

Abstract Abstract
Atlantic Forest, in the eastern coast of Brazil, is a hotspot of biodiversity of mammals, and Parque Estadual da Serra do Mar (PESM) is the largest continuous area of this biome. Here, we characterized the karyotype composition of the small mammals from Santa Virgínia, a region in the northern part of PESM. Specimens were collected from July 2008 to September 2009. We identified 17 species (13 rodents and 4 marsupials) from which 7 exhibited species-specific karyotypes, illustrating the importance of karyotype information in cytotaxonomy. We report for first time the karyotype of Monodelphis scalops (Thomas, 1888) and two new records for PESM: Akodon montensis Thomas, 1913 and Brucepattersonius soricinus Hershkovitz, 1998. Cytogenetic polymorphisms were detected for some species trapped in the area. Our results show the importance of Santa Virgínia / PESM in addressing studies for the conservation of small mammal wildlife in the Atlantic Forest.
Keywords: Atlantic Forest, conservation, cytotaxonomy, Monodelphis scalops
Introduction
The Atlantic Forest is the fourth biodiversity hotspot in the world (Myers et al. 2000, Ceballos and Ehrlich 2006, Carnaval et al. 2009). Geographical aspects combined with the large altitudinal and longitudinal ranges have favored the emergence of high endemism and species richness in this biome (Leal and De Gusmão Câmara 2003, Ribeiro et al. 2009). Nevertheless, the remaining forest represents only approximately 11% of the original extent, which highlights the biome as a priority for biodiversity conservation (Ribeiro et al. 2009).
The Parque Estadual da Serra do Mar (PESM), located in the state of São Paulo, Brazil was created in 1977, and is considered the largest remaining block of Atlantic Forest with 315.390 hectares (Instituto Florestal 2006).
Studies the mammal fauna of this park are scarce and the majority of the reports were presented in undergraduate theses and master’s dissertations, focusing on large mammals (Wang 2002, Norris et al. 2012). The most comprehensive article about small mammals from PESM was performed in Picinguaba (Northern of PESM) and reported morphology and karyotype information of 27 species belonging to the orders Didelphimorphia, Carnivora, and Rodentia (Pinheiro and Geise 2008).
According to Paglia et al. (2012), small mammals of the orders Rodentia and Didelphimorphia are important components of the Atlantic Forest mammal fauna, representing approximately 40% of the species. Morphological studies combined with cytogenetics and geographical distribution information allow the proper identification of taxa, particularly in cases of cryptic or morphologically similar species. Moreover, cytogenetic study can reveal genetic variability within and among individuals.
This study aims to characterize the karyotype composition and contribute to the identification of small rodents and marsupials from Santa Virgínia, since there is only one published study focusing on small mammals of this area. Data about geographical distribution of trapped species are also given.
Material and methods
Study area
Santa Virgínia (lat. 23°24.00'S to 23°17.00'S, long. 45°03.00'W to 45°11.00'W) is located in the Northern of PESM (Fig. 1) covering an area of 17,000 hectares (Instituto Florestal 2006), and altitudes ranging from 870 to 1,100 meters (Tabarelli and Mantovani 1999). The vegetation is defined as a dense montane humid forest (‘Floresta Ombrófila Densa Montana’) (Veloso et al. 1991) and the annual precipitation is about 2200 mm. The annual mean temperature varies from 18°C to 22°C.
Figure 1.
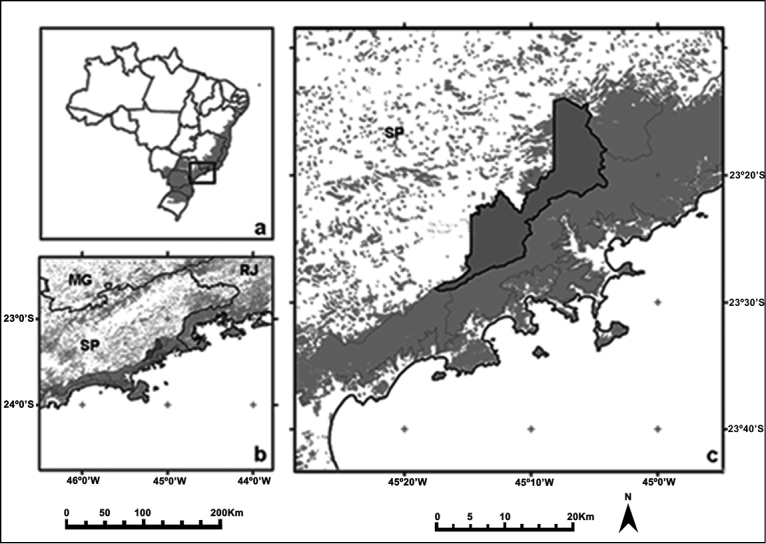
a Map of Brazil with original Atlantic Forest cover in grey and the region of Parque Estadual da Serra do Mar (PESM) indicated (square) b Parque Estadual da Serra do Mar (PESM) in grey c Santa Virgínia is highlighted (extracted and modified from Instituto Florestal 2006).
Field work
Small mammals were sampled by commercial live-traps (Sherman and Tomahawk-like traps) and pitfall-traps. In July 2008, a pilot experiment was performed from one to three nights, with a total sampling effort of 300 live-traps/night. From September 2008 to September 2009, field survey was carried out bimonthly during five consecutive nights. During this period, we set up six grids with 30 live-traps per grid and 12 transects of pitfall-traps. Live-traps were arranged in a 0.6 ha grids (60 × 100 m each) with 24 trap stations spaced every 20 meters. Each trap station received one Sherman of different size, randomly set (small, 25 × 7.5 × 9.5 cm; medium, 30 × 7.5 × 9.5 cm; large, 37.5 × 10 × 12 cm; H.B. Sherman Trap®, Inc., Tallahassee, Florida, USA). We also set randomly a Tomahawk-like trap (45 × 16 × 16 cm; Rosaminas Serviço Engenharia e Comércio Ltda. Piraúba, Minas Gerais, Brazil) at six trapping stations. Overall, we had 6300 live-trap/night.
The 12 transects of pitfall-traps were pairwise 30 meters apart, from November 2008 to September 2009. Each transect received four plastic buckets (60L, 40 cm top diameter, 35 cm bottom diameter, and 56 cm depth) buried with the rim at ground level, spaced every 10 meters each. The buckets on each line were connected with a 0.5 meters tall plastic drift fence that extended an additional 10 meters at each end, totaling 50 meters of fence. In total, we used 48 buckets, resulting in 1,440 pitfall-traps/night.
Different sizes and models of traps were used to optimize the sampling, aiming to reduce the selectivity based on body size and/or habits of the animals. Attractive baits (mashed bananas, peanut butter, bacon and corn meal) were placed in both kinds of traps. All traps were checked daily, preferably on the first hours in the morning.
Trapping and handling were carried out under ICMBio licence (number 14428-2) of Instituto Chico Mendes de Conservação da Biodiversidade.
Animals were euthanized according to the protocol of the “Animal experimentation ethics” (Carpenter et al. 1996) and under permission of Instituto Butantan Ethics Committee (242/05). The skins, skulls and partial skeletons were deposited in the Museu de Zoologia da Universidade de São Paulo (MZUSP) (still without MZUSP number), Museu Nacional da Universidade Federal do Rio de Janeiro (MN) and Coleção de Mamíferos da Universidade Federal do Espírito Santo (UFES) (Table 1).
Table 1.
