R. Verhotz, Shaun Toor, Garrett Garber, and Joshua Newman for their technical assistance.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Financial Disclosures All authors report no biomedical financial interests or potential conflicts of interest.
References
- 1.Kalivas PW, Volkow ND. The neural basis of addiction: a pathology of motivation and choice. Am J Psychiatry. 2005;162:1403–13. doi: 10.1176/appi.ajp.162.8.1403. [DOI] [PubMed] [Google Scholar]
- 2.Koob GF, Volkow ND. Neurocircuitry of addiction. Neuropsychopharmacology. 2009;35:217–238. doi: 10.1038/npp.2009.110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Nestler EJ. The neurobiology of cocaine addiction. Sci Pract Perspect. 2005;3:4–10. doi: 10.1151/spp05314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Pierce RC, Kumaresan V. The mesolimbic dopamine system: the final common pathway for the reinforcing effect of drugs of abuse. Neurosci Behav Rev. 2006;30:215–238. doi: 10.1016/j.neubiorev.2005.04.016. [DOI] [PubMed] [Google Scholar]
- 5.Willuhn I, Wanat MJ, Clark JJ, Phillips PE. Dopamine signaling in the nucleus accumbens of animals self-administering drugs of abuse. Curr Top Behav Neurosci. 2010;3:29–71. doi: 10.1007/7854_2009_27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Carboni E, Imperato A, Perezzani L, Di Chiara G. Amphetamine, cocaine, phencyclidine and nomifensine increase extracellular dopamine concentrations preferentially in the nucleus accumbens of freely moving rats. Neuroscience. 1989;28(3):653–61. doi: 10.1016/0306-4522(89)90012-2. [DOI] [PubMed] [Google Scholar]
- 7.Di Chiari G, Imperato A. Drugs abused by humans preferentially increase synaptic dopamine concentrations in the mesolimbic system of freely moving rats. Proc Natl Acad Sci. 1988;85:5274–5278. doi: 10.1073/pnas.85.14.5274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Hernandez L, Hoebel BG. Food reward and cocaine increase extracellular dopamine in the nucleus accumbens as measured by microdialysis. Life Sci. 1988;42:1705–1712. doi: 10.1016/0024-3205(88)90036-7. [DOI] [PubMed] [Google Scholar]
- 9.Bathcell RK, Whisler K, Karanian D, Self DW. Effects of intranucleus accumbens shell administration of dopamine agonists and antagonists on cocaine-taking and cocaine-seeking behaviors in the rat. Psychopharmacology. 2005;183:41–53. doi: 10.1007/s00213-005-0133-1. [DOI] [PubMed] [Google Scholar]
- 10.Caine SB, Heinrichs SC, Coffin VL, Koob GF. Effects of the dopamine D-1 antagonist SCH 23390 microinjected into the accumbens, amygdala or striatum on cocaine self-administration in the rat. Brain Res. 1995;692(1–2):47–56. doi: 10.1016/0006-8993(95)00598-k. [DOI] [PubMed] [Google Scholar]
- 11.McGregor A, Roberts DC. Dopaminergic antagonism within the nucleus accumbens or the amygdala produces differential effects on the intravenous cocaine self-administration under fixed and progressive ratio schedules of reinforcement. Brain Res. 1993;624:245–52. doi: 10.1016/0006-8993(93)90084-z. [DOI] [PubMed] [Google Scholar]
- 12.Bari AA, Pierce RC. D1-like and D2 dopamine receptor antagonists administered into the shell subregion of the rat nucleus accumbens decrease cocaine, but not food, reinforcement. Neuroscience. 2005;135:959–68. doi: 10.1016/j.neuroscience.2005.06.048. [DOI] [PubMed] [Google Scholar]
