

Abstract
Skin cancer is still a major cause of morbidity and mortality worldwide. Skin overexposure to ultraviolet irradiations, chemicals, and several viruses has a capability to cause severe skin-related disorders including immunosuppression and skin cancer. These factors act in sequence at various steps of skin carcinogenesis via initiation, promotion, and/or progression. These days cancer chemoprevention is recognized as the most hopeful and novel approach to prevent, inhibit, or reverse the processes of carcinogenesis by intervention with natural products. Phytochemicals have antioxidant, antimutagenic, anticarcinogenic, and carcinogen detoxification capabilities thereby considered as efficient chemopreventive agents. Considerable efforts have been done to identify the phytochemicals which may possibly act on one or several molecular targets that modulate cellular processes such as inflammation, immunity, cell cycle progression, and apoptosis. Till date several phytochemicals in the light of chemoprevention have been studied by using suitable skin carcinogenic in vitro and in vivo models and proven as beneficial for prevention of skin cancer. This revision presents a comprehensive knowledge and the main molecular mechanisms of actions of various phytochemicals in the chemoprevention of skin cancer.
1. Introduction
Skin cancer is currently the most common type of human cancer worldwide. In the United States, the annual incidence rate of skin cancer is increasing each year, representing a growing public concern [1]. It has also been anticipated from the current records that nearly half of all Americans are susceptible to develop skin cancer at least once up to the age of 65. According to an estimate by the American Cancer Society 76,250 men and women have been diagnosed with skin cancer and 12,190 men and women could have been died (9,180 from melanoma and 3,010 from other nonepithelial skin cancers) in the United States in year 2012. Even if considerable progress has been done in developing effective skin cancer treatment modalities, skin cancer is still a global health problem of concern [2]. The main types of skin cancer are the following. (i) Basal cell carcinoma, the most common easily treated form of skin cancers and accounts for 90% of all cases. Long-term exposure to sunlight is the main cause of it. (ii) Squamous cell carcinoma is the second most common type of skin cancer. It is easily treated when found early, but in a small percentage of cases this cancer has metastasis potential. Both basal cell and squamous cell carcinoma are together known as nonmelanoma type of skin cancer. (iii) Melanoma is responsible for 75% of all skin cancer-related deaths [3] despite the fact that it accounts for less than 5% of all skin cancer cases. Approximately 65–90% of melanomas are caused by exposure to ultraviolet (UV) light [4]. (iv) Kaposi's sarcoma, a more slow-growing form of skin cancer, occurs in elderly men of Italian or Jewish ancestry and is caused by a Herpes family virus. It is an aggressive AIDS-related form affects about one third of patients with AIDS.
2. Cause of Skin Carcinogenesis
Skin, a major environmental interface for the body, is unintentionally or occupationally exposed to a number of harmful stimuli such as UV irradiation, viruses, and several chemical carcinogenic agents (Figures 1 and 2).
Figure 1.
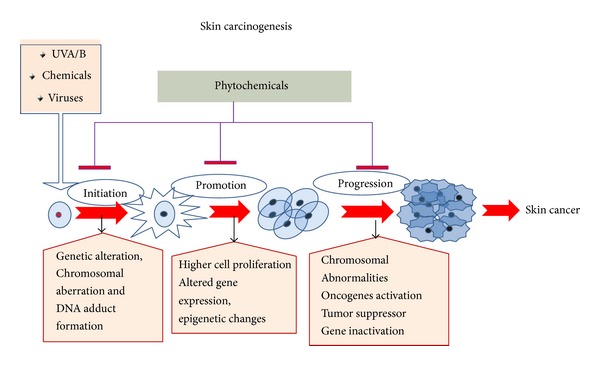
Skin cancer develops in series of events in multiple steps; however in most of studies, skin carcinogenesis is progressed in three key steps, that is, initiation, promotion, and progression, and many phytochemicals could prevent the abrupt changes in each of the steps to reverse the process of developing skin cancer.
Figure 2.
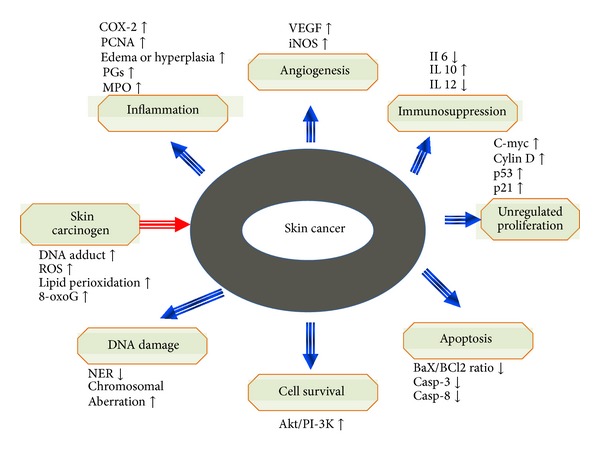
Molecular pathways are altered due to modulation of signaling pathway in skin cancer. Skin carcinogens lead to higher level of DNA adducts formation, ROS, lipid perioxidation, and 8-oxoG. Afterwards, major events like inflammation, angiogenesis, immunosuppression, higher proliferation, lesser apoptosis, enhanced DNA damage, and cell survival take place. Upregulated and downregulated molecules are depicted by arrows represented in upper and lower direction, respectively.
2.1. UV Radiation
Epidemiological data around the world have been evocative of that the exposure of skin to UV radiation is the main ecological carcinogen in the development of both melanoma and nonmelanoma types of skin cancers. UV irradiation stimulates clonal expansion of aberrant skin cells, resulting in skin carcinogenesis via involvement of multiple cellular signaling pathways [5]. The skin exposures to both ultraviolet B (UVB, wavelength 280–320 nm) and ultraviolet A (UVA, wavelength 320–400 nm) radiation have been shown to cause DNA damage and immunosuppression. In comparison to UVA radiation, UVB is more lethal and acts as complete carcinogen. It has been well recognized that UVB induced oxidative stress contributes to several adverse biological effects on the skin [6–8]. UVB is absorbed by proteins or DNA of epidermis, whereas UVA radiation penetrates deeply into the skin and reaches the lower epidermis and dermal fibroblasts. Other chromophores also absorb UV radiation to generate reactive oxygen species (ROS) and cause oxidative damage to DNA such as 8-oxo-7, 8-dihydroguanine (8-oxoG). Several tumor suppressor genes and oncogenes have been reported to be activated by UV (p53, PTCH1, BRM, and Ras) and implicated in photocarcinogenesis [9]. The alteration of the signaling molecules regulating cell proliferation, differentiation, and death is associated with UVB radiation induced skin cancer [10]. UVB radiation is known to modulate the cell cycle regulatory proteins (cyclins, cyclin-dependent kinases), antiapoptotic proteins (Akt, Bcl-2), transcription factors (AP-1, NF-κB, and STAT3), proapoptotic proteins (caspases, PARP), various protein kinases (Akt, Erk, IKK, JNK, PI3K, and p38), inflammatory enzymes (COX-2), and growth factor signaling pathways (EGFR, TNF, IGF).
2.2. Chemicals
Over the last few years occupational skin cancer cases are reported and have mainly been owing to industrial exposure of human being to chemical carcinogens such as polycyclic aromatic hydrocarbons (PAH) and arsenic. PAH from shale oil, tar, pitch, raw paraffin, creosote, asphalt, and chimney soot have been associated with risk to skin cancers. Workers from industries in which PAH are produced (coal, coke and aluminium production plants, steel and iron foundries, and exposure to diesel engine exhaust fumes) are at higher risk of skin cancer [11]. UV radiation is reported to be a cocarcinogen to PAH induced skin cancers [12]. Arsenic in pesticides represents a potential skin cancer risk for agriculture workers. Arsenic mining and smelting industries are also place for its exposure. Being a strong mutagen, it induces large chromosomal mutations and also appears to act as a cocarcinogen with UV. Occupational coexposure to arsenic and sunlight creates an increased risk of NMSC [13].
2.3. Viruses
Viruses are another agent which transforms keratinocytes by activation of cancer-promoting genes. Apart from this, several viral proteins act as oncogenes which lead to the cellular proliferation. Viral skin carcinogenesis is highly concerned with immune deficient host, in such cases lower T cell reactivity and antigen presenting cells in skin assist in viral escape and tumorigenesis [14]. Among major viral agents (i) human papilloma virus (HPV) which is linked to squamous cell carcinoma (SCC) of the skin and its role in nonmelanoma skin cancer of immune competent hosts is not yet proved; (ii) Kaposi's sarcoma associated herpes virus which has wide repertoire of genes that regulate angiogenesis, inflammation, and cell cycle through which it initiates carcinogenesis; and (iii) human T-cell leukemia virus type 1 (HTLV-1) which majorly causes adult T-cell leukemia are common for skin cancer.
3. Animal Model for Skin Carcinogenesis Studies
In the current understanding of skin carcinogenesis, several animal models which are based on two stage carcinogenesis model framework have been developed. Among them in 7,12-dimethylbenz(a)anthracene (DMBA) or chronic UVB/A initiated and 12-O-tetradecanoylphorbol-13-acetate (TPA) promoted rodent models are widely used. The most commonly employed chemical carcinogen for inducing experimental carcinogenesis is DMBA [15]. DMBA induced experimental skin carcinogenesis is preceded by a sequence of hyperplasia, dysplasia, and carcinoma. DMBA mediates skin carcinogenesis through formation of DNA adducts, DNA damage, ROS generation, and chronic inflammation [15]. It is suggested that DMBA mediated changes are similar to those noted in human cancers [16]. DMBA initiated carcinogenesis may so be used as suitable model to study the chemopreventive potential of compounds.
4. Mechanistic Pathways of Skin Carcinogenesis
4.1. Mitogen-Activated Protein Kinases (MAPKs) Signaling Pathway
MAPKs, a family of serine/threonine-specific protein kinases belonging to the CMGC (CDK/MAPK/GSK3/CLK) kinase group [17], play a major role in translating extracellular signals into intracellular pathway through phosphorylation cascade [18]. MAPKs work mainly through the extracellular signal-regulated protein kinases (ERK), the c-Jun N-terminal kinases (JNK), and p38 kinases pathways. These are involved in regulation of cellular metabolism and gene expression and different vital actions including growth and development, disease, apoptosis, and cellular responses to external stimuli. The Raf/MEK/ERK cascade is a key signaling element in development, growth, and inflammation via the transmission of signals from cell surface receptors to the nucleus [19]. The Raf/ERK pathway is the most critical mediator of oncogene Ras-dependent carcinogenesis and UV irradiation is reported to act as inducer of it [20, 21]. Inhibition of MEK/ERK activities inhibited UV induced skin carcinogenesis [22]. In addition, Raf is one of the signaling molecules involved in the UVA induced production of matrix metalloproteinase 1 (MMP-1). MKK4, another important element of stress activated MAPK signaling pathways, phosphorylates and activates both JNK1/2 and p38 but not ERK1/2 in response to cellular stresses and inflammatory cytokines. In addition, the activation of JNK by inflammatory cytokines or by stress is frequently accompanied by the nuclear translocation of NF-κB, and many genes require the concomitant activation of AP-1 and NF-κB [23]. JNK1−/− mice were highly susceptible to DMBA/TPA-induced skin carcinogenesis as indicated by the increased rates of tumor growth kinetic and progression into carcinomas as compared to the wild type counterparts [24]. p38 signaling is critical for solar radiation induced skin carcinogenesis [25]. UVB irradiation results in the activation of this pathway and subsequently AP-1 activation and cyclooxygenase-2 (Cox-2) expression [26]. Hence, targeting of MAPK pathway seems as an attractive approach for the development of skin cancer chemotherapeutic agents.
4.2. Phosphatidylinositol-3-kinase (PI3K)/Akt Pathway
PI3K is a member of lipid kinase family and generates phosphatidylinositol-3,4,5-trisphosphate (PI(3, 4, 5)P3). PI(3, 4, 5)P3 is a second messenger essential for the translocation of Akt to the plasma membrane where it is phosphorylated and activated by phosphoinositide-dependent kinase (PDK) 1 and PDK2. Activated Akt alter the function of several substrates involved in the regulation of cell survival, cell cycle progression, and cellular growth. In recent years, it has been shown that PI3K/Akt signalling pathway components are frequently altered in cancers [27]. UV irradiation triggers the activation of EGFR and subsequently upregulates the phosphorylation of Akt, which is involved in enhanced survival of mutated cells, thereby promoting skin cancer [28]. Further, activated Akt promotes cell survival by inhibiting apoptosis by the inactivation of several proapoptotic factors Bad. The Akt level is reported to be increased during chemically induced skin carcinogenesis in mouse due to increased activity of PI3K [29, 30]. Studies have also shown that Akt regulates the activation of NF-κB and AP-1 [31, 32]. PI3K has been proven to be necessary for Ras-induced malignant transformation; mice with mutations in the PI3K catalytic subunit p110α that block its ability to interact with Ras are highly resistant to endogenous oncogenic H-Ras-induced skin carcinogenesis [33]. Thereby, the targeting of this pathway will provide an effective means of cancer therapy.
4.3. The JAK-STAT Pathway
It is an important oncogenic signaling cascade that consists of the Janus kinase (JAK) family of nonreceptor tyrosine kinases and the signal transducer of activator of transcription (STAT) family of transcription factors. Under physiological conditions, the ligand-dependent activation of the JAK-STAT pathway is transient and securely regulated. The JAK family plays a critical role in cell survival, proliferation, differentiation, and apoptosis. However, in most malignancies, STAT proteins and particularly STAT3 are aberrantly activated (tyrosine phosphorylation). STAT3 regulates the expression of a variety of genes in response to cellular stimuli and thus plays a key role in cell growth and apoptosis. The activation and interaction between STAT3 and NF-κB play vital roles in control of the communication between cancer cells and inflammatory cells. STAT3 and NF-κB are reported as two major factors controlling the ability of preneoplastic as well as malignant cells to resist apoptosis-based tumor-surveillance and regulating tumor angiogenesis and invasiveness. Therefore, the JAK-STAT3 pathway has been suggested as playing a critical role in cell transformation and carcinogenesis. Intervention of this pathway will offer opportunities for the design of new chemopreventive and chemotherapeutic approaches.
