

Abstract
Context
Research has implicated an addictive process in the development and maintenance of obesity. Although parallels in neural functioning between obesity and substance dependence have been found, no studies have examined the neural correlates of addictive-like eating behavior.
Objective
To test the hypothesis that elevated “food addiction” scores are associated with similar patterns of neural activation as substance dependence.
Design
Between-Subjects fMRI study.
Participants
Forty-eight healthy adolescent females ranging from lean to obese recruited for a healthy weight maintenance trial.
Main Outcome Measure
The relation between elevated “food addiction” scores and blood oxygen level-dependent fMRI activation in response to receipt and anticipated receipt of palatable food (chocolate milkshake).
Results
Food addiction scores (N = 39) correlated with greater activation in the anterior cingulate cortex (ACC), medial orbitofrontal cortex (OFC), and amygdala in response to anticipated receipt of food (P <0.05, false-discovery rate (FDR) corrected for multiple comparisons in small volumes). Participants with higher (n=15) versus lower (n=11) food addiction scores showed greater activation in the dorsolateral prefrontal cortex (DLPFC) and the caudate in response to anticipated receipt of food, but less activation in the lateral OFC in response to receipt of food (pFDR <0.05).
Conclusions
Similar patterns of neural activation are implicated in addictive-like eating behavior and substance dependence; elevated activation in reward circuitry in response to food cues and reduced activation of inhibitory regions in response to food intake.
One-third of American adults are now obese1 and obesity-related disease is the second leading cause of preventable death2. Unfortunately, most obesity treatments do not result in lasting weight loss, as most patients regain their lost weight within five years3.
Based on numerous parallels in neural functioning associated with substance dependence1 and obesity, theorists have proposed that addictive processes may be involved in the etiology of obesity4,5. Food and drug use both result in dopamine release in mesolimbic regions and the degree of release correlates with subjective reward from both food and drug use5,6. Similar patterns of brain activation in response to food and drug cues have also been found. Individuals with versus without substance dependence show greater activation in brain regions that encode the reward value of stimuli (e.g., the orbitofrontal cortex (OFC), amygdala, insula, striatum, anterior cingulate cortex (ACC) and dorsolateral prefrontal cortex (DLPFC))7,8 and greater dopamine release in the dorsal striatum in response to drug cues9. Similarly, obese versus lean individuals show greater activation in the OFC, amygdala, ACC, striatum and mediodorsal thalamus in response to food cues7 and greater activation in regions associated with cue-related craving for drugs, such as the ACC, striatum, insula and the DLPFC in response to anticipated receipt of palatable foods10,11,12.
Although obese and substance dependent individuals show hyper-responsivity of reward learning regions to food and substance cues respectively, actual intake of food and drugs is associated with reduced reward circuitry activation. Obese versus lean individuals show less dorsal striatal and medial OFC activation in response to palatable food intake13,14, echoing evidence that substance dependent individuals exhibit blunted dopaminergic release during actual drug consumption and report weaker subjective reward relative to healthy controls15,16,17,18. Results dovetail with evidence of reduced D2 receptor availability in obese and substance dependent individuals versus healthy controls19,20. These findings have prompted the theory that individuals who experience less reward from food intake may overeat to compensate for this reward deficit19,21.
Although there are strong parallels in brain regions that encode reward from drugs and palatable foods and in neural abnormalities associated with substance dependence and obesity, these findings may tell us little about true “food addiction” (FA). Obesity is strongly linked to excess food consumption, but other factors contribute to unhealthy weight gain, such as physical inactivity22. Additionally, excess consumption is not necessarily indicative of substance dependence; whereas 40% of college students binge drink23, only 6% meet criteria for alcohol dependence24. Thus, to more directly assess FA it would be useful to identify participants who may exhibit signs of dependence in their eating behavior. Currently, a diagnosis of substance dependence is given when sufficient behavioral criteria are met (see Table 1). The Yale Food Addiction Scale (YFAS) was developed to operationalize the construct of palatable food dependence based upon the DSM-IV-TR25 substance dependence criteria26. The identification of individuals who exhibit FA symptoms would allow for more direct examinations of the neurobiological similarities between substance dependence and compulsive food consumption.
Table 1.
Diagnostic Criteria for Substance Dependence as Stated by the DSM-IV-TR25
1. Tolerance, as defined by either of the following:
|
2. Withdrawal, as manifested by either of the following:
|
| 3. Taking the substance often in larger amounts or over a longer period than was intended. |
| 4. There is a persistent desire or unsuccessful effort to cut down or control substance use. |
| 5. Spending a great deal of time in activities necessary to obtain or use the substance or to recover from its effects. |
| 6. Giving up social, occupational, or recreational activities because of substance use. |
| 7. Continuing the substance use with the knowledge that it is causing or exacerbating a persistent or recurrent physical or psychological problem. |
In the present study we examined the relation of food addiction symptoms, as assessed by the YFAS, with neural activation in response to: 1) cues signaling impending delivery of a highly palatable food (chocolate milkshake) versus a tasteless control solution and 2) intake of chocolate milkshake versus tasteless solution among healthy adolescent females ranging from lean to obese. Based on previous findings, we hypothesized that participants exhibiting elevated FA symptoms would show greater activation in response to food cues in the amygdala, striatum, OFC, DLPFC, thalamus, midbrain, insula, and anterior cingulate gyrus. Further, we hypothesized that, during consumption of a highly palatable food, the high versus low FA group would demonstrate less activation in the dorsal striatum and OFC, analogous to the reduced activation demonstrated in substance dependent participants upon receipt of a drug.
METHODS
Participants
Participants were 48 young women (M age = 20.8, SD = 1.31); M Body Mass Index [BMI; Kg/M2] = 28.0, SD = 3.0, range 23.8 - 39.2) who enrolled in a program developed to help people achieve and maintain a healthy weight on a long-term basis. Data from this sample has been published previously14,27. Individuals who reported DSM-IV binge eating or compensatory behaviors (e.g., vomiting for weight control), use of psychotropic medications or illicit drugs over the past three months, smoking, head injury with a loss of consciousness, or current (past three months) Axis I psychiatric disorder were excluded. Written informed consent was obtained from participants. The local Institutional Review Board approved this study.
Measures
Body Mass
The body mass index (BMI = kg/m2) was used to reflect adiposity. After removal of shoes and coats, height was measured to the nearest millimeter using a stadiometer and weight was assessed to the nearest 0.1 kg using a digital scale. Two measures of height and weight were obtained and averaged.
Yale Food Addiction Scale (YFAS)
The Yale Food Addiction Scale26 is a 25-item measure developed to operationalize FA by assessing signs of substance dependence symptoms (e.g., tolerance, withdrawal, loss of control) in eating behavior. The YFAS has shown internal consistency (α = .86), as well as convergent and incremental validity. The YFAS provides two scoring options; a symptom count version and a diagnostic version. To receive a “diagnosis” of FA, it is necessary to report experiencing three or more symptoms in the past year and clinically significant impairment or distress. The version of the YFAS used in the current study measured all items on a Likert scale. In line with YFAS scoring instructions, five of the Likert scale items were dichotomized, such that participants who indicated that they had “never” experienced the symptom were assigned a value of zero, and those reporting ever experiencing the symptom in the past year were assigned a value of one.