- 13.Maldonado R, Robledo P, Chover AJ, Caine SB, Koob GF. D1 dopamine receptors in the nucleus accumbens modulate cocaine self-administration in the rat. Pharmacol Biochem Behav. 1993;45(1):239–42. doi: 10.1016/0091-3057(93)90112-7. [DOI] [PubMed] [Google Scholar]
- 14.Robledo P, Maldonado-Lopez R, Koob GF. Role of dopamine receptors in the nucleus accumbens in the rewarding properties of cocaine. Ann N Y Acad Sci. 1992;654:509–512. doi: 10.1111/j.1749-6632.1992.tb26015.x. [DOI] [PubMed] [Google Scholar]
- 15.Baker DA, Fuchs RA, Specio SE, Khroyan TV, Neisewander JL. Effects of intraaccumbens administration of SCH-23390 on cocaine-induced locomotion and conditioned place preference. Synapse. 1998;30(2):181–93. doi: 10.1002/(SICI)1098-2396(199810)30:2<181::AID-SYN8>3.0.CO;2-8. [DOI] [PubMed] [Google Scholar]
- 16.Laurier LG, Corrigall WA, George SR. Dopamine receptor density, sensitivity and mRNA levels are altered following self-administration of cocaine in the rat. Brain Res. 1994;634(1):31–40. doi: 10.1016/0006-8993(94)90255-0. [DOI] [PubMed] [Google Scholar]
- 17.Moore RJ, Vinsant SL, Nader MA, Porrino LJ, Friedman DP. Effect of cocaine self-administration on striatal dopamine D1 receptors in rhesus monkeys. Synapse. 1998;28(1):1–9. doi: 10.1002/(SICI)1098-2396(199801)28:1<1::AID-SYN1>3.0.CO;2-G. [DOI] [PubMed] [Google Scholar]
- 18.Graziella De Montis M, Co C, Dworkin SI, Smith JE. Modifications of dopamine D1 receptor complex in rats self-administering cocaine. Eur J Pharmacol. 1998;362(1):9–15. doi: 10.1016/s0014-2999(98)00731-6. [DOI] [PubMed] [Google Scholar]
- 19.Lynch WJ, Taylor JR. Persistent changes in motivation to self-administer cocaine following modulation of cyclic AMP-dependent protein kinase A (PKA) activity in the nucleus accumbens. Eur J Neurosci. 2005;22:1214–1220. doi: 10.1111/j.1460-9568.2005.04305.x. [DOI] [PubMed] [Google Scholar]
- 20.Self DW, Genova LM, Hope BT, Barnhart WJ, Spencer JJ, Nestler EJ. Involvement of cAMP-dependent protein kinase in the nucleus accumbens in cocaine self-administration and relapse of cocaine-seeking behavior. J Neurosci. 1998;18:1848–1859. doi: 10.1523/JNEUROSCI.18-05-01848.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Caine SB, Thomsen M, Gabriel KI, Berkowitz JS, Gold LH, Koob GF, et al. Lack of self-administration of cocaine in dopamine D1 receptor knock-out mice. J Neurosci. 2007;27(48):13140–50. doi: 10.1523/JNEUROSCI.2284-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Zhang J, Zhang L, Jiao J, Zhang Q, Zhang D, Xu M, et al. c-Fos facilitates the acquisition and extinction of cocaine-induced persistent changes. J Neurosci. 2006;26:13287–13296. doi: 10.1523/JNEUROSCI.3795-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Anderson SM, Pierce RC. Cocaine-induced alterations in dopamine receptor signaling: implications for reinforcement and reinstatement. Pharmacol Ther. 2005;106:389–403. doi: 10.1016/j.pharmthera.2004.12.004. [DOI] [PubMed] [Google Scholar]
- 24.Koeltzow TE, White FJ. Behavioral depression during cocaine withdrawal is associated with decreased spontaneous activity of ventral tegmental area dopamine neurons. Behav Neurosci. 2003;117:860. doi: 10.1037/0735-7044.117.4.860. [DOI] [PubMed] [Google Scholar]
- 25.Maisonneuve IM, Ho A, Kreek MJ. Chronic administration of a cocaine “binge” alters basal extracellular levels in male rats: an in vivo microdialysis study. J Pharmacol Exp Ther. 1995;272:652–657. [PubMed] [Google Scholar]