A list of cytogenetically studied small mammals from Santa Virgínia, Parque Estadual da Serra do Mar, state of São Paulo, Brazil. N: number of individuals analyzed. Specimens voucher/Museum Field number: ROD and MARS - Laboratório de Ecologia e Evolução Instituto Butantan, Brazil; MN - Museu Nacional, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil; UFES - Coleção de Mamíferos da Universidade Federal do Espírito Santo, Brazil. 2n: diploid number, and FNa: number of autosomes arms. Morphologies: A=acrocentric; M=metacentric; SM=submetacentric; ST=subtelocentric. Grey cells correspond to species-specific karyotypes.
| ORDER Family Tribe Species |
N | Specimens voucher/ museum field number | Distribution | 2n | FNa | Autosome pairsa | Sex chromosomes | Variable cytogenetic characteristics | Karyotype reference | Figure No. |
|---|---|---|---|---|---|---|---|---|---|---|
|
ORDER RODENTIA Family Cricetidae Tribe Akodontini Akodon montensis |
3♀6♂ | ROD 3*, 6*, 11*, 28*, 29* UFES 2235-2237, 2239 | From Rio de Janeiro to Rio Grande do Sul and Minas Gerais, Brazil 1, 2 | 24, 24 (+ 1B) | 42 | 9 large to medium M/SM; 1 A; 1 small M | X: medium A Y: small A |
X chromosome polymorphism (enlarged short arm), 1 SM B-chromosome | Kasahara and Yonenaga-Yassuda (1982) | 3a |
| Blarinomys breviceps | 1♀ | UFES 2263 | Endemic of Atlantic Forest, Brazil1, 2 | 29 (+2B) | 50 | 11 medium M/SM 1 A Heteromorphic pair:1 M + 2 A |
X: large A | Heteromorphic pair, 2 M B-chromosomes | Ventura et al. (2012) | See Ventura et al. (2012) |
| Brucepattersonius soricinus | 1♀, 1♂ | MN 78955, 78956 | Southeastern Brazil, exclusively in Atlantic Forest1, 2, 3 | 52 | 52 | 24 medium to small A; 1 small SM | X: large ST Y: small A |
- | Bonvicino et al. (1998) | 3b–d |
| Thaptomys nigrita | 2♂ | ROD 2*, 4* | South Bahia to the north of Rio Grande do Sul, Brazil1, 2 | 52 | 52 | 24 medium to small A; 1 small SM |
X: large A Y: small SM |
- | Kasahara and Yonenaga-Yassuda (1984) | 5a |
| Tribe Oryzomyini Drymoreomys albimaculatus |
1♀, 1♂ | UFES 2271, 2272 | Endemic of Atlantic Forest, Brazil 4 | 62 | 62 | 29 medium to small A; 1 small M | X: large SM Y large SM, smaller than the X |
- | Suárez-Villota et al. (2013) | See Suárez-Villota et al. (2013) |
| Euryoryzomys russatus | 1♀, 7♂ | ROD 5*, 12*, 30* UFES 2242- 2244, 2265-2266 |
Coastal region of Brazil from Bahia to Rio Grande do Sul1, 2 | 80 | 86 | 35 A decreasing in size; 4 small M | X: large SM Y: small A or small ST |
Sex chromosomes polymorphisms | Andrades-Miranda et al. (2000) | 5b |
| Nectomys squamipes | 1♀ | UFES 2270 | Eastern Brazil 2 | 56 (+2B) | 56 | 26 A decreasing in size; 1 small M | X: large SM | 2 small SM B-chromosomes | Silva and Yonenaga-Yassuda (1998) | 4a |
| Oligoryzomys nigripes | 4♀, 4♂ | ROD 34*, UFES 2274-2280 | From South Bahia to Rio Grande do Sul, Brazil1, 2 | 62 | 80–82 | 11 M/SM decreasing in size; 19 A decreasing in size | X: large SM or large M Y: medium M or medium SM |
Pericentric inversions in pair 3, sex chromosomes polymorphisms | Paresque et al. (2007) | 4b |
| Sooretamys angouya | 1♀, 4♂ | UFES 2262, 2282-2285 | From Espírito Santo to Santa Catarina, Brazil2 | 58 | 60 | 26 A decreasing in size; 2 small M | X: large A Y: medium A |
- | Andrades-Miranda et al. (2000) | 5c |
| Tribe Phyllotini Calomys tener |
1♂ | UFES 2264 | Widespread in the state of São Paulo, Brazil1, 2 | 66 | 66 | 31 medium to small A; 1 M | X: large SM Y: medium A |
- | Mattevi et al. (2005) | 6a |
| Tribe Thomasomyini Rhipidomys itoan |
1♀ | UFES 2281 | PESM5, 6 | 44 | 50 | 17 A decreasing in size; 1 medium SM; 3 small M | X: large SM | - |
Pinheiro and Geise (2008); Costa et al. (2011) |
4c |
|
Incertae sedis Juliomys pictipes |
3♂ | UFES 2267-2269 | Minas Gerais to Rio Grande do Sul, Brazil1, 2 | 36 | 34 | 17 A decreasing in size | X: medium A Y: small A |
- | Bonvicino and Otazu (1999) | 6b–c |
|
Family Echimyidae Trinomys iheringi |
2♀, 1♂ | ROD 7*, 10*, UFES 2286 | West of Rio de Janeiro, São Paulo to north of Paraná, Brazil2, 7 | 60+1B, 60+4B | 116 | 29 M or SM decreasing in size | X: large SM Y: small SM |
1 or 4 dot-like B-chromosomes; Secondary constriction on pair 7 |
Yonenaga-Yassuda et al. (1985) | 4d |
|
ORDER DIDELPHIMORPHIA Family Didelphidae Marmosops incanus |
2♀, 1♂ | MARS 1*, 5*, 6* | Eastern Brazil8 | 14 | 24 | 6 SM decreasing in size | X: small SM Y: small A |
- | Carvalho et al. (2002) | 7a |
| Micoureus paraguayanus | 1♀, 1♂ | MARS 3*, 4* | Atlantic Forest; Eastern Brazil, until Rio Grande do Sul state8 | 14 | 20 | 4 M or SM 2 A |
X: medium A Y: medium A, smaller than X |
- | Pereira et al. (2008) | 7b |
| Monodelphis scalops | 1♂ | MN 78961 | Espírito Santo, Rio de Janeiro and São Paulo, Brazil8 | 18 | 30 | 4 SM 3 ST 1 A |
X: small ST Y: minute A |
- | Present study | 2 |
| Philander frenatus | 1♀, 1♂ | UFES 2287-2288 | From Bahia to Santa Catarina, Brazil8 | 22 | 20 | 10 A | X: medium A Y: small A |
Pereira et al. (2008) | 7c |
Geographic distribution according to: 1. Musser and Carleton (2005); 2. Bonvicino et al. (2008); 3. Bonvicino et al. (1998); 4. Percequillo et al. (2011); 5. De Vivo et al. (2011); 6. Pinheiro and Geise (2008); 7. Woods and Kilpatrick (2005); 8. Gardner (2005).
*Specimens voucher deposited in Museu de Zoologia da Universidade de São Paulo (MZUSP) without catalog number yet. ª Autosomal morphologies do not include Bs.
The nomenclature used in this work follows Gardner (2005), Musser and Carleton (2005), Weksler et al. (2006) and Percequillo et al. (2011). External morphologic traits of marsupials were compared with voucher specimens preserved at MZUSP.