Data Management
The YFAS exhibited a normal distribution (skew and kurtosis coefficients <2). Four participants with significant missing data on the YFAS and five participants who showed excessive head movement during the scan were excluded, resulting in a final N = 39. The primary aim was to test whether YFAS scores correlate with neural activation in brain regions associated with substance dependence. We expected YFAS scores to correlate positively with activation in regions that encode the reward value of stimuli in response to anticipated receipt of palatable food, but negatively with activation in these regions in response to food intake. Secondary analyses explored potential differences in activation of participants who likely experienced FA compared to healthy controls. Few participants reported experiencing clinically significant impairment or distress on the YFAS (n = 2), potentially due to the exclusion of participants with eating disorders and Axis I disorders. To more closely approximate those exhibiting signs of food-related substance dependence versus healthy eating behaviors, participants were placed into a high FA group with three or more symptoms (n=15) and a low FA group with one or fewer symptoms (n=11). Participants reporting two symptoms were omitted from these analyses (n=13) to ensure adequate separation between high and low FA groups.
Procedures
fMRI paradigm
Participants were scanned at baseline. Participants were asked to consume regular meals, but to refrain from eating or drinking (including caffeinated beverages) for 4-6 hours immediately preceding their imaging session. This deprivation period was selected to capture the hunger state that most individuals experience as they approach their next meal, which is a time when individual differences in food reward would logically impact caloric intake. Most participants completed the paradigm between 10:00 a.m. and 1:00 p.m., but a subset completed scans between 2:00 and 4:00 p.m. Before the imaging session, participants were familiarized with the fMRI paradigm through practice on a separate computer.
The milkshake paradigm was designed to examine activation in response to consumption and anticipated consumption of palatable food (Figure 1). Stimuli consisted of 2 images (glass of milkshake and glass of water) that signaled the delivery of either 0.5 ml of a chocolate milkshake (4 scoops of Häagen-Daz vanilla ice cream, 1.5 cups of 2% milk, and 2 tablespoons of Hershey’s chocolate syrup) or a calorie-free tasteless solution, designed to mimic the natural taste of saliva (25 mM KCl and 2.5 mM NaHCO3 diluted in 500ml of distilled water). We used artificial saliva because water has a taste that activates the taste cortex28. Order of presentation was randomized across participants. Images were presented for 2 seconds using MATLAB followed by a jitter of 1-3 secs during which a blank screen with a cross-hair at the center for fixation was presented (to eliminate random eye-movement). Taste delivery occurred 5 seconds after onset of the cue and lasted 5 seconds. On 40% of the chocolate and tasteless solution trials the taste was not delivered as expected to allow the investigation of the neural response to anticipation of a taste that was not confounded with actual food receipt (unpaired trials). Each run consisted of 30 events each of milkshake cue and milkshake intake and 20 events each of tasteless solution cue and of tasteless solution intake. Fluids were delivered using programmable syringe pumps (Braintree Scientific BS-8000) controlled by MATLAB to ensure consistent volume, rate, and timing of taste delivery. Sixty ml syringes filled with chocolate milkshake and tasteless solution were connected via Tygon tubing through a wave guide to a manifold attached to the scanner head coil. The manifold fit into the participants’ mouths and delivered the taste to a consistent segment of the tongue. Participants were instructed to swallow when they saw the ‘swallow’ cue. Images were presented with a digital projector/reverse screen display system at the end of the MRI scanner bore, visible via a head coil mounted mirror. Before the scan, participants consumed the milkshake and tasteless solution and rated desire for, perceived pleasantness, edibility, and intensity of the tastes on cross-modal visual analog scales. This procedure has been successfully used in the past to deliver liquids in the scanner, as described in detail elsewhere14.
Figure 1.
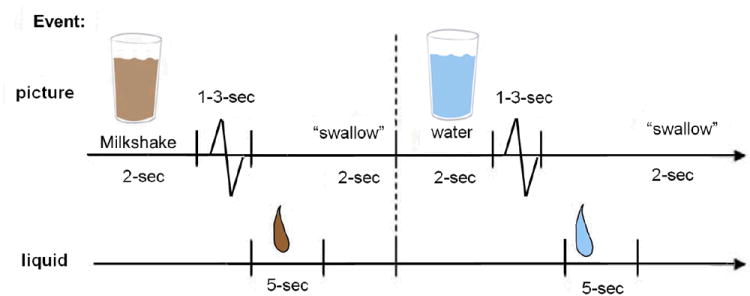
Example of timing and ordering of presentation of pictures and drinks during the run. Drops represent delivery of chocolate milkshake (brown) or tasteless solution (blue).
Imaging and statistical analysis
Scanning was performed with a 3 Tesla head-only MRI scanner. A standard birdcage coil acquired data from the entire brain. A thermo foam vacuum pillow and padding restricted head motion. In total, 229 volumes were collected during each functional run. Functional scans used a T2* weighted gradient single-shot echo planar imaging (EPI) sequence (TE=30 ms, TR = 2000 ms, flip angle=80°) with an in plane resolution of 3.0 × 3.0 mm2 (64 × 64 matrix; 192 × 192 mm2 field of view). To cover the whole brain, 32 4mm slices (interleaved acquisition, no skip) were acquired along the AC-PC transverse, oblique plane as determined by the midsagittal section. Structural scans were collected using an inversion recovery T1 weighted sequence (MP-RAGE) in the same orientation as the functional sequences to provide detailed anatomic images aligned to the functional scans. High-resolution structural MRI sequences (FOV = 256 × 256 mm2, 256 × 256 matrix, thickness = 1.0 mm, slice number ≈ 160) were acquired.
Data were pre-processed and analyzed using SPM5 software29 in MATLAB30,31. Images were time-acquisition corrected to the slice obtained at 50% of the TR. Functional images were realigned to the mean. Anatomical and functional images were normalized to the standard MNI template brain implemented in SPM5 (ICBM152, based on an average of 152 normal MRI scans). Normalization resulted in a voxel size of 3 mm3 for functional images and 1 mm3 for structural images. Functional images were smoothed with a 6 mm FWHM isotropic Gaussian kernel. Excessive movement was probed using the realignment parameters and was defined as movement >1 mm in any direction during the paradigm. To identify brain regions activated by anticipation of food receipt, we contrasted BOLD response during unpaired milkshake cue versus unpaired tasteless solution cue. We analyzed data from unpaired cue presentation in which the tastes were not delivered to ensure that taste receipt would not influence our definition of anticipatory activation. We contrasted BOLD response during receipt of milkshake versus tasteless solution to identify brain regions activated in response to food consumption. We considered the arrival of a taste in the mouth to be consummatory reward, rather than when the solution was swallowed, but acknowledge that post-ingestive effects contribute to the reward value of food32. Condition-specific effects at each voxel were estimated using general linear models. Vectors of the onsets for each event of interest were compiled and entered into the design matrix so that event-related responses could be modeled by the canonical hemodynamic response function (HRF), as implemented in SPM5, consisting of a mixture of 2 gamma functions that emulate the early peak at 5 seconds and the subsequent undershoot. To account for the variance induced by swallowing the solutions, we included the time of the swallow cue as a control variable. We also included temporal derivatives of the hemodynamic function to obtain a better model of the data33. A 128 second high-pass filter (per SPM5 convention) removed low-frequency noise and slow drifts in the signal.