4.4. Cyclooxygenase (Cox)
Derived prostaglandins (PGs) exhibit multiple functions in severe and chronic skin inflammation provoked by various physical (UV light, wounding) and chemical (TPA, arachidonic acid) injurious stimuli. COX has two main isoforms as COX-1 and COX-2. COX-1 is constitutively expressed in most tissues, whereas COX-2 is inducible by a variety of tumor promoting agents [34]. It has been observed that Cox-2 is upregulated after UV exposure in skin cells and is involved in the development of skin cancer [35]. Lack of one allele of Cox-2 mice had a 50–65% reduction of tumor multiplicity and a marked decrease in tumor size in a TPA-induced murine tumorigenesis model. Topical administration of indomethacin after each UV exposure was also effective, suggesting that a postexposure approach to skin cancer prevention may be effective [36]. Thus, the interruption of COX-2 is an effective strategy to treat and chemoprevent the skin cancer.
4.5. Hypoxia-Inducible Factor-1 (HIF-1)
Heterodimeric factor consisting of two subunits α and β is a master regulator of the transcriptional response of cells to oxygen deprivation. Since its discovery in the early 1990s, HIF-1 has quickly attracted interest both for its involvement in fundamental biological processes, including tumor metabolism, angiogenesis, metastasis, and inflammation, and for its potential role as therapeutic target [37]. In addition, HIF-1 can induce in an oxygen-independent manner by various cytokines through the PI3-K/Akt signal transduction pathway [38]. Deregulation of HIF-1alpha is associated with UVB-induced hyperplasia of the epidermis [39]. UV radiation induces HIF-1alpha and VEGF expression via the EGFR/PI3K/DEC1 signaling pathway [40]. Thus, developing an agent that can suppress HIF1 expression could be a critical strategy for the chemoprevention of skin cancer.
Many angiogenic factors, including vascular endothelial growth factor (VEGF) and matrixmetalloproteinases (MMPs), are also upregulated in skin cancer.
5. Skin Cancer Chemoprevention by Using Phytochemicals: Mechanistic Studies
Cancer chemoprevention by using phytochemicals to prevent or suppress the process of carcinogenesis is still an area of active investigation. Epidemiological studies have provided persuasive evidence that phytochemicals can prevent this process. Laboratory researches have further established the effectiveness of a number of bioactive natural components that have the capacity to check cancer and other chronic diseases. Skin carcinogenesis, a multistep process, allows for the possible curative intervention; thus, cancer chemoprevention is a very promising approach to successfully achieve this goal. Lots of compounds belonging to diverse chemical classes have been identified and proved as potential skin cancer chemopreventive agents. Fruits, vegetables, seeds, flowers, leaves, and bark represent huge reservoirs of phytochemicals such as polyphenols, flavonoids, isoflavonoids, proanthocyanidins, phytoalexins, anthocyanidins, and carotenoids. Many of them have already been studied extensively for their potential anticancer or chemopreventive efficacy and some of them are at clinics now. According to estimates, on average 0.2–1 g/d of these agents are consumed as part of a regular diet [41]. Thus, the better understanding of the knowledge of underneath molecular mechanisms of cancer chemoprevention not only is important for the safe and effective use of these compounds in cancer patients but also allows for further development of novel and better therapy for cancer patients. In this review we summarized the update knowledge of photochemical reported to have skin cancer chemopreventive prospective (Figure 3 and Table 1).
Figure 3.
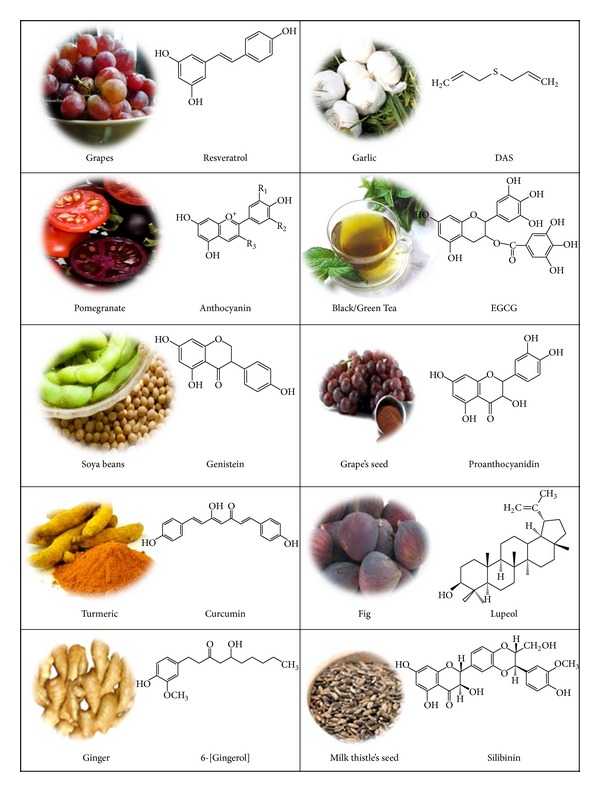
Phytochemicals and their major source associated with chemoprevention of skin cancer as discussed in text.
Table 1.
| Number | Name of phytochemicals | Rich source | Concerned pathways in skin cancer | References |
|---|---|---|---|---|
| 1 | Myricetin | Walnuts (Juglans regia) | Inhibits Akt activity to induce apoptosis (HaCaT cells) and also inhibits Fyn kinase activity | Kim et al. 2010 [58]; Jung et al. 2008 [59] |
| 2 | Caffeic acid | Coffee (Coffea canephora) | Inhibits Fyn kinase activity | Kang et al. 2009 [60] |
| 3 | Alpha-santalol | Sandalwood oil | Activates proapoptotic caspases and p53 and also cleave poly (ADP-ribose) polymerase through activating upstream caspase-8 and caspase-9. | Arasada et al. 2008 [61] |
| 4 | Nicotinamide | Peas, Asparagus, Mushroom, Squash | Repair UV induced DNA damage | Surjana et al. 2013 [62] |
| 5 | Norathyriol | Mango, Hypericum elegans,Tripterospermum lanceolatum, and so forth | Inhibit AKT activation, epidermal growth factor (EGF), and ERk1/2 to attenuate UVB-induced phosphorylation in MAPK signaling | Li et al. 2012 [63] |
| 6 | Retinoids (Acitretin and isotretinoin) | Vitamin A rich food source like Carrot, Spinach, pumpkin, and so forth | Prevention of nonmelanoma skin cancer isotretinoin is preferred in xeroderma pigmentosum and nevoid basal cell carcinoma syndrome, whereas acitretin is more used in transplant recipients, psoriasis, and severe sun damage | Bettoli et al. 2013 [64] |
| 7 | Shikonin | Lithospermum erythrorhizon | Reverse AMPK activity in TPA induced suppression | Li et al., 2012 [65] |
| 8 | Ferulic acid (FA) | Rich source in Phaseolus vulgaris, Rice and Maize Bran | Inhibit MMP-2 and MMP-9 protein expression | Staniforth et al. 2012 [66] |
| 9 | Honokiol | Magnolia plant | Inhibit UVB-induced expression of cyclooxygenase-2, prostaglandin E(2), PCNA, and proinflammatory cytokines (TNF-α, IL-1β and IL-6) | Vaid et al. 2010 [67] |
| 10 | Pycnogenol B7 | French maritime pine, Pinus pinaster | Offered protection against sun UV induced acute inflammation, immunosuppression, and carcinogenesis | Sime and Reeve, 2004 [68] |
| 11 | Guggulsterone | Commiphora mukul | attenuates Cox-2 and i NOS protein expression | Sarfaraz et al., 2008 [69] |
| 12 | Humulone | Humulus lupulus | Inhibit TPA-induced epidermal COX-2 expression by blocking upstream kinases IKK and JNK and activates NF-kappaB and AP-1. | Lee et al., 2007 [70] |
| 13 | Apigenin | Parsley and Onions | Inhibits UVB-induced COX-2 expression by targeting src Kinase | Byun et al., 2012 [71] |
| 14 | 5-Hydroxy-3,6,7,8,3′,4′-hexamethoxyflavone (5-OH-HxMF), | Fruits of citrus genus specially Peels of sweet orange | Inhibitory effects on TPA-induced expression of iNOS and COX-2 in mouse skin and activates NF-κB and STAT3 | Lai et al., 2007 [72] |
| 15 | 3,3′-Diindolylmethane (DIM) | Cruciferous vegetables like broccoli, cabbage, and so forth | Inhibited the TPA-induced increases in COX-2, iNOS, chemokine (C-X-C motif) ligand (CXCL) 5, IL-6, NF-κB, IKK, and ERK-1/2 | Kim et al., 2010 [73] |
| 16 | Xanthorrhizol | Curcuma xanthorrhiza | Reduces ODC, iNOS, and COX-2 level, regulated by the NF-κB, MAPK/Akt | Chung et al., 2007 [74] |
| 17 | Oleandrin | Nerium oleander | Reverts PI3K, Akt phosphorylation, and activation of NF-κB | Afaq et al., 2004 [75] |
| 18 | Delphinidin | genus Delphinium | Decreases cell viability and PCNA expression, peroxidation, and formation of 8-OHdG in HaCaT cells and SKH-1 animals | Afaq et al., 2007 [76] |
5.1. Proanthocyanidins
Grapes (Vitis vinifera) are one of the most widely consumed fruits in the world and its seeds are the rich source of proanthocyanidins (oligomeric flavonoids) which are well known to exert anti-inflammatory, antioxidant, antiarthritic, and antiallergic activities, prevent skin aging, and inhibit UV radiation-induced peroxidation activity. Other rich sources of proanthocyanidins are red wine, cocoa, apples, peanuts, almonds, cranberries, blueberries, and bark of the maritime pine. Grape seed proanthocyanidins (GSPs) are promising bioactive phytomolecules that have shown antiskin carcinogenic effects and reveal no apparent toxicity in vivo [42–44]. The results of acute and subchronic toxicity testing in rats, in vitro genotoxicity testing in mammalian cell, and in vivo mouse micronucleus test reveal a low toxicity and no genotoxic potential of proanthocyanidins-rich extract from grape seeds [45]. The bioavailability of proanthocyanidins is largely influenced by their degree of polymerization. The absorption rate of proanthocyanidin dimers is 5–10% of that of (−)-epicatechin [46]. Trimers and tetramers had lower absorption rates than dimers. Absorbed intact dimers, trimers, and tetramers undergo limited phase II metabolism in the intestine and liver in rats [46]. Employing HPLC nine individual polyphenols is identified in GSPs as catechin, epicatechin, procyanidins B1–B5 and C1, and procyanidin B5-3′-gallate which are potent antioxidant in terms of inhibition of epidermal lipid peroxidation [47]. Intake of GSPs prevents skin cancer through their anti-inflammatory, antioxidative, and DNA repair mechanisms in both UV radiation-induced and DMBA-initiated/TPA-promoted mouse skin tumors. Mittal et al. [42] reported that dietary intake of GSPs (0.2 and 0.5%, w/w) resulted in prevention against UVB-induced complete initiation and promotion stages of photocarcinogenesis in terms of tumor incidence (20–95%), multiplicity (46–95%), and size (29–94%). Treatment of GSP significantly inhibited UVB or Fe3+ induced lipid peroxidation by 57–66% (P < 0.01) and 41–77% (P < 0.05–0.001), respectively, thus suggesting the strong antioxidant mechanism of photoprotection offered by GSPs [42]. According to Sharma et al. [48, 49] same doses of GSPs have the ability to protect the skin from the adverse effects of UVB radiation via modulation of the MAPK and NF-κB pathways. GSPs inhibited UVB-induced infiltration of proinflammatory leukocytes and the levels of myeloperoxidase (MPO), Cox-2, PGE2, Cyclin D1, and PCNA in the skin and skin tumors compared to non-GSPs-treated UVB irradiated counterpart [50]. GSPs reduced the UVB-induced increase in immunosuppressive cytokine interleukin (IL-) 10 in skin and draining lymph nodes compared with mice that did not receive GSPs. In contrast, GSPs enhanced the production of immune stimulatory cytokine IL-12 in the draining lymph nodes. GSPs fed mice exhibit a reduction in UV-induced suppression of allergic contact hypersensitivity, a prototypic T-cell-mediated response [51]. Results suggest that GSPs prevent UVB-induced immunosuppression through DNA repair dependent functional activation of dendritic cells and reexpression of tumor suppressor genes RASSF1A, p16INK4a, and Cip1/p21. Skin painted with GSPs before UV radiation showed fewer sunburn cells and mutant p53-positive epidermal cells and more Langerhans cells compared with skin treated with UV radiation only [52]. Dose dependent antitumor-promoting effects of GSPs were evident in terms of a reduction in tumor incidence (35 and 60% inhibition), multiplicity (61 and 83% inhibition), and volume (67 and 87% inhibition) at both 0.5 and 1.5 mg GSP, respectively, in DMBA/TPA treated SENCAR mouse skin model [48]. Pretreatment of CD-1 mouse skin with GSP (5–30 mg doses) resulted in a dose-dependent reduction in TPA-induced ODC and MPO activity [53, 54].