Chromosome preparation
Metaphases were obtained from bone marrow and spleen after in vivo injection of a 0.1% colchicine solution (1mL/100g of weight). Cells were suspended in 0.075M KCl solution for 20 minutes at 37°C and fixed in three washes of methanol: acetic acid (3:1). GTG and CBG-banding were performed according to Seabright (1971) and Sumner (1972), respectively. At least 20 metaphases per individual were analyzed to define the diploid number (2n) and fundamental number of autosome arms (FNa). Chromosomes were measured using the program ImageJ version 1.46 (Rasband 2011) to establish the fundamental number, according to Levan et al. (1964). Karyotypes were set up according to the literature, when available.
Specimen identification was carried out through a comparison of our data with previous cytogenetic information, external morphological characteristics, and geographic distribution (see Table 1 references).
Results
A total of 706 small mammal specimens were captured (600 rodents and 106 marsupials) and 54 specimens were selected for chromosome preparations (46 rodents and 8 marsupials, Table 1).
On the whole, 13 species of rodents belonging to two families were cytogenetically analyzed (Table 1): Akodon montensis Thomas, 1913; Blarinomys breviceps (Winge, 1887); Brucepattersonius soricinus Hershkovitz, 1998; Thaptomys nigrita (Lichtenstein, 1829); Drymoreomys albimaculatus Percequillo, Weksler & Costa, 2011; Euryoryzomys russatus (Wagner, 1848); Nectomys squamipes (Brants, 1827); Oligoryzomys nigripes (Olfers, 1818); Sooretamys angouya (Fischer, 1814); Calomys tener (Winge, 1887); Rhipidomys itoan Costa, Geise, Pereira and Costa, 2011; Juliomys pictipes (Osgood, 1933) of family Cricetidae, and Trinomys iheringi (Thomas, 1911) of family Echimyidae.
Four marsupial species (Didelphimorphia) were karyotyped: Marmosops incanus (Lund, 1840); Micoureus paraguayanus (Tate, 1931); Monodelphis scalops (Thomas, 1888) and Philander frenatus (Olfers, 1818) (Table 1).
First cytogenetic information for Monodelphis scalops
Eight individuals were collected, although only one male had been cytogenetically studied. Morphological data and geographic distribution comparisons allow us to identify all as Monodelphis scalops. The morphological traits of these individuals are similar to voucher specimens of Monodelphis scalops preserved at MZUSP under catalogue numbers 1528, 30702, 30712 and 30757. This species has also been reported in São Paulo state, Brazil (Gardner 2005), agreeing to our collecting site (Fig. 1).
Here we present, for the first time, the karyotype of Monodelphis scalops. The karyotype of a male showed 2n=18, FNa=30. Pair 1 is a large submetacentric, pair 2 is a medium metacentric, pairs 3, 4 and 6 are medium subtelocentric, pair 5 is a medium acrocentric and pairs 7 and 8 are medium submetacentric. X chromosome is a small subtelocentric, and the Y is a minute acrocentric (Fig. 2). The short arm of pairs 4 and 6 are difficult to see depending on the condensation of the chromosome and so it was necessary to analyze and measure more than 30 metaphases to define their morphology.
Figure 2.
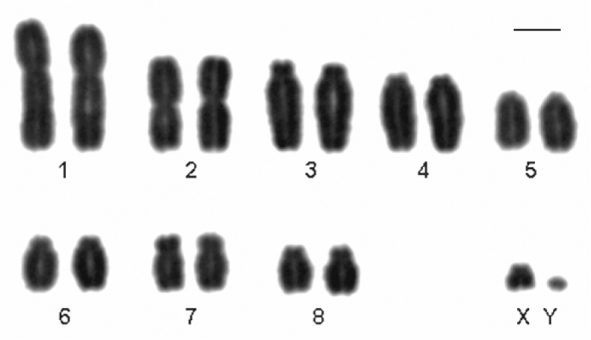
Conventional stained karyotype of Monodelphis scalops (2n=18, FNa=30, male). Bar = 10µm.
New records for PESM
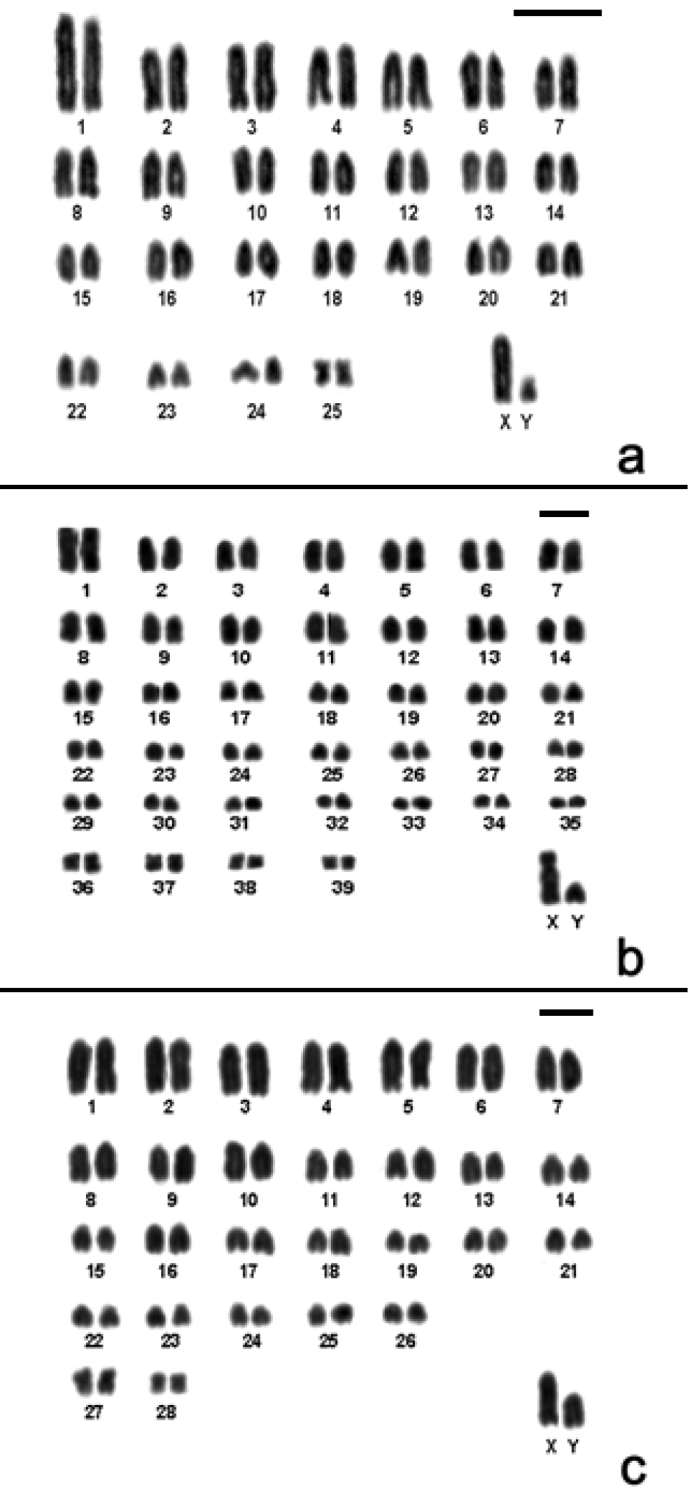
Cytogenetic data helped us to report for first time the presence of Akodon montensis, and Brucepattersonius soricinus in PESM. Cytogenetic information of these species are shown in Fig. 3, Table 1. Briefly, Akodon montensis showed 2n=24, 25 (24+1B), FNa=42 and one individual showed a heteromorphic X chromosome with an enlarged short arm. We also detected one small supernumerary submetacentric (B) in three out of nine individuals analyzed (Fig. 3a).
