

Summary
To combat potentially deadly viral infections, prokaryotic microbes enlist small RNA-based adaptive immune systems (CRISPR-Cas systems) that protect through sequence-specific recognition and targeted destruction of viral nucleic acids (either DNA or RNA depending on the system). Here, we summarize rapid progress made in redirecting the nuclease activities of these microbial immune systems to bind and cleave DNA or RNA targets of choice, by reprogramming the small guide RNAs of the various CRISPR-Cas complexes. These studies have demonstrated the potential of Type II CRISPR-Cas systems both as efficient and versatile genome editing tools and as potent and specific regulators of gene expression in a very broad range of cell types (including human) and organisms. Progress is also being made in developing a Type III RNA-targeting CRISPR-Cas system as a novel gene knockdown platform to investigate gene function and modulate gene expression for metabolic engineering in microbes.
Keywords: CRISPR, Cas, genome editing, RNA silencing, biotechnology, metabolic engineering
CRISPR-Cas: Prokaryotic small RNA-based defense systems
Prokaryotic microbes (bacteria and archaea) are under constant attack by potentially deadly viruses and other harmful invading mobile genetic elements. The viruses evolve rapidly and are estimated to outnumber the prokaryotes 10 to 1[1]. Not surprisingly, prokaryotes have evolved a variety of innate defense mechanisms to ward off these predators (reviewed in [2–5]). The CRISPR-Cas systems are the most recently discovered armament in the prokaryotic arsenal (recently reviewed in [4, 6–9]). The unique features of these RNA-guided adaptive immune systems have captivated the scientific community, and the molecular mechanisms that underlie the pathways (Figure 1) are the subject of intense research. In turn, the newly acquired knowledge is being rapidly deployed in the development of exciting new CRISPR-based technologies with important biotechnological, industrial, and biomedical applications.
Figure 1.

Overview of the CRISPR-Cas pathways. To obtain the ability to recognize and silence a specific virus or other foreign genetic element, a short fragment of invader DNA is acquired and integrated into the host CRISPR locus near the leader (Adaptation). The CRISPR locus contains copies of a short direct repeat sequence (black) that separate the invader-derived sequences (multiple colors). Cas genes encoding the protein components of the system are typically encoded adjacent to CRISPR loci (not shown). Protospacer adjacent motifs (PAMs) are found immediately adjacent to viral sequences selected for CRISPR integration (protospacers). CRISPR locus transcripts are processed to generate multiple individual crRNAs (crRNA Biogenesis). Mature crRNAs retain repeat sequence at the 5′ end (shown) and/or 3′ end of the invader-derived guide sequence (colored region). crRNAs associate with 1 to 6 Cas proteins (depending on the system) to form crRNA-Cas protein (crRNP) effector complexes. The crRNP specifically recognizes foreign DNA or RNA (depending on the system) via base-pairing of the crRNA and cleaves (Invader Silencing). PAMs provide important auxiliary signals for the recognition and silencing of invaders for some Type I and II DNA-targeting systems.
Components of the multiple types of CRISPR-Cas systems: Arsenal inventory
At least eleven distinct CRISPR-Cas systems have been identified [4, 6–11]. The systems can be grouped into three broad classes, Types I, II and III, which include eleven distinct subtypes: Types I A–F, Types II A–C and Types III A and B [6, 11–13] (Table 1). Each of the systems and its specific Cas protein components are referred to by a name derived from a model organism for the system (e.g., Cse or Cas subtype ME. coli for Type I–E) [10, 14]. Most of the CRISPR-Cas systems that have been characterized target invader DNA [4, 15–21]; however, the Type III-B Cmr system cleaves target RNAs [22–26].
Table 1.
Multiple CRISPR-Cas systems with distinct crRNAs, Cas proteins, and mechanism of action against target nucleic acids.
| CRISPR/CAS system | crRNA | Known or predicted Cas proteins of effector crRNP | Target | Effector Nuclease | |
|---|---|---|---|---|---|
| I | A | 251658240 |
Cas8a, Csa5, Cas7, Cas5, Cas3′+″ | DNA | Cas3 |
| B | Cas8b, Cas7, Cas5, Cas3 | ||||
| C | Cas8c, Cas7, Cas5, Cas3 | ||||
| D | Cas10d, Csc1, Csc2, | ||||
| E | 251658240 |
Cse1-2, Cas7, Cas5, Cas6e, Cas3 | |||
| F | Csy1-3, Cas6y, Cas2/3 | ||||
| II | A,B,C | 251658240
|
Cas9 (+ tracrRNA) | DNA | Cas9 |
| III | A | 251658240 |
Csm1(Cas10a)-Csm5 | DNA | ? |
| B | Cmr1, Cmr2 (Cas10b), Cmr3-6 | RNA | ? | ||
The guide RNAs of the CRISPR-Cas systems arise from the CRISPRs (clustered regularly interspaced short palindromic repeats): loci present in prokaryotic genomes that contain tandem arrays of a short direct repeat sequence (~30–40 basepairs) separated by a series of similarly-sized “spacer” sequences acquired from viruses and other invaders (where the same sequence is known as a “protospacer”) [16, 27–31] (Figure 1). Transcription of CRISPR loci and subsequent processing of CRISPR transcripts produces mature CRISPR (cr)RNAs that each contain an invader-derived (guide) sequence flanked by the signature CRISPR repeat (tag or handle) sequence at the 5′ and/or 3′ end, depending on the system [24, 32–38].
crRNAs interact with CRISPR-associated (Cas) proteins to form ribonucleoprotein (crRNP) effector complexes that recognize (via crRNA basepairing) and cleave the DNA or RNA of the invader [17, 18, 23, 32, 39–43](Figure 1). Each CRISPR-Cas system includes a characteristic module of Cas proteins typically encoded near the CRISPR locus (Table 1). Some of the Cas proteins are members of broad superfamilies found in multiple CRISPR-Cas systems [10, 11, 14]. Cas3 and Cas9 superfamily proteins are known effector nucleases (i.e., DNases that destroy the invader DNA) for Type I and Type II systems, respectively [17, 18, 27, 32, 44–46]. The identity of the effector nuclease component of Type III CRISPR-Cas systems is not yet known.