Individual maps were constructed to compare the activations within each participant for the contrasts “unpaired milkshake cue – unpaired tasteless cue” and “milkshake receipt – tasteless receipt”, which were then regressed against total YFAS scores using SPM5. To detect group differences, two second level 2 × 2 ANOVAs were conducted: (high FA group vs. low FA group) by (milkshake receipt– tasteless receipt) and (high FA group vs. low FA group) by (unpaired milkshake – unpaired tasteless). T-map threshold was set at P uncorrected = 0.001 and a 3-voxel cluster size. We performed small volume correction (SVC) analyses using peaks with the highest volumes (mm3) and z-values identified previously in the cue-induced craving and drug administration literatures8,34,35 as well as in food cue/food administration studies14, 36, 37. To test our hypothesis that participants exhibiting more FA symptoms would demonstrate greater activation in response to food cues, search volumes were restricted within a 10-mm radius of coordinates in the OFC (42, 46, -16; -8, 60, -14), caudate (9, 0, 21), amygdala (-12, -10,-16), ACC (-10,24, 30; -4, 30, 16), DLPFC (-30, 36, 42), thalamus (-7,-26,9), midbrain (-12, -20, -22; 3, -28, -13), and insula (36, 12, 2). To test our hypothesis that during consumption of a highly palatable food, the high versus low FA group would demonstrate less activation in reward-related brain regions, search volumes were restricted within a 10-mm radius of coordinates in the OFC (± 42,46, -16; ± 41, 34, -19; ±8, 60, -14) and caudate (±9, 0, 21; ±2, -9, 34). Predicted activations were considered to be significant at p<.0.05 after correcting for multiple comparisons (pFDR) across the voxels within the a priori defined small volumes. Bonferroni corrections were then used to correct for the number of regions of interest tested. Because Dreher et al. (2007)38 reported that women in the mid-follicular phase (4-8 d after first period) show greater response in reward regions compared to those in the luteal phase, we attempted to run the scans for all women during the same period of the menstrual cycle. However, because of scheduling difficulties, two participants were scanned during the mid-follicular phase. When these individuals were excluded, the relations between YFAS and BOLD responses to food intake and anticipated intake remained significant.
Results
On average, high FA participants endorsed approximately four FA symptoms (M = 3.60, SD = .63), whereas the low FA group all endorsed one FA symptom. There were no significant differences found between the high and low FA groups on age (F (1, 24) = 2.25, p = .147), BMI (F (1, 24) = 1.14, p = .296), or on pleasantness ratings of the milkshake administered during the study (F (1, 24) = .013, p = .910). YFAS scores in the current study correlated with the emotional eating (rs = .34, p = .03) and external eating (rs = .37, p = .02) subscales of the Dutch Eating Behavior Questionnaire39.
Correlations between FA Symptoms and Response to Anticipation and Intake of Palatable Food2
YFAS scores (N =39) showed positive correlations with activation in the left ACC (Figure 2), left medial OFC (Figure 3), and left amygdala in response to anticipated intake of palatable food (Table 2). The activation in the left ACC and left OFC survived the more stringent Bonferroni correction (0.05/11 regions of interest = 0.0045). We derived the effect sizes (r) from the Z-values (Z/√N). Effect sizes were all medium to large per Cohen’s criteria40 (M r = .60). There were no significant correlations in the hypothesized regions in response to consumption of palatable food.
Figure 2.
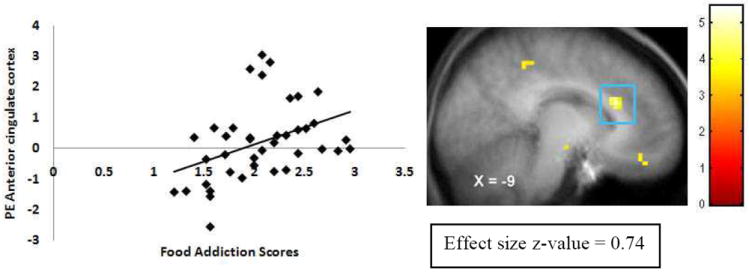
Activation in a region of the anterior cingulate cortex (-9, 24, 27, Z = 4.64, pFDR < .001) during milkshake cues – tasteless cues as a function of YFAS scores with the graph of parameter estimates (PE) from that peak.
Figure 3.
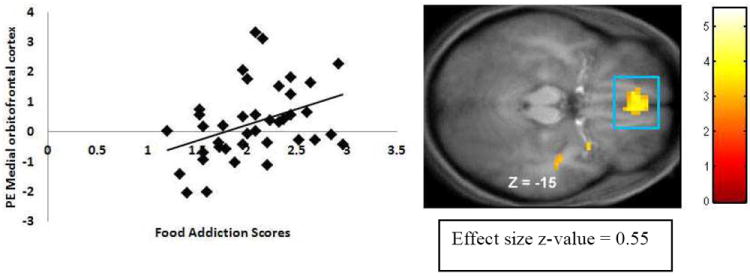
Activation in a region of the medial orbitofrontal cortex (3, 42, -15, Z = 3.47 pFDR = .004) during milkshake cues – tasteless cues as a function of YFAS scores with the graph of parameter estimates (PE) from that peak.
Table 2.
Regions Responding during Anticipatory Food Reward and Consummatory Food Reward as a Function of YFAS scores (N = 39)
| Reference coordinates1 | X | Y | Z | Cluster | Z | P FDR corrected | Effect size z-value | |
|---|---|---|---|---|---|---|---|---|
| Milkshake cue – tasteless solution cue | ||||||||
| Anterior cingulate cortex | -10,24,30 | -9 | 24 | 27 | 16 | 4.64 | .00 | 0.74 |
| Medial orbitofrontal cortex | -8, 60, -14 | -6 | 45 | -15 | 31 | 3.61 | .004 | 0.58 |
| Amygdala | -12,-10,-16 | -15 | -9 | -9 | 3 | 3.41 | .007 | 0.55 |
Reference coordinates from Gilman et al., 2008, McBride et al., 2006, Risinger et al., 2005, Small et al., 2001, or Stice et al., 2008
Statistically significant results that survived the Bonferroni correction are highlighted in bold.
Response to Anticipation and Intake of Palatable Food for Participants with High versus Low FA Scores
Participants in the High FA group versus the Low FA group showed greater activation in the left DLPFC (Figure 4) and right caudate (Figure 5). The activation in the right caudate survived the Bonferroni correction (0.05/11 regions of interest = 0.0045). Further, the High FA group showed less activation in the left lateral OFC (Figure 6) during food intake than the Low FA group (Table 3). This peak also survived the Bonferroni correction (0.05/3 regions of interest = 0.017). Effect sizes from these analyses were large (M r = .71).