Figure 3.

Karyotypes of the new records for PESM. a Conventional stained karyotype of Akodon montensis (2n=24+1B, FNa=42, male) b Conventional stained karyotype of Brucepattersonius soricinus (2n=52, FNa=52, female). Inset: sex chromosomes of a male c CBG-banding pattern of Brucepattersonius soricinus (2n=52, FNa=52, female) d GTG-banding pattern of Brucepattersonius soricinus (2n=52, FNa=52, female). Bar = 10µm.
Brucepattersonius soricinus had 2n=52, FNa=52 (Fig. 3b) and this is the first time that banding-pattern is presented in this species. The CBG-banding pattern in the female specimen showed rather pronounced amount of pericentromeric heterochromatin in all chromosomes (Fig. 3c). GTG-banding allowed the identification of all autosomic pairs and X chromosomes (Fig. 3d).
Chromosomal variability and species-specific karyotypes
The remaining species studied in this work have already been recorded in PESM and their karyotypes are in accordance to the literature. Karyotype information of all species analyzed and the chromosomal variability found in this work is shown in Table 1 and Figs 4–7.
Figure 4.

CBG-banding pattern of Nectomys squamipes (2n=56 + 2B, FNa=56, female) b Conventional stained karyotype of Oligoryzomys nigripes (2n=62, FNa=80, male). Inset: different forms of pair 3: heteromorphic (3H) and homomorphic metacentric (3M) c Conventional stained karyotype of Rhipidomys itoan (2n=44, FNa=50, female) d Conventional stained karyotype of Trinomys iheringi (2n=60+4Bs, FNa=116, female). Inset: sex chromosomes of a male. Bar = 10µm.
Figure 5.
Conventional stained karyotypes: a Thaptomys nigrita (2n=52, FNa=52, male) b Euryoryzomys russatus (2n=80, FNa=86, male) c Sooretamys angouya (2n=58, FNa=60, male). Bar = 10µm.
Figure 6.
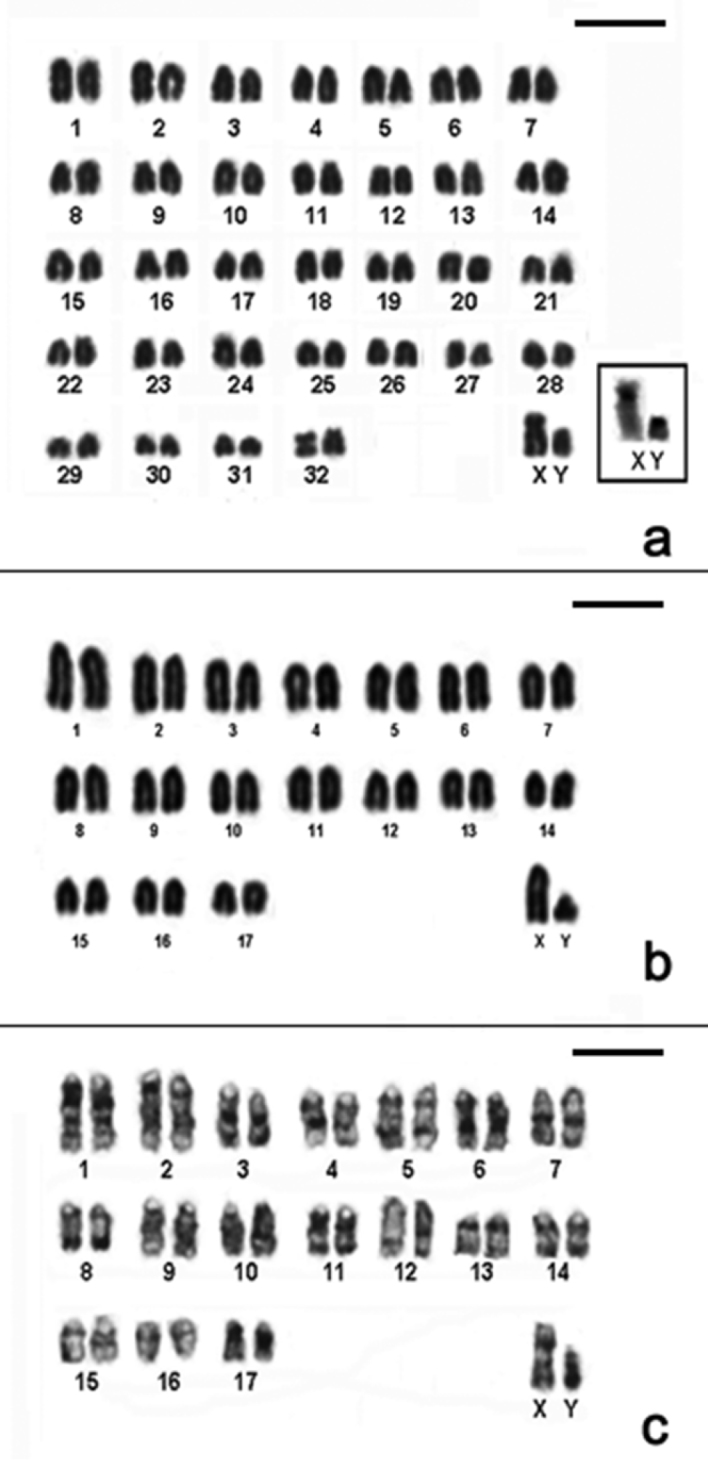
a Conventional stained karyotype of Calomys tener (2n=66, FNa=66, male). Inset: Sex chromosomes CBG-banded b Conventional stained karyotype of Juliomys pictipes (2n=36, FNa=36, male) c GTG-banding pattern of Juliomys pictipes (2n=36, FNa=36, male). Bar = 10µm.
Figure 7.
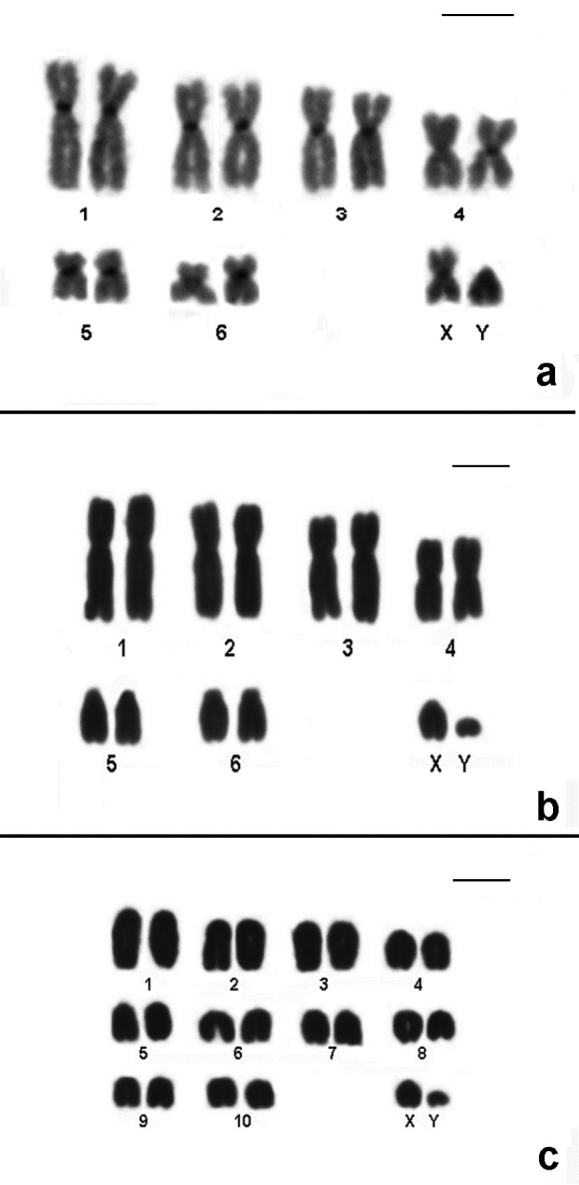
a CBG-banding pattern of Marmosops incanus (2n=14, FNa=24, male) b Conventional stained karyotype of Micoureus paraguayanus (2n=14, FNa=20, male) c Conventional stained karyotype of Philander frenatus (2n=22, FNa=20, male). Bar = 10µm.
