

Abstract
Background and Aim
Intermedin (IMD) is a member of calcitonin/calcitonin gene-related peptide (CGRP) family together with adrenomedullin (AM) and amylin. It has a wide distribution in the central nervous system (CNS) especially in hypothalamic paraventricular nucleus (PVN). Cardiac sympathetic afferent reflex (CSAR) is enhanced in chronic heart failure (CHF) rats. The aim of this study is to determine the effect of IMD in the PVN on CSAR and its related mechanisms in CHF rats.
Methodology/Principal Findings
Rats were subjected to left descending coronary artery ligation to induce CHF or sham-operation (Sham). Renal sympathetic nerve activity (RSNA), mean arterial pressure (MAP) and heart rate (HR) were recorded. CSAR was evaluated by the RSNA and MAP responses to epicardial application of capsaicin. Acute experiments were carried out 8 weeks after coronary ligation or sham surgery under anesthesia. IMD and angiotensin II (Ang II) levels in the PVN were up-regulated in CHF rats. Bilateral PVN microinjection of IMD caused greater decreases in CSAR and the baseline RSNA and MAP in CHF rats than those in Sham rats. The decrease of CSAR caused by IMD was prevented by pretreatment with AM receptor antagonist AM22-52, but not CGRP receptor antagonist CGRP8-37. Ang II in the PVN significantly enhanced CSAR and superoxide anions level, which was inhibited by PVN pretreatment with IMD or tempol (a superoxide anions scavenger) in Sham and CHF rats.
Conclusion
IMD in the PVN inhibits CSAR via AM receptor, and attenuates the effects of Ang II on CSAR and superoxide anions level in CHF rats. PVN superoxide anions involve in the effect of IMD on attenuating Ang II-induced CSAR response.
Introduction
Cardiac sympathetic afferent reflex (CSAR), a sympatho-excitatory reflex, can be induced by stimulation of cardiac sympathetic afferents with exogenous or endogenous chemicals from myocardium during myocardial ischemia [1], [2]. Chronic heart failure (CHF) is accompanied by the enhanced sympathetic nerve activity (SNA) [3]–[5], and the enhanced CSAR partially contributes to the sympathetic activation in CHF rats [5]–[7]. Suppression of sympatho-excitation has been considered as a strategy in treating patients with CHF [8], [9], so the inhibition of CSAR may be a good target for decreasing sympathetic activation in CHF.
Intermedin (IMD) is widely distributed in peripheral organs and central nervous system (CNS) and belongs to calcitonin/calcitonin gene-related peptide (CGRP) family together with adrenomedullin (AM) and amylin [10]–[13]. IMD, CGRP and AM share the receptor system including calcitonin receptor-like receptor (CRLR) and receptor activity-modifying proteins (RAMPs). The CRLR/RAMP1 complex forms the CGRP receptor, whereas CRLR/RAMP2 or CRLR/RAMP3 complex forms the AM receptor [10]. In the present, no unique receptor has been identified for IMD, but it can non-selectively bind to all three CRLR/RAMP complexes. CRLR and RAMPs have been found in the paraventricular nucleus (PVN) of hypothalamus, and there is abundant IMD-like immunoreactivity in the PVN including both parvocellular and magnocellular cells [11], [14]–[16].
PVN is an integrative site in regulating sympathetic outflow and cardiovascular activity [17], [18], and it is also a component of central neurocircuitry of the CSAR [19], [20]. Previous studies have shown that PVN is involved in excessive sympathetic activation and enhanced CSAR in CHF [21]–[25]. Angiotensin II (Ang II), AT1 receptors and reactive oxygen species (ROS) in the PVN play an important role in the central modulation of CSAR, contribute to the pathogenesis of enhanced CSAR in CHF [23]–[25], and AT1 receptor mRNA antisense normalizes the enhanced CSAR in CHF rats [26]. IMD has different effects on SNA depending on where it is applied in the brain. Administration of IMD into the lateral cerebroventricle or NTS increased SNA [27]–[29], but we recently found that IMD in the PVN attenuated sympathetic activity [30] and CSAR (data not published) in hypertensive Rats. It is interesting to know the roles and mechanisms of IMD in the PVN in CSAR in CHF rats because of the importance of the PVN in the pathogenesis of CHF.
Ang II in the PVN promotes the enhanced CSAR in rats with CHF [25], [31], [32], and the decrease of endogenous Ang II by angiotensin converting enzyme inhibitor captopril in the PVN normalizes the enhanced CSAR in rats with CHF [24]. Ang II in the PVN increases superoxide anions level which mediates CSAR and the effect of Ang II in the PVN on CSAR in CHF rats [23]. Ang II exposure induces remarkable increases in the expression of endogenous IMD and its receptor components in H9c2 cell cultures [33]. IMD exerts an antihypertrophic effect caused by Ang II on neonatal cardiomyocytes by reducing the level of superoxide [34]. In many animal disease models, IMD has a protective effects on some tissues or cells via the inhibition of oxidative stress [35]–[40]. It is not known whether PVN superoxide anions involve in the effect of IMD on attenuating Ang II-induced CSAR response. Therefore, the present study was designed to determine whether IMD in the PVN inhibits SNA and CSAR and whether IMD in the PVN attenuates Ang II-induced CSAR response and its related mechanisms in CHF rats.
Methods
Animals
Experiments were carried out on male Sprague-Dawley rats weighing 300–400 g. The procedures were approved by the Experimental Animal Care and Use Committee of Nanjing Medical University and complied with the Guide for the Care and Use of Laboratory Animals (the 8th edition, 2011). The rats were maintained on a cycle of 12 h light and 12 h darkness in a temperature-controlled room and provided free access to laboratory chow and water.