Figure 4.
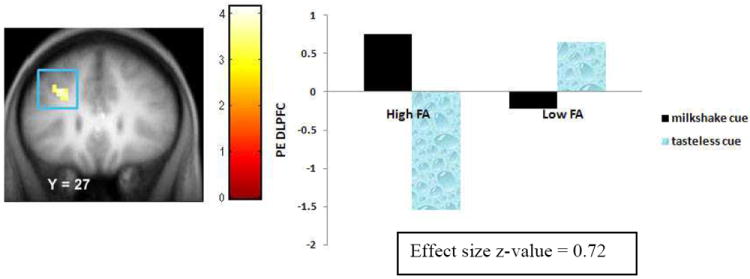
Activation in a region of the dorsolateral prefrontal cortex (-27, 27, 36, Z = 3.72, pFDR = .007) during anticipatory food reward (milkshake cue – tasteless cue) in the High FA group versus Low FA group with the bar graphs of parameter estimates from that peak.
Figure 5.
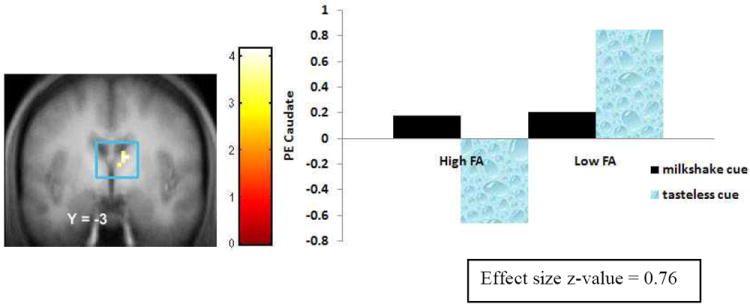
Activation in a region of the caudate (9, -3, 21, Z = 3.96, pFDR = .004) during anticipatory food reward (milkshake cue – tasteless cue) in the High FA group versus Low FA group with the bar graphs of parameter estimates from that peak.
Figure 6.
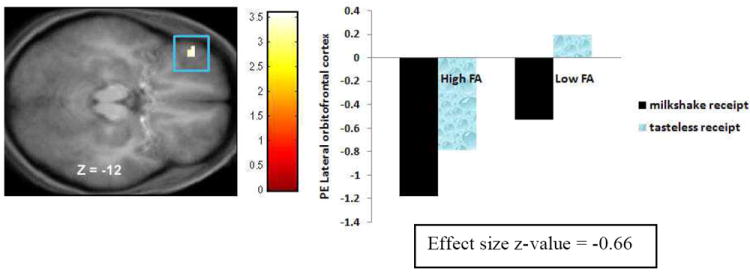
Activation in a region of the lateral orbitofrontal cortex (-42, 42, -12, Z = -3.45, pFDR = .009) during consummatory reward (milkshake receipt – tasteless receipt) in the High FA group versus Low FA group with the bar graphs of parameter estimates from that peak.
Table 3.
Regions Showing Activation during Anticipatory Food Reward and Consummatory Food Reward in High FA Individuals (N = 15) compared to Low FA Individuals (N = 11)
| Reference coordinates1 | X | Y | Z | Cluster | Z | P FDR corrected | Effect size z-value | |
|---|---|---|---|---|---|---|---|---|
| Milkshake cue – tasteless solution cue | ||||||||
| Dorsolateral Prefrontal cortex | -30, 36, 42 | -27 | 27 | 36 | 18 | 3.72 | .007 | .72 |
| Caudate | 9, 0, 21 | 9 | -3 | 21 | 11 | 3.96 | .004 | .76 |
| Receipt milkshake – receipt tasteless solution | ||||||||
| Lateral orbitofrontal cortex | 42, 46, -16 | -42 | 42 | -12 | 5 | -3.45 | .009 | -.66 |
Reference coordinates from Gilman et al., 2008, McBride et al., 2006, Risinger et al., 2005, Small et al., 2001, or Stice et al., 2008
Statistically significant results that survived the Bonferroni correction are highlighted in bold.
Discussion
In the current study, lean and obese participants with higher FA scores demonstrated a differential pattern of neuronal activation from participants with lower FA scores. Although studies have explored the association of anticipatory and consummatory reward with BMI 12,13,14, this is the first study to examine the relation between FA and neural activation of reward circuitry to intake and anticipated intake of palatable food. FA scores correlated positively with activation in the ACC, medial OFC, and amygdala in response to anticipated intake of palatable food, but were not significantly related to activation in response to palatable food intake. Further, high versus low FA participants demonstrated greater activation in the DLPFC and caudate during anticipated palatable food intake and reduced activation in the lateral OFC during palatable food intake.
As predicted, elevated FA scores were associated with greater activation of regions that play a role in encoding the motivational value of stimuli in response to food cues. The ACC and medial OFC have both been implicated in motivation to feed41,42, and to consume drugs among individuals with substance dependence43. Elevated ACC activation in response to alcohol-related cues is also associated with reduced D2 receptor availability44 and increased risk for relapse45. Similarly, increased activation in the amygdala is associated with increased appetitive motivation46 and exposure to foods with greater motivational and incentive value47. In addition, the DLPFC is associated with memory, planning48, attentional control49, and goal-directed behavior49. Hare and colleagues50 found that participants who attempted to resist pleasurable foods also exhibited elevated DLPFC activation, which was linked to reduced activity in areas implicated in coding food reward, such as the ventromedial prefrontal cortex. Thus, participants with higher FA scores may respond to increased appetitive motivation for food by attempting to implement self-control strategies. It has also been suggested that DLPFC activation by drug cues relates to integration of information about internal state (craving, withdrawal), motivation, expectancy, and cues in the regulation and planning of drug-seeking behavior7. Similarly, the caudate also appears to play a role in enhanced motivation. Elevated caudate activation is associated with expectation of a positive reward51, exposure to cues with increased incentive value52, and exposure to drug-stimuli for substance dependent participants53. Thus, greater FA scores may be related to stronger motivations to seek out food in response to food-related cues.
Neural activation of regions that appear to play a role in the encoding of craving was also positively correlated to FA scores. For example, activation in the ACC and medial OFC is associated with craving in substance use disorders54,55. The amygdala has also been commonly implicated in drug cue reactivity56 and drug craving57. Further, activation in the caudate is associated with craving for palatable foods58, as well as craving in response to drug cues in substance dependent participants53, 59. Thus, FA scores may be associated with greater cue-triggered food cravings.