Mantena and Katiyar [54] attempted to define the photoprotective mechanism of action of GSPs. Researchers observed that in vitro treatment of NHEK with GSPs resulted in the prevention of UVB-induced depletion of antioxidant defense enzymes (GPx, catalase, SOD, and GSH) and H2O2 production. Further, GSPs inhibit H2O2-induced phosphorylation of ERK1/2, JNK, and p38 proteins. Meeran and Katiyar [55] reported that GSPs inhibited skin cancer cell (A431 cells) proliferation which was mediated through the inhibition of cyclin-dependent kinases (Cdk) Cdk2, Cdk4, and Cdk6 and cyclin D1, D2, and E, increase in cyclin-dependent kinase inhibitors (Cdki), Cip1/p21 and Kip1/p27, and enhance binding of Cdki-Cdk. GSPs have the ability to inhibit highly metastasis-specific human melanoma cells invasion/migration by targeting the endogenous Cox-2 expression and PGE2 production and reversing the process of epithelial-to-mesenchymal transition [56]. The precise epigenetic molecular mechanisms of anticancer potential of GSPs have been explored by Vaid et al. [57]. Treatment of skin cancer cells with GSPs decreased the levels of global DNA methylation, 5-methylcytosine, DNA methyltransferase (DNMT) activity, and mRNA and protein levels of DNMT1, DNMT3a, and DNMT3b in treated cells.
5.2. Tea Polyphenols
Tea (Camellia sinensis; Theaceae) has been consumed as a popular beverage worldwide and skin photoprotection by green tea polyphenols (GTPs) has been widely investigated. Besides health beneficial effects toxicity of tea polyphenols is attributed to their role as prooxidants in the presence of redox-active chemicals and lead to the formation of reactive oxygen species which can cause damage to DNA, lipids and other biological molecules [77, 78]. Oral feeding of a GTP or water extract of green tea affords protection against UVB radiation induced inflammatory responses and carcinogenesis in animals [79]. GTP (0.2% w/v) as the sole source of drinking water for 30 days to SKH-1 hairless mice followed by irradiation with UVB (900 mJ/cm2) offered a significant protection against UVB radiation induced cutaneous edema, epidermal ODC and Cox-2 activities, and depletion of the antioxidant-defense system in epidermis [80]. Further elevated levels of nucleotide excision repair (NER) genes show a novel mechanism by which drinking GTPs prevents UV-induced immunosuppression [81]. Prevention of photocarcinogenesis by GTPs is mediated through IL-12-dependent DNA repair and a subsequent reduction in skin inflammation [82]. GTPs (0.2%, w/v) in drinking water significantly reduced UVB-induced tumor development, markers of inflammation (Cox-2, PGE2, PCNA, and cyclin D1) and proinflammatory cytokines (TNF-α, IL-6, and IL-1β) in wild-type mice, when compared with IL-12-knockout mice. Topically repeated doses of GTP (5 mg/mouse) resulted in significant decrease in UVB (180 mJ/cm2) induced bi-fold-skin thickness, skin edema and infiltration of leukocytes, and modulations in MAPK and NF-κB signaling pathways [83]. Application of epigallocatechin gallate (EGCG) protects against radiation induced immunosuppression by tolerance induction, blocking the migration of antigen-presenting cells from the skin to draining lymph nodes (DLN) and markedly inducing IL-12 dependent DNA repair mechanism [81, 84–86]. In addition EGCG decreased the number of UVB-induced increases in H2O2-producing cells and inducible nitric oxide synthase (iNOS-) expressing cells and the production of H2O2 and NO in both exposed part of epidermis and dermis [87]. Findings of a study done by Vayalil et al. [88] suggest the long-term photopreventive effects of EGCG as a cream based formulation of GTP which was for the first time tested in this study to explore the possibility of its use for the humans. Topical application of EGCG (1 mg/cm2) in hydrophilic ointment before single (180 mJ/cm2) or multiple (180 mJ/cm2, daily for 10 days) doses of UVB exposure resulted in prevention of UVB-induced depletion of antioxidant enzymes levels. Further, treatment of EGCG to mouse skin resulted in marked inhibition of a single UVB induced phosphorylation of ERK1/2 (16–95%), JNK (46–100%), and p38 (100%) proteins in a time-dependent manner. Photoprotective efficacy of GTP given in drinking water (0.2%, w/v) was also compared with that of topical treatment of EGCG and GTP by Vayalil et al. [88]. It was noted that lesser photoprotective efficacy of GTP in drinking water in comparison with topical application may be due to its less bioavailability in skin target cells. Pretreatment with EGCG was also found to restore the UV-induced decrease in GSH level and afforded protection to the antioxidant enzyme GPx in human skin [87].
Topical application of black tea polyphenols (BTP) (6 mg/0.2 mL acetone/animal) 30 min prior to TPA application on to the mouse skin resulted in inhibition against TPA-induced epidermal edema, hyperplasia, leukocytes infiltration, and induction of epidermal ODC and proinflammatory cytokine IL-1alpha mRNA expression and Cox2 activities [89]. Thearubigins or polymeric black tea polyphenols (PBPs) have been shown to possess antitumor-promoting effects in two-stage skin carcinogenesis. Pretreatment with PBP fractions exhibits antitumor promoting effects as evidenced by differentially altered latency, multiplicity, and incidence of skin papillomas [90]. PBPs pretreatment decreased TPA-induced translocation of PKC isozymes (α, β, η, γ, and ε) from cytosol to membrane and PKC phosphorylation [88]. These antipromoting effects of PBPs are due to modulation of TPA-induced PI3K-mediated signal transduction [91]. PBPs decreased TPA-induced cell proliferation by decreasing the activation of signaling kinases (c-Jun, ERK, p38, and Akt), transcription factors (AP-1 and NF-κB), and inflammatory protein (Cox2) [90].
In vitro doses of EGCG to NHEK cells before UVB exposure inhibits UVB-induced H2O2 production and activation of MAPK [92] and NF-κB pathways [90]. EGCG treatment resulted in a dose-dependent inhibition of cell growth and induction of apoptosis mediated through NF-κB inhibition in A431 cells but not in normal cells [93, 94]. Recent findings suggest that GTP reduce skin cancer cells survival by influencing polycomb group proteins (PcG-) mediated epigenetic regulatory mechanisms [95, 96]. EGCG and DZNep, independently and in combination, reduce the level of PcG proteins, namely, Ezh2, eed, Suz12, Mel18, and Bmi-1 [95]. This change in PcG protein expression was associated with reduced expression of cyclin-dependent kinases (cdk) (1, 2, and 4), cyclin (D1, E, A, and B1), and increased expression of cell-cycle inhibitory proteins (p21 and p27) [93]. Apoptosis was evidenced by increased cleavage of caspases (9, 8, and 3) and increased cleavage of PARP [95].
A population-based, case-control study conducted on 450 individuals showed that tea concentration specifically black tea, brewing time, and beverage temperature has major influences on the potential protective effects of tea in relation to skin squamous cell carcinoma [97]. According to Bickers and Athar, topical application of tea extracts to human volunteers protects against UVB-induced erythema [97]. These studies indicate that tea extracts are effective in reducing UVB- and PUVA-mediated DNA damage and expression of early response genes and early inflammatory changes in human skin [98]. Katiyar et al. in year 2000 conducted a human study and reported that topical treatment with GTP (~1 mg/cm2 of skin area) 20 min before human buttock skin (sun-protected site) exposure to UVB inhibited CPD formation in epidermis by 81, 70, 60, and 60% at 0.5, 1.0, 2.0, and 4.0 minimal erythema dose of UV exposure, respectively [99]. Further, treatment of human skin with varying doses of GTP (1–4 mg/2.5 cm2 of skin area) before a single dose of UVB exposure (4.0 minimal erythema dose) decreased dose dependently the formation of UVB-induced CPDs in both epidermis and dermis.
5.3. Pomegranate Fruit Extract
Pomegranate fruit (Punica granatum L.) possesses extraordinary medicinal properties and has been named as the “nature's power fruit” since ancient time. Traditionally pomegranate including its roots, tree bark, fruit juice, leaves, and flowers has been used to treat several ailments in human. P. granatum contains anthocyanins (delphinidin, cyaniding, and pelargonidin) and hydrolyzable tannins (punicalin, pedunculagin, punicalagin, and gallagic and ellagic acid esters of glucose), reported to have strong antioxidant and anti-inflammatory properties [100]. Pomegranate fruit extract (PFE) possesses antitumor promoting effects in a mouse model of skin carcinogenesis induced by chemicals as well as UV radiations [101, 102]. Application of PFE (2 mg/mouse) prior to TPA on CD-1 mouse skin afforded significant inhibition in TPA-induced increase skin edema and hyperplasia, epidermal ODC activity, and protein expression of ODC and Cox-2 [103]. Further, PFE resulted in inhibition of TPA-induced activation of ERK1/2, p38, JNK1/2, NF-κB, and IKKα proteins. The effects of skin application of pomegranate seed oil and PFE on two stage skin carcinogenesis model (DMBA-initiated and TPA-promoted) were also evaluated by the same researchers [103, 104]. Pomegranate seed oil (5%) and PFE application prior to TPA on mouse skin resulted in delay in latency period from 9 to 14 weeks and afforded considerable protection in terms of tumor incidence and tumor multiplicity [103, 104]. Feeding of PFE (0.2% w/v) for 14 days inhibited UVB-induced skin edema, hyperplasia, infiltration of leukocytes, lipid peroxidation, H2O2, ODC activity, and expression of ODC, Cox-2, PCNA iNOS, cyclin D1 and MMP-2, MMP-3 and MMP-9 proteins in SKH-1 mice [105, 106]. PFE supplemented diet also enhanced the repair of UVB-mediated formation of both CPDs and 8-oxodG [105] and these events were associated with PFE induced inhibition of MAPK and NF-κB pathways [105, 106].
In vitro treatment of PFE (10–40 μg/mL) also protects against the adverse effects of UVB (40 mJ/cm2) radiation by inhibiting UVB-induced modulations of NF-κB and MAPK pathways and provides a molecular basis for the photochemopreventive effects of PFE [107]. Treatment of NHEK cells with PFE (60–100 μg/mL for 24 h) prior to UVA exposure resulted in a dose-dependent inhibition of UVA-mediated phosphorylation of STAT3 at Tyr705, AKT at Ser473, ERK1/2 and mTOR at Thr2448, and p70S6K at Thr421/Ser424 [108]. Further, PFE pretreatment of NHEK was found to increase the G1 phase cell-cycle arrest and the expression of Bax and Bad with downregulation of Bcl-xL [108]. Pretreatment of PFA (10–40 μg/mL) protects HaCaT cells against UVB (15–30 mJ/cm2-) induced oxidative stress and markers of photoaging and could be a useful supplement in skin care products [109]. In a study, Pacheco-Palencia et al. [110] investigated potential protective effects of a PFE against both UVA and UVB-induced damage in human skin fibroblast (SKU-1064) cells. PFE (5 to 60 mg/L) was effective in protecting UV induced cell death and likely related to a reduced activation of NF-κB, a downregulation of caspase-3, and an increased G0/G1 phase. Authors further suggested that higher polyphenolic concentration (500–10000 mg/L) is needed to achieve a noteworthy reduction in UV-induced ROS levels.
Afaq and colleagues [111] determined the effects of pomegranate-derived products, POMx juice, POMx extract, and pomegranate oil (POMo), against UVB-mediated damage using reconstituted human skin (EpiDerm(TM) FT-200). Pretreatment of Epiderm resulted in inhibition of UVB-induced CPD, 8-OHdG, protein oxidation, and expression of collagenase (MMP-1), gelatinase (MMP-2, MMP-9), stromelysin (MMP-3), marilysin (MMP-7), elastase (MMP-12), tropoelastin, c-Fos, c-Jun, and PCNA. Collectively, these results suggest that all three pomegranate-derived products may be useful against UVB-induced damage to human skin.
A double-blind, placebocontrolled trial to clinically evaluate the protective and ameliorative effects of ellagic acid-rich pomegranate extract on pigmentation in the skin after UV irradiation, using female subjects of age 20–40 s, was performed by Kasai et al. [112]. Ellagic acid-rich pomegranate extract ingested orally by the subjects has an inhibitory effect on a slight pigmentation in the human skin caused by UV irradiation, suggesting the relevance of laboratory data in human too.
5.4. Resveratrol
Resveratrol, a trans-3,5,4′-trihydroxy-trans-stilbene, is a phytoalexin, produced by many plants and the skin of red grapes exhibits versatile biological effects. Besides its protection of the cardiovascular system, it can affect the essential stages of carcinogenesis, involving tumor initiation, promotion, and progression. Topical doses of resveratrol either pre- or post- UVB irradiation has been tested for its efficacy against the development of skin cancer. Topical doses of resveratrol to SKH-1 hairless mice significantly inhibited single or multiple doses of UVB (180 mJ/cm2-) mediated phototoxicity as evidenced by reversal of bi-fold skin thickness, skin edema, and hyperplasia [113]. Similarly, in mouse skin DMBA-TPA model, treatment with resveratrol doses showed up to a 98% reduction in skin tumors [114] and 60% reduction in papillomas [115]. These effects were related to its cytotoxic and free radical scavenging activities [116]. According to Fu et al, [117] diet containing resveratrol shown to inhibit DMBA/croton oil-induced skin papillomas correlated with prolonging tumor latency period and inhibiting croton oil-induced epidermal ODC activities. Resveratrol doses in SKH-1 mice skin decreased the expression of cyclins D1 and D2, Cdk 2, 4, and 6, and PCNA and increased expression of p21WAF1/CIP [118]. Further, activation of p53 and Bax along with reduced expression of antiapoptotic proteins (survivin and Bcl-2) and markers of tumor promotion (Cox-2 and ODC) was observed [119, 120]. Kundu et al. [121] findings suggest that resveratrol targets IKK in blocking TPA-induced NF-κB activation and Cox-2 expression in mouse skin. In a study by Roy et al. [122] chemopreventive properties of resveratrol in DMBA induced mouse skin tumors were also reflected by delay in onset of tumorigenesis, reduced cumulative number of tumors, and reduction in tumor volume. DMBA suppressed (p53, Bax, and Apaf-1) and increased (Bcl-2 and survivin) expression of proteins were modulated by resveratrol treatment. Furthermore, resveratrol supplementation resulted in regulation of PI3K/AKT pathway [122]. Results of a study by Yusuf et al. [123] indicate that toll-like receptors (TLR4) are an important mediator of resveratrol induced chemoprevention in DMBA skin tumorigenesis. Resveratrol combinations with ellagic acid, grape seed extract, and other phytochemicals are very potent inhibitors of DMBA induced skin tumorgenesis [124].