- 26.Paulson PE, Camp DM, Robinson TE. Time course of transient behavioral depression and persistent behavioral sensitization in relation to regional brain monoamine concentrations during amphetamine withdrawal in rats. Psychopharmacology. 1991;103:480–492. doi: 10.1007/BF02244248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Chao J, Nestler EJ. Molecular neurobiology of drug addiction. Annu Rev Med. 2004;55:113–132. doi: 10.1146/annurev.med.55.091902.103730. [DOI] [PubMed] [Google Scholar]
- 28.Melis M, Spiga S, Diana M. The dopamine hypothesis of drug addiction: hypodopaminergic state. Int Rev Neurobiol. 2005;63:101–154. doi: 10.1016/S0074-7742(05)63005-X. [DOI] [PubMed] [Google Scholar]
- 29.Wolf ME, Mangiavacchi S, Sun X. Mechanisms by which dopamine receptors may influence synaptic plasticity. Acad Sci. 2003;1003:241–9. doi: 10.1196/annals.1300.015. [DOI] [PubMed] [Google Scholar]
- 30.Roberts DC, Morgan D, Liu Y. How to make a rat addicted to cocaine. Prog Neuropsychopharmacol Biol Psychiatry. 2007;31:1614–1624. doi: 10.1016/j.pnpbp.2007.08.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ahmed SH. The science of making drug-addicted animals. Neuroscience. 2012;211:107–125. doi: 10.1016/j.neuroscience.2011.08.014. [DOI] [PubMed] [Google Scholar]
- 32.Liu Y, Roberts DC, Morgan D. Sensitization of the reinforcing effects of self-administered cocaine in rats: effects of dose and intravenous injection speed. Eur J Neurosci. 2005;22:195–200. doi: 10.1111/j.1460-9568.2005.04195.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Morgan D, Brebner K, Lynch WJ, Roberts DC. Increases in the reinforcing efficacy of cocaine after particular histories of reinforcement. Behav Pharmacol. 2002;13:389–396. doi: 10.1097/00008877-200209000-00012. [DOI] [PubMed] [Google Scholar]
- 34.Lynch WJ, Taylor JR. Sex differences in the behavioral effects of 24-hr access to cocaine under a discrete trial procedure. Neuropsychopharmacology. 2004;29:943–951. doi: 10.1038/sj.npp.1300389. [DOI] [PubMed] [Google Scholar]
- 35.Ramôa CP, Doyle SE, Naim DW, Lynch WJ. Estradiol as a mechanism for sex differences in the development of an addicted phenotype following extended access cocaine self-administration. Neuropsychopharmacology. 2013 doi: 10.1038/npp.2013.68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Segarra A, Agosto-Rivera J, Febo M, Lugo-Escobar N, Menéndez-Delmestre R, Puig-Ramos A, et al. Estradiol: a key biological substrate mediating the response to cocaine in female rats. Horm Behav. 2010;58:33–43. doi: 10.1016/j.yhbeh.2009.12.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Hu M, Becker JB. Acquisition of cocaine self-administration in ovariectomized female rats: effect of estradiol dose or chronic estradiol administration. Drug Alcohol Depend. 2008;94:56–62. doi: 10.1016/j.drugalcdep.2007.10.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Lynch WJ, Roth ME, Carroll ME. Biological basis of sex differences in drug abuse: preclinical and clinical studies. Psychopharmacology. 2002;164:121–137. doi: 10.1007/s00213-002-1183-2. [DOI] [PubMed] [Google Scholar]
- 39.Larson EB, Anker JJ, Gliddon LA, Fons KS, Carroll ME. Effects of estrogen and progesterone on the escalation of cocaine self-administration in female rats during extended access. Exper Clin Psychopharmacol. 2007;15:461. doi: 10.1037/1064-1297.15.5.461. [DOI] [PubMed] [Google Scholar]