Finally, FA scores were associated with activation in regions that play a role in disinhibition and satiety. Interestingly, although FA was correlated positively with activation in the medial OFC during anticipatory food reward, FA scores were correlated negatively with activation in the lateral OFC during receipt of food. These findings are consistent with research showing very different patterns of response in these regions. Specifically, Small et al. (2001)36 found that the medial and lateral caudal OFC showed opposite patterns of activity during chocolate consumption prompting the suggestion that this pattern occurs as participants’ desire to eat decreases and their behavior (eating) comes to be inconsistent with their desires. Thus, lateral OFC activity occurs when the desire to stop eating is suppressed. Similar dissociations between the medial and lateral OFC have also been found in substance dependence. Unlike the medial OFC, which is more closely related to subjective evaluation of reward60, increased activation in the lateral OFC is associated with greater inhibitory control46,61 and a greater ability to suppress previously rewarded responses62. Substance dependent participants typically exhibit increased activation in the medial OFC in response to drug cues54,55, but also exhibit hypoactivation in the lateral OFC46, suggesting less inhibitory control in response to reward cues. The reduced activation in the lateral OFC in high FA individuals observed herein may be related to either less inhibitory control during intake of palatable food or a reduced satiety response during palatable food intake.
In sum, these findings support the theory that compulsive food consumption may be driven in part by an enhanced anticipation of the rewarding properties of food12. Similarly, addicted individuals are more likely to be physiologically, psychologically, and behaviorally reactive to substance-related cues63, 64. This process may be due in part to incentive salience which suggests that cues associated with the substance (in this case food) may begin triggering the release of dopamine and driving consumption65,66. Brain regions associated with dopaminergic release also showed significantly greater activation during cue exposure in high FA participants. The possibility that food-related cues may develop pathological properties is especially concerning in the current food environment where palatable foods are constantly available and heavily marketed.
In contrast to our initial hypotheses, there were limited differences in reward circuitry activation between high FA and low FA participants during food intake. These findings provide little support for the notion that abnormal reward response to food intake drives food addiction. Instead, the high FA group exhibited patterns of neural activation associated with reduced inhibitory control. Previous studies have found that administration of a priming dose can trigger excessive consumption in participants with substance use problems67,68 and eating pathology 69,70,71. The current results, taken in concert with these previous findings, suggest that consumption of a palatable food may override desires to limit caloric food consumption in high FA participants, resulting in disinhibited food consumption.
Interestingly, no significant correlation was found between YFAS scores and BMI. Thus, the current findings suggest that FA scores and related neural functioning can occur among individuals with a range of body weights. In the initial validation, the YFAS was also not significantly related to BMI, but was associated with binge eating, emotional eating, and problematic eating attitudes26. Similarly, herein, the YFAS was correlated with emotional eating and external eating. It is possible that some individuals experience compulsive eating behavior, but engage in compensatory behaviors to maintain a lower weight. An alternative possibility is that lean participants that endorse FA are at risk for future weight-gain. Given the young age of the sample, the probability of future weight gain may be especially likely. Either possibility suggests that examining FA in lean participants may be beneficial in identifying individuals at-risk for weight gain or disordered eating and that the YFAS may provide important information above and beyond current BMI.
It is important to consider the limitations of this study. First, potentially due to the exclusion of participants with eating disorders and Axis I disorders, few participants met the clinically significant distress or impairment criteria of the YFAS, which is required for a FA “diagnosis.” Thus, the current study should be considered a conservative test and future studies of the neural correlates of FA should include participants with more severe scores. Second, although we asked participants to refrain from eating 4 to 6 hours prior to their scan session, we did not measure hunger. Fasting and hunger are associated with similar patterns of neural response, such as increased activation in the medial OFC and amygdala72,73. It is possible that participants with higher FA scores experienced more hunger. If this was the case, it may have contributed to some of the observed effects. It is also possible that increased hunger may interact with FA, as both addiction and hunger are associated with elevated drive74. Future studies should examine the relation between FA, hunger, and reward circuitry response to food intake and anticipated intake. Third, the current study was conducted solely with female participants, thus results should be generalized with caution to males. Fourth, this study is cross-sectional, which did not allow us to evaluate the time course of the development of FA and related neural correlates. A longitudinal design would allow for a greater understanding of the antecedents and consequences of FA. Fifth, regions implicated in the current study are also implicated in non-addictive reward-related behavior, thus future studies would benefit from collecting addiction-related measures during the scan, such as craving and loss of control. Finally, the sample size of the current study is relatively small, thus there may have been limited power to detect other effects, such as individual differences in neural response to food intake.
The current findings have implications regarding future research directions. First, given that some types of eating behavior may be driven by food cues, it will be important to examine neural activation in response to food advertisements. In addition, to further explore the role of disinhibition in FAs, it will be useful to measure feelings of loss of control and ad libitum food consumption. Further, the use of fMRI technology does not allow for the direct measurement of dopamine release or dopamine receptors. It will be important to examine induced dopamine release and D2 receptor availability in participants who report indicators of FA. Finally, although dopamine is implicated in both feeding and addictive behaviors, other neurotransmitters are also likely to play an important role (e.g., opioid, GABA). Thus, future studies on the association between FA and neural activation associated with these neurotransmitters will also be important.
Despite the aforementioned limitations, the current findings suggest that FA is associated with reward-related neural activation that is frequently implicated in substance dependence. This is the first study to link indicators of addictive eating behavior with a specific pattern of neural activation. The current study also provides evidence that objectively measured biological differences are related to variations in YFAS scores, thus providing further support for the validity of the scale. Further, if certain foods are addictive, this may partially explain the difficulty people experience in achieving sustainable weight loss. If food cues take on enhanced motivational properties in a manner analogous to drug cues, efforts to change the current food environment may be critical to successful weight loss and prevention efforts. Ubiquitous food advertising and the availability of inexpensive palatable foods may make it extremely difficult to adhere to healthier food choices because the omnipresent food cues trigger the reward system. Finally, if palatable food consumption is accompanied by disinhibition, the current emphasis on personal responsibility as the anecdote to increasing obesity rates may have minimal effectiveness.
Acknowledgments
This project was supported by the following grant: Roadmap Supplement R1MH64560A.
Ms. Gearhardt is the corresponding author and she takes responsibility for the integrity of the data and the accuracy of the data analysis, and states that all authors had full access to all the data in the study.
Footnotes
In the current paper, the terms substance dependence and addiction are used interchangeably to represent a diagnosis of substance dependence as defined by the Diagnostic and Statistical Manual IV-TR25.
All peaks remained significant when BMI was statistically controlled for in analyses.
All authors report no conflict of interest with respect to the content of this paper.