CHF model
CHF was induced by coronary artery ligation method as previously described [5]. Briefly, rats were anesthetized with sodium pentobarbital (60 mg/kg, i.p.) and subjected to the ligation of the left anterior descending coronary artery or sham operation using sterile techniques. Sham-operated rats (Sham) received similar surgery except their coronary arteries were not ligated. The criterion for CHF was that left ventricle end-diastolic pressure (LVEDP) was higher than 12 mmHg. At the end of experiment, infarct size was expressed as a percentage of the left ventricle (LV) surface area [26].
General procedures of acute experiment
Acute experiment was carried out 8 weeks after surgery. Each rat was anaesthetized with intraperitoneal injection of urethane (800 mg/kg) and a-chloralose (40 mg/kg). A midline incision in the neck was made to expose trachea and carotid artery. The trachea was intubated and connected to a rodent ventilator (Model 683, Harvard Apparatus Inc, Holliston, MA, USA) for mechanical ventilation. A pressure transducer was used via a catheter in the right carotid artery for the measurement of mean arterial pressure (MAP). The MAP and renal sympathetic nerve activity (RSNA) were simultaneously recorded with a PowerLab data acquisition system (8/35, AD Instruments, Castle Hill, Australia). Supplemental doses of anesthetic agents were applied to maintain an adequate depth of anesthesia during the experiments.
RSNA recording
A retroperitoneal incision was made for isolation of the left renal sympathetic nerve. The nerve was cut distally to eliminate its afferent activity and placed on a pair of silver electrodes which were immersed in warm mineral oil. The RSNA was amplified with a four channel AC/DC differential amplifier (DP-304, Warner Instruments, Hamden, CT, USA) with a high pass filter at 100 Hz and a low pass filter at 3,000 Hz. The RSNA was integrated at a time constant of 100 ms. The background noise was determined as previously reported [41].
Evaluation of CSAR
The rats used to evaluate the CSAR were subjected to vagotomy and baroreceptor denervation [23]. To expose the heart, a limited left lateral thoracotomy was performed and then the pericardium was removed. The CSAR was induced by stimulating cardiac sympathetic afferents with a piece of filter paper (3 mm×3 mm) containing capsaicin (Cap, 1.0 nmol in 2.0 μl) on the non-infarct area of the left ventricle. The filter paper was removed 1 minute later, and the ventricular surface was rinsed three times with normal saline at 37°C. The CSAR was evaluated by the RSNA and MAP responses to the epicardial application of Cap [23].
PVN microinjection
The stereotaxic coordinates for the PVN were 1.8 mm caudal from bregma, 0.4 mm lateral to the midline and 7.9 mm ventral to the dorsal surface using a stereotaxic frame (Stoelting, Chicago, IL, USA). The bilateral PVN microinjections were carried out with two glass micropipettes (about 50 μm tip diameter) and completed within 1 min. Microinjection volume was 50 nl for each microinjection site. At the end of the experiment, same volume of Evans Blue (2%) was injected into the microinjection site for histological identification. Only the data from rats whose microinjection sites were within the boundaries of the PVN were used for analysis. Rats with microinjection sites outside the PVN or at the margin of PVN were excluded from data analysis. Total 14 rats in Sham and CHF groups were excluded from data analysis because the microinjection sites were not within the PVN (Figure 1). A representative coronal photograph of brain section which showed bilateral microinjection sites evaluated by 50 nl of Evans blue diffusion (upper panel) and a schematic representation of microinjection sites (lower panel) in the PVN of a rat were in Figure 1.
Figure 1. Microinjection sites in PVN region.

Upper panel, a representative photo of microinjection sites in the PVN evaluated by 50: paraventricular nucleus of hypothalamus; 3V: the third ventricle.
PVN sample preparation
The rat was anaesthetized with an overdose of pentobarbital (100 mg/kg, i.p.). The brain of the rat was quickly removed, frozen in liquid nitrogen and stored at −80 °C until being used. Brain tissue was cut into 450 μm coronal section within the levels from 1.5 mm to 2.0 mm caudal from bregma with frozen microtome (Leica CM1900-1-1, Wetzlar, Hessen, Germany).
Measurement of IMD and Ang II levels
PVN sample tissue was homogenized and the total protein in the homogenate supernatant was extracted and measured by using protein assay kit (BCA; Pierce). The IMD and Ang II levels in PVN tissue homogenate supernatant were measured using an enzyme-linked immunoassay kit (USCN Life Science Inc., USA). For both assays, the manufacturer's instructions were followed.
Measurement of superoxide anions
The superoxide anions level in the PVN was detected using the lucigenin-derived chemiluminescence method [23]. Dark-adapted lucigenin (5 μM) reacts with superoxide anions and results in photon emission which can be measured with a luminometer (20/20n, Turner, CA, USA) once every minute for 10 min. The values were expressed as the MLU per minute per milligram of protein.
Chemicals
IMD, AM22-52 and CGRP8-37 were obtained from Bachem (Hauptstrasse, Bubendorf, Switzerland), capsaicin, Ang II, tempol and lucigenin were purchased from Sigma Chemical (St. Louis, MO, USA).
Experimental design
Experiment 1
IMD and Ang II levels in the PVN were determined with Elisa method in both Sham and CHF rats (n = 6 for each group).
Experiment 2
Effects of PVN microinjection of IMD (30 pmol) on the baseline RSNA, MAP and HR responses were investigated in Sham and CHF rats (n = 6 for each group).
Experiment 3
Effects of IMD, CGRP8-37, AM22-52 and Ang II on the CSAR were respectively investigated in Sham rats and CHF rats. The PVN microinjection of saline, two doses of IMD (3 or 30 pmol), CGRP8-37 (0.2 nmol), AM22-52 (1 nmol) or Ang II (0.3 nmol) were carried out in 6 groups of Sham rats and 6 groups of CHF rats (n = 6 for each group). To exclude the possibility that the effects of IMD were caused by diffusion to other brain area, the effects of microinjection of IMD (30 pmol) into the anterior hypothalamic area which is adjacent to the PVN were determined in CHF rats (n = 3).