Exploiting the CRISPR-Cas systems: strain surveillance and protection
The discovery and understanding of the CRISPR-Cas systems used by prokaryotes to defend themselves against invaders encountered in their environments is being leveraged in a number of ways [47–49]. CRISPR locus sequences can evolve rapidly in response to local predators and thus vary among isolates of related strains. This feature is being exploited for fast and precise strain identification in epidemiological investigations (e.g., to track pathogenic strains involved in disease outbreaks) and to probe microbial diversity and host-virus interactions in natural environments including human microbiomes [50–55].
A variety of bacteria and archaea are employed to make important products, including pharmaceuticals, biofuels, commodity chemicals, and food. Unique CRISPR locus sequences can be used to tag and identify valuable industrial strains [47]. Moreover, priming or manipulation of CRISPR-Cas systems can generate phage-resistant commercial strains, improving product quality and reducing economic losses due to culture infections [51]. Resistant strains can be isolated by deliberate exposure to phages and selection of strains with arrays of phage-targeting sequences in their CRISPR loci [51]. Alternatively, resistant strains can be deliberately engineered by introduction of invader sequences into existing CRISPR loci or by introduction of entire CRISPR-Cas modules into strains lacking defense [56, 57].
As described below, individual CRISPR-Cas pathway enzymes are also being repurposed as powerful weapons for the research and medical fronts.
Repurposing Type II CRISPR-Cas effector complexes: powerful new RNA-guided genome editing tools
The ability to eliminate, replace, or modify sequences within a genome in a highly targeted manner (genome editing) has a broad range of important basic and applied applications including fundamental gene discovery, plant and microbe bioengineering and correction of human gene disorders (gene therapy). Targeted gene editing in vivo can be accomplished through ectopic expression of artificial protein nucleases (typically, zinc finger nucleases (ZFNs) or transcription activator-like effector nucleases (TALENs)) individually designed to recognize a specific target DNA site and introduce a double-stranded (ds)DNA break [8, 58–60]. Different types of mutations can be obtained depending upon which endogenous DNA repair pathway is engaged to repair the dsDNA break. If the dsDNA break occurs in the presence of a co-introduced, editing template (donor DNA fragment that shares homology with target DNA), this can trigger DNA repair by the high fidelity, homologous recombination (HR) pathway to replace, remove, or alter/correct the target gene in a manner specified by the editing template. In the absence of an editing template, cells typically repair the dsDNA break by the relatively error-prone mechanism of non-homologous end joining (NHEJ), which typically leads to short gene insertions or deletions (indels) and thus can yield gene inactivation mutations.
The “Cas9” Type II CRISPR-Cas immune system effector complex has recently emerged as a powerfully adaptable, next-generation genome editing tool: an attractive alternative to the development of individual protein enzymes to induce individual genome modifications (Figure 2) [48, 61–63]}. In a short period of time, a hail of studies have reported successful RNA-guided genome editing with Cas9 crRNP complexes in diverse cell types and model organisms, including several cultured human cell lines [63–75], rodents [69, 76–80], zebra fish [81–84], frog [85, 86], plants [86–93], fruit fly [94–99], worm [100–108], yeast [109] and bacteria [110]. Cas9-based genome editing systems have been used for large scale (including genome-wide) loss-of-function genetic screens in cultured human cells [74, 75] as well as for correction of disease-associated genetic mutations in adult human stem cell-derived organoids and mouse zygotes [73, 80].
Figure 2.