References
- 1.Yach D, Stuckler D, Brownell KD. Epidemiologic and economic consequences of the global epidemics of obesity and diabetes. Nature. 2006;12:62–65. doi: 10.1038/nm0106-62. [DOI] [PubMed] [Google Scholar]
- 2.Mokdad AH, Marks JS, Stroup MF, Gerberding JL. Actual causes of death in the United States, 2000. JAMA. 2004;291:1238–1245. doi: 10.1001/jama.291.10.1238. [DOI] [PubMed] [Google Scholar]
- 3.Wadden TA, Butryn ML, Byrne KJ. Efficacy of lifestyle modification for long term weight control. Obes Res. 2004;12:151–162. doi: 10.1038/oby.2004.282. [DOI] [PubMed] [Google Scholar]
- 4.Volkow ND, O’Brien CP. Issues for DSM-V: Should obesity be included as a brain disorder? Am J Psychiatry. 2007;164:708–10. doi: 10.1176/ajp.2007.164.5.708. [DOI] [PubMed] [Google Scholar]
- 5.Volkow ND, Wang GJ, Fowler JS, Telang F. Overlapping neuronal circuits in addiction and obesity: evidence of systems pathology. Philos Trans R Soc Lond B Biol Sci. 2008;363:3191–3200. doi: 10.1098/rstb.2008.0107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Volkow ND, Wang GJ, Fowler JS, Logan J, Jayne M, Franceschi D, Wong C, Gatley SJ, Gifford AN, Ding Y-S, Pappas N. “Nonhedonic” food motivation in humans involves dopamine in the dorsal striatum and methvlphenidate amplifies this effect. Synapse. 2002;44:175–180. doi: 10.1002/syn.10075. [DOI] [PubMed] [Google Scholar]
- 7.McBride D, Barrett SP, Kelly JT, Aw A, Dagher A. Effects of expectancy and abstinence on the neural response to smoking cues in cigarette smokers: An fMRI study. Neuropsychopharmacology. 2006;31:2728–2738. doi: 10.1038/sj.npp.1301075. [DOI] [PubMed] [Google Scholar]
- 8.Franklin TF, Wang Z, Wang J, Sciortino N, Harper D, Li Y, Ehrman R, Kampman K, O’Brien C, Detre JA, Childress AR. Limbic activation to cigarette smoking cues independent of nicotine withdrawal: a perfusion fMRI study. Neuorpsychopharmacology. 2007;32:2301–9. doi: 10.1038/sj.npp.1301371. [DOI] [PubMed] [Google Scholar]
- 9.Volkow ND, Wang GJ, Telang F, Fowler JS, Logan J, Childress AR, Jayne M, Ma Y, Wong C. Cocaine cues and dopamine in dorsal striatum: mechanism of craving in cocaine addiction. J Neurosci. 2006;26:6583–6588. doi: 10.1523/JNEUROSCI.1544-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Rothemund Y, Preuschhof C, Bohner G, Bauknecht H, Klingebiel R, Flor H, Klapp BF. Differential activation of the dorsal striatum by high-calorie visual food stimuli in obese individuals. NeuroImage. 2007;37:410–421. doi: 10.1016/j.neuroimage.2007.05.008. [DOI] [PubMed] [Google Scholar]
- 11.Stoeckel LE, Weller RE, Cook EW, Twieg DB, Knowlton RC, Cox JF. Widespread reward-system activation in obese women in response to pictures of high-calorie foods. NeuroImage. 2008;41:636–647. doi: 10.1016/j.neuroimage.2008.02.031. [DOI] [PubMed] [Google Scholar]
- 12.Stice E, Spoor S, Ng J, Zald DH. Relation of obesity to consummatory and anticipatory food reward. Physiology & Behavior. 2009;97:551–560. doi: 10.1016/j.physbeh.2009.03.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Stice E, Spoor S, Bohon C, Small DM. Relation between obesity and blunted striatal response to food is moderated by TaqlA1 DRD2 gene. Science. 2008;322:449–452. doi: 10.1126/science.1161550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Stice E, Spoor S, Bohon C, Veldhuizen M, Small DM. Relation of reward from food intake and anticipated intake to obesity: A functional magnetic resonance imaging study. J Abnorm Psychol. 2008;117:924–935. doi: 10.1037/a0013600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Volkow ND, Wang GJ, Fowler JS, Logan J, Gatley SJ, Hitzemann R, Chen AD, Dewey SL, Pappas N. Decreasd striatal dopaminergic responsivity in detoxified cocaine abusers. Nature. 1997;386:830–33. doi: 10.1038/386830a0. [DOI] [PubMed] [Google Scholar]
- 16.Volkow ND, Wang G-J, Telang F, Fowler JS, Logan J, Jayne M, Ma Y, Pradhan K, Wong C. Profound decreases in dopamine release in striatum in detoxified alcoholics: possible orbitofronal involvement. J Neurosci. 2007;27:12700–12706. doi: 10.1523/JNEUROSCI.3371-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Martinez D, Gil R, Slifstein M, Hwang DR, Huang YY, Perez A, Kegeles L, Talbot P, Evans S, Krystal J, Laruelle M, Abi-Dargham A. Alcohol dependence is associated with blunted dopamine transmission in the ventral striatum. Biol Psychiatry. 2005;58:779–786. doi: 10.1016/j.biopsych.2005.04.044. [DOI] [PubMed] [Google Scholar]
- 18.Martinez D, Narendran R, Foltin RW, Slifstein M, Hwang D-R, Broft A, Huang Y, Cooper TB, Fischman MW, Kleber HD, Laruelle M. Amphetamine-induced dopamine release: Markedly blunted in cocaine dependence and predictive of the choice to self-administer cocaine. Am J Psychiatry. 2007;164:622–629. doi: 10.1176/ajp.2007.164.4.622. [DOI] [PubMed] [Google Scholar]
- 19.Wang G-J, Volkow ND, Logan J, Pappas NR, Wong CT, Zhu W, Netusil N, Fowler JS. Brain dopamine and obesity. Lancet. 2001;357:354–357. doi: 10.1016/s0140-6736(00)03643-6. [DOI] [PubMed] [Google Scholar]
- 20.Volkow ND, Fowler JS, Wang G-J, Swanson JM. Dopamine in drug abuse and addiction: results from imaging studies and treatment implications. Mol Psychiatry. 2004;9:557–569. doi: 10.1038/sj.mp.4001507. [DOI] [PubMed] [Google Scholar]
- 21.Volkow ND, Wang GJ, Telang F, Fowler JS, Thanos PK, Logan J, Alexoff D, Ding YS, Wong C, Ma Y, Pradhan K. Low dopamine striatal D2 receptors are associated with prefrontal metabolism in obese subjects: possible contributing factors. NeuroImage. 2008;42:1537–1543. doi: 10.1016/j.neuroimage.2008.06.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Marcus MD, Wildes JE. Obesity: Is it a mental disorder? International Journal of Eating Disorders. 2009;42:739–53. doi: 10.1002/eat.20725. [DOI] [PubMed] [Google Scholar]
- 23.O’Malley PM, Johnston LD. Epidemiology of alcohol and other drug use among American college students. J Stud Alcohol. 2002;14:23–39. doi: 10.15288/jsas.2002.s14.23. [DOI] [PubMed] [Google Scholar]