- 40.Lynch WJ, Roth ME, Mickelberg JL, Carroll ME. Role of estrogen in the acquisition of intravenously self-administered cocaine in female rats. Pharmacol Biochem Behav. 2001;68(4):641–6. doi: 10.1016/s0091-3057(01)00455-5. [DOI] [PubMed] [Google Scholar]
- 41.Jackson LR, Robinson TE, Becker JB. Sex differences and hormonal influences on acquisition of cocaine self-administration in rats. Neuropsychopharmacology. 2006;31:129–138. doi: 10.1038/sj.npp.1300778. [DOI] [PubMed] [Google Scholar]
- 42.Becker JB. Estrogen rapidly potentiates amphetamine-induced striatal dopamine release and rotational behavior during microdialysis. Neurosci Lett. 1990;118:169–171. doi: 10.1016/0304-3940(90)90618-j. [DOI] [PubMed] [Google Scholar]
- 43.Becker JB. Gender differences in dopaminergic function in striatum and nucleus accumbens. Pharmacol Biochem Behav. 1999;64:803–812. doi: 10.1016/s0091-3057(99)00168-9. [DOI] [PubMed] [Google Scholar]
- 44.Xiao L, Becker JB. Quantitative microdialysis determination of extracellular striatal dopamine concentration in male and female rats: effects of estrous cycle and gonadectomy. Neurosci Lett. 1994;180:155–158. doi: 10.1016/0304-3940(94)90510-x. [DOI] [PubMed] [Google Scholar]
- 45.Taylor JR, Lynch WJ, Sanchez H, Olausson P, Nestler EJ, Bibb JA. Inhibition of Cdk5 in the nucleus accumbens enhances the locomotor-activating and incentive-motivational effects of cocaine. Proc Natl Acad Sci. 2007;104(10):4147–52. doi: 10.1073/pnas.0610288104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Ben-Shahar O, Keeley P, Cook M, Brake W, Joyce M, Nyffeler M, et al. Changes in levels of D1, D2, or NMDA receptors during withdrawal from brief or extended daily access to IV cocaine. Brain Res. 2007;1131:220–228. doi: 10.1016/j.brainres.2006.10.069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Briand LA, Flagel SB, Garcia-Fuster MJ, Watson SJ, Akil H, Sarter M, et al. Persistent alterations in cognitive function and prefrontal dopamine D2 receptors following extended, but not limited, access to self-administered cocaine. Neuropsychopharmacology. 2008;33:2969–2980. doi: 10.1038/npp.2008.18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Mateo Y, Lack CM, Morgan D, Roberts DC, Jones SR. Reduced dopamine terminal function and insensitivity to cocaine following cocaine binge self-administration and deprivation. Neuropsychopharmacology. 2005;30:1455–1463. doi: 10.1038/sj.npp.1300687. [DOI] [PubMed] [Google Scholar]
- 49.Ahmed SH, Lin D, Koob GF, Parsons LH. Escalation of cocaine self-administration does not depend on altered cocaine-induced nucleus accumbens dopamine levels. J Neurochem. 2003;86:102–113. doi: 10.1046/j.1471-4159.2003.01833.x. [DOI] [PubMed] [Google Scholar]
- 50.Ahmed SH, Koob GF. Changes in response to a dopamine receptor antagonist in rats with escalating cocaine intake. Psychopharmacology. 2004;172:450–454. doi: 10.1007/s00213-003-1682-9. [DOI] [PubMed] [Google Scholar]
- 51.Anker JJ, Carroll ME. Females are more vulnerable to drug abuse than males: evidence from preclinical studies and the role of ovarian hormones. Curr Top Behav Neurosci. 2011;8:73–96. doi: 10.1007/7854_2010_93. [DOI] [PubMed] [Google Scholar]