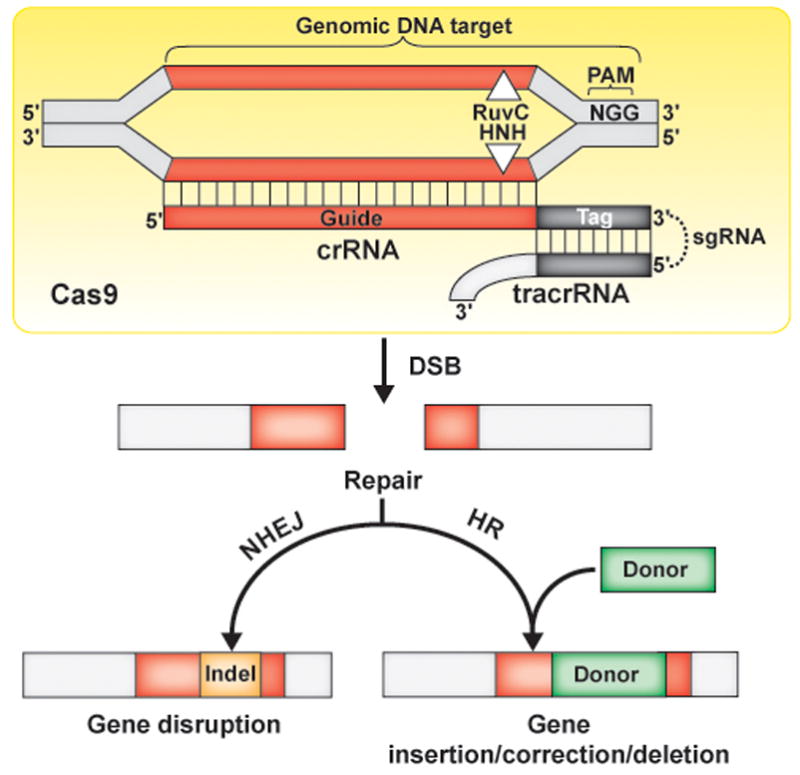
RNA-programmable genome editing by Type II Cas9 CRISPR-Cas systems. The Cas9 endonuclease (yellow) has two nuclease active sites (RuvC and HNH) that cleave opposing strands of a DNA molecule to generate a blunt-ended, double-strand DNA break (DSB). The cleavage is directed by the associated crRNA that contains a guide region that recognizes one strand of the target sequence by base-pairing. The enzyme requires a short protospacer adjacent motif (PAM) immediately adjacent to the region of homology as an auxiliary target identification signal. Cas9 activity also requires a second RNA (tracrRNA) that base-pairs with the repeat-derived tag of the crRNA. The crRNA and tracrRNA can be combined through covalent linkage into a single-guide RNA (sgRNA). In human and many other cell types, the Cas9-induced, double-strand DNA breaks undergo repair by either non-homologous end joining (NHEJ) or homologous recombination (HR). NHEJ repair results in indels (short insertions or deletions) and provides a means to selectively disrupt or inactivate target genes. When provided with a homologous donor DNA, HR repair can affect sequence replacement to insert, modify or delete sequence at the site.
Type II CRISPR-Cas system effector complexes are comprised of a single protein: Cas9, an RNA-guided endonuclease that catalyzes blunt-ended dsDNA breaks [16–18, 35] (Figure 2). The system utilizes two RNAs: a crRNA that guides Cas9 to DNA target sequences via base-pairing and a tracrRNA that pairs with the crRNA and plays an unspecified but essential role in target DNA cleavage [18, 35]. The two essential RNAs (crRNA and tracrRNA) can be experimentally joined (by a connecting RNA loop) into a single guide RNA (sgRNA) [18] to simplify the system to just one protein (Cas9) and one RNA (sgRNA).
The only known constraint on target selection for Cas9 crRNPs is the requirement for a very short sequence motif known as the PAM (protospacer adjacent motif) immediately downstream of the desired target site in the genome (protospacer). The most widely used Cas9 enzyme is derived from the bacterium Streptococcus pyogenes, and its activity simply requires a 5′-GG-3′ (or, with reduced activity, 5′-AG-3′) PAM sequence located one base-pair downstream of the target site [13, 18, 63, 65, 110, 111]. Cas9 from Streptococcus thermophilus [13, 17, 46, 65, 112, 113]} Neisseria meningitidus [13, 67, 113]} and Treponema denticola [113] require different PAMs (5′-NNAGAAW-3′, 5′-NNNNGA/CTT-3′, 5′-NAAAAN-3′ respectively), providing additional options for targeting a specific genome site. Codon-optimized versions of Cas9 are used to facilitate expression of the protein in heterologous cells or organisms, and nuclear localization signals are required for function when eukaryotic species are used.
RNA-directed Cas9 genome editing enzymes have been found to perform with comparable and typically improved efficiencies relative to ZFNs or TALENs in side-by-side comparisons in multiple systems [63–66, 81]. Moreover, Cas9 RNA-based genome editing offers several advantages over protein-guided ZFN and TALEN systems. Targeting new DNA sites with Cas9 simply entails designing an sgRNA that is complementary to the target DNA making the approach much faster, cheaper, and easier to implement (as compared to developing a new ZNF and TALEN protein for each target site). In addition, the Cas9 platform allows simultaneous editing of more than one site (multiplexing) through expression of more than one guide crRNA [65, 66, 110]. The multiplex approach has been used to create mice carrying five different mutant genes in a single step [79], for example, and to introduce large deletions of genomic regions by directing Cas9 cleavages at the two sites flanking the desired deletion [65]. The independent mechanisms by which Cas9 cuts the two strands of a DNA target also provide a distinct advantage. Cas9 employs different nuclease active sites (RuvC and HNH nuclease domains) to cut each DNA strand (Figure 2), which can be independently inactivated (by mutation) to generate a Cas9 enzyme that cleaves one strand [11, 17, 18, 46 Jinek, 2013 #216]. Single-strand nicking variants of Cas9 used for genome editing have been found to lower NHEJ (error prone) DNA repair and favor more predictable changes made as the results of HR DNA repair in human cells [65, 66]. Thus, wildtype Cas9 can be used for double-stranded DNA cleavage, while active site mutants (RuvC or HNH) cleave a single strand, offering a way of reliably introducing predictable mutations into genomes (Table 2).
Table 2.