Experiment 4
Effects of PVN pretreatment with saline, CGRP receptor antagonist CGRP8-37 (0.2 nmol) and adrenomedullin (AM) receptor antagonist AM22-52 (1 nmol) on the CSAR response to the PVN microinjection of IMD (30 pmol) were investigated in 6 groups of Sham rats and 6 groups of CHF rats (n = 6 for each group).
Experiment 5
Effects of PVN pretreatment with saline, IMD (30 pmol) and tempol (20 nmol) on the CSAR response to the PVN microinjection of Ang II (0.3 nmol) were investigated in 4 groups of Sham rats and 4 groups of CHF rats (n = 6 for each group). The PVN microinjection of Ang II was carried out 28 or 8 min after IMD or tempol pretreatment respectively, and CSAR was determined 2 min after Ang II microinjection.
Experiment 6
Effects of the PVN microinjection of saline, IMD (30 pmol) and Ang II (0.3 nmol) on the superoxide anions level in the PVN were investigated in 3 groups of Sham rats and 3 groups of CHF rats (n = 6 for each group).
Experiment 7
Effects of PVN pretreatment with saline, IMD (30 pmol) and tempol (20 nmol) on the superoxide anions level response to the PVN microinjection of Ang II (0.3 nmol) were investigated in 4 groups of Sham rats and 4 groups of CHF rats (n = 6 for each group).
Statistics
Comparisons between two groups were made by Student's t test. One-way ANOVA followed by the Bonferroni test for post hoc analysis was used when multiple comparisons were made. The values were expressed as the mean±SE. P<0.05 was considered statistically significant.
Results
Anatomical and hemodynamic data
Mean infarct area was 33.4% of the LV in CHF rats, and there was no obvious infarct in Sham rats. Compared with Sham rats, heart weight, heart-to-body weight ratio and LVEDP were significantly increased in CHF rats, but LV peak systolic pressure (LVSP) and the maximum of the first differentiation of LV pressure (dp/dtmax) were markedly decreased in CHF rats (Table 1).
Table 1. Anatomic and hemodynamic data at the 8th week in Sham and CHF rats.
| Sham | CHF | |
| Body weight (g) | 367±10 | 359±8 |
| Heart weight (mg) | 1287±48 | 1546±54* |
| HW/BW (mg/g) | 3.37±0.21 | 4.30±0.29* |
| MAP (mm Hg) | 94.7±3.4 | 92.7±2.8 |
| HR (beats/min) | 388±7 | 396±6 |
| Infarct size (% LV area) | 0 | 33.4±2.9* |
| LVSP (mm Hg) | 131±6 | 112±5* |
| LVEDP(mm Hg) | −2.08±0.28 | 20.8±2.0* |
| LV dp/dtmax (mm Hg/s) | 3580±146 | 2062±102* |
HW: heart weight; BW: body weight; MAP: mean arterial pressure; LV: left ventricle; LVSP: left ventricle peak systolic pressure; LVEDP: left ventricle end-diastolic pressure; LVdp/dtmax: maximal rise rate of LV pressure. Values are mean±SE. *P<0.05 versus Sham. n = 7 for each group.
CSAR
In this study, the CSAR was induced by epicardial application of Cap and evaluated by the RSNA and MAP responses to Cap. The CSAR was significantly enhanced in CHF rats compared with Sham rats (Figure 2).
Figure 2. CSAR induced by epicardial application of capsaicin was evaluated by the RSNA and MAP responses to epicardial application of capsaicin (1 nmol).
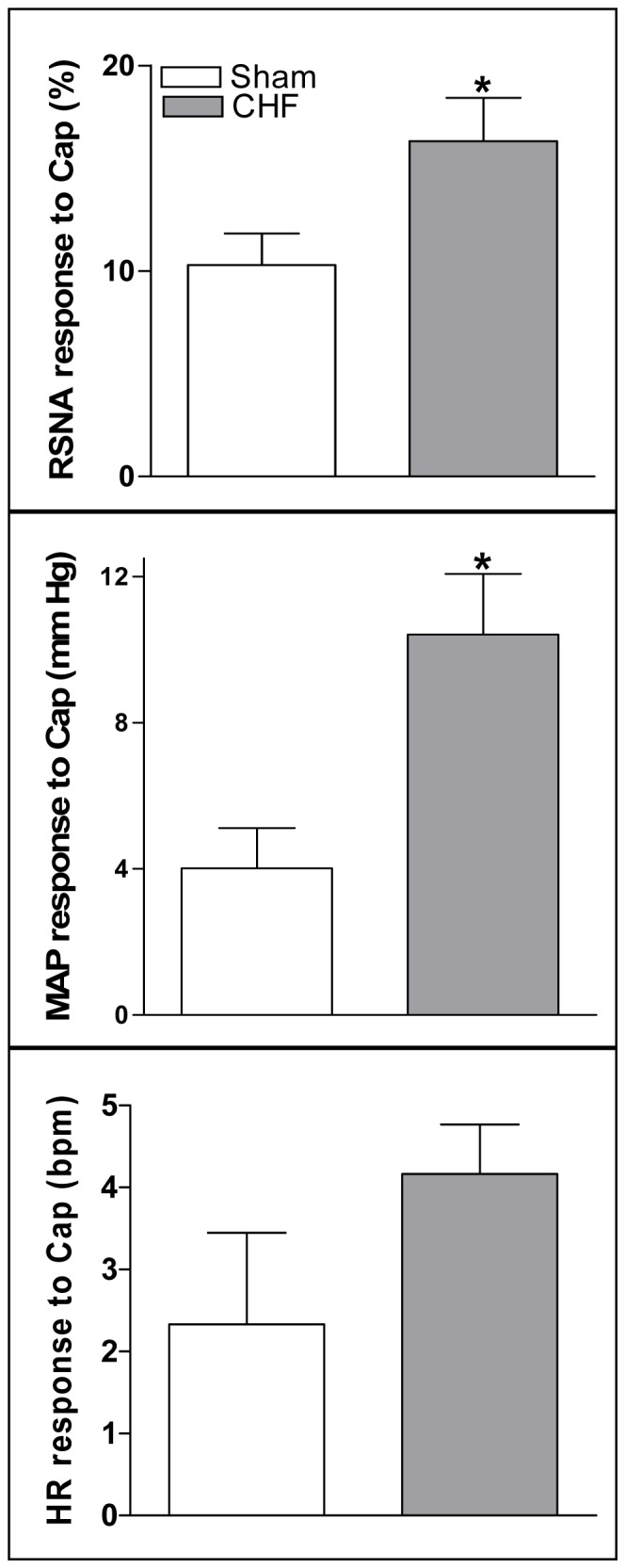
CSAR: cardiac sympathetic afferent reflex; RSNA: renal sympathetic nerve activity; MAP: mean arterial pressure; Cap: capsaicin. Values are mean±SE. *P<0.05 vs. Sham. n = 6 for each group.