Seven out of the 13 rodent species showed species-specific karyotypes: Akodon montensis, Drymoreomys albimaculatus, Oligoryzomys nigripes, Sooretamys angouya, Calomys tener, Juliomys pictipes and Trinomys iheringi (grey cells in Table 1). The identification of the remaining species (Blarinomys breviceps, Brucepattersonius soricinus, Thaptomys nigrita, Euryoryzomys russatus, Nectomys squamipes, and Rhipidomys itoan) required additional morphological and molecular investigation and geographic distribution information (Table 1).
Marsupials presented conserved diploid numbers of 14, 18 and 22 and were identified here by external morphological comparisons.
Discussion
Importance of cytogenetic study for Neotropical rodents
We proved the cytogenetic analyses as a taxonomic tool, since 7 out of 13 rodent species present species-specific karyotypes (53.8%). Besides, we identified 94% of all species, when cytogenetic data were combined with information of external morphology and geographical distribution (Table 1).
Cryptic species are relatively common in some Neotropical rodent groups and cytogenetic information was indispensable for identifying such species. For instance, Akodon montensis is morphologically indistinguishable from Akodon cursor (Winge, 1887) and both species occur in sympatry in the Atlantic Forest (Christoff et al. 2000). In addition, the occurrence of Akodon cursor previously recorded in Santa Virgínia/PESM (Instituto Florestal 2006) was doubtful till this study, as we proved the occurrence of Akodon montensis by karyotypic analysis.
Another cryptic species case occurs in the genus Thaptomys. Thaptomys sp. (2n=50) and Thaptomys nigrita (2n=52) are morphologically identical, so the karyotypes are the diagnostic information to distinguish both species (Ventura et al. 2004, 2010).
By contrast, Thaptomys nigrita and Brucepattersonius soricinus present very similar karyotypes (2n=52, FNa=52) however their identification can be safely done at the level of genera by external morphological characters. An accurate observation on the karyotypes of Brucepattersonius soricinus and Thaptomys nigrita showed that the pair 1 of Thaptomys nigrita is the largest of the chromosome set (Fig. 5a) meanwhile Brucepattersonius soricinus has the pair 1 similar in size to the others of the set (Figs 3b–d). We also noticed differences regarding sex chromosome morphologies of both species (Table 1). This feature could be a diagnostic tool to differentiate each karyotype, but additional cytogenetic studies (including comparative and molecular cytogenetic data) are needed to support these first observations.
Blarinomys breviceps presents a peculiar karyotype and it could not be considered species-specific due to the great variability in 2n and FNa (Geise et al. 2008, Ventura et al. 2012). Moreover, Ventura et al. (2012) suggested the existence of more species for the monotypic genus Blarinomys in Atlantic Forest since molecular phylogenetic analyses showed two geographically distinct lineages.
Euryoryzomys russatus does not have species-specific karyotype also. Euryoryzomys emmonsae Musser, Carleton, Brothers and Gardner, 1998, and Euryoryzomys nitidus (Thomas, 1884) share the same 2n=80, NFa=86 (Bonvicino and Geise 2006). However, when cytogenetic information is combined with morphologic and geographic distribution data, Euryoryzomys russatus can be confirmed.
Concerning Nectomys squamipes, it is not possible to affirm that this species possess species-specific karyotype with classical cytogenetic data because, when compared to Holochilus brasiliensis (Desmarest 1819), both karyotypes are identical (Yonenaga-Yassuda et al. 1987). Nevertheless, the association of cytogenetic, geographic distribution and external morphological characters allows the recognition of Nectomys squamipes as occurring at PESM (Bonvicino et al. 2008). Nectomys squamipes was considered for years as a carrier of two basic distinct karyotypes: 2n=56 (1 to 3Bs) and 2n=52 (1 to 3Bs), and only after crossings in laboratory, Bonvicino et al. (1996) noticed that two different species could be diagnosed - Nectomys squamipes (2n=56) and Nectomys rattus (Pelzeln, 1883), (2n=52).
The karyotype of Rhipidomys itoan presented here (2n=44, FNa=50 Fig. 4c) is the same one as described by Zanchin et al. (1992) and Silva and Yonenaga-Yassuda (1999). Pinheiro and Geise (2008) also found an identical karyotype for a species referred as Rhipidomys sp., trapped in Picinguaba (PESM), and De Vivo et al. (2011) reported an undescribed species of Rhipidomys that occurs at the Parque Estadual da Serra do Mar. Recently, two new species from Atlantic Forest were described: Rhipidomys tribei Costa, Geise, Pereira and Costa, 2011 and Rhipidomys itoan; and the latter presented 2n=44, FNa=48, 49, 50 (Costa et al. 2011). Santa Virgínia is embedded in the geographical distribution described for this species and molecular analyzes confirmed that this sample belongs to Rhipidomys itoan species. Nevertheless, we do not consider this karyotype species-specific.
Finally, cytogenetic analysis was useful in identifying Thaptomys iheringi as two species – Thaptomys iheringi and Thaptomys dimidiatus (Günther, 1876) - occur in Atlantic Forest. Despite the regular chromosome set of Thaptomys iheringi (not considering B chromosomes) is identical to the one described for the species Thaptomys dimidiatus (2n=60, FNa=116) by Pessoa et al. (2005), the presence of at least one B and the morphology of Y chromosome in Thaptomys iheringi represent good characters to diagnose the species.
Chromosome variations
Mammals have remarkable diversity in species karyotypes, and rodents exhibit noteworthy variability of diploid chromosome number (O’Brien et al. 2006, Romanenko et al. 2012). For instance, in this work, diploid numbers of rodents ranged from 24 in Akodon montensis to 80 in Euryoryzomys russatus.
The chromosome variation observed here is due to the presence of supernumerary chromosomes (B chromosomes), sex chromosome heteromorphism and/or polymorphism, as well as autosomal polymorphisms. This chromosome variability does not cause a problem in characterizing the species, except in the case of Thaptomys iheringi, in which the presence of at least one B chromosome is sufficient to confirm its identity.