In the normal skin keratinocytes, resveratrol doses (5, 10, and 25 μM) blocked UVB-mediated (40 mJ/cm2) activation of NF-κB, phosphorylation and degradation of IκBα, and activation of IKKα [125]. In vitro data demonstrate that resveratrol induces G1-phase cell-cycle arrest accompanied by p21WAF1/CIP1 induction and inhibition of MEK1, ERK1/and AP-1 signalling in A431 cells [126–128]. Resveratrol doses arbitrated TGF-β2 downregulation; this event appears to occur through the inhibition of both TGF-β2/Smad dependent and independent pathways and thus suppressed the invasiveness of A431 cells [129]. Through the establishment of A431 cells xenografts in nude mice, Hao et al. [130] noted that the anticancer mechanism of resveratrol was through inducing apoptosis as it altered p53 and survivin expression. In a study, Bastianetto et al. [131] aimed to investigate the possible existence of specific binding sites in skin that mediate the protective action of resveratrol. Using human skin tissue, researcher reported here the presence of specific [(3)H]-resveratrol binding sites (K(D) = 180 nM) that are mainly located in the epidermis. Cottart et al. reviewed the bioavailability and toxicity of resveratrol; it seem to be well tolerated and without any marked toxicity [132]. These data are important in the context of human effectiveness studies, and authors provide further support for the use of resveratrol as a pharmacological drug in human medicine.
Pterostilbene (trans-3,5-dimethoxy-4-hydroxystilbene), a natural analogue of resveratrol, found in blueberries and grapes is a powerful natural antioxidant. Tsai et al. [133] for the first time suggested pterostilbene as a novel functional agent capable of preventing inflammation-associated skin tumorigenesis which functions by downregulating inflammatory iNOS and Cox-2 gene. Pterostilbene significantly inhibited DMBA/TPA-induced skin tumor formation via suppressing activation of NF-κB, AP-1, ERK1/2, JNK1/2, and PI3K/Akt.
5.5. Silymarin
Since long, silymarin and its major constituent silibinin, isolated from the medicinal plant Silybum marianum (commonly called as milk thistle), have been used for the treatment of hepatic ailments. Recently, these orally active flavonoids have also been interrogated for their significant anticancer effects in a variety of in vitro and in vivo systems of carcinomas including skin. Notably, it is well endured at doses up to 1% w/w or 750 mg/kg body wt in diet and showed no adverse effects to mice [134]. Topical treatment of silymarin inhibited DMBA-initiated and several tumor promoters like TPA, mezerein, benzoyal peroxide, and okadaic acid induced mouse skin carcinogenesis [135]. Mechanism of such effects involves inhibition of promoter-induced edema, hyperplasia, cell proliferation, and oxidative stress [135]. Application of silymarin (3, 6, and 12 mg/animal) prior to TPA resulted in a great protection against tumor promotion in DMBA initiated mouse skin and was apparent in terms of reduction in tumor incidence, multiplicity, and volume [135]. Singh et al. [136] demonstrated the mechanistic rationale of silymarin against skin tumors in in vivo study. In this study DMBA-TPA established that skin papilloma/tumor-bearing SENCAR mice were fed with 0.5% silymarin and both tumor growth and regression were monitored during entire feeding schedule. Silymarin feeding significantly inhibited tumor growth, decreased proliferation index, increased apoptotic index, and decreased phospho-ERK1/2. In UVB-induced carcinogenesis the effect of silymarin was also profound in terms of tumor incidence, multiplicity, and tumor volume [137]. Results from another short term experiments suggested that silymarin application resulted in inhibition of UVB-caused sunburn and apoptotic cell formation, skin edema, depletion of catalase activity, and induction of Cox and ODC activities and ODC mRNA expression [137]. Preventive efficacy of silibinin against UVB induced photocarcinogenesis involves the inhibition of DNA synthesis, cell proliferation, and cell cycle progression and an induction of apoptosis [138, 139]. Dietary feeding of silibinin to mice before UVB irradiation affords strong protection against UV-induced damage in epidermis via decrease in thymine dimer positive cells and increase in p53-p21/Cip1 proteins expression [139–141]. Silibinin also showed a strong phosphorylation of ERK1/2, JNK1/2, and p38 MAPK but inhibited Akt [140]. Topical treatment of silymarin (1 mg/cm2) inhibited UV-induced (90 mJ/cm2) oxidative stress through inhibition of infiltrating CD11b+ cells (showing higher production of ROS in both epidermis and dermis than CD11b-cells) in C3H/HeN mice [142]. The inhibitory effect of silymarin, in spite of whether it is given before or after UV irradiation, was of similar degree [141]. Silibinin treatment (50 mg/kg/day for 4 days) offered UVB induced improper autophagy intervention as evidenced by reversed dermal and epidermal autophagy levels [142, 143].
Anticancer and cancer preventive efficacy of these compounds are also apparent by in vitro studies. Treatment of skin cancer A431 cells by silibinin resulted in cell growth inhibition and death, which was found to be coupled with a decrease in MAPK/ERK1/2 levels and an upregulation of SAPK/JNK1/2 and p38 MAPK activation [136]. Studies suggest that both silymarin and silibinin are equally beneficial in the removal of UVB and UVA damaged skin cells. Pretreatment inhibited UV-induced apoptosis in HaCaT cells [144]. Following the doses of silibinin (500 μM) the expression of Fas-associating protein with death domain was totally abolished and was associated with inhibition of cleavage of procaspase-8, decreased release of cytochrome c, and reduced expression of inhibitor of caspase-activated DNase and PARP. In a study by Roy et al. [145] silibinin pretreatment accelerated the repair of CPD induced by moderate dose of UVB (50 mJ/cm2) and p53-mediated upregulation of GADD45α is suggested as key mechanism. Treatment of UVA irradiated HaCaT (20 J/cm2) with silymarin (0.7–34 mg/L, 4 h) resulted in concentration-dependent diminution of UVA-caused oxidative stress [146]. Silymarin application extensively reduced formation of DNA single strand breaks, ROS production, lipid peroxidation, GSH depletion, and caspase-3 activity in irradiated cells. Silibinin (75 μm 2 h before UVA exposure) enhances ER stress-mediated apoptosis in HaCaT cells by increasing the expression of CHOP protein [147].
5.6. Lupeol
Lupeol, a phytosterol and triterpene, is widely found in edible fruits and vegetables. Extensive research over the last few years has revealed several noticeable pharmacological activities of lupeol. Studies suggest that lupeol has a potential to act as an anti-inflammatory, antimicrobial, antiprotozoal, antiproliferative, anti-invasive, antiangiogenic, and cholesterol lowering agent. Effects of lupeol on TPA-induced markers of skin tumor promotion were evaluated [148]. Topical application of lupeol (1-2 mg/mouse) prior to TPA onto the skin of CD-1 mice afforded significant inhibition against TPA-mediated increase in skin edema and hyperplasia, epidermal ODC activity, and protein expression of ODC, Cox-2, and nitric oxide synthase (NOSs). The animals pretreated with lupeol showed significantly reduced tumor incidence, tumor burden, and delay in the latency period for tumor appearance. Further, lupeol doses resulted in the inhibition of TPA-induced activation of PI3K, phosphorylation of Akt, activation of NF-κB and IKKα, and degradation and phosphorylation of IκBα [149]. Both pre- and posttreatment of lupeol (200 μg/mouse) showed significant (P < 0.001) preventive effects in DMBA induced DNA strand breaks in dose and time dependent manner [148]. Nigam et al. [150] demonstrated the effects of lupeol on DMBA-induced alterations in mouse skin. Cell-cycle analysis showed that lupeol-induced G2/M-phase arrest mediated through inhibition of the cyclin-B-regulated signaling pathway involving p53, p21/WAF1, cdc25C, cdc2, and cyclin-B gene. Lupeol induced apoptosis was associated with upregulation of bax and caspase-3 genes and downregulation of bcl-2 and survivin genes.
In vitro apoptosis inducing effects of lupeol were studied in A431 skin carcinoma cells by Prasad et al. [151]. Lupeol-induced apoptosis was associated with caspase dependent mitochondrial cell death pathway through activation of Bax, caspases, and Apaf1, decrease in Bcl-2, and subsequent cleavage of PARP.
5.7. Curcumin
Curcurmin (diferuloylmethane) is a yellow odorless pigment isolated from the rhizome of turmeric (Curcuma longa L.), reported to have strong anti-inflammatory, anticancer, and antioxidant properties. Anticarcinogenic and chemopreventive effects of curcurmin are attributed to its effect on several apoptotic molecules, transcription factors, cellular signaling pathways, and so forth. It has been found that topical application of curcurmin in TPA-pretreated skin of CD-1 mice significantly inhibited UVA induced ODC activity and ODC gene expression [152]. Further, the inhibitory effects of curcurmin were credited to its capacity to scavenge UVA induced ROS by several researchers [152, 153]. Oral and tropically doses of curcumin were equally potent to inhibit UV induced skin tumor in SKH-1 mice [154].
In vitro doses of curcumin have been reported to prevent UV induced apoptotic changes in human skin cancer (A431) cells, accompanied with release of cytochrome c and activation caspase-3. Moreover, UV induced ROS generation was also abolished by curcurmin [155]. In HaCaT cells curcurmin and UVR synergistically induced apoptotic cell death through activation of caspases 8, 3, and 9 and release of cytochrome c. Treatment with curcumin strongly inhibited COX-2 mRNA and protein expression in UVB irradiated HaCaT cell. Curcurmin inhibited UVB-induced AP-1 transcripcional activation and p38/JNK signaling pathway [156].
Phase I clinical trial of curcumin conducted by Chen et al. demonstrated that curcumin is not toxic to humans up to 8,000 mg/day when taken by mouth for 3 months and has potential effect in the cancer chemoprevention [157].
5.8. Ginger
Ginger (Zingiber officinale Roscoe) is one of the most heavily consumed dietary spices in the world. [6]-gingerol (oleoresin from the root of ginger), a major pharmacologically active component of ginger, has been shown to inhibit several cancers including skin cancer. This compound has antibacterial, anti-inflammatory, analgesic, and antitumor activity. It has been found to possess potent antioxidant activity. A study by Kim et al. [158] suggested that [6]-gingerol could be an effective therapeutic agent providing protection against UVB-induced skin disorders. Topical doses of gingerol prior to UVB irradiation in SKH-1 mice inhibited the induction of COX-2 mRNA and protein, as well as NF-κB translocation.
In JB6 cells [6]-gingerol block EGF-induced cell transformation and inhibited EGF-induced AP-1 DNA binding activity [159]. In vitro, pretreatment with gingerol reduced UVB-induced intracellular ROS levels, activation of caspases (3, 8, -9), and Fas expression. It also reduced UVR-induced expression and transactivation of COX-2 [158]. Translocation of NF-κB from cytosol to nucleus in HaCaT cells was inhibited by [6]-gingerol via suppression of IκBα phosphorylation.
5.9. Soy Isoflavone
Genistein, (4′,5,7-trihydroxyisoflavone) the most abundant isoflavone of the soy derived phytoestrogen compounds, is a potent antioxidant and inhibitor of tyrosine kinase activity. Genistein, daidzein (4′,7-dihydroxyisoflavone), and their 7-glycosides, genistin, and daidzin are the major isoflavonoids of soybeans and soy products. Soybean containing diet is associated with cancer chemoprevention. Genistein exerts its anti-initiational and promotional effects on skin carcinogenesis probably through blockage of DNA adduct formation and inhibition of oxidative (H2O2) and inflammatory (ODC activity) events in mouse skin [160]. In another study pretreatment of animals with 10 μM of genistein 1 h prior to UVB exposure significantly inhibited UVB-induced H2O2 and MDA in skin and 8-OHdG in epidermis as well as internal organs [161]. Genistein protects the skin from long-term psoralen plus ultraviolet radiation induced skin damages [162]. Additionally, the inhibitory effect of soy isoflavones on TPA-induced cutaneous inflammation includes inhibition of proinflammatory cytokines, attenuation of oxidative stress, and activation of NF-κB and expression of Cox-2 [163]. Moore et al. [164] studied genistein's photoprotective efficacy within the context of full thickness human reconstituted skin. Genistein (10, 20, and 50 μM) 1 h prior to UVB (20 and 60 mJ/cm2) radiation inhibited UV-induced DNA damage and dose dependently preserved cutaneous proliferation and repair mechanics in skin samples.
5.10. Diallyl Sulphide (DAS)
DAS is an organosulfur compound derived from garlic (Allium sativum L.) and a few other genus of Allium plants and is well known to be beneficial for the prevention of lifestyle-related diseases. The therapeutic uses of garlic in cancer have been widely studied. Modulation in p53 and p21/waf1 expression by topical application of DAS was recorded in DMBA-induced mouse skin tumours [165, 166]. DMBA induced antiapoptotic proteins (survivin and bcl-2), oncoprotein (Ras), and signaling molecules activation (PI3K/Akt and MAPKs) were significantly downregulated by DAS supplementation [167]. Topical application of liposome-based formulation of DAS (250 microg/mouse), one hour after DMBA exposure, not only resulted in delay of tumorogenesis but also reduced the cumulative number and size of papillomas [168]. The pretreatment of DAS (10 mg/kg body-weight) showed 68.35% protection and posttreatment showed 59.49% protection, at an intermittent period of 48 h, against DMBA-induced DNA strand breakage in mouse skin [169].
6. Miscellaneous
Several other phytochemicals also have been reported for their imperative effects against skin cancer. These phytochemicals are enlisted in table with their respective sources and their mode of action in skin cancer prevention.