IMD and Ang II levels in the PVN
IMD and Ang II levels in the PVN were much higher in CHF rats than Sham rats (Figure 3).
Figure 3. Levels of IMD and Ang II in the PVN in Sham rats and CHF rats.

Values are mean±SE. *P<0.05 vs. Sham. n = 6 for each group.
Effects of IMD on the baseline RSNA and MAP and CSAR
IMD (30 pmol) significantly decreased the baseline RSNA and MAP in CHF rats compared with Sham rats (Figure 4). The effects of IMD on the baseline RSNA and MAP peaked at about 30 min and lasted at least 60 min. Microinjection of IMD (3 or 30 pmol) into the PVN caused greater decrease in the CSAR in Sham and CHF rats (Figure 5). IMD in the PVN had no effects on the baseline HR and HR response to Cap in Sham and CHF rats (Figure2, 4). Microinjection of IMD (30 pmol) into the anterior hypothalamic area, which is adjacent to the PVN, had no significant effects on the CSAR and the baseline RSNA and MAP in CHF rats (data not shown). The representative recordings showed that PVN microinjection of IMD (30 pmol) decreased CSAR in CHF rats (Figure 6).
Figure 4. Effects of PVN microinjection of saline and IMD (30 pmol) on the baseline RSNA, MAP and HR in Sham rats and CHF rats.
RSNA: renal sympathetic nerve activity; MAP: mean arterial pressure, HR: heart rate. Values are mean±SE. *P<0.05 vs. Saline; #P<0.05 vs. Sham. n = 6 for each group.
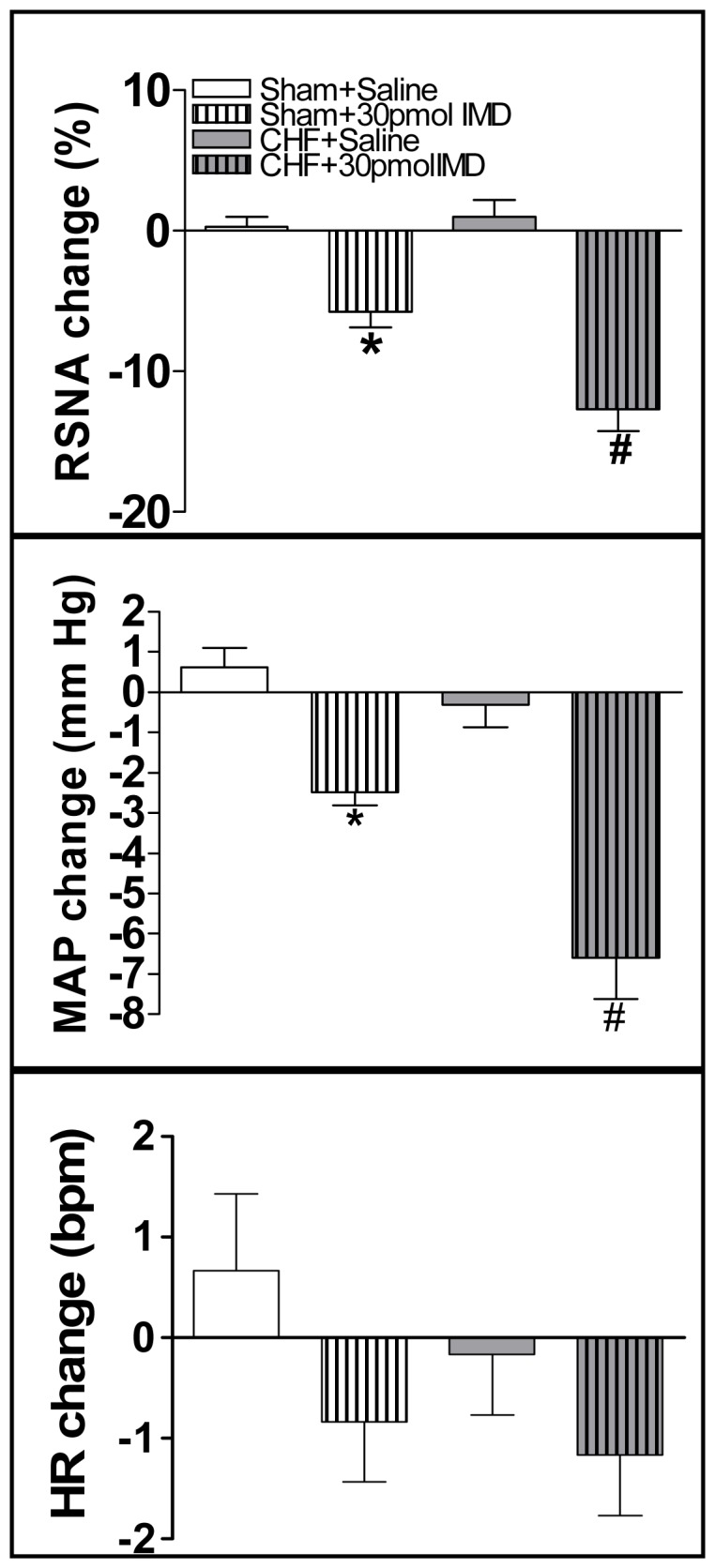
Figure 5. Effect of PVN microinjection of saline, two doses of IMD (3 or 30 pmol), CGRP receptor antagonist CGRP8-37 (0.2 nmol), AM receptor antagonist AM22-52 (1 nmol) or Ang II (0.3 nmol) on the CSAR.