Structural rearrangements may explain much of the observed karyotype diversity in rodents. In this regard, Robertsonian fusions/fissions (whole-arm translocations) and pericentric inversions, have long been considered the predominant rearrangements in natural populations of rodents (Patton and Sherwood 1983). Nevertheless, studies with more refined techniques such as fluorescent in situ hybridization and chromosome painting demonstrate that tandem fusions, reciprocal translocations, and paracentric inversions are much more common than previously thought (Hass et al. 2008, Ventura et al. 2009, Romanenko et al. 2012).
Our data showed two species with pericentric inversion rearrangements, Oligoryzomys nigripes and Rhipidomys itoan. Oligoryzomys nigripes showed variation in autosomal pair 3 (Fig. 4b) but this rearrangement had also been reported in pairs 2, 4 and 8, which places this species as one of the most polymorphic within Neotropical rodents (Paresque et al. 2007). The genus Rhipidomys frequently shows 2n=44, except for the 2n=50 reported by Silva and Yonenaga-Yassuda (1999) from Amazonas, in contrast with differences in the FNa (Zanchin et al. 1992, Costa et al. 2011). The variation of FNa, which represents the commonest chromosome change observed for the genus, may be a consequence of pericentric inversion events.
Karyotype diversity is also enhanced in mammals due to the presence of B chromosomes. B chromosomes are extra elements found in the karyotypes of many eukaryotic species. Their functions and molecular composition remain obscure but, apparently in mammals, these chromosomes neither promote phenotypic alterations nor affect fitness of individuals (Jones and Rees 1982, Trifonov et al. 2010). B chromosomes are known in nine Brazilian rodent species (Silva and Yonenaga-Yassuda 2004, Ventura et al. 2012). Herein, we found B chromosomes in four out of 13 species of rodents (30,76%, i.e. almost a third of the total): Akodon montensis, Brucepattersonius breviceps, Nectomys squamipes and Thaptomys iheringi. Silva and Yonenaga-Yassuda (2004) found B chromosomes in Sooretamys angouya (referred at that time as Oryzomys angouya), however, in our sample, B chromosomes were not observed for this species (Fig. 5c).
Sex chromosome heteromorphisms/polymorphisms were found in Akodon montensis and Oligoryzomys nigripes, and the variation is due to addition/deletion of constitutive heterochromatin, as described by Kasahara and Yonenaga-Yassuda (1982) and Paresque et al. (2007), respectively.
Marsupials
Cytogenetic data exposed three diploid numbers for the family Didelphidae: 2n=14, 18 and 22 (Reig et al. 1977, Carvalho et al. 2002). As the karyotypes of American marsupials are conserved, cytogenetic analyses cannot be considered as a diagnostic tool to identify species. However, differences in banding patterns could help in the characterization of some taxa, for instance, Marmosops incanus (Svartman 2009).
In the present paper we report for the first time the karyotype of Monodelphis scalops which is similar to the one described for Monodelphis kunsi Pine, 1975 and Monodelphis brevicauda (Erxleben, 1777) by Carvalho et al. (2002), except for the morphology of the sex chromosomes (Fig. 2). Besides, Monodelphis scalops karyotype differs from Monodelphis rubida (Thomas, 1899) (2n=18, FNa=32) (Pereira et al. 2008) due to the presence of one acrocentric pair (#5) instead of a biarmed pair (Fig. 2).
Final considerations
Our species list is an evidence of the limited knowledge of small mammals in PESM since the karyotype of Monodelphis scalops is reported for the first time and Akodon montensis and Brucepattersonius soricinus are new records for the park. According to De Vivo et al. (2011), it is important to increase samples in areas of dense humid forest since these areas are poorly surveyed. The number of species collected during the period of 14 months in Santa Virgínia should be considered highly representative, and this effort brought to light new findings. This includes the specimen of Blarinomys breviceps herein mentioned which was added to a larger sample with animals collected from different localities of Brazil and the diploid numbers ranged from 28 to 52 (Ventura et al. 2012), as well as Drymoreomys albimaculatus which was studied by Suárez-Villota et al. (2013).
The improvements to the list of mammals of PESM could be attributed to different methods of capture (live and pitfall traps) to enhance the success of trapping in different habitats. The multidisciplinary approach employed is also evidently important in some cases as presented above. Additionally, data on diversity and geographical distribution of species are essential to reach conservation strategies, and the significance of Santa Virgínia / PESM in the preservation of the Neotropical fauna becomes more clear.
Acknowledgments
The authors are greatly indebted to the head of Santa Virgínia (João Paulo Villani), Dr. Mauro Galetti for supporting CLN field work, Adelita Santiago for technical support, and Elkin Suárez-Villota for critical review of the early version of the manuscript. This work was supported by the Program “Jovem Pesquisador” FAPESP for MJJS (Proc. 05/04557-3) and BIOTA-FAPESP for MG (Proc. 07/03392-6). Grants for CBDN (FAPESP Proc. 08/00493-9 and 10/03432-0), CLN (CNPq Proc. 131891/2008-1), and JFV (101.822/2011 FAPERJ) are also acknowledged. We thank the anonymous reviewers and the editor for their helpful comments.
Citation
Di-Nizo CB, Neves CL, Vilela JF, Silva MJJ (2014) New karyologycal data and cytotaxonomic considerations on small mammals from Santa Virgínia (Parque Estadual da Serra do Mar, Atlantic Forest, Brazil). Comparative Cytogenetics 8(1): 11–30. doi: 10.3897/CompCytogen.v8i1.6430
Supplementary materials
References
- Andrades-Miranda J, Zanchin NIT, Oliveira LFB, Langguth AR. (2000) Cytogenetic studies in nine taxa of the genus Oryzomys (Rodentia, Sigmodontinae) from Brazil. Mammalia 65: 461–472. doi: 10.1515/mamm.2001.65.4.461
- Bonvicino CR, D’Andrea PS, Cerqueira R, Seuánez HN. (1996) The chromosomes of Nectomys (Rodentia, Cricetidae) with 2n= 52, 2n= 56, and interspecific hybrids (2n= 54). Cytogenetic Genome Research 73: 190-193 doi: 10.1159/000134337 [DOI] [PubMed] [Google Scholar]
- Bonvicino CR, Geise L. (2006) Relevância dos estudos cariológicos na taxonomia de alguns gêneros de Oryzomyini (Rodentia, Sigmodontinae). In: Freitas TRO, Vieira E, Pacheco S, Christoff AU. (Eds). Mamíferos do Brasil: genética, sistemática, ecologia e conservação.Sociedade Brasileira de Genética, Ribeirão Preto: 27-37
- Bonvicino CR, Oliveira JA, D’Andrea PS. (2008) Guia dos roedores do Brasil, com chaves para gêneros baseada em caracteres externos. Centro Pan-Americano de Febre Aftosa – OPAS/OMS, Rio de Janeiro, 120 pp. [Google Scholar]
- Bonvicino CR, Otazu I. (1999) The Wilfredomys pictipes (Rodentia, Sigmodontinae) karyotype with comments on the karyosystematics of Brazilian Thomasomyini. Acta Theriologica 44: 329–332.