- 52.Becker JB, Hu M. Sex differences in drug abuse. Front Neuroendocrinol. 2008;29:36–47. doi: 10.1016/j.yfrne.2007.07.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Festa ED, Quinones-Jenab V. Gonadal hormones provide the biological basis for sex differences in behavioral responses to cocaine. Horm Behav. 2004;46(5):509–19. doi: 10.1016/j.yhbeh.2004.04.009. [DOI] [PubMed] [Google Scholar]
- 54.Hu M, Crombag HS, Robinson T, Becker JB. Biological basis of sex differences in the propensity to self-administer cocaine. Neuropsychopharmacology. 2004;29:81–5. doi: 10.1038/sj.npp.1300301. [DOI] [PubMed] [Google Scholar]
- 55.Lynch WJ, Carroll ME. Sex differences in the acquisition of intravenously self-administered cocaine and heroin rats. Psychopharmacology. 1999;144:77–82. doi: 10.1007/s002130050979. [DOI] [PubMed] [Google Scholar]
- 56.Becker JB, Hu M. Sex differences in drug abuse. Front Neuroendocrinol. 2008;29:36–47. doi: 10.1016/j.yfrne.2007.07.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Thompson TL, Moss RL. Estrogen Regulation of Dopamine Release in the Nucleus Accumbens: Genomic- and Nongenomic-Mediated Effects. J Neurochem. 1994;62:1750–1756. doi: 10.1046/j.1471-4159.1994.62051750.x. [DOI] [PubMed] [Google Scholar]
- 58.Festa E, Jenab S, Weiner J, Nazarian A, Niyomchai T, Russo SJ, et al. Cocaine-induced sex differences in D1 receptor activation and binding levels after acute cocaine administration. Brain Res Bull. 2006;68:277–284. doi: 10.1016/j.brainresbull.2005.08.023. [DOI] [PubMed] [Google Scholar]
- 59.Lynch WJ, Kiraly DD, Caldarone BJ, Picciotto MR, Taylor JR. Effect of cocaine self-administration on striatal PKA-regulated signaling in male and female rats. Psychopharmacology. 2007;191(2):263–71. doi: 10.1007/s00213-006-0656-0. [DOI] [PubMed] [Google Scholar]
- 60.Auger AP, Meredith JM, Snyder GL, Blaustein JD. Oestradiol increases phosphorylation of a dopamine-and cyclic AMP-regulated phosphoprotein (DARPP-32) in female rat brain. J Neuroend. 2001;13:761–768. doi: 10.1046/j.1365-2826.2001.00700.x. [DOI] [PubMed] [Google Scholar]
- 61.Meredith JM, Moffatt CA, Auger AP, Snyder GL, Greengard P, Blaustein JD. Mating-related stimulation induces phosphorylation of dopamine-and cyclic AMP-regulated phosphoprotein-32 in progestin receptor-containing areas in the female rat brain. J Neurosci. 1998;18:10189–10195. doi: 10.1523/JNEUROSCI.18-23-10189.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Woolverton WL. Effects of a D1 and a D2 dopamine antagonist on the self-administration of cocaine and piribedil by rhesus monkeys. Pharmacol Biochem Behav. 1986;24:531–535. doi: 10.1016/0091-3057(86)90553-8. [DOI] [PubMed] [Google Scholar]
- 63.Bergman J, Kamien JB, Spealman RD. Antagonism of cocaine self-administration by selective dopamine D(1) and D(2) antagonists. Behav Pharmacol. 1990;1:355–363. doi: 10.1097/00008877-199000140-00009. [DOI] [PubMed] [Google Scholar]
- 64.Pickens CL, Airavaara M, Theberge F, Fanous S, Hope BT, Shaham Y. Neurobiology of the incubation of drug craving. Trends Neurosci. 2011;34:411–420. doi: 10.1016/j.tins.2011.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Kalivas PW, McFarland K. Brain circuitry and the reinstatement of cocaine-seeking behavior. Psychopharmacology. 2003;168:44–56. doi: 10.1007/s00213-003-1393-2. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.