CSAR: cardiac sympathetic afferent reflex; RSNA: renal sympathetic nerve activity; MAP: mean arterial pressure; HR: heart rate; Cap: capsaicin. Values are mean±SE. *P<0.05 vs. Saline; #P<0.05 vs. Sham. n = 6 for each group.
Figure 6. Tracing showing the effect of PVN microinjection of Saline in Sham and CHF rats and IMD (30 pmol) in CHF rats on CSAR.

CSAR was induced by epicardial application of capsaicin. The CSAR in CHF rats (B) was stronger than Sham rats (A). IMD attenuated the enhanced CSAR in CHF rats (C). RSNA: renal sympathetic nerve activity; ABP: arterial blood pressure; MAP: mean arterial pressure.
Effects of AM22-52 and CGRP8-37
AM22-52 microinjection into the PVN significantly increased the CSAR in both Sham and CHF rats but not CGRP8-37 (Figure 5). The reducement of CSAR response to IMD was inhibited by the pretreatment with AM22-52 (1 nmol) in the PVN in both Sham and CHF rats but not CGRP8-37 (Figure 7).
Figure 7. Effect of PVN pretreatment with saline, CGRP receptor antagonist CGRP8-37 (0.2 nmol) or AM receptor antagonist AM22-52 (1 nmol) on the CSAR response to the PVN microinjection of IMD (30 pmol).

CSAR: cardiac sympathetic afferent reflex; RSNA: renal sympathetic nerve activity; MAP: mean arterial pressure; Cap: capsaicin. Values are mean±SE. *P<0.05 vs. Saline+Saline; $P<0.05 vs. Saline+IMD; #P<0.05 vs. Sham. n = 6 for each group.
Effect of IMD on Ang II-induced increase in CSAR
Ang II (0.3 nmol) microinjection into the PVN significantly increased the CSAR in CHF rats compared with Sham rats (Figure 5). The augmented response of Ang II on CSAR was inhibited by the pretreatment with IMD (30 pmol) or tempol (20 nmol) in the PVN in both Sham and CHF rats (Figure 8).
Figure 8. Effect of PVN pretreatment with saline, IMD (30 pmol) or tempo (20 nmol) on the CSAR response to the PVN microinjection of Ang II (0.3 nmol).

CSAR: cardiac sympathetic afferent reflex; RSNA: renal sympathetic nerve activity; MAP: mean arterial pressure; Cap: capsaicin. Values are mean±SE. *P<0.05 vs. Saline+Saline; $P<0.05 vs. Saline+Ang II; #P<0.05 vs. Sham. n = 6 for each group.
Superoxide anions level in the PVN
Microinjection of IMD (30 pmol) into the PVN significantly decreased the superoxide anions level in the PVN in CHF rats, but not in Sham rats (Figure 9). Microinjection of Ang II (0.3 nmol) into the PVN significantly increased the superoxide anions level in the PVN in Sham and CHF rats (Figure 9). PVN pretreatment with IMD (30 pmol) or tempol (20 nmol) significantly decreased Ang II-induced increase in superoxide anions level in the PVN in both Sham and CHF rats (Figure 9).
Figure 9. Effect of PVN microinjection of saline, IMD (30 pmol) or Ang II (0.3 nmol) on the superoxide anions level in the PVN and effect of PVN pretreatment with saline, IMD (30 pmol) or tempo (20 nmol) on the superoxide anions level caused by Ang II (0.3 nmol) in the PVN.

Values are mean±SE. *P<0.05 vs. Saline; #P<0.05 vs. Sham. n = 6 for each group (upper panel). Values are mean±SE. *P<0.05 vs. Saline+Saline; $P <0.05 vs. Saline+Ang II; #P<0.05 vs. Sham. n = 6 for each group (lower panel).
Discussion
The primary findings in this study were that the levels of IMD and Ang II in the PVN were increased in CHF rats; IMD in the PVN significantly lowered the CSAR and the baseline RSNA and MAP in CHF rats than those in Sham rats; PVN pretreatment with IMD suppressed the enhanced CSAR and superoxide anions responses to Ang II in the PVN in Sham and CHF rats. AM receptor antagonist AM22-52 increased the CSAR and inhibited the effect of IMD on CSAR in Sham and CHF rats. These data indicate that exogenous IMD in the PVN inhibits CSAR via AM receptor in CHF rats, and attenuates the effect of Ang II in the PVN on CSAR, which may be through inhibiting Ang II-induced superoxide anions increase in the PVN.