- Bonvicino CR, Penna-firme V, Seuanez HN. (1998) The karyotype of Brucepattersonius griserufescens Hershkovitz, 1998 (Rodentia, Sigmodontinae) with comments on distribution and taxonomy. Zeitschrift für Säugetierkunde 63: 329-335 [Google Scholar]
- Carnaval AC, Hickerson MJ, Haddad CF, Rodrigues MT, Moritz C. (2009) Stability predicts genetic diversity in the Brazilian Atlantic forest hotspot. Science 323: 785-789 doi: 10.1126/science.1166955 [DOI] [PubMed] [Google Scholar]
- Carpenter J, Mashima T, Rupiper D. (1996) Exotic Animal Formulary. Greystone Publicashions, Philadelphia, PA, 310 pp. [Google Scholar]
- Carvalho BA, Oliveira LFB, Nunes AP, Mattevi MS. (2002) Karyotypes of nineteen marsupial species from Brazil. Journal of Mammalogy 83: 58-70 doi: 10.1644/1545-1542(2002)083<0058:KONMSF>2.0.CO;2 [Google Scholar]
- Ceballos G, Ehrlich PR. (2006) Global mammal distributions, biodiversity hotspots, and conservation. Proceedings of the National Academy of Sciences 103: 19374-19379 doi: 10.1073/pnas.0609334103 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Christoff AU, Fagundes V, Sbalqueiro IJ, Mattevi MS, Yonenaga-Yassuda Y. (2000) Description of a new species of Akodon (Rodentia: Sigmodontinae) from southern Brazil. Journal of Mammalogy 81: 838-851 doi: 10.1644/1545-1542(2000)081<0838:DOANSO>2.3.CO;2 [Google Scholar]
- Costa BMA, Geise L, Pereira LG, Costa LP. (2011) Phylogeography of Rhipidomys (Rodentia: Cricetidae: Sigmodontinae) and description of two new species from southeastern Brazil. Journal of Mammalogy 92: 945-962 doi: 10.1644/10-MAMM-A-249.1 [Google Scholar]
- De Vivo M, Carmignotto AP, Gregorin R, Hingst-Zaher E, Iack-Ximenes GE, Miretzki M, Percequillo AR, Rollo M, Rossi RV, Taddei VA. (2011) Checklist dos mamíferos do Estado de São Paulo, Brasil. Biota Neotropica 11. http://www.biotaneotropica.org.br/v11n1a/en/abstract?inventory+bn0071101a2011
- Gardner A. (2005) Order Didelphimorphia. In: Wilson D, Reeder D. (Eds). Mammal species of the World: a taxonomic and geographic reference.Baltimore, Maryland: John Hopkins University Press: 3-18
- Geise L, Bergallo HG, Esbérard CEL, Rocha CFD, Van Sluys M. (2008) The karyotype of Blarinomys breviceps (Mammalia: Rodentia: Cricetidae) with comments on its morphology and some ecological notes. Zootaxa 1907: 47-60 [Google Scholar]
- Hass I, Sbalqueiro IJ, Müller S. (2008) Chromosomal phylogeny of four Akodontini species (Rodentia, Cricetidae) from Southern Brazil established by Zoo-FISH using Mus musculus (Muridae) painting probes. Chromosome Research 16: 75-88 doi: 10.1007/s10577-007-1211-5 [DOI] [PubMed] [Google Scholar]
- Instituto Florestal (2006) Plano de manejo do Parque Estadual da Serra do Mar. São Paulo, 63 pp.
- Jones RN, Rees H. (1982) B chromosomes. New York: Academic Press, 266 pp.
- Kasahara S, Yonenaga-Yassuda Y. (1982) Chromosomal variability in Akodon sp. (Rodentia, Cricetidae). Cytologia 47: 317-324 [DOI] [PubMed] [Google Scholar]
- Kasahara S, Yonenaga-Yassuda Y. (1984) A progress report of cytogenetic data on Brazilian rodents. Revista Brasileira de Genética 7: 509-533 doi: 10.1508/cytologia.47.317 [Google Scholar]
- Leal CG, de Gusmão Câmara I. (2003) The Atlantic Forest of South America: biodiversity status, threats, and outlook. Island Press, 953–955. doi: 10.1007/s10980-005-1788-z
- Levan A, Fredga K, Sandberg AA. (1964) Nomenclature for centromeric position on chromosomes. Hereditas 52: 201-220 doi: 10.1111/j.1601-5223.1964.tb01953.x [Google Scholar]
- Mattevi MS, Haag T, Oliveira LFB, Langguth AR. (2005) Chromosome characterization of Brazilian species of Calomys Waterhouse, 1837 from Amazon, Cerrado and Pampas domains (Rodentia, Sigmodontinae). Arquivos do Museu Nacional 63: 175-181 [Google Scholar]
- Musser G, Carleton M. (2005) Superfamily Muroidea. In: Wilson D, Reeder D (Eds) Mammal species of the world: a taxonomic and geographic reference Baltimore, Maryland: Johns Hopkins University Press, 894–1531.
- Myers N, Mittermeier RA, Mittermeier CG, Fonseca GA, Kent J. (2000) Biodiversity hotspots for conservation priorities. Nature 403: 853-858 doi: 10.1038/35002501 [DOI] [PubMed] [Google Scholar]
- Norris D, Ramírez JM, Zacchi C, Galetti M. (2012) A Survey of mid and large bodied mammals in Núcleo Caraguatatuba, Serra do Mar State Park, Brazil. Biota Neotropica 12: 127-133 doi: 10.1590/S1676-06032012000200013 [Google Scholar]
- O’Brien SJ, Menninger JC, Nash WG. (2006) Atlas of Mammalian Chromosomes. John Wiley & Sons, 679 pp. doi: 10.1002/0471779059
- Paglia AP, Fonseca GAB, Rylands AB, Herrmann G, Aguiar LMS, Chiarello AG, Leite YLR, Costa LP, Siciliano S, Kierulff MCM, Mendes SL, Tavares VC, Mittermeier RA, Patton JL. (2012) Lista Anotada dos Mamíferos do Brasil/Annotated Checklist of Brazilian Mammals. In: Occasional Papers in Conservation Biology. Arlington, 76 pp.