Cas9 variants for genome editing or gene expression control applications.
| Cas9 | Activity | Application |
|---|---|---|
| Wild-type (Cas9 RuvC+, HNH+) | Double-stranded DNA break (cleavage) | Genome editing |
| Single active site mutants (Cas9 RuvC−, HNH+or Cas9 RuvC+, HNH−) | Single-stranded DNA break (nickase) | Genome editing |
| Double active site mutant (Cas9 RuvC−, HNH−) | DNA binding (no cutting) | Gene knockdown/interference (CRISPRi) |
| Double active site mutants with fusion proteins (Cas9, RuvC−, HNH− & transcription activator or repressor domain) | DNA binding (no cutting) & transcriptional control | Modulation of gene expression (activation or repression) |
Weapons modification: nuclease-defective Cas9 enzymes developed to control transcription
Inactivation of both nuclease domains of Cas9 (by double mutation) produces a protein capable of RNA-guided transcription repression in E. coli, and less reliably, in a human cell line [114]. This novel platform, dubbed “CRISPRi” (for CRISPR interference), appears to function via the Cas9 complex binding (but not cleaving) the DNA target site and blocking transcription initiation or elongation (Table 2). Targeting (of either DNA strand) at key transcriptional promoter elements results in significant impairment of gene expression (presumably blocking initiation) [114]. By contrast, targeting only the non-template strand of intragenic sites results in gene expression repression (presumably blocking elongation). An innovative inducible Cas9-based CRISPRi system permitted controlled and reversible target gene inhibition in E. coli. Targeting expression of regulatory and structural genes of the well-characterized lactose utilization pathway produced expected phenotypes, demonstrating the effectiveness and utility of the CRISPRi knockdown technology [114]. The nuclease-defective Cas9 is also capable of activating (rather than repressing) gene expression when directed (by the sgRNA) to interact with transcriptional repressor binding sites [114].
The Cas9 complex has been further modified by the fusion of specific transcription activators or repressors to the nuclease-defective protein to precisely activate or silence expression of a target gene (Table 2) [115–118]. These “CRISPR transcription factors” (Cas9-transcriptional effector domain fusion proteins) can be programmed with sgRNAs that recognize specific transcription control elements to obtain predictable effects on endogenous gene expression at targeted loci. The effects can be modulated by multiplex targeting (employing multiple sgRNAs) at different positions along the promoter region of the gene [115, 116]. CRISPR transcription factors have been successfully employed in human [115–117], mouse [115], yeast [116, 117], and bacterial [118] cells.
The Cas9 complex provides a versatile platform for RNA-guided delivery of various agents and Cas9 fusion proteins with other functionalities will certainly be developed. For example, epigenetic effector domain fusions could be constructed, such as a methyltransferase fusion for RNA-guided DNA methylation or a histone modifying enzyme fusion to effect gene-specific changes in chromatin organization. In addition, multiple Cas9-based tools can be independently deployed in a single cell (e.g., to activate transcription of one gene and inhibit transcription of another) using Cas9 proteins from different organisms and corresponding sgRNAs (which differ in the repeat and tracrRNA sequences) [13, 113].
Assessment of target specificity of Cas9 systems: Avoiding collateral damage
Clearly, the various RNA-programmable Cas9 systems hold tremendous promise in site-specific genome editing and gene expression control applications. A critical issue that remains to be thoroughly addressed is the degree to which off-target effects (cleavage or binding of unintended DNA sequences) occur in the various cellular contexts where these Cas9-based tools are being used and how these can be mitigated.
Both the homology between the ~20 nucleotide guide region of the sgRNA and the requirement for the immediately adjacent PAM motif in the target DNA contribute to the specificity of the Cas9 reagents, however some off-target mutations have been documented. DNA cleavage has been observed at a few unintended sites with various extents of sgRNA/target sequence mismatch in mammalian cells [70, 72]. Studies aimed at deciphering Cas9 targeting rules indicate that extensive - but not necessarily 100% – homology with the sgRNA guide element is required for targeting [17, 18, 65, 69–71, 110, 112, 119]. The most recent in-depth studies reveal that Cas9-sgRNA targeting is quite complex, with mismatch tolerance depending upon the exact number, type, and position of RNA/DNA mismatches and varying for different sgRNA/target combinations [65, 69–71, 110, 119]. The mismatch tolerance of the Cas9 complex may be advantageous for silencing rapidly evolving viral target sequences.
Evidence indicates that off-target effects in Cas9 applications can be reduced by judicious choice of target sites (guided by recently established targeting rules) and by optimizing the expression levels of the Cas9-sgRNA complexes [71]. In addition, a “double-nicking” approach has been developed to reduce the possibility of off-target effects. Single active site mutants of Cas9 are used with two distinct sgRNAs to simultaneously generate offset single-strand nicks (yielding a 5′ overhang) on each of the opposing two strands of the target sequence [69, 111]. This co-nicking strategy appears to generate double-stranded breaks with efficiency comparable to wildtype Cas9 (with a single sgRNA), but with greatly improved target site specificity [69].
Another weapon in the arsenal: deploying the Type III effector complex for gene knockdown
Another CRISPR-Cas effector complex is in development for applied RNA-guided gene knockdown. The Type III-B Cmr complex is the only CRISPR-Cas complex so far found to target RNAs [22, 23] (Figure 3). Functional Cmr complexes can be reconstituted with the six recombinant Cmr proteins (Cmr1-6) and synthetic crRNAs [23]. The crRNAs have an essential 8-nucleotide 5′ tag sequence (derived from the CRISPR repeat) and can have guide regions of various lengths [23, 24]. crRNAs found in native Cmr complexes in P. furiosus contain 31- and 37-nucleotide guide elements [23, 24]. The Cmr complexes specifically cleave RNA targets in the region base-paired by the crRNA guide sequence [22–25]. Identifying the nuclease component may allow further refinement of this complex as a tool. The overall architecture and subunit organization of functional Cmr complexes was recently revealed by a combination of cryo electron microscopy and biochemical analyses [22, 25, 26].