IMD shares the receptor system consisting of CRLR and RAMPs with CGRP and AM [10]. In our recent study, all the receptor components were expressed in the PVN [30], CRLR, RAMP2 and RAMP3 protein expressions are apparent in the PVN, but the RAMP1 mRNA and protein expressions are low. In this study, the effect of IMD on CSAR was blocked with AM receptor antagonist AM22-52 but not CGRP receptor antagonist CGRP8-37, one possibility is that the level of RAMP1 expression remained small in CHF state and it was not enough to block the effect of IMD, the other is that IMD displayed its action on the CSAR through AM receptor not CGRP receptor. Furthermore, AM22-52 increased the CSAR in both sham rats and CHF rats, whereas CGRP8-37 had no significant effect. These results suggest that the AM receptor (CRLR/RAMP2 or CRLR/RAMP3) rather than CGRP receptor (CRLR/RAMP1) is involved in the CSAR in sham and CHF rats. The endogenous IMD in the PVN may have inhibitory effect on the CSAR via AM receptor in CHF. It is speculated that the increased IMD or AM receptor (CRLR/RAMP2 or CRLR/RAMP3) expression by various strategies in the PVN may be beneficial for the attenuation of CSAR. The higher level of IMD may be an important compensatory change in the PVN in CHF rats, which may contribute to the long-lasting sympathetic inhibition by attenuating the CSAR in CHF. In the future study, we should investigate whether endogenous AM could also inhibit the enhanced CSAR by binding with its receptor complex (CRLR/RAMP2 or CRLR/RAMP3) in the PVN in CHF rats and whether the effect of IMD on CSAR is associated with unidentfied receptor in the PVN.
Ang II and superoxide anions in the PVN causes exaggerated increases in SNA and CSAR in rats with CHF [23], [25], [42], and Ang II in the PVN promotes the increase of superoxide anions which can strengthen its effect on the CSAR in CHF rats [23]. AM-knockout mice showed Ang II/salt-loading-induced cardiovascular injury, all associated with enhanced ROS generation [43]. IMD is known to inhibit rat cardiac fibroblast activation induced by angiotensin II [44], and it exerts an antihypertrophic effect caused by Ang II on neonatal cardiomyocytes by reducing the level of superoxide [34]. These results suggest that IMD may have an inhibitory effect on Ang II-induced CSAR response by lowing superoxide anions level. In this study, the levels of Ang II and superoxide anions in the PVN were increased in CHF rats, IMD in the PVN decreased superoxide anions level and attenuated Ang II-induced CSAR response and superoxide anions increase in the PVN in CHF rats, which indicate that the inhibitory effect of IMD on CSAR is partially through inhibiting Ang II-induced superoxide anions.
The level of IMD is augmented in the presence of oxidative stress in hypertrophied cardiomyocytes [45]. IMD has a potential protective role against oxidative stress in human aortic endothelial cells [46] and rat cerebral endothelial cells [40]. It ameliorated vascular and renal injury in DOCA-salt hypertensive rats [39] and attenuated chemia/reperfusion-induced myocardial injury by inhibiting oxidative stress [37]. NAD(P)H oxidase in the PVN is a major source of the ROS in modulating the CSAR, and Ang II microinjecrion into the PVN significantly increases NAD(P)H oxidase activity which contributes to the effect of Ang II on CSAR [47]. The inhibition of NADPH oxidase involves in the effects of IMD on attenuating myocardial oxidative stress injury induced by ischemia/reperfusion [37] and ventricular myocyte hypertrophy induced by Ang II in neonatal rat [34]. These findings suggest that the inhibition of NADPH oxidase may be one of the mechanisms of IMD in decreasing superoxide anions in the PVN in CHF rats or attenuating Ang II-induced superoxide anions in the PVN, which needs to be explored in future study.
The enhanced CSAR is involved in the sympathetic over-activation in CHF, and superoxide anions and Ang II in the PVN promote the increases in SNA and CSAR in CHF rats. In the present study, we found that IMD and Ang II in the PVN were increased in CHF rats; IMD in the PVN inhibited SNA and attenuated the CSAR, and blockade of AM receptors in the PVN partially prevented its effect; PVN IMD decreased superoxide anions and attenuated the effects of Ang II on CSAR and superoxide anions level in the PVN in CHF rats. These results indicate that IMD inhibits SNA in CHF rats, which is through attenuating the enhanced CSAR response to superoxide anions and Ang II in the PVN. Activation of AM receptors in the PVN mediates the IMD's effects and IMD in the PVN is an important mechanism in attenuating CSAR and sympathetic activation in CHF.
Funding Statement
This work was supported by the funds from the National Natural Science Foundation of China (grant no. 81000106), the Jiangsu Higher Education Institutions of China (grant no. 10KJB310004), the Nanjing Medical University of China (grant no. 09JC013, JX 10131801056 and 09NJMUZ04), and the Collaborative Innovation Center for Cardiovascular Disease Translational Medicine. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Wang W, Ma R (2000) Cardiac sympathetic afferent reflexes in heart failure. Heart Fail Rev 5: 57–71. [DOI] [PubMed] [Google Scholar]