7. Conclusions
The phytochemicals possess antioxidant and antiinflammatory properties through which they contribute to their anticarcinogenic effects and to modulate various biochemical processes and molecular signaling pathways induced or mediated by carcinogens. Based on laboratory and epidemiological evidences it can be concluded that routine consumption/application of these natural agents may provide proficient safeguard against damaging effects of solar ultraviolet radiation as well as other skin carcinogens present in environment. Likewise, these natural agents in combination with sunscreens or skin care cosmetics may offer a rational approach for reducing skin cancers and other skin diseases.
Conflict of Interests
The authors declare that there is no conflict of interests regarding the publication of this paper.
References
- 1.Newell GR, Sider JG, Bergfelt L, Kripke ML. Incidence of cutaneous melanoma in the United States by histology with special reference to the face. Cancer Research. 1988;48(17):5036–5041. [PubMed] [Google Scholar]
- 2.American Cancer Society. Cancer Facts and Figures 2012. 2012, http://www.cancer.org/acs/groups/content/@epidemiologysurveilance/documents/document/acspc-031941.pdf.
- 3.Jerant AF, Johnson JT, Sheridan CD, Caffrey TJ. Early detection and treatment of skin cancer. American Family Physician. 2000;62(2):357–382. [PubMed] [Google Scholar]
- 4.Armstrong BK, Kricker A. How much melanoma is caused by sun exposure. Melanoma Research. 1993;3(6):395–401. doi: 10.1097/00008390-199311000-00002. [DOI] [PubMed] [Google Scholar]
- 5.Tanos T, Marinissen MJ, Leskow FC, et al. Phosphorylation of c-Fos by members of the p38 MAPK family: role in the AP-1 response to UV light. Journal of Biological Chemistry. 2005;280(19):18842–18852. doi: 10.1074/jbc.M500620200. [DOI] [PubMed] [Google Scholar]
- 6.Miller DL, Weinstock MA. Nonmelanoma skin cancer in the United States: incidence. Journal of the American Academy of Dermatology. 1994;30(5):774–778. doi: 10.1016/s0190-9622(08)81509-5. [DOI] [PubMed] [Google Scholar]
- 7.Kligman LH. Photoaging. Manifestations, prevention, and treatment. Dermatologic Clinics. 1986;4(3):517–528. [PubMed] [Google Scholar]
- 8.Mukhtar H, Elmets CA. Photocarcinogenesis: mechanisms, models and human health implications. Photochemistry and Photobiology. 1996;63(4):355–447. doi: 10.1111/j.1751-1097.1996.tb03040.x. [DOI] [PubMed] [Google Scholar]
- 9.Chen AC, Halliday GM, Damian DL. Non-melanoma skin cancer: carcinogenesis and chemoprevention. Pathology. 2013;45(3):331–341. doi: 10.1097/PAT.0b013e32835f515c. [DOI] [PubMed] [Google Scholar]
- 10.Nichols JA, Katiyar SK. Skin photoprotection by natural polyphenols: anti-inflammatory, antioxidant and DNA repair mechanisms. Archives of Dermatological Research. 2010;302(2):71–83. doi: 10.1007/s00403-009-1001-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Boffetta P, Jourenkova N, Gustavsson P. Cancer risk from occupational and environmental exposure to polycyclic aromatic hydrocarbons. Cancer Causes and Control. 1997;8(3):444–472. doi: 10.1023/a:1018465507029. [DOI] [PubMed] [Google Scholar]
- 12.Millard LG. Multiple pigmented papular basal cell carcinomas: a new pattern of industrial tar induced skin tumours. British Journal of Industrial Medicine. 1986;43(2):134–136. doi: 10.1136/oem.43.2.134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Surdu S, Fitzgerald EF, Bloom MS, et al. Occupational exposure to arsenic and risk of non-melanoma skin cancer in a multinational European study. International Journal of Cancer. 2013;133(9):2182–2191. doi: 10.1002/ijc.28216. [DOI] [PubMed] [Google Scholar]
- 14.Molho-Pessach V, Lotem M. Viral carcinogenesis in skin cancer. Current Problems in Dermatology. 2007;35:39–51. doi: 10.1159/000106409. [DOI] [PubMed] [Google Scholar]
- 15.Filler RB, Roberts SJ, Girardi M. Cutaneous two-stage chemical carcinogenesis. CSH Protocol. 2007;2007 doi: 10.1101/pdb.prot4837.pdb.prot4837 [DOI] [PubMed] [Google Scholar]
- 16.Miyata M, Furukawa M, Takahashi K, Gonzalez FJ, Yamazoe Y. Mechanism of 7,12-dimethylbenz[a]anthracene-induced immunotoxicity: role of metabolic activation at the target organ. Japanese Journal of Pharmacology. 2001;86(3):302–309. doi: 10.1254/jjp.86.302. [DOI] [PubMed] [Google Scholar]
- 17.Pearson G, Robinson F, Gibson TB, et al. Mitogen-activated protein (MAP) kinase pathways: regulation and physiological functions. Endocrine Reviews. 2001;22(2):153–183. doi: 10.1210/edrv.22.2.0428. [DOI] [PubMed] [Google Scholar]
- 18.Cowan KJ, Storey KB. Mitogen-activated protein kinases: new signaling pathways functioning in cellular responses to environmental stress. Journal of Experimental Biology. 2003;206(7):1107–1115. doi: 10.1242/jeb.00220. [DOI] [PubMed] [Google Scholar]
- 19.Cobb MH, Goldsmith EJ. How MAP kinases are regulated. Journal of Biological Chemistry. 1995;270(25):14843–14846. doi: 10.1074/jbc.270.25.14843. [DOI] [PubMed] [Google Scholar]
- 20.Rajagopalan H, Bardelli A, Lengauer C, Kinzler KW, Vogelstein B, Velculescu VE. RAF/RAS oncogenes and mismatch-repair status. Nature. 2002;418(6901):p. 934. doi: 10.1038/418934a. [DOI] [PubMed] [Google Scholar]
- 21.Djavaheri-Mergny M, Dubertret L. UV-A-induced AP-1 activation requires the Raf/ERK pathway in human NCTC 2544 keratinocytes. Experimental Dermatology. 2001;10(3):204–210. doi: 10.1034/j.1600-0625.2001.010003204.x. [DOI] [PubMed] [Google Scholar]
- 22.Hoyos B, Imam A, Korichneva I, Levi E, Chua R, Hammerling U. Activation of c-Raf kinase by ultraviolet light: regulation by retinoids. Journal of Biological Chemistry. 2002;277(26):23949–23957. doi: 10.1074/jbc.M110750200. [DOI] [PubMed] [Google Scholar]
- 23.Verma IM, Stevenson JK, Schwarz EM, van Antwerp D, Miyamoto S. Rel/NF-κB/IκB family: intimate tales of association and dissociation. Genes and Development. 1995;9(22):2723–2735. doi: 10.1101/gad.9.22.2723. [DOI] [PubMed] [Google Scholar]
- 24.Zhang JY, Selim MA. The role of the c-Jun N-terminal Kinase signaling pathway in skin cancer. American Journal of Cancer Research. 2012;2(6):691–698. [PMC free article] [PubMed] [Google Scholar]
- 25.Liu K, Yu D, Cho YY, et al. Sunlight UV-induced skin cancer relies upon activation of the p38α signaling pathway. Cancer Research. 2013;73(7):2181–2188. doi: 10.1158/0008-5472.CAN-12-3408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Dickinson SE, Olson ER, Zhang J, et al. P38 MAP kinase plays a functional role in UVB-Induced mouse skin carcinogenesis. Molecular Carcinogenesis. 2011;50(6):469–478. doi: 10.1002/mc.20734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Cantley LC. The phosphoinositide 3-kinase pathway. Science. 2002;296(5573):1655–1657. doi: 10.1126/science.296.5573.1655. [DOI] [PubMed] [Google Scholar]
- 28.Wan YS, Wang ZQ, Shao Y, Voorhees JJ, Fisher GJ. Ultraviolet irradiation activates PI 3-kinase/AKT survival pathway via EGF receptors in human skin in vivo. International journal of oncology. 2001;18(3):461–466. doi: 10.3892/ijo.18.3.461. [DOI] [PubMed] [Google Scholar]
- 29.Samuels Y, Wang Z, Bardelli A, et al. High Frequency of Mutations of the PIK3CA Gene in Human Cancers. Science. 2004;304(5670):p. 554. doi: 10.1126/science.1096502. [DOI] [PubMed] [Google Scholar]
- 30.Brunet A, Bonni A, Zigmond MJ, et al. Akt promotes cell survival by phosphorylating and inhibiting a forkhead transcription factor. Cell. 1999;96(6):857–868. doi: 10.1016/s0092-8674(00)80595-4. [DOI] [PubMed] [Google Scholar]
- 31.Los M, Maddika S, Erb B, Schulze-Osthoff K. Switching akt: from survival signaling to deadly response. BioEssays. 2009;31(5):492–495. doi: 10.1002/bies.200900005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Ozes ON, Mayo LD, Gustin JA, Pfeffer SR, Pfeffer LM, Donner DB. NF-κB activation by tumour necrosis factor requires tie Akt serine-threonine kinase. Nature. 1999;401(6748):82–85. doi: 10.1038/43466. [DOI] [PubMed] [Google Scholar]
- 33.Castellano E, Downward J. Ras interaction with PI3K: more than just another effector pathway. Genes and Cancer. 2011;2(3):261–274. doi: 10.1177/1947601911408079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Müller-Decker K. Cyclooxygenase-dependent signaling is causally linked to non-melanoma skin carcinogenesis: pharmacological, genetic, and clinical evidence. Cancer and Metastasis Reviews. 2011;30(3-4):343–361. doi: 10.1007/s10555-011-9306-z. [DOI] [PubMed] [Google Scholar]
- 35.Rundhaug JE, Mikulec C, Pavone A, Fischer SM. A role for cyclooxygenase-2 in ultraviolet light-induced skin carcinogenesis. Molecular Carcinogenesis. 2007;46(8):692–698. doi: 10.1002/mc.20329. [DOI] [PubMed] [Google Scholar]
- 36.Fischer SM. Is cyclooxygenase-2 important in skin carcinogenesis? Journal of Environmental Pathology, Toxicology and Oncology. 2002;21(2):183–191. [PubMed] [Google Scholar]
- 37.Giaccia A, Siim BG, Johnson RS. HIF-1 as a target for drug development. Nature Reviews Drug Discovery. 2003;2(10):803–811. doi: 10.1038/nrd1199. [DOI] [PubMed] [Google Scholar]
- 38.Strieter RM. Masters of angiogenesis. Nature Medicine. 2005;11(9):925–927. doi: 10.1038/nm0905-925. [DOI] [PubMed] [Google Scholar]
- 39.Cho Y-S, Kim C-H, Park J-W. Involvement of HIF-1α in UVB-induced epidermal hyperplasia. Molecules and Cells. 2009;28(6):537–543. doi: 10.1007/s10059-009-0148-2. [DOI] [PubMed] [Google Scholar]
- 40.Li Y, Bi Z, Yan B, Wan Y. UVB radiation induces expression of HIF-1α and VEGF through the EGFR/PI3K/DEC1 pathway. International Journal of Molecular Medicine. 2006;18(4):713–719. [PubMed] [Google Scholar]
- 41.Hertog MGL, Hollman PCH, Katan MB, Kromhout D. Intake of potentially anticarcinogenic flavonoids and their determinants in adults in The Netherlands. Nutrition and Cancer. 1993;20(1):21–29. doi: 10.1080/01635589309514267. [DOI] [PubMed] [Google Scholar]
- 42.Mittal A, Elmets CA, Katiyar SK. Dietary feeding of proanthocyanidins from grape seeds prevents photocarcinogenesis in SKH-1 hairless mice: relationship to decreased fat and lipid peroxidation. Carcinogenesis. 2003;24(8):1379–1388. doi: 10.1093/carcin/bgg095. [DOI] [PubMed] [Google Scholar]
- 43.Meeran SM, Vaid M, Punathil T, Katiyar SK. Dietary grape seed proanthocyanidins inhibit 12-O-tetradecanoyl phorbol-13-acetate-caused skin tumor promotion in 7,12-dimethylbenz[a]anthracene-initiated mouse skin, which is associated with the inhibition of inflammatory responses. Carcinogenesis. 2009;30(3):520–528. doi: 10.1093/carcin/bgp019. [DOI] [PubMed] [Google Scholar]
- 44.Nandakumar V, Singh T, Katiyar SK. Multi-targeted prevention and therapy of cancer by proanthocyanidins. Cancer Letters. 2008;269(2):378–387. doi: 10.1016/j.canlet.2008.03.049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Yamakoshi J, Saito M, Kataoka S, Kikuchi M. Safety evaluation of proanthocyanidin-rich extract from grape seeds. Food and Chemical Toxicology. 2002;40(5):599–607. doi: 10.1016/s0278-6915(02)00006-6. [DOI] [PubMed] [Google Scholar]
- 46.Manach C, Scalbert A, Morand C, Rémésy C, Jiménez L. Polyphenols: food sources and bioavailability. The American Journal of Clinical Nutrition. 2004;79(5):727–747. doi: 10.1093/ajcn/79.5.727. [DOI] [PubMed] [Google Scholar]
- 47.Zhao J, Wang J, Chen Y, Agarwal R. Anti-tumor-promoting activity of a polyphenolic fraction isolated from grape seeds in the mouse skin two-stage initiation-promotion protocol and identification of procyanidin B5-3′-gallate as the most effective antioxidant constituent. Carcinogenesis. 1999;20(9):1737–1745. doi: 10.1093/carcin/20.9.1737. [DOI] [PubMed] [Google Scholar]