- Paresque R, Silva MJJ, Yonenaga-Yassuda Y, Fagundes V. (2007) Karyological geographic variation of Oligoryzomys nigripes Olfers, 1818 (Rodentia, Cricetidae) from Brazil. Genetics and Molecular Biology 30: 43-53 doi: 10.1590/S1415-47572007000100010 [Google Scholar]
- Patton JL, Sherwood SW. (1983) Chromosome evolution and speciation in rodents. Annual Review of Ecology and Systematics 14: 139-158 doi: 10.1146/annurev.es.14.110183.001035 [Google Scholar]
- Percequillo AR, Weksler M, Costa LP. (2011) A new genus and species of rodent from the Brazilian Atlantic Forest (Rodentia: Cricetidae: Sigmodontinae: Oryzomyini), with comments on oryzomyine biogeography. Zoological Journal of the Linnean Society 161: 357-390 doi: 10.1111/j.1096-3642.2010.00643.x [Google Scholar]
- Pereira NP, Ventura K, Silva Júnior MC, Silva DM, Yonenaga-Yassuda Y, Pellegrino K. (2008) Karyotype characterization and nucleolar organizer regions of marsupial species (Didelphidae) from areas of Cerrado and Atlantic Forest in Brazil. Genetics and Molecular Biology 31: 887-892 doi: 10.1590/S1415-47572008005000012 [Google Scholar]
- Pessoa LM, Corrêa M, Bittencourt E, Reis S. (2005) Chromosomal characterization of taxa of the genus Trinomys Thomas, 1921 (Rodentia, Echimyidae) in the states of Rio de Janeiro and São Paulo. Arquivos do Museu Nacional 63: 161-168 [Google Scholar]
- Pinheiro PS, Geise L. (2008) Non-volant mammals of Picinguaba, Ubatuba, state of São Paulo, southeastern Brazil. Boletim do Museu Biológico Mello Leitão 23: 51-59 [Google Scholar]
- Rasband W. (2011) ImageJ. 1.46r ed, National Institute of Health, USA. http://imagej.nih.gov/ij/
- Reig OA, Gardner AL, Bianchi NO, Patton JL. (1977) The chromosomes of the Didelphidae (Marsupialia) and their evolutionary significance. Biological Journal of the Linnean Society 9: 191-216 doi: 10.1111/j.1095-8312.1977.tb00265.x [Google Scholar]
- Ribeiro MC, Metzger JP, Martensen AC, Ponzoni FJ, Hirota MM. (2009) The Brazilian Atlantic Forest: How much is left, and how is the remaining forest distributed? Implications for conservation. Biological conservation 142: 1141-1153 doi: 10.1016/j.biocon.2009.02.021 [Google Scholar]
- Romanenko S, Perelman P, Trifonov V, Graphodatsky A. (2012) Chromosomal evolution in Rodentia. Heredity 108: 4-16 doi: 10.1038/hdy.2011.110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seabright M. (1971) A rapid banding technique for human chromosomes. The Lancet 298: 971-972 doi: 10.1016/S0140-6736(71)90287-X [DOI] [PubMed] [Google Scholar]
- Silva MJJ, Yonenaga-Yassuda Y. (1998) Heterogeneity and meiotic behaviour of B and sex chromosomes, banding patterns and localization of (TTAGGG)n sequences by fluorescence in situ hybridization in the neotropical water rat Nectomys (Rodentia, Cricetidae). Chromosome Research 6: 455-462 doi: 10.1023/A:1009248311530 [DOI] [PubMed] [Google Scholar]
- Silva MJJ, Yonenaga-Yassuda Y. (1999) Autosomal and sex chromosomal polymorphisms with multiple rearrangements and a new karyotype in the genus Rhipidomys (Sigmodontinae, Rodentia). Hereditas 131: 211-220 doi: 10.1111/j.1601-5223.1999.00211.x [DOI] [PubMed] [Google Scholar]
- Silva MJJ, Yonenaga-Yassuda Y. (2004) B chromosomes in Brazilian rodents. Cytogenet Genome Res 106: 257-263 doi: 10.1159/000079296 [DOI] [PubMed] [Google Scholar]
- Suárez-Villota EY, Di-Nizo CB, Neves CL, Silva MJJ. (2013) First cytogenetic information for Drymoreomys albimaculatus (Rodentia, Cricetidae), a recently described genus from Brazilian Atlantic Forest. Zookeys: 65 76. doi: 10.3897/zookeys.303.4873 [DOI] [PMC free article] [PubMed]
- Sumner AT. (1972) A simple technique for demonstrating centromeric heterochromatin. Experimental Cell Research 75: 304-306 doi: 10.1016/0014-4827(72)90558-7 [DOI] [PubMed] [Google Scholar]
- Svartman M. (2009) American marsupials chromosomes: why study them? Genetics and Molecular Biology 32: 675–687. doi: 10.1590/S1415-47572009005000084 [DOI] [PMC free article] [PubMed]
- Tabarelli M, Mantovani W. (1999) A regeneração de uma floresta tropical montana após corte e queima (São Paulo-Brasil). Revista Brasileira de Biologia 59: 239-250 doi: 10.1590/S0034-71081999000200008 [Google Scholar]
- Trifonov VA, Dementy’eva PV, Beklemisheva VR, Yudkin DV, Vorobi’eva NV, Graphodatsky AS. (2010) Supernumerary chromosomes, segmental duplications, and evolution. Russian Journal of Genetics 46: 1234-1236 doi: 10.1134/S1022795410090206 [PubMed] [Google Scholar]
- Veloso HP, Rangel Filho ALR, Lima JCA. (1991) Classificação da vegetação brasileira, adaptada a um sistema universal. Ministério da Economia, Fazenda e Planejamento, Fundação Instituto Brasileiro de Geografia e Estatística, Diretoria de Geociências, Departamento de Recursos Naturais e Estudos Ambientais.
- Ventura K, O’Brien PC, Yonenaga-Yassuda Y, Ferguson-Smith MA. (2009) Chromosome homologies of the highly rearranged karyotypes of four Akodon species (Rodentia, Cricetidae) resolved by reciprocal chromosome painting: the evolution of the lowest diploid number in rodents. Chromosome Research 17: 1063-1078 doi: 10.1007/s10577-009-9083-5 [DOI] [PubMed] [Google Scholar]
- Ventura K, Sato-Kuwabara Y, Fagundes V, Geise L, Leite YL, Costa LP, Silva MJJ, Yonenaga-Yassuda Y, Rodrigues MT. (2012) Phylogeographic structure and karyotypic diversity of the Brazilian shrew mouse (Blarinomys breviceps, Sigmodontinae) in the Atlantic Forest. Cytogenet Genome Res 138: 19-30 doi: 10.1159/000341887 [DOI] [PubMed] [Google Scholar]
- Ventura K, Silva MJJ, Fagundes V, Pardini R, Yonenaga-Yassuda Y. (2004) An undescribed karyotype for Thaptomys (2n= 50) and the mechanism of differentiation from Thaptomys nigrita (2n= 52) evidenced by FISH and Ag-NORs. Caryologia 57: 89-97 doi: 10.1080/00087114.2004.10589376 [Google Scholar]
- Ventura K, Silva MJJ, Yonenaga-Yassuda Y. (2010) Thaptomys Thomas 1915 (Rodentia, Sigmodontinae, Akodontini) with karyotypes 2n= 50, FN= 48, and 2n= 52, FN= 52: Two monophyletic lineages recovered by molecular phylogeny. Genetics and Molecular Biology 33: 256-261 doi: 10.1590/S1415-47572010005000053 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang E. (2002) Diets of ocelots (Leopardus pardalis), margays (L. wiedii), and oncillas (L. tigrinus) in the Atlantic rainforest in southeast Brazil. Studies on Neotropical Fauna and Environment 37: 207-212 doi: 10.1076/snfe.37.3.207.8564 [Google Scholar]
- Weksler M, Percequillo AR, Voss RS. (2006) Ten new genera of oryzomyine rodents (Cricetidae: Sigmodontinae). American Museum Novitates: 1–29. doi: 10.1206/0003-0082(2006)3537[1:TNGOOR]2.0.CO;2
- Woods C, Kilpatrick C. (2005) Infraorder Hystricognathi. In: Wilson D, Reeder D (Eds) Mammal species of the world: a taxonomic and geographic reference Baltimore, Maryland: Johns Hopkins University Press, 1538–1600.
- Yonenaga-Yassuda Y, Prado RC, Mello DA. (1987) Supernumerary chromosomes in Holochilus brasiliensis and comparative cytogenetic analysis with Nectomys squamipes (Cricetidae, Rodentia). Revista Brasileira de Genética 10: 209-220 [Google Scholar]
- Yonenaga-Yassuda Y, Souza M, Kasahara S, L’abbate M, Chu H. (1985) Supernumerary system in Proechimys iheringi iheringi (Rodentia: Echimyidae) from the State of Sao Paulo, Brazil. Caryologia 38: 179-194 [Google Scholar]
- Zanchin NI, Langguth A, Mattevi MS. (1992) Karyotypes of Brazilian species of Rhipidomys (Rodentia, Cricetidae). Journal of Mammalogy 73: 120-122 doi: 10.2307/1381872 [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.