- 2. Gan XB, Duan YC, Xiong XQ, Li P, Cui BP, et al. (2011) Inhibition of cardiac sympathetic afferent reflex and sympathetic activity by baroreceptor and vagal afferent inputs in chronic heart failure. PLoS One 6: e25784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Cohn JN, Levine TB, Olivari MT, Garberg V, Lura D, et al. (1984) Plasma norepinephrine as a guide to prognosis in patients with chronic congestive heart failure. N Engl J Med 311: 819–823. [DOI] [PubMed] [Google Scholar]
- 4. Packer M, Bristow MR, Cohn JN, Colucci WS, Fowler MB, et al. (1996) The effect of carvedilol on morbidity and mortality in patients with chronic heart failure. U.S. Carvedilol Heart Failure Study Group. N Engl J Med 334: 1349–1355. [DOI] [PubMed] [Google Scholar]
- 5. Swedberg K, Eneroth P, Kjekshus J, Wilhelmsen L (1990) Hormones regulating cardiovascular function in patients with severe congestive heart failure and their relation to mortality. CONSENSUS Trial Study Group. Circulation 82: 1730–1736. [DOI] [PubMed] [Google Scholar]
- 6. Malliani A, Montano N (2002) Emerging excitatory role of cardiovascular sympathetic afferents in pathophysiological conditions. Hypertension 39: 63–68. [DOI] [PubMed] [Google Scholar]
- 7. Du YH, Chen AF (2007) A “love triangle” elicited by electrochemistry: complex interactions among cardiac sympathetic afferent, chemo-, and baroreflexes. J Appl Physiol 102: 9–10. [DOI] [PubMed] [Google Scholar]
- 8. Floras JS (2009) Sympathetic nervous system activation in human heart failure: clinical implications of an updated model. J Am Coll Cardiol 54: 375–385. [DOI] [PubMed] [Google Scholar]
- 9. Grassi G, Seravalle G, Quarti-Trevano F, Dell'oro R (2009) Sympathetic activation in congestive heart failure: evidence, consequences and therapeutic implications. Curr Vasc Pharmacol 7: 137–145. [DOI] [PubMed] [Google Scholar]
- 10. Roh J, Chang CL, Bhalla A, Klein C, Hsu SY (2004) Intermedin is a calcitonin/calcitonin gene-related peptide family peptide acting through the calcitonin receptor-like receptor/receptor activity-modifying protein receptor complexes. J Biol Chem 279: 7264–7274. [DOI] [PubMed] [Google Scholar]
- 11. Takahashi K, Kikuchi K, Maruyama Y, Urabe T, Nakajima K, et al. (2006) Immunocytochemical localization of adrenomedullin 2/intermedin-like immunoreactivity in human hypothalamus, heart and kidney. Peptides 27: 1383–1389. [DOI] [PubMed] [Google Scholar]
- 12. Fujisawa Y, Nagai Y, Miyatake A, Miura K, Shokoji T, et al. (2006) Roles of adrenomedullin 2 in regulating the cardiovascular and sympathetic nervous systems in conscious rats. Am J Physiol Heart Circ Physiol 290: H1120–127. [DOI] [PubMed] [Google Scholar]
- 13. Yuan Y, Wang X, Zeng Q, Wu HM, Qi YF, et al. (2012) Effects of continuous intermedin infusion on blood pressure and hemodynamic function in spontaneously hypertensive rats. J Geriatr Cardiol 9: 17–27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Oliver KR, Kane SA, Salvatore CA, Mallee JJ, Kinsey AM, et al. (2001) Cloning, characterization and central nervous system distribution of receptor activity modifying proteins in the rat. Eur J Neurosci 14: 618–628. [DOI] [PubMed] [Google Scholar]
- 15. Takahashi K, Morimoto R, Hirose T, Satoh F, Totsune K (2011) Adrenomedullin 2/intermedin in the hypothalamo-pituitary-adrenal axis. J Mol Neurosci 43: 182–192. [DOI] [PubMed] [Google Scholar]
- 16. Hashimoto H, Kitamura K, Kawasaki M, Saito T, Suzuki H, et al. (2008) Adrenomedullin 2/intermedin-like immunoreactivity in the hypothalamus and brainstem of rats. Auton Neurosci 139: 46–54. [DOI] [PubMed] [Google Scholar]
- 17. Coote JH (2005) A role for the paraventricular nucleus of the hypothalamus in the autonomic control of heart and kidney. Exp Physiol 90: 169–173. [DOI] [PubMed] [Google Scholar]
- 18. Li DP, Pan HL (2007) Glutamatergic inputs in the hypothalamic paraventricular nucleus maintain sympathetic vasomotor tone in hypertension. Hypertension 49: 916–925. [DOI] [PubMed] [Google Scholar]
- 19. Kang YM, Gao F, Li HH, Cardinale JP, Elks C, et al. (2011) NF-kappaB in the paraventricular nucleus modulates neurotransmitters and contributes to sympathoexcitation in heart failure. Basic Res Cardiol 106: 1087–1097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Zhong MK, Duan YC, Chen AD, Xu B, Gao XY, et al. (2008) Paraventricular nucleus is involved in the central pathway of cardiac sympathetic afferent reflex in rats. Exp Physiol 93: 746–753. [DOI] [PubMed] [Google Scholar]
- 21. Badoer E (2010) Role of the hypothalamic PVN in the regulation of renal sympathetic nerve activity and blood flow during hyperthermia and in heart failure. Am J Physiol Renal Physiol 298: F839–F846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Gao J, Zhong MK, Fan ZD, Yuan N, Zhou YB, et al. (2012) SOD1 overexpression in paraventricular nucleus improves post-infarct myocardial remodeling and ventricular function. Pflugers Arch 463: 297–307. [DOI] [PubMed] [Google Scholar]
- 23. Han Y, Shi Z, Zhang F, Yu Y, Zhong MK, et al. (2007) Reactive oxygen species in the paraventricular nucleus mediate the cardiac sympathetic afferent reflex in chronic heart failure rats. Eur J Heart Fail 9: 967–973. [DOI] [PubMed] [Google Scholar]
- 24. Wang HJ, Zhang F, Zhang Y, Gao XY, Wang W, et al. (2005) AT1 receptor in paraventricular nucleus mediates the enhanced cardiac sympathetic afferent reflex in rats with chronic heart failure. Auton Neurosci 121: 56–63. [DOI] [PubMed] [Google Scholar]