Figure 3.

RNA-programmable gene knockdown by Type III-B Cmr CRISPR-Cas systems. The Cmr effector complex (blue) consists of six Cmr proteins and a crRNA. The guide region of the crRNA base-pairs with the homologus sequence in the mRNA target and the target RNA is cleaved by the complex. Applications of the Cmr system include RNA-directed gene knockdown to investigate gene function or to facilitate metabolic engineering.
The Cmr complex can be programmed to cleave selected target RNAs using customized crRNAs in vitro [24] suggesting that Cmr complexes can be engineered to destroy selected mRNAs and manipulate gene expression similar to eukaryotic RNAi technologies (Figure 3). As a basic research tool, the Cmr technology has the potential to rapidly advance gene discovery and pathway analysis (especially in prokaryotes which lack the RNAi machinery naturally present in many eukaryotic systems). crRNA-mediated knockdowns would allow rapid screening of multiple gene targets in parallel. In industry, the potential of the Cmr system to simultaneously modulate the expression of several genes in a pathway could be of tremendous value in metabolic engineering of microbes to optimize production of biofuels, pharmaceuticals, foods, etc. Relative to DNA-targeting technologies, the Cmr system offers more straightforward capabilities for partial and temporary gene knockdown. Furthermore, the Cmr complex appears not to have target site limitations imposed by the requirement for a PAM sequence flanking the target sequence (unlike Cas9-based reagents and other DNA targeting CRISPR-Cas systems) [24]. Although the potential for off-target effects has not been experimentally tested, the Cmr complexes employ longer guide sequences for target recognition than the Cas9-based reagents.
Enlistment of crRNA biogenesis nucleases: novel endoribonucleases
The site-specific RNA cleavage capabilities of Cas6 superfamily crRNA processing endoribonucleases are also being exploited as valuable research tools. These protein enzymes recognize and cleave at specific sites within the CRISPR repeat sequences in CRISPR locus transcripts to release individual crRNAs [32, 34, 38]. There are multiple potential applications for the Cas6 site-specific endoribonucleases. The Cas6y (or Csy4) nuclease has been used to optimize protein expression in E. coli expression systems [120]. Cleavages at Cas6y sites inserted upstream of the ribosomal binding sites in monocistronic or polycistronic mRNAs leads to significantly improved predictable expression [120]. An inventive, cleavable RNA affinity tagging procedure to isolate and purify RNA-associated proteins has also being developed using Cas6y [121]. Using the related Cas6e (or Cse3) enzyme, this general approach has been adapted to generate T7 in vitro transcripts with homogenous 5′ ends [122].
Concluding remarks
The effector complexes of the CRISPR-Cas immune systems utilize guide RNAs to effectively recognize and target foreign DNAs or RNAs for destruction. RNA-guided recognition makes these immune systems highly and rapidly adaptable to diverse targets – recognizing a new target simply requires a new guide RNA sequence (that can be obtained directly from the invader). This same attribute makes the CRISPR-Cas effector complexes exceptionally flexible and accessible tools for the multiple applications to which they can be adapted, which currently include incredibly valuable uses in genome editing and modulation of gene expression. We are currently witnessing the power of a naturally evolved prokaryotic defense system being repurposed to obtain effective new weapons for industry, research and medicine. Notably, our understanding of the multiple CRISPR-Cas systems is still far from complete and additional tools and applications are yet to come from these fertile research grounds.
Highlights.
CRISPR-Cas systems use RNA-guided effector complexes to destroy invading nucleic acid
Powerful technologies are rapidly arising from these prokaryotic defense systems
Uses range from pathogen tracking to genetic defect repair to metabolic engineering
The RNA-guided effector complexes offer particularly adaptable application platforms
Acknowledgments
This work was funded by NIH grants RO1 GM54682 and RO1 GM99876 to R.M.T. and M.P.T. We thank Claiborne Glover (University of Georgia) and members of the Terns lab for critical review of the manuscript.
Glossary
- Cas
CRISPR-associated. Refers to a conserved gene or corresponding protein, that functions with (and is often encoded adjacent to) the CRISPR locus as part of a CRISPR-Cas immune system that protects prokaryotes from viruses and other molecular invaders.
- CRISPR
Clustered regularly interspaced short palindromic repeat. A CRISPR locus consists of a regular array of copies of a short direct repeat interspaced with similarly-sized sequences that typically originate from viruses or plasmids termed spacers.
- crRNA
CRISPR RNA. Small non-coding RNAs that are produced by transcription and processing of CRISPR transcripts. crRNAs contain guide sequence (complementary to an invasive nucleic acid) and a CRISPR-repeat-derived sequence or tag at the 5′ and/or 3′ end.
- crRNA tag
CRISPR RNA repeat-derived sequence present at the 5′ and/or 3′ end of a mature crRNA. The tag element contributes to specific assembly of crRNPs and is required for invader silencing. Also referred to as handles in the literature.
- crRNP
A complex between a crRNA and cognate Cas proteins responsible for recognition and defense against invading nucleic acids.
- Protospacer
The sequence in the foreign target DNA that corresponds with the (spacer) sequence integrated into the CRISPR locus and is recognized by the guide region of the encoded crRNA in the DNA targeting systems.
- PAM
Protospacer adjacent motif. A short (typically 2–5 nucleotide) conserved sequence element that is located immediately adjacent to the protospacer in the foreign target DNA. This motif (or a close sequence variant) is important for target DNA identification and cleavage in Type I and II CRISPR-Cas systems.