- 48.Sharma SD, Meeran SM, Katiyar SK. Dietary grape seed proanthocyanidins inhibit UVB-induced oxidative stress and activation of mitogen-activated protein kinases and nuclear factor-κB signaling in in vivo SKH-1 hairless mice. Molecular Cancer Therapeutics. 2007;6(3):995–1005. doi: 10.1158/1535-7163.MCT-06-0661. [DOI] [PubMed] [Google Scholar]
- 49.Sharma SD, Katiyar SK. Dietary grape seed proanthocyanidins inhibit UVB-induced cyclooxygenase-2 expression and other inflammatory mediators in UVB-exposed skin and skin tumors of SKH-1 hairless mice. Pharmaceutical Research. 2010;27(6):1092–1102. doi: 10.1007/s11095-010-0050-9. [DOI] [PubMed] [Google Scholar]
- 50.Vaid M, Singh T, Prasad R, Elmets CA, Xu H, Katiyar SK. Bioactive grape proanthocyanidins enhance immune reactivity in UV-irradiated skin through functional activation of dendritic cells in mice. Cancer Prevention Research. 2013;6(3):242–252. doi: 10.1158/1940-6207.CAPR-12-0320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Yuan X-Y, Liu W, Hao J-C, Gu W-J, Zhao Y-S. Topical grape seed proanthocyandin extract reduces sunburn cells and mutant p53 positive epidermal cell formation, and prevents depletion of langerhans cells in an acute sunburn model. Photomedicine and Laser Surgery. 2012;30(1):20–25. doi: 10.1089/pho.2011.3043. [DOI] [PubMed] [Google Scholar]
- 52.Bomser JA, Singletary KW, Wallig MA, Smith MAL. Inhibition of TPA-induced tumor promotion in CD-1 mouse epidermis by a polyphenolic fraction from grape seeds. Cancer Letters. 1999;135(2):151–157. doi: 10.1016/s0304-3835(98)00289-4. [DOI] [PubMed] [Google Scholar]
- 53.Bomser J, Singletary K, Meline B. Inhibition of 12-O-tetradecanoylphorbol-13-acetate (TPA)-induced mouse skin ornithine decarboxylase and protein kinase C by polyphenolics from grapes. Chemico-Biological Interactions. 2000;127(1):45–59. doi: 10.1016/s0009-2797(00)00170-8. [DOI] [PubMed] [Google Scholar]
- 54.Mantena SK, Katiyar SK. Grape seed proanthocyanidins inhibit UV-radiation-induced oxidative stress and activation of MAPK and NF-κB signaling in human epidermal keratinocytes. Free Radical Biology and Medicine. 2006;40(9):1603–1614. doi: 10.1016/j.freeradbiomed.2005.12.032. [DOI] [PubMed] [Google Scholar]
- 55.Meeran SM, Katiyar SK. Grape seed proanthocyanidins promote apoptosis in human epidermoid carcinoma A431 cells through alterations in Cdki-Cdk-cyclin cascade, and caspase-3 activation via loss of mitochondrial membrane potential. Experimental Dermatology. 2007;16(5):405–415. doi: 10.1111/j.1600-0625.2007.00542.x. [DOI] [PubMed] [Google Scholar]
- 56.Vaid M, Singh T, Katiyar SK. Grape seed Proanthocyanidins inhibit Melanoma cell invasiveness by reduction of PGE2 synthesis and reversal of Epithelial-to-Mesenchymal transition. PLoS ONE. 2011;6(6) doi: 10.1371/journal.pone.0021539.e21539 [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 57.Vaid M, Prasad R, Singh T, Jones V, Katiyar SK. Grape seed proanthocyanidins reactivate silenced tumor suppressor genes in human skin cancer cells by targeting epigenetic regulators. Toxicology and Applied Pharmacology. 2012;263(1):122–130. doi: 10.1016/j.taap.2012.06.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Kim W, Yang JH, Youn H, Yun YJ, Seong KM, Youn B. Myricetin inhibits Akt survival signaling and induces badmediated apoptosis in a low dose ultraviolet (UV)-B-irradiated hacat human immortalized keratinocytes. Journal of Radiation Research. 2010;51(3):285–296. doi: 10.1269/jrr.09141. [DOI] [PubMed] [Google Scholar]
- 59.Jung SK, Lee KW, Byun S, et al. Myricetin suppresses UVB-induced skin cancer by targeting Fyn. Cancer Research. 2008;68(14):6021–6029. doi: 10.1158/0008-5472.CAN-08-0899. [DOI] [PubMed] [Google Scholar]
- 60.Kang NJ, Lee KW, Shin BJ, et al. Caffeic acid, a phenolic phytochemical in coffee, directly inhibits Fyn kinase activity and UVB-induced COX-2 expression. Carcinogenesis. 2009;30(2):321–330. doi: 10.1093/carcin/bgn282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Arasada BL, Bommareddy A, Zhang X, Bremmon K, Dwivedi C. Effects of α-santalol on proapoptotic caspases and p53 expression in UVB irradiated mouse skin. Anticancer Research. 2008;28(1):129–132. [PubMed] [Google Scholar]
- 62.Surjana D, Halliday GM, Damian DL. Nicotinamide enhances repair of ultraviolet radiation-induced DNA damage in human keratinocytes and ex vivo skin. Carcinogenesis. 2013;34(5):1144–1149. doi: 10.1093/carcin/bgt017. [DOI] [PubMed] [Google Scholar]
- 63.Li J, Malakhova M, Mottamal M, et al. Norathyriol suppresses skin cancers induced by solar ultraviolet radiation by targeting ERK kinases. Cancer Research. 2012;72(1):260–270. doi: 10.1158/0008-5472.CAN-11-2596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Bettoli V, Zauli S, Virgili A. Retinoids in the chemoprevention of non-elanoma skin cancers: why, when and how. Journal of Dermatological Treatment. 2013;24(3):235–237. doi: 10.3109/09546634.2012.746634. [DOI] [PubMed] [Google Scholar]
- 65.Li W, Liu J, Zhao Y. PKM2 inhibitor shikonin suppresses TPA-induced mitochondrial malfunction and proliferation of skin epidermal JB6 cells. Molecular Carcinogenesis. 2012 doi: 10.1002/mc.21988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Staniforth V, Huang W-C, Aravindaram K, Yang N-S. Ferulic acid, a phenolic phytochemical, inhibits UVB-induced matrix metalloproteinases in mouse skin via posttranslational mechanisms. Journal of Nutritional Biochemistry. 2012;23(5):443–451. doi: 10.1016/j.jnutbio.2011.01.009. [DOI] [PubMed] [Google Scholar]
- 67.Vaid M, Sharma SD, Katiyar SK. Honokiol, a phytochemical from the Magnolia plant, inhibits photocarcinogenesis by targeting UVB-induced inflammatory mediators and cell cycle regulators: development of topical formulation. Carcinogenesis. 2010;31(11):2004–2011. doi: 10.1093/carcin/bgq186. [DOI] [PubMed] [Google Scholar]
- 68.Sime S, Reeve VE. Protection from inflammation, immunosuppression and carcinogenesis induced by UV radiation in mice by topical pycnogenol B7. Photochemistry and Photobiology. 2004;79(2):193–198. doi: 10.1562/0031-8655(2004)079<0193:pfiiac>2.0.co;2. [DOI] [PubMed] [Google Scholar]
- 69.Sarfaraz S, Siddiqui IA, Syed DN, Afaq F, Mukhtar H. Guggulsterone modulates MAPK and NF-κB pathways and inhibits skin tumorigenesis in SENCAR mice. Carcinogenesis. 2008;29(10):2011–2018. doi: 10.1093/carcin/bgn180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Lee J-C, Kundu JK, Hwang D-M, Na H-K, Surh Y-J. Humulone inhibits phorbol ester-induced COX-2 expression in mouse skin by blocking activation of NF-κB and AP-1: IκB kinase and c-Jun-N-terminal kinase as respective potential upstream targets. Carcinogenesis. 2007;28(7):1491–1498. doi: 10.1093/carcin/bgm054. [DOI] [PubMed] [Google Scholar]
- 71.Byun S, Park J, Lee E, et al. Src kinase is a direct target of apigenin against UVB-induced skin inflammation. Carcinogenesis. 2013;34(2):397–405. doi: 10.1093/carcin/bgs358. [DOI] [PubMed] [Google Scholar]
- 72.Lai C-S, Li S, Chai C-Y, et al. Inhibitory effect of citrus 5-hydroxy-3,6,7,8,3′,4′-hexamethoxyflavone on 12-O-tetradecanoylphorbol 13-acetate-induced skin inflammation and tumor promotion in mice. Carcinogenesis. 2007;28(12):2581–2588. doi: 10.1093/carcin/bgm231. [DOI] [PubMed] [Google Scholar]
- 73.Kim J, Park H, Kim J, Park JHY. 3,3′-Diindolylmethane suppresses 12-O-tetradecanoylphorbol-13-acetate-induced inflammation and tumor promotion in mouse skin via the downregulation of inflammatory mediators. Molecular Carcinogenesis. 2010;49(7):672–683. doi: 10.1002/mc.20640. [DOI] [PubMed] [Google Scholar]
- 74.Chung WY, Park JH, Kim MJ, et al. Xanthorrhizol inhibits 12-O-tetradecanoylphorbol-13-acetate-induced acute inflammation and two-stage mouse skin carcinogenesis by blocking the expression of ornithine decarboxylase, cyclooxygenase-2 and inducible nitric oxide synthase through mitogen-activated protein kinases and/or the nuclear factor-κB. Carcinogenesis. 2007;28(6):1224–1231. doi: 10.1093/carcin/bgm005. [DOI] [PubMed] [Google Scholar]
- 75.Afaq F, Saleem M, Aziz MH, Mukhtar H. Inhibition of 12-O-tetradecanoylphorbol-13-acetate-induced tumor promotion markers in CD-1 mouse skin by oleandrin. Toxicology and Applied Pharmacology. 2004;195(3):361–369. doi: 10.1016/j.taap.2003.09.027. [DOI] [PubMed] [Google Scholar]
- 76.Afaq F, Syed DN, Malik A, et al. Delphinidin, an anthocyanidin in pigmented fruits and vegetables, protects human HaCaT keratinocytes and mouse skin against UVB-mediated oxidative stress and apoptosis. Journal of Investigative Dermatology. 2007;127(1):222–232. doi: 10.1038/sj.jid.5700510. [DOI] [PubMed] [Google Scholar]
- 77.Decker EA. Phenolics: prooxidants or antioxidants? Nutrition Reviews. 1997;55(11):396–398. doi: 10.1111/j.1753-4887.1997.tb01580.x. [DOI] [PubMed] [Google Scholar]
- 78.Jain A, Manghani C, Kohli S, Nigam D, Rani V. Tea and human health: the dark shadows. Toxicology Letters. 2013;220(1):82–87. doi: 10.1016/j.toxlet.2013.04.010. [DOI] [PubMed] [Google Scholar]
- 79.Wang ZY, Agarwal R, Bickers DR, Mukhtar H. Protection against ultraviolet B radiation-induced photocarcinogenesis in hairless mice by green tea polyphenols. Carcinogenesis. 1991;12(8):1527–1530. doi: 10.1093/carcin/12.8.1527. [DOI] [PubMed] [Google Scholar]
- 80.Agarwal R, Katiyar SK, Khan SG, Mukhtar H. Protection against ultraviolet B radiation-induced effects in the skin of SKH-1 hairless mice by a polyphenolic fraction isolated from green tea. Photochemistry and Photobiology. 1993;58(5):695–700. doi: 10.1111/j.1751-1097.1993.tb04954.x. [DOI] [PubMed] [Google Scholar]
- 81.Katiyar SK, Vaid M, van Steeg H, Meeran SM. Green tea polyphenols prevent uv-induced immunosuppression by rapid repair of DNA damage and enhancement of nucleotide excision repair genes. Cancer Prevention Research. 2010;3(2):179–189. doi: 10.1158/1940-6207.CAPR-09-0044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Meeran SM, Akhtar S, Katiyar SK. Inhibition of UVB-induced skin tumor development by drinking green tea polyphenols is mediated through DNA repair and subsequent inhibition of inflammation. Journal of Investigative Dermatology. 2009;129(5):1258–1270. doi: 10.1038/jid.2008.354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Afaq F, Ahmad N, Mukhtar H. Suppression of UVB-induced phosphorylation of mitogen-activated protein kinases and nuclear factor kappa B by green tea polyphenol in SKH-1 hairless mice. Oncogene. 2003;22(58):9254–9264. doi: 10.1038/sj.onc.1207035. [DOI] [PubMed] [Google Scholar]
- 84.Katiyar SK, Challa A, McCormick TS, Cooper KD, Mukhtar H. Prevention of UVB-induced immunosuppression in mice by the green tea polyphenol (-)-epigallocatechin-3-gallate may be associated with alterations in IL-10 and IL-12 production. Carcinogenesis. 1999;20(11):2117–2124. doi: 10.1093/carcin/20.11.2117. [DOI] [PubMed] [Google Scholar]
- 85.Meeran SM, Mantena SK, Katiyar SK. Prevention of ultraviolet radiation—induced immunosuppression by (-)-epigallocatechin-3-gallate in mice is mediated through interleukin 12-dependent DNA repair. Clinical Cancer Research. 2006;12(7, part 1):2272–2280. doi: 10.1158/1078-0432.CCR-05-2672. [DOI] [PubMed] [Google Scholar]
- 86.Meeran SM, Mantena SK, Elmets CA, Katiyar SK. (-)-Epigallocatechin-3-gallate prevents photocarcinogenesis in mice through interleukin-12-dependent DNA repair. Cancer Research. 2006;66(10):5512–5520. doi: 10.1158/0008-5472.CAN-06-0218. [DOI] [PubMed] [Google Scholar]
- 87.Katiyar SK, Mukhtar H. Green tea polyphenol (-)-epigallocatechin-3-gallate treatment to mouse skin prevents UVB-induced infiltration of leukocytes, depletion of antigen-presenting cells, and oxidative stress. Journal of Leukocyte Biology. 2001;69(5):719–726. [PubMed] [Google Scholar]