- 25. Zhu GQ, Gao L, Patel KP, Zucker IH, Wang W (2004a) ANG II in the paraventricular nucleus potentiates the cardiac sympathetic afferent reflex in rats with heart failure. J Appl Physiol 97: 1746–1754. [DOI] [PubMed] [Google Scholar]
- 26. Zhu GQ, Gao L, Li Y, Patel KP, Zucker IH, et al. (2004b) AT1 receptor mRNA antisense normalizes enhanced cardiac sympathetic afferent reflex in rats with chronic heart failure. Am J Physiol Heart Circ Physiol 287: H1828–H1835. [DOI] [PubMed] [Google Scholar]
- 27. Taylor MM, Bagley SL, Samson WK (2005) Intermedin/adrenomedullin-2 acts within central nervous system to elevate blood pressure and inhibit food and water intake. Am J Physiol Regul Integr Comp Physiol 288: R919–R927. [DOI] [PubMed] [Google Scholar]
- 28. Li P, Sun HJ, Han Y, Wang JJ, Zhang F, et al. (2013) Intermedin enhances sympathetic outflow via receptor-mediated cAMP/PKA signaling pathway in nucleus tractus solitarii of rats. Peptides 47: 1–6. [DOI] [PubMed] [Google Scholar]
- 29. Cui H, Kohsaka A, Waki H, Gouraud S, Nakamura T, et al. (2008) Adrenomedullin 2 microinjection into the nucleus tractus solitarius elevates arterial pressure and heart rate in rats. Auton Neurosci 142: 45–50. [DOI] [PubMed] [Google Scholar]
- 30. Zhou YB, Sun HJ, Chen D, Liu TY, Han Y, et al. (2014) Intermedin in Paraventricular Nucleus Attenuates Sympathetic Activity and Blood Pressure via Nitric Oxide in Hypertensive Rats. Hypertension 63: 330–337. [DOI] [PubMed] [Google Scholar]
- 31. Sharma NM, Zheng H, Mehta PP, Li YF, Patel KP (2011) Decreased nNOS in the PVN leads to increased sympathoexcitation in chronic heart failure: role for CAPON and Ang II. Cardiovasc Res 92: 348–357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Zhang S, Zhang F, Sun H, Zhou Y, Han Y (2012) Enhanced sympathetic activity and cardiac sympathetic afferent reflex in rats with heart failure induced by adriamycin. J Biomed Res 26: 425–431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Chen H, Wang X, Tong M, Wu D, Wu S, et al. (2013) Intermedin suppresses pressure overload cardiac hypertrophy through activation of autophagy. PLoS One 8: e64757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Liu K, Deng X, Gong L, Chen X, Wang S, et al. (2013) The effect of intermedin on angiotensin II and endothelin-1 induced ventricular myocyte hypertrophy in neonatal rat. Clin Lab 59: 589–596. [DOI] [PubMed] [Google Scholar]
- 35. Li H, Bian Y, Zhang N, Guo J, Wang C, et al. (2013) Intermedin protects against myocardial ischemia-reperfusion injury in diabetic rats. Cardiovasc Diabetol 12: 91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Qiao X, Li RS, Li H, Zhu GZ, Huang XG, et al. (2013) Intermedin protects against renal ischemia-reperfusion injury by inhibition of oxidative stress. Am J Physiol Renal Physiol 304: F112–F119. [DOI] [PubMed] [Google Scholar]
- 37. Zhao L, Peng DQ, Zhang J, Song JQ, Teng X, et al. (2012) Extracellular signal-regulated kinase 1/2 activation is involved in intermedin1-53 attenuating myocardial oxidative stress injury induced by ischemia/reperfusion. Peptides 33: 329–335. [DOI] [PubMed] [Google Scholar]
- 38. Bell D, Campbell M, Ferguson M, Sayers L, Donaghy L, et al. (2012) AM1-receptor-dependent protection by intermedin of human vascular and cardiac non-vascular cells from ischaemia-reperfusion injury. J. Physiol Mar 590: 1181–1197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Hagiwara M, Bledsoe G, Yang ZR, Smith RS Jr, Chao L, et al. 2008. Intermedin ameliorates vascular and renal injury by inhibition of oxidative stress. Am J Physiol Renal Physiol 295: F1735–F1743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Chen L, Kis B, Hashimoto H, Busija DW, Takei Y, et al. (2006) Adrenomedullin 2 protects rat cerebral endothelial cells from oxidative damage in vitro. Brain Res 1086: 42–49. [DOI] [PubMed] [Google Scholar]
- 41. Chen AD, Xiong XQ, Gan XB, Zhang F, Zhou YB, et al. (2012) Endothelin-1 in paraventricular nucleus modulates cardiac sympathetic afferent reflex and sympathetic activity in rats. PLoS One 7: e40748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Gao L, Wang W, Li YL, Schultz HD, Liu D, et al. (2004) Superoxide mediates sympathoexcitation in heart failure: roles of angiotensin II and NAD(P)H oxidase. Circ Res 95: 937–944. [DOI] [PubMed] [Google Scholar]
- 43. Ando K, Shimosawa T, Fujita T (2004) Adrenomedullin in vascular diseases. Curr Hypertens Rep 6: 55–59. [DOI] [PubMed] [Google Scholar]
- 44. Yang JH, Cai Y, Duan XH, Ma CG, Wang X, et al. (2009) Intermedin 1–53 inhibits rat cardiac fibroblast activation induced by angiotensin II. Regul Pept 158: 19–25. [DOI] [PubMed] [Google Scholar]
- 45. Bell D, Zhao Y, McCoy FP, Devine A, McDermott BJ (2008) Expression of the counter-regulatory peptide intermedin is augmented in the presence of oxidative stress in hypertrophied cardiomyocytes. Cell Physiol Biochem 21: 409–420. [DOI] [PubMed] [Google Scholar]
- 46. Pearson LJ, Yandle TG, Nicholls MG, Evans JJ (2009) Intermedin (adrenomedullin-2): a potential protective role in human aortic endothelial cells. Cell Physiol Biochem 23: 97–108. [DOI] [PubMed] [Google Scholar]
- 47. Zhang Y, Yu Y, Zhang F, Zhong MK, Shi Z, et al. (2006) NAD(P)H oxidase in paraventricular nucleus contributes to the effect of angiotensin II on cardiac sympathetic afferent reflex. Brain Res 1082: 132–141. [DOI] [PubMed] [Google Scholar]