- sgRNA
An engineered chimeric RNA in which the crRNA and tracrRNA of Type II CRISPR-Cas systems are linked in a single molecule. Capable of directing Cas9 to cleave a corresponding target DNA sequence.
- Spacer
Historical term for a DNA sequence found between the repeat elements of the CRISPR locus. Spacer sequences in the CRISPR locus match protospacer sequences in invading DNAs and encode the guide elements of the crRNAs.
- tracrRNA
Trans-activating crRNA. A small non-coding RNA of Type II CRISPR-Cas systems that base-pairs with the 3′ tag of the crRNA and is required for Cas9 DNA targeting activity.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Hatfull GF. Bacteriophage genomics. Current opinion in microbiology. 2008;11:447–453. doi: 10.1016/j.mib.2008.09.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Labrie SJ, et al. Bacteriophage resistance mechanisms. Nature reviews Microbiology. 2010;8:317–327. doi: 10.1038/nrmicro2315. [DOI] [PubMed] [Google Scholar]
- 3.Stern A, Sorek R. The phage-host arms race: shaping the evolution of microbes. BioEssays : news and reviews in molecular, cellular and developmental biology. 2011;33:43–51. doi: 10.1002/bies.201000071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Westra ER, et al. The CRISPRs, they are a-changin’: how prokaryotes generate adaptive immunity. Annual review of genetics. 2012;46:311–339. doi: 10.1146/annurev-genet-110711-155447. [DOI] [PubMed] [Google Scholar]
- 5.Samson JE, et al. Revenge of the phages: defeating bacterial defences. Nature reviews Microbiology. 2013;11:675–687. doi: 10.1038/nrmicro3096. [DOI] [PubMed] [Google Scholar]
- 6.Sorek R, et al. CRISPR-mediated adaptive immune systems in bacteria and archaea. Annual review of biochemistry. 2013;82:237–266. doi: 10.1146/annurev-biochem-072911-172315. [DOI] [PubMed] [Google Scholar]
- 7.Terns MP, Terns RM. CRISPR-based adaptive immune systems. Current opinion in microbiology. 2011;14:321–327. doi: 10.1016/j.mib.2011.03.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Bhaya D, et al. CRISPR-Cas systems in bacteria and archaea: versatile small RNAs for adaptive defense and regulation. Annual review of genetics. 2011;45:273–297. doi: 10.1146/annurev-genet-110410-132430. [DOI] [PubMed] [Google Scholar]
- 9.Wiedenheft B, et al. RNA-guided genetic silencing systems in bacteria and archaea. Nature. 2012;482:331–338. doi: 10.1038/nature10886. [DOI] [PubMed] [Google Scholar]
- 10.Haft DH, et al. A guild of 45 CRISPR-associated (Cas) protein families and multiple CRISPR/Cas subtypes exist in prokaryotic genomes. PLoS Comput Biol. 2005;1:e60. doi: 10.1371/journal.pcbi.0010060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Makarova KS, et al. Evolution and classification of the CRISPR-Cas systems. Nature reviews Microbiology. 2011;9:467–477. doi: 10.1038/nrmicro2577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Chylinski K, et al. The tracrRNA and Cas9 families of type II CRISPR-Cas immunity systems. RNA Biol. 2013;10:726–737. doi: 10.4161/rna.24321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Fonfara I, et al. Phylogeny of Cas9 determines functional exchangeability of dual-RNA and Cas9 among orthologous type II CRISPR-Cas systems. Nucleic acids research. 2013 doi: 10.1093/nar/gkt1074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Makarova KS, et al. A putative RNA-interference-based immune system in prokaryotes: computational analysis of the predicted enzymatic machinery, functional analogies with eukaryotic RNAi, and hypothetical mechanisms of action. Biology direct. 2006;1:7. doi: 10.1186/1745-6150-1-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Marraffini LA, Sontheimer EJ. CRISPR interference limits horizontal gene transfer in staphylococci by targeting DNA. Science. 2008;322:1843–1845. doi: 10.1126/science.1165771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Garneau JE, et al. The CRISPR/Cas bacterial immune system cleaves bacteriophage and plasmid DNA. Nature. 2010;468:67–71. doi: 10.1038/nature09523. [DOI] [PubMed] [Google Scholar]
- 17.Gasiunas G, et al. Cas9-crRNA ribonucleoprotein complex mediates specific DNA cleavage for adaptive immunity in bacteria. Proceedings of the National Academy of Sciences of the United States of America. 2012;109:E2579–2586. doi: 10.1073/pnas.1208507109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Jinek M, et al. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science. 2012;337:816–821. doi: 10.1126/science.1225829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Fischer S, et al. An Archaeal Immune System Can Detect Multiple Protospacer Adjacent Motifs (PAMs) to Target Invader DNA. J Biol Chem. 2012;287:33351–33363. doi: 10.1074/jbc.M112.377002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Gudbergsdottir S, et al. Dynamic properties of the Sulfolobus CRISPR/Cas and CRISPR/Cmr systems when challenged with vector-borne viral and plasmid genes and protospacers. Molecular microbiology. 2011;79:35–49. doi: 10.1111/j.1365-2958.2010.07452.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Manica A, et al. In vivo activity of CRISPR-mediated virus defence in a hyperthermophilic archaeon. Molecular microbiology. 2011;80:481–491. doi: 10.1111/j.1365-2958.2011.07586.x. [DOI] [PubMed] [Google Scholar]