- 24.Knight JR, Wechsler H, Kuo M, Seibring M, Weitzman ER, Schuckit MA. Alcohol abuse and dependence among U.S. college students. J Stud Alcohol. 2002;63(3):263–270. doi: 10.15288/jsa.2002.63.263. [DOI] [PubMed] [Google Scholar]
- 25.American Psychiatric Association. Diagnostic and statistical manual of mental disorders. 4. Washington, DC: 2000. text revision. [Google Scholar]
- 26.Gearhardt AN, Corbin WR, Brownell KD. Preliminary validation of the Yale Food Addiction Scale. Appetite. 2009;52:430–436. doi: 10.1016/j.appet.2008.12.003. [DOI] [PubMed] [Google Scholar]
- 27.Stice E, Yokum S, Blum K, Bohon C. Weight gain is associated with reduced striatal response to palatable food. J Neurosci. 2010;30:13105–13109. doi: 10.1523/JNEUROSCI.2105-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Zald DH, Pardo JV. Cortical activation induced by introral stimulation with water in humans. Chem Senses. 2000;25:267–75. doi: 10.1093/chemse/25.3.267. [DOI] [PubMed] [Google Scholar]
- 29.Wellcome Department of Imaging Neuroscience. London, UK: [Google Scholar]
- 30.Mathworks, Inc.; Sherborn, MA: [Google Scholar]
- 31.Worsley KJ, Marrett S, Neelin P, Vandal AC, Friston KJ, Evans AC. A unified statistical approach for determining signals in images of cerebral activation. Hum Brain Mapp. 1996;4:58–73. doi: 10.1002/(SICI)1097-0193(1996)4:1<58::AID-HBM4>3.0.CO;2-O. [DOI] [PubMed] [Google Scholar]
- 32.O’Doherty JP, Deichmann R, Critchley HD, Dolan RJ. Neural responses during anticipation of a primary taste reward. Neuron. 2002;33:815–826. doi: 10.1016/s0896-6273(02)00603-7. [DOI] [PubMed] [Google Scholar]
- 33.Henson RN, Price CJ, Rugg MD, Turner R, Friston KJ. Detecting latency differences in event-related BOLD responses: Application to words versus nonwords and initial versus repeated face presentations. NeuroImage. 2002;15:83–97. doi: 10.1006/nimg.2001.0940. [DOI] [PubMed] [Google Scholar]
- 34.Gilman JM, Ramchandani VA, Davis MB, Bjork JM, Hommer DM. Why we like to drink: A functional magnetic resonance imaging study of the rewarding and anxiolytic effects of alcohol. J Neurosci. 2008;28:4583–4591. doi: 10.1523/JNEUROSCI.0086-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Risinger RC, Salmeron BJ, Ross TJ, Amen SL, Sanfilipo M, Hoffmann RG, Bloom AS, Garavan H, Stein EA. Neural correlates of high and craving during cocaine self-administration using BOLD fMRI. Neuroimage. 2005;26:1097–1108. doi: 10.1016/j.neuroimage.2005.03.030. [DOI] [PubMed] [Google Scholar]
- 36.Small DM, Zatorre RJ, Dagher A, Evans AC, Jones-Gotman M. Changes in brain activity related to eating chocolate: from pleasure to aversion. Brain. 2001;124:1720–1733. doi: 10.1093/brain/124.9.1720. [DOI] [PubMed] [Google Scholar]
- 37.Friston KJ, Buechel C, Fink GR, Morris J, Rolls E, Dolan RJ. Psychophysiological and modulatory interactions in neuroimaging. Neuroimage. 1997;6:218–229. doi: 10.1006/nimg.1997.0291. [DOI] [PubMed] [Google Scholar]
- 38.Dreher J-S, Schmidt PJ, Kohn P, Furman D, Rubinow D, Berman KF. Menstrual cycle phase modulates reward-related neural function in women. PNAS. 2007;104:2465–70. doi: 10.1073/pnas.0605569104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Van Strien T, Frijters JER, Van Staveren WA, Defares PB, Deurenberg P. The Dutch Eating Behavior Questionnaire for assessment of restrained, emotional and external eating behavior. IJED. 1986;5:295–315. [Google Scholar]
- 40.Cohen J. Statistical power analysis for the behavioral sciences. 2. Hillsdale, NJ: Erlbaum; 1988. [Google Scholar]
- 41.Rolls ET. The orbitofrontal cortex and reward. Cerebral Cortex. 2000;10:284–294. doi: 10.1093/cercor/10.3.284. [DOI] [PubMed] [Google Scholar]
- 42.de Araujo IET, Rolls ET. The representation in the human brain of food texture and oral fat. J Neurosci. 2004;24:3086–3093. doi: 10.1523/JNEUROSCI.0130-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Volkow ND, Fowler JS, Wang G-J, Swanson JM, Telang F. Dopamine in drug abuse and addiction. Arch Neurol. 2007;64:1575–9. doi: 10.1001/archneur.64.11.1575. [DOI] [PubMed] [Google Scholar]
- 44.Heinz A, Siessmeier T, Wrase J, Hermann D, Klein S, Grüsser-Sinopoli SM, Flor H, Braus DF, Buchholz HG, Gründer G, Schreckenberger M, Smolka MN, Rösch F, Mann K, Bartenstein P. Correlation between dopamine D2 receptors in the ventral striatum and central processing of alcohol cues and craving. Am J Psychiatry. 2004;161:1783–1789. doi: 10.1176/appi.ajp.161.10.1783. [DOI] [PubMed] [Google Scholar]
- 45.Grüsser SM, Wrase J, Klein S, Hermann D, Smolka MN, Ruf M, Weber-Fahr W, Flor H, Mann K, Braus DF, Heinz A. Cue-induced activation of the striatum and medial prefrontal cortex is associated with subsequent relapse in abstinent alcoholics. Psychopharmacology. 2004;175:296–302. doi: 10.1007/s00213-004-1828-4. [DOI] [PubMed] [Google Scholar]
- 46.Goldstein RZ, Tomasi D, Alia-Klein N, Cottone LA, Zhang L, Telang F, Volkow ND. Subjective sensitivity to monetary gradients is associated with frontolimbic activation to reward in cocaine abusers. Drug Alcohol Depend. 2007;87:233–40. doi: 10.1016/j.drugalcdep.2006.08.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Arana FS, Parkinson JA, Hinton E, Holland AJ, Owen AM, Roberts AC. Dissociable contributions of the human amygdala and orbitofrontal cortex to incentive motivation and goal selection. J Neurosci. 2003;23:9632–9638. doi: 10.1523/JNEUROSCI.23-29-09632.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Petrides M. Frontal lobes and working memory: evidence from investigations of the effects of cortical excisions in nonhuman primates. In: Boller F, Grafman J, editors. Handbook of Neuropsychology. Elsevier; Amsterdam: 1994. pp. 59–82. [Google Scholar]
- 49.Heller W. Emotion. In: Banich MT, editor. Cognitive neuroscience and neuropsychology. Boston, MA: Houghton Mifflin Company; 2004. pp. 393–428. [Google Scholar]
- 50.Hare TA, Camerer CF, Rangel A. Self-control in decision-making involves modulation of the vmPFC valuation system. Science. 2009;324:646–648. doi: 10.1126/science.1168450. [DOI] [PubMed] [Google Scholar]