- 88.Vayalil PK, Elments CA, Katiyar SK. Treatment of green tea polyphenols in hydrophilic cream prevents UVB-induced oxidation of lipids and proteins, depletion of antioxidant enzymes and phosphorylation of MAPK proteins in SKH-1 hairless mouse skin. Carcinogenesis. 2003;24(5):927–936. doi: 10.1093/carcin/bgg025. [DOI] [PubMed] [Google Scholar]
- 89.Katiyar SK, Mukhtar H. Inhibition of phorbol ester tumor promoter 12-O-tetradecanoylphorbol-13-acetate-caused inflammatory responses in SENCAR mouse skin by black tea polyphenols. Carcinogenesis. 1997;18(10):1911–1916. doi: 10.1093/carcin/18.10.1911. [DOI] [PubMed] [Google Scholar]
- 90.Patel R, Krishnan R, Ramchandani A, Maru G. Polymeric black tea polyphenols inhibit mouse skin chemical carcinogenesis by decreasing cell proliferation. Cell Proliferation. 2008;41(3):532–553. doi: 10.1111/j.1365-2184.2008.00528.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Kumar G, Dange P, Kailaje V, Vaidya MM, Ramchandani AG, Maru GB. Polymeric black tea polyphenols modulate the localization and activity of 12-O-tetradecanoylphorbol-13-acetate-mediated kinases in mouse skin: mechanisms of their anti-tumor-promoting action. Free Radical Biology & Medicine. 2012;53(6):1358–1370. doi: 10.1016/j.freeradbiomed.2012.07.017. [DOI] [PubMed] [Google Scholar]
- 92.Katiyar SK, Afaq F, Azizuddin K, Mukhtar H. Inhibition of UVB-induced oxidative stress-mediated phosphorylation of mitogen-activated protein kinase signaling pathways in cultured human epidermal keratinocytes by green tea polyphenol ( - )-epigallocatechin-3-gallate. Toxicology and Applied Pharmacology. 2001;176(2):110–117. doi: 10.1006/taap.2001.9276. [DOI] [PubMed] [Google Scholar]
- 93.Afaq F, Adhami VM, Ahmad N, Mukhtar H. Inhibition of ultraviolet B-mediated activation of nuclear factor κB in normal human epidermal keratinocytes by green tea Constituent (-)-epigallocatechin-3-gallate. Oncogene. 2003;22(7):1035–1044. doi: 10.1038/sj.onc.1206206. [DOI] [PubMed] [Google Scholar]
- 94.Ahmad N, Gupta S, Mukhtar H. Green tea polyphenol epigallocatechin-3-gallate differentially modulates nuclear factor κB in cancer cells versus normal cells. Archives of Biochemistry and Biophysics. 2000;376(2):338–346. doi: 10.1006/abbi.2000.1742. [DOI] [PubMed] [Google Scholar]
- 95.Balasubramanian S, Adhikary G, Eckert RL. The Bmi-1 polycomb protein antagonizes the (-)-epigallocatechin-3-gallate-dependent suppression of skin cancer cell survival. Carcinogenesis. 2010;31(3):496–503. doi: 10.1093/carcin/bgp314. [DOI] [PMC free article] [PubMed] [Google Scholar] [Research Misconduct Found]
- 96.Choudhury SR, Balasubramanian S, Chew YC, Han B, Marquez VE, Eckert RL. (-)-Epigallocatechin-3-gallate and DZNep reduce polycomb protein level via a proteasome-dependent mechanism in skin cancer cells. Carcinogenesis. 2011;32(10):1525–1532. doi: 10.1093/carcin/bgr171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Bickers DR, Athar M. Novel approaches to chemoprevention of skin cancer. Journal of Dermatology. 2000;27(11):691–695. doi: 10.1111/j.1346-8138.2000.tb02259.x. [DOI] [PubMed] [Google Scholar]
- 98.Hakim IA, Harris RB, Weisgerber UM. Tea intake and squamous cell carcinoma of the skin: influence of type of tea beverages. Cancer Epidemiology Biomarkers and Prevention. 2000;9(7):727–731. [PubMed] [Google Scholar]
- 99.Katiyar SK, Perez A, Mukhtar H. Green tea polyphenol treatment to human skin prevents formation of ultraviolet light B-induced pyrimidine dimers in DNA. Clinical Cancer Research. 2000;6(10):3864–3869. [PubMed] [Google Scholar]
- 100.Jurenka JS. Therapeutic applications of pomegranate (Punica granatum L.): a review. Alternative Medicine Review. 2008;13(2):128–144. [PubMed] [Google Scholar]
- 101.Park HM, Moon E, Kim A-J, et al. Extract of Punica granatum inhibits skin photoaging induced by UVB irradiation. International Journal of Dermatology. 2010;49(3):276–282. doi: 10.1111/j.1365-4632.2009.04269.x. [DOI] [PubMed] [Google Scholar]
- 102.Adhami VM, Khan N, Mukhtar H. Cancer chemoprevention by pomegranate: laboratory and clinical evidence. Nutrition and Cancer. 2009;61(6):811–815. doi: 10.1080/01635580903285064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Afaq F, Saleem M, Krueger CG, Reed JD, Mukhtar H. Anthocyanin- and hydrolyzable tannin-rich pomegranate fruit extract modulates MAPK and NF-κB pathways and inhibits skin tumorigenesis in CD-1 mice. International Journal of Cancer. 2005;113(3):423–433. doi: 10.1002/ijc.20587. [DOI] [PubMed] [Google Scholar]
- 104.Hora JJ, Maydew ER, Lansky EP, Dwivedi C. Chemopreventive effects of pomegranate seed oil on skin tumor development in CD1 mice. Journal of Medicinal Food. 2003;6(3):157–161. doi: 10.1089/10966200360716553. [DOI] [PubMed] [Google Scholar]
- 105.Afaq F, Khan N, Syed DN, Mukhtar H. Oral feeding of pomegranate fruit extract inhibits early biomarkers of UVB radiation-induced carcinogenesis in SKH-1 hairless mouse epidermis. Photochemistry and Photobiology. 2010;86(6):1318–1326. doi: 10.1111/j.1751-1097.2010.00815.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Khan N, Syed DN, Pal HC, Mukhtar H, Afaq F. Pomegranate fruit extract inhibits UVB-induced inflammation and proliferation by modulating NF-κB and MAPK signaling pathways in mouse skin. Photochemistry and Photobiology. 2012;88(5):1126–1134. doi: 10.1111/j.1751-1097.2011.01063.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Afaq F, Malik A, Syed D, Maes D, Matsui MS, Mukhtar H. Pomegranate fruit extract modulates UV-B-mediated phosphorylation of mitogen-activated protein kinases and activation of nuclear factor kappa B in normal human epidermal keratinocytes. Photochemistry and Photobiology. 2005;81(1):38–45. doi: 10.1562/2004-08-06-RA-264. [DOI] [PubMed] [Google Scholar]
- 108.Syed DN, Malik A, Hadi N, Sarfaraz S, Afaq F, Mukhtar H. Photochemopreventive effect of pomegranate fruit extract on UVA-mediated activation of cellular pathways in normal human epidermal keratinocytes. Photochemistry and Photobiology. 2006;82(2):398–405. doi: 10.1562/2005-06-23-RA-589. [DOI] [PubMed] [Google Scholar]
- 109.Zaid MA, Afaq F, Syed DN, Dreher M, Mukhtar H. Inhibition of UVB-mediated oxidative stress and markers of photoaging in immortalized HaCaT keratinocytes by pomegranate polyphenol extract POMx. Photochemistry and Photobiology. 2007;83(4):882–888. doi: 10.1111/j.1751-1097.2007.00157.x. [DOI] [PubMed] [Google Scholar]
- 110.Pacheco-Palencia LA, Noratto G, Hingorani L, Talcott ST, Mertens-Talcott SU. Protective effects of standardized pomegranate (Punica granatum L.) polyphenolic extract in ultraviolet-irradiated human skin fibroblasts. Journal of Agricultural and Food Chemistry. 2008;56(18):8434–8441. doi: 10.1021/jf8005307. [DOI] [PubMed] [Google Scholar]
- 111.Afaq F, Zaid MA, Khan N, Dreher M, Mukhtar H. Protective effect of pomegranate-derived products on UVB-mediated damage in human reconstituted skin. Experimental Dermatology. 2009;18(6):553–561. doi: 10.1111/j.1600-0625.2008.00829.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Kasai K, Yoshimura M, Koga T, Arii M, Kawasaki S. Effects of oral administration of ellagic acid-rich pomegranate extract on ultraviolet-induced pigmentation in the human skin. Journal of Nutritional Science and Vitaminology. 2006;52(5):383–388. doi: 10.3177/jnsv.52.383. [DOI] [PubMed] [Google Scholar]
- 113.Afaq F, Adhami VM, Ahmad N. Prevention of short-term ultraviolet B radiation-mediated damages by resveratrol in SKH-1 hairless mice. Toxicology and Applied Pharmacology. 2003;186(1):28–37. doi: 10.1016/s0041-008x(02)00014-5. [DOI] [PubMed] [Google Scholar]
- 114.Jang M, Cai L, Udeani GO, et al. Cancer chemopreventive activity of resveratrol, a natural product derived from grapes. Science. 1997;275(5297):218–220. doi: 10.1126/science.275.5297.218. [DOI] [PubMed] [Google Scholar]
- 115.Soleas GJ, Grass L, Josephy PD, Goldberg DM, Diamandis EP. A comparison of the anticarcinogenic properties of four red wine polyphenols. Clinical Biochemistry. 2002;35(2):119–124. doi: 10.1016/s0009-9120(02)00275-8. [DOI] [PubMed] [Google Scholar]
- 116.Kapadia GJ, Azuine MA, Tokuda H, et al. Chemopreventive effect of resveratrol, sesamol, sesame oil and sunflower oil in the Epstein-Barr virus early antigen activation assay and the mouse skin two-stage carcinogenesis. Pharmacological Research. 2002;45(6):499–505. doi: 10.1006/phrs.2002.0992. [DOI] [PubMed] [Google Scholar]
- 117.Fu Z-D, Cao Y, Wang K-F, Xu S-F, Han R. Chemopreventive effect of resveratrol to cancer. Chinese Journal of Cancer. 2004;23(8):869–873. [PubMed] [Google Scholar]
- 118.Reagan-Shaw S, Afaq F, Aziz MH, Ahmad N. Modulations of critical cell cycle regulatory events during chemoprevention of ultraviolet B-mediated responses by resveratrol in SKH-1 hairless mouse skin. Oncogene. 2004;23(30):5151–5160. doi: 10.1038/sj.onc.1207666. [DOI] [PubMed] [Google Scholar]
- 119.Aziz MH, Afaq F, Ahmad N. Prevention of ultraviolet-B radiation damage by resveratrol in mouse skin is mediated via modulation in survivin. Photochemistry and Photobiology. 2005;81(1):25–31. doi: 10.1562/2004-08-13-RA-274. [DOI] [PubMed] [Google Scholar]
- 120.Kalra N, Roy P, Prasad S, Shukla Y. Resveratrol induces apoptosis involving mitochondrial pathways in mouse skin tumorigenesis. Life Sciences. 2008;82(7-8):348–358. doi: 10.1016/j.lfs.2007.11.006. [DOI] [PubMed] [Google Scholar]
- 121.Kundu JK, Shin YK, Kim SH, Surh Y-J. Resveratrol inhibits phorbol ester-induced expression of COX-2 and activation of NF-κB in mouse skin by blocking IκB kinase activity. Carcinogenesis. 2006;27(7):1465–1474. doi: 10.1093/carcin/bgi349. [DOI] [PubMed] [Google Scholar]
- 122.Roy P, Kalra N, Prasad S, George J, Shukla Y. Chemopreventive potential of resveratrol in mouse skin tumors through regulation of mitochondrial and PI3K/AKT signaling pathways. Pharmaceutical Research. 2009;26(1):211–217. doi: 10.1007/s11095-008-9723-z. [DOI] [PubMed] [Google Scholar]
- 123.Yusuf N, Nasti TH, Meleth S, Elmets CA. Resveratrol enhances cell-mediated immune response to DMBA through TLR4 and prevents DMBA induced cutaneous carcinogenesis. Molecular Carcinogenesis. 2009;48(8):713–723. doi: 10.1002/mc.20517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Kowalczyk MC, Kowalczyk P, Tolstykh O, Hanausek M, Walaszek Z, Slaga TJ. Synergistic effects of combined phytochemicals and skin cancer prevention in SENCAR mice. Cancer Prevention Research. 2010;3(2):170–178. doi: 10.1158/1940-6207.CAPR-09-0196. [DOI] [PubMed] [Google Scholar]
- 125.Adhami VM, Afaq F, Ahmad N. Suppression of ultraviolet B exposure-mediated activation of NF-κB in normal human keratinocytes by resveratrol. Neoplasia. 2003;5(1):74–82. doi: 10.1016/s1476-5586(03)80019-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Adhami VM, Afaq F, Ahmad N. Involvement of the retinoblastoma (pRb)—E2F/DP pathway during antiproliferative effects of resveratrol in human epidermoid carcinoma (A431) cells. Biochemical and Biophysical Research Communications. 2001;288(3):579–585. doi: 10.1006/bbrc.2001.5819. [DOI] [PubMed] [Google Scholar]
- 127.Ahmad N, Adhami VM, Afaq F, Feyes DK, Mukhtar H. Resveratrol causes WAF-1/p21-mediated G1-phase arrest of cell cycle and induction of apoptosis in human epidermoid carcinoma A431 cells. Clinical Cancer Research. 2001;7(5):1466–1473. [PubMed] [Google Scholar]
- 128.Kim AL, Zhu Y, Zhu H, et al. Resveratrol inhibits proliferation of human epidermoid carcinoma A431 cells by modulating MEK1 and AP-1 signalling pathways. Experimental Dermatology. 2006;15(7):538–546. doi: 10.1111/j.1600-0625.2006.00445.x. [DOI] [PubMed] [Google Scholar]