- 22.Zhang J, et al. Structure and mechanism of the CMR complex for CRISPR-mediated antiviral immunity. Molecular cell. 2012;45:303–313. doi: 10.1016/j.molcel.2011.12.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Hale CR, et al. RNA-guided RNA cleavage by a CRISPR RNA-Cas protein complex. Cell. 2009;139:945–956. doi: 10.1016/j.cell.2009.07.040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hale CR, et al. Essential features and rational design of CRISPR RNAs that function with the Cas RAMP module complex to cleave RNAs. Molecular cell. 2012;45:292–302. doi: 10.1016/j.molcel.2011.10.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Spilman M, et al. Structure of an RNA Silencing Complex of the CRISPR-Cas Immune System. Molecular cell. 2013;52:146–152. doi: 10.1016/j.molcel.2013.09.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Staals RH, et al. Structure and Activity of the RNA-Targeting Type III-B CRISPR-Cas Complex of Thermus thermophilus. Molecular cell. 2013;52:135–145. doi: 10.1016/j.molcel.2013.09.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Barrangou R, et al. CRISPR provides acquired resistance against viruses in prokaryotes. Science. 2007;315:1709–1712. doi: 10.1126/science.1138140. [DOI] [PubMed] [Google Scholar]
- 28.Datsenko KA, et al. Molecular memory of prior infections activates the CRISPR/Cas adaptive bacterial immunity system. Nature communications. 2012;3:945. doi: 10.1038/ncomms1937. [DOI] [PubMed] [Google Scholar]
- 29.Erdmann S, Garrett RA. Selective and hyperactive uptake of foreign DNA by adaptive immune systems of an archaeon via two distinct mechanisms. Molecular microbiology. 2012;85:1044–1056. doi: 10.1111/j.1365-2958.2012.08171.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Swarts DC, et al. CRISPR interference directs strand specific spacer acquisition. PloS one. 2012;7:e35888. doi: 10.1371/journal.pone.0035888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Yosef I, et al. Proteins and DNA elements essential for the CRISPR adaptation process in Escherichia coli. Nucleic acids research. 2012;40:5569–5576. doi: 10.1093/nar/gks216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Brouns SJ, et al. Small CRISPR RNAs guide antiviral defense in prokaryotes. Science. 2008;321:960–964. doi: 10.1126/science.1159689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Carte J, et al. Binding and cleavage of CRISPR RNA by Cas6. Rna. 2010;16:2181–2188. doi: 10.1261/rna.2230110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Carte J, et al. Cas6 is an endoribonuclease that generates guide RNAs for invader defense in prokaryotes. Genes & development. 2008;22:3489–3496. doi: 10.1101/gad.1742908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Deltcheva E, et al. CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III. Nature. 2011;471:602–607. doi: 10.1038/nature09886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Garside EL, et al. Cas5d processes pre-crRNA and is a member of a larger family of CRISPR RNA endonucleases. Rna. 2012;18:2020–2028. doi: 10.1261/rna.033100.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Hatoum-Aslan A, et al. Mature clustered, regularly interspaced, short palindromic repeats RNA (crRNA) length is measured by a ruler mechanism anchored at the precursor processing site. Proceedings of the National Academy of Sciences of the United States of America. 2011;108:21218–21222. doi: 10.1073/pnas.1112832108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Haurwitz RE, et al. Sequence- and structure-specific RNA processing by a CRISPR endonuclease. Science. 2010;329:1355–1358. doi: 10.1126/science.1192272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Hale C, et al. Prokaryotic silencing (psi)RNAs in Pyrococcus furiosus. Rna. 2008;14:2572–2579. doi: 10.1261/rna.1246808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Lintner NG, et al. Structural and functional characterization of an archaeal clustered regularly interspaced short palindromic repeat (CRISPR)-associated complex for antiviral defense (CASCADE) J Biol Chem. 2011;286:21643–21656. doi: 10.1074/jbc.M111.238485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Wiedenheft B, et al. Structures of the RNA-guided surveillance complex from a bacterial immune system. Nature. 2011;477:486–489. doi: 10.1038/nature10402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Wiedenheft B, et al. RNA-guided complex from a bacterial immune system enhances target recognition through seed sequence interactions. Proceedings of the National Academy of Sciences of the United States of America. 2011;108:10092–10097. doi: 10.1073/pnas.1102716108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Jore MM, et al. Structural basis for CRISPR RNA-guided DNA recognition by Cascade. Nature structural & molecular biology. 2011;18:529–536. doi: 10.1038/nsmb.2019. [DOI] [PubMed] [Google Scholar]
- 44.Beloglazova N, et al. Structure and activity of the Cas3 HD nuclease MJ0384, an effector enzyme of the CRISPR interference. The EMBO journal. 2011;30:4616–4627. doi: 10.1038/emboj.2011.377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Sinkunas T, et al. Cas3 is a single-stranded DNA nuclease and ATP-dependent helicase in the CRISPR/Cas immune system. The EMBO journal. 2011;30:1335–1342. doi: 10.1038/emboj.2011.41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Sapranauskas R, et al. The Streptococcus thermophilus CRISPR/Cas system provides immunity in Escherichia coli. Nucleic acids research. 2011;39:9275–9282. doi: 10.1093/nar/gkr606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Horvath P, et al. Applications of the Versatile CRISPR-Cas Systems. In: heidelberg S-VB, editor. CRISPR-Cas Systems RNA-mediated Adative Immunity in Bacteria and Archaea. 2012. pp. 267–278. [Google Scholar]