- 51.Kawagoe R, Takikawa Y, Hikosaka Reward-predicted activity of dopamine and caudate neurons – a possible mechanism of motivational control of saccadic eye movement. J Neurophysiol. 2004;91:1013–1024. doi: 10.1152/jn.00721.2003. [DOI] [PubMed] [Google Scholar]
- 52.Delagado MR, Stenger VA, Fiez JA. Motivation-dependent responses in the human caudate nucleus. Cerebral Cortex. 2004;14:1022–1033. doi: 10.1093/cercor/bhh062. [DOI] [PubMed] [Google Scholar]
- 53.Garavan H, Pankiewicz J, Bloom A, Cho J, Sperry L, Ross TJ, Salmeron BJ, Risinger R, Kelley D, Stein EA. Cue-induced cocaine craving: neuroanatomical specificity for for drug users and drug cocaine craving: neuroanatomical specificity for drug users and drug stimuli. Am J Psychiatry. 2000;157:1789–1798. doi: 10.1176/appi.ajp.157.11.1789. [DOI] [PubMed] [Google Scholar]
- 54.Grant S, London ED, Newlin DB, Villemagne VL, Liu X, Contoreggi C, Phillips RL, Kimes AS, Margolin A. Activation of memory circuits during cue-elicited cocaine craving. Proc Natl Acad Sci USA. 1996;93:12040–12045. doi: 10.1073/pnas.93.21.12040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Wang G-J, Volkow ND, Fowler JS, Cervany P, Hitzemann RJ, Pappas N, Wong CT, Felder C. Regional brain metabolic activation during craving elicited by recall of previous drug experiences. Life Sci. 1999;64:775–784. doi: 10.1016/s0024-3205(98)00619-5. [DOI] [PubMed] [Google Scholar]
- 56.Wilson SJ, Sayette MA, Fieze JA. Prefrontal responses to drug cues: a neurocognitive analyses. Nat Neurosci. 2004;7:211–214. doi: 10.1038/nn1200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Childress AR, Mozley PD, McElgin W, Fitzgerald J, Reivich M, O’Brien CP. Limbic activation during cue-induced cocaine craving. Am J Psychiatry. 1999;156:11–18. doi: 10.1176/ajp.156.1.11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Pelchat ML, Johnson A, Chan R, Valdez J, Ragland JD. Images of desire: food-craving activation during fMRI. Neuroimage. 2004;23:1486–1493. doi: 10.1016/j.neuroimage.2004.08.023. [DOI] [PubMed] [Google Scholar]
- 59.Modell JG, Mountz JM. Focal cerebral blood flow change during craving for alcohol measured by SPECT. J Neuropsychiatry Clin N. 1995;7:15–22. doi: 10.1176/jnp.7.1.15. [DOI] [PubMed] [Google Scholar]
- 60.Berridge KC, Kringlebach ML. Affective neuroscience of pleasure: Reward in humans and animals. Psychopharmacology. 2008;199:457–480. doi: 10.1007/s00213-008-1099-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Boettiger CA, Mitchell JM, Tavares VC, Robertson M, Joslyn G, D’Esposito M, Fields HL. Immediate reward bias in humans: Fronto-parietal networks and a role for the catechol-O-methyltransferase. J Neurosci. 2007;27:14383–14391. doi: 10.1523/JNEUROSCI.2551-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Elliot R, Dolan RJ, Frith CD. Dissociable functions in the medial and lateral orbitofrontal cortex: Evidence from human neuroimaging studies. Cerebral Cortex. 2000;10:308–317. doi: 10.1093/cercor/10.3.308. [DOI] [PubMed] [Google Scholar]
- 63.Chiamulera C. Cue reactivity in nicotine and tobacco dependence: a “multiple-action” model of nicotine as primary reinforcement and as an enhancer of the effects of smoking-associated stimuli. Brain Res Rev. 2005;48:74–97. doi: 10.1016/j.brainresrev.2004.08.005. [DOI] [PubMed] [Google Scholar]
- 64.Shalev U, Grimm JW, Shaham Y. Neurobiology of relapse to heroin and cocaine seeking: a review. Pharmacol Rev. 2002;54:1–42. doi: 10.1124/pr.54.1.1. [DOI] [PubMed] [Google Scholar]
- 65.Robinson TE, Berridge KC. Incentive-sensitization and addiction. Addiction. 2001;96:103–114. doi: 10.1046/j.1360-0443.2001.9611038.x. [DOI] [PubMed] [Google Scholar]
- 66.Robinson TE, Berridge KC. The psychology and neurobiology of addiction: An incentive-sensitization view. Addiction. 2000;95:91–117. doi: 10.1080/09652140050111681. [DOI] [PubMed] [Google Scholar]
- 67.Fillmore MT, Rush CR. Alcohol effects on inhibitory and activational response strategies in the acquisition of alcohol and other reinforcers: priming the motivation to drink. J Stud Alc. 2001;62:646–656. doi: 10.15288/jsa.2001.62.646. [DOI] [PubMed] [Google Scholar]
- 68.Fillmore MT. Cognitive preoccupation with alcohol and binge drinking in college students: alcohol-induced priming of the motivation to drink. Psychol Addict Behav. 2001;15:325–332. [PubMed] [Google Scholar]
- 69.Fedoroff IDC, Polivy J, Herman CP. The effect of pre-exposure to food cues on the eating behavior of restrained and unrestrained eaters. Appetite. 1997;28:33–47. doi: 10.1006/appe.1996.0057. [DOI] [PubMed] [Google Scholar]
- 70.Jansen A, van den Hout M. On being led into temptation: “Counterregulation” of dieters after smelling a “preload”. Addictive Behaviors. 1991;16:247–253. doi: 10.1016/0306-4603(91)90017-c. [DOI] [PubMed] [Google Scholar]
- 71.Rogers PJ, Hill AJ. Breakdown of dietary restraint following mere exposure to food stimuli: Interrelationships between restraint, hunger, salivation, and food intake. Addictive Behaviors. 1989;14:387–397. doi: 10.1016/0306-4603(89)90026-9. [DOI] [PubMed] [Google Scholar]
- 72.Führer D, Zysset S, Stumvoll M. Brain activity in hunger and satiety: an exploratory visually stimulated fMRI study. Obesity. 2008;16:945–950. doi: 10.1038/oby.2008.33. [DOI] [PubMed] [Google Scholar]
- 73.Siep N, Roefs A, Roebroeck A, Havermans R, Bonte ML, Jansen A. Hunger is the best spice: An fMRI study of the effects of attention, hunger and calorie content on food reward processing in the amygdala and orbitofrontal cortex. Behav Brain Res. 2009;109:149–158. doi: 10.1016/j.bbr.2008.10.035. [DOI] [PubMed] [Google Scholar]
- 74.Berridge KC, Ho C-Y, Richard JM, DiFeliceantonio AG. The tempted brain eats: pleasure and desire circuits in obesity and eating disorders. Brain Res. 2010;1350:43–64. doi: 10.1016/j.brainres.2010.04.003. [DOI] [PMC free article] [PubMed] [Google Scholar]