- 129.Kim KH, Back JH, Zhu Y, et al. Resveratrol targets transforming growth factor-Β2 signaling to block UV-induced tumor progression. Journal of Investigative Dermatology. 2011;131(1):195–202. doi: 10.1038/jid.2010.250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Hao Y, Huang W, Liao M, et al. The inhibition of resveratrol to human skin squamous cell carcinoma A431 xenografts in nude mice. Fitoterapia. 2013;86:84–91. doi: 10.1016/j.fitote.2013.02.005. [DOI] [PubMed] [Google Scholar]
- 131.Bastianetto S, Dumont Y, Duranton A, Vercauteren F, Breton L, Quirion R. Protective action of resveratrol in human skin: possible involvement of specific receptor binding sites. PLoS ONE. 2010;5(9) doi: 10.1371/journal.pone.0012935.e12935 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Cottart C-H, Nivet-Antoine V, Laguillier-Morizot C, Beaudeux J-L. Resveratrol bioavailability and toxicity in humans. Molecular Nutrition and Food Research. 2010;54(1):7–16. doi: 10.1002/mnfr.200900437. [DOI] [PubMed] [Google Scholar]
- 133.Tsai ML, Lai CS, Chang YH, Chen WJ, Ho CT, Pan MH. Pterostilbene, a natural analogue of resveratrol, potently inhibits 7, 12-dimethylbenz[a]anthracene (DMBA)/12-O-tetradecanoylphorbol-13-acetate (TPA)-induced mouse skin carcinogenesis. Food & Function. 2012;3(11):1185–1194. doi: 10.1039/c2fo30105a. [DOI] [PubMed] [Google Scholar]
- 134.Rajamanickam S, Velmurugan B, Kaur M, Singh RP, Agarwal R. Chemoprevention of intestinal tumorigenesis in APCmin/+ mice by silibinin. Cancer Research. 2010;70(6):2368–2378. doi: 10.1158/0008-5472.CAN-09-3249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Lahiri-Chatterjee M, Katiyar SK, Mohan RR, Agarwal R. A flavonoid antioxidant, silymarin, affords exceptionally high protection against tumor promotion in the SENCAR mouse skin tumorigenesis model. Cancer Research. 1999;59(3):622–632. [PubMed] [Google Scholar]
- 136.Singh RP, Tyagi AK, Zhao J, Agarwal R. Silymarin inhibits growth and causes regression of established skin tumors in SENCAR mice via modulation of mitogen-activated protein kinases and induction of apoptosis. Carcinogenesis. 2002;23(3):499–510. doi: 10.1093/carcin/23.3.499. [DOI] [PubMed] [Google Scholar]
- 137.Katiyar SK, Korman NJ, Mukhtar H, Agarwal R. Protective effects of silymarin against photocarcinogenesis in a mouse skin model. Journal of the National Cancer Institute. 1997;89(8):556–566. doi: 10.1093/jnci/89.8.556. [DOI] [PubMed] [Google Scholar]
- 138.Gu M, Dhanalakshmi S, Singh RP, Agarwal R. Dietary feeding of silibinin prevents early biomarkers of UVB radiation-induced carcinogenesis in SKH-1 hairless mouse epidermis. Cancer Epidemiology Biomarkers and Prevention. 2005;14(5):1344–1349. doi: 10.1158/1055-9965.EPI-04-0664. [DOI] [PubMed] [Google Scholar]
- 139.Mallikarjuna G, Dhanalakshmi S, Singh RP, Agarwal C, Agarwal R. Silibinin protects against photocarcinogenesis via modulation of cell cycle regulators, mitogen-activated protein kinases, and Akt signaling. Cancer Research. 2004;64(17):6349–6356. doi: 10.1158/0008-5472.CAN-04-1632. [DOI] [PubMed] [Google Scholar]
- 140.Dhanalakshmi S, Mallikarjuna GU, Singh RP, Agarwal R. Silibinin prevents ultraviolent radiation-caused skin damages in SKH-1 hairless mice via a decrease in thymine dimer positive cells and an up-regulation of p53-p21/Cip1 in epidermis. Carcinogenesis. 2004;25(8):1459–1465. doi: 10.1093/carcin/bgh152. [DOI] [PubMed] [Google Scholar]
- 141.Katiyar SK, Meleth S, Sharma SD. Silymarin, a flavonoid from milk thistle (Silybum marianum L.), inhibits UV-induced oxidative stress through targeting infiltrating CD11b+ cells in mouse skin. Photochemistry and Photobiology. 2008;84(2):266–271. doi: 10.1111/j.1751-1097.2007.00241.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Wang Q, Liu W, Zeng H, et al. p53-mediated autophagy adjustment is involved in the protection of silibinin against murine dermal inflammation and epidermal apoptosis induced by UVB irradiation. Journal of Asian Natural Products Research. 2013;15(2):117–129. doi: 10.1080/10286020.2012.739616. [DOI] [PubMed] [Google Scholar]
- 143.Wang Q, Ye Y, Liu W, et al. Dual effects of silibinin treatment on autophagy-regulated dermal apoptosis retardation and epidermal apoptosis up-regulation in UVB-induced skin inflammation. Journal of Asian Natural Products Research. 2012;14(7):688–699. doi: 10.1080/10286020.2012.685725. [DOI] [PubMed] [Google Scholar]
- 144.Li L-H, Wu L-J, Tashiro S-I, Onodera S, Uchiumi F, Ikejima T. Silibinin prevents UV-induced HaCaT cell apoptosis partly through inhibition of caspase-8 pathway. Biological and Pharmaceutical Bulletin. 2006;29(6):1096–1101. doi: 10.1248/bpb.29.1096. [DOI] [PubMed] [Google Scholar]
- 145.Roy S, Deep G, Agarwal C, Agarwal R. Silibinin prevents ultraviolet B radiation-induced epidermal damages in JB6 cells and mouse skin in a p53-GADD45α-dependent manner. Carcinogenesis. 2012;33(3):629–636. doi: 10.1093/carcin/bgr299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Svobodová A, Zdarilová A, Malisková J, Mikulková H, Walterová D, Vostalová J. Attenuation of UVA-induced damage to human keratinocytes by silymarin. Journal of Dermatological Science. 2007;46(1):21–30. doi: 10.1016/j.jdermsci.2006.12.009. [DOI] [PubMed] [Google Scholar]
- 147.Narayanapillai S, Agarwal C, Tilley C, Agarwal R. Silibinin is a potent sensitizer of UVA radiation-induced oxidative stress and apoptosis in human keratinocyte haCaT cells. Photochemistry and Photobiology. 2012;88(5):1135–1140. doi: 10.1111/j.1751-1097.2011.01050.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Nigam N, Prasad S, Shukla Y. Preventive effects of lupeol on DMBA induced DNA alkylation damage in mouse skin. Food and Chemical Toxicology. 2007;45(11):2331–2335. doi: 10.1016/j.fct.2007.06.002. [DOI] [PubMed] [Google Scholar]
- 149.Saleem M, Afaq F, Adhami VM, Mukhtar H. Lupeol modulates NF-κB and PI3K/Akt pathways and inhibits skin cancer in CD-1 mice. Oncogene. 2004;23(30):5203–5214. doi: 10.1038/sj.onc.1207641. [DOI] [PubMed] [Google Scholar]
- 150.Nigam N, Prasad S, George J, Shukla Y. Lupeol induces p53 and cyclin-B-mediated G2/M arrest and targets apoptosis through activation of caspase in mouse skin. Biochemical and Biophysical Research Communications. 2009;381(2):253–258. doi: 10.1016/j.bbrc.2009.02.033. [DOI] [PubMed] [Google Scholar]
- 151.Prasad S, Madan E, Nigam N, Roy P, George J, Shukla Y. Induction of apoptosis by lupeol in human epidermoid carcinoma A431 cells through regulation of mitochondrial, Akt/PKB and NFκB signaling pathways. Cancer Biology and Therapy. 2009;8(17):1632–1639. doi: 10.4161/cbt.8.17.9204. [DOI] [PubMed] [Google Scholar]
- 152.Ishizaki C, Oguro T, Yoshida T, Wen CQ, Sueki H, Iijima M. Enhancing effect of ultraviolet A on ornithine decarboxylase induction and dermatitis evoked by 12-o-tetradecanoylphorbol-13-acetate and its inhibition by curcumin in mouse skin. Dermatology. 1996;193(4):311–317. doi: 10.1159/000246276. [DOI] [PubMed] [Google Scholar]
- 153.Oguro T, Yoshida T. Effect of ultraviolet A on ornithine decarboxylase and metallothionein gene expression in mouse skin. Photodermatology Photoimmunology and Photomedicine. 2001;17(2):71–78. doi: 10.1034/j.1600-0781.2001.017002071.x. [DOI] [PubMed] [Google Scholar]
- 154.Phillips JM, Clark C, Herman-Ferdinandez L, et al. Curcumin inhibits skin squamous cell carcinoma tumor growth in vivo. International Journal of Dermatology. 2010;49(6):608–622. doi: 10.1177/0194599811400711. [DOI] [PubMed] [Google Scholar]
- 155.Chan W-H, Wu C-C, Yu J-S. Curcumin inhibits UV irradiation-induced oxidative stress and apoptotic biochemical changes in human epidermoid carcinoma A431 cells. Journal of Cellular Biochemistry. 2003;90(2):327–338. doi: 10.1002/jcb.10638. [DOI] [PubMed] [Google Scholar]
- 156.Cho J-W, Park K, Kweon GR, et al. Curcumin inhibits the expression of COX-2 in UVB-irradiated human keratinocytes (HaCaT) by inhibiting activation of AP-1: p38 MAP kinase and JNK as potential upstream targets. Experimental and Molecular Medicine. 2005;37(3):186–192. doi: 10.1038/emm.2005.25. [DOI] [PubMed] [Google Scholar]
- 157.Chen A-L, Hsu C-H, Lin J-K, et al. Phase I clinical trial of curcumin, a chemopreventive agent, in patients with high-risk or pre-malignant lesions. Anticancer Research. 2001;21(4):2895–2900. [PubMed] [Google Scholar]
- 158.Kim J-K, Kim Y, Na K-M, Surh Y-J, Kim T-Y. [6]-gingerol prevents UVB-induced ROS production and COX-2 expression in vitro and in vivo. Free Radical Research. 2007;41(5):603–614. doi: 10.1080/10715760701209896. [DOI] [PubMed] [Google Scholar]
- 159.Bode AM, Dong Z. Signal transduction pathways: targets for chemoprevention of skin cancer. Lancet Oncology. 2000;1(3):181–188. doi: 10.1016/s1470-2045(00)00029-2. [DOI] [PubMed] [Google Scholar]
- 160.Wei H, Bowen R, Zhang X, Lebwohl M. Isoflavone genistein inhibits the initiation and promotion of two-stage skin carcinogenesis in mice. Carcinogenesis. 1998;19(8):1509–1514. doi: 10.1093/carcin/19.8.1509. [DOI] [PubMed] [Google Scholar]
- 161.Wei H, Zhang X, Wang Y, Lebwohl M. Inh olet light-induced oxidative events in the skin and internal organs of hairless mice by isoflavone genistein. Cancer Letters. 2002;185(1):21–29. doi: 10.1016/s0304-3835(02)00240-9. [DOI] [PubMed] [Google Scholar]
- 162.Shyong EQ, Lu Y, Lazinsky A, et al. Effects of the isoflavone 4′,5,7,-trihydroxyisoflavone (genistein) on psoralen plus ultraviolet A radiation (PUVA)-induced photodamage. Carcinogenesis. 2002;23(2):317–321. doi: 10.1093/carcin/23.2.317. [DOI] [PubMed] [Google Scholar]
- 163.Khan AQ, Khan R, Rehman MU, et al. Soy isoflavones (daidzein & genistein) inhibit 1-O-tetradecanoylphorbol-13-acetate (TPA)-induced cutaneous inflammation via modulation of COX-2 and NF-κB in Swiss albino mice. Toxicology. 2012;302(2-3):2266–2274. doi: 10.1016/j.tox.2012.08.008. [DOI] [PubMed] [Google Scholar]
- 164.Moore JO, Wang Y, Stebbins WG, et al. Photoprotective effect of isoflavone genistein on ultraviolet B-induced pyrimidine dimer formation and PCNA expression in human reconstituted skin and its implications in dermatology and prevention of cutaneous carcinogenesis. Carcinogenesis. 2006;27(8):1627–1635. doi: 10.1093/carcin/bgi367. [DOI] [PubMed] [Google Scholar]
- 165.Arora A, Siddiqui IA, Shukla Y. Modulation of p53 in 7,12-dimethylbenz[a]anthracene-induced skin tumors by diallyl sulfide in Swiss albino mice. Molecular Cancer Therapeutics. 2004;3(11):1459–1466. [PubMed] [Google Scholar]
- 166.Arora A, Kalra N, Shukla Y. Regulation of p21/ras protein expression by diallyl sulfide in DMBA induced neoplastic changes in mouse skin. Cancer Letters. 2006;242(1):28–36. doi: 10.1016/j.canlet.2005.10.049. [DOI] [PubMed] [Google Scholar]
- 167.Kalra N, Arora A, Shukla Y. Involvement of multiple signaling pathways in diallyl sulfide mediated apoptosis in mouse skin tumors. Asian Pacific Journal of Cancer Prevention. 2006;7(4):556–562. [PubMed] [Google Scholar]
- 168.Khan A, Shukla Y, Kalra N, et al. Potential of diallyl sulfide bearing pH-sensitive liposomes in chemoprevention against DMBA-induced skin papilloma. Molecular Medicine. 2007;13(7-8):443–451. doi: 10.2119/2006-00111.Khan. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Nigam N, Shukla Y. Preventive effects of diallyl sulfide on 7,12-dimethylbenz[a]anthracene induced DNA alkylation damage in mouse skin. Molecular Nutrition and Food Research. 2007;51(11):1324–1328. doi: 10.1002/mnfr.200700140. [DOI] [PubMed] [Google Scholar]