- 48.Pennisi E. The CRISPR craze. Science. 2013;341:833–836. doi: 10.1126/science.341.6148.833. [DOI] [PubMed] [Google Scholar]
- 49.Richter H, et al. Exploiting CRISPR/Cas: interference mechanisms and applications. International journal of molecular sciences. 2013;14:14518–14531. doi: 10.3390/ijms140714518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Andersson AF, Banfield JF. Virus population dynamics and acquired virus resistance in natural microbial communities. Science. 2008;320:1047–1050. doi: 10.1126/science.1157358. [DOI] [PubMed] [Google Scholar]
- 51.Barrangou R, Horvath P. CRISPR: new horizons in phage resistance and strain identification. Annual review of food science and technology. 2012;3:143–162. doi: 10.1146/annurev-food-022811-101134. [DOI] [PubMed] [Google Scholar]
- 52.Shariat N, et al. The combination of CRISPR-MVLST and PFGE provides increased discriminatory power for differentiating human clinical isolates of Salmonella enterica subsp. enterica serovar Enteritidis. Food microbiology. 2013;34:164–173. doi: 10.1016/j.fm.2012.11.012. [DOI] [PubMed] [Google Scholar]
- 53.Delannoy S, et al. Use of clustered regularly interspaced short palindromic repeat sequence polymorphisms for specific detection of enterohemorrhagic Escherichia coli strains of serotypes O26:H11, O45:H2, O103:H2, O111:H8, O121:H19, O145:H28, and O157:H7 by real-time PCR. Journal of clinical microbiology. 2012;50:4035–4040. doi: 10.1128/JCM.02097-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Stern A, et al. CRISPR targeting reveals a reservoir of common phages associated with the human gut microbiome. Genome research. 2012;22:1985–1994. doi: 10.1101/gr.138297.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Shariat N, Dudley EG. CRISPRs: Molecular Signatures Used for Pathogen Subtyping. Applied and environmental microbiology. 2014;80:430–439. doi: 10.1128/AEM.02790-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Elmore JR, et al. Programmable plasmid interference by the CRISPR-Cas system in Thermococcus kodakarensis. RNA Biol. 2013;10:828–840. doi: 10.4161/rna.24084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Pougach K, et al. Transcription, processing and function of CRISPR cassettes in Escherichia coli. Molecular microbiology. 2010;77:1367–1379. doi: 10.1111/j.1365-2958.2010.07265.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Urnov FD, et al. Genome editing with engineered zinc finger nucleases. Nature reviews Genetics. 2010;11:636–646. doi: 10.1038/nrg2842. [DOI] [PubMed] [Google Scholar]
- 59.Perez-Pinera P, et al. Advances in targeted genome editing. Current opinion in chemical biology. 2012;16:268–277. doi: 10.1016/j.cbpa.2012.06.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Joung JK, Sander JD. TALENs: a widely applicable technology for targeted genome editing. Nature reviews Molecular cell biology. 2013;14:49–55. doi: 10.1038/nrm3486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Gasiunas G, Siksnys V. RNA-dependent DNA endonuclease Cas9 of the CRISPR system: Holy Grail of genome editing? Trends Microbiol. 2013;21:562–567. doi: 10.1016/j.tim.2013.09.001. [DOI] [PubMed] [Google Scholar]
- 62.Mali P, et al. Cas9 as a versatile tool for engineering biology. Nature methods. 2013;10:957–963. doi: 10.1038/nmeth.2649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Jinek M, et al. RNA-programmed genome editing in human cells. eLife. 2013;2:e00471. doi: 10.7554/eLife.00471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Cho SW, et al. Targeted genome engineering in human cells with the Cas9 RNA-guided endonuclease. Nature biotechnology. 2013;31:230–232. doi: 10.1038/nbt.2507. [DOI] [PubMed] [Google Scholar]
- 65.Cong L, et al. Multiplex genome engineering using CRISPR/Cas systems. Science. 2013;339:819–823. doi: 10.1126/science.1231143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Mali P, et al. RNA-guided human genome engineering via Cas9. Science. 2013;339:823–826. doi: 10.1126/science.1232033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Hou Z, et al. Efficient genome engineering in human pluripotent stem cells using Cas9 from Neisseria meningitidis. Proceedings of the National Academy of Sciences of the United States of America. 2013;110:15644–15649. doi: 10.1073/pnas.1313587110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Ebina H, et al. Harnessing the CRISPR/Cas9 system to disrupt latent HIV-1 provirus. Scientific reports. 2013;3:2510. doi: 10.1038/srep02510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Ran FA, et al. Double nicking by RNA-guided CRISPR Cas9 for enhanced genome editing specificity. Cell. 2013;154:1380–1389. doi: 10.1016/j.cell.2013.08.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Fu Y, et al. High-frequency off-target mutagenesis induced by CRISPR-Cas nucleases in human cells. Nature biotechnology. 2013;31:822–826. doi: 10.1038/nbt.2623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Hsu PD, et al. DNA targeting specificity of RNA-guided Cas9 nucleases. Nature biotechnology. 2013;31:827–832. doi: 10.1038/nbt.2647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Cradick TJ, et al. CRISPR/Cas9 systems targeting beta-globin and CCR5 genes have substantial off-target activity. Nucleic acids research. 2013;41:9584–9592. doi: 10.1093/nar/gkt714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Schwank G, et al. Functional Repair of CFTR by CRISPR/Cas9 in Intestinal Stem Cell Organoids of Cystic Fibrosis Patients. Cell stem cell. 2013;13:653–658. doi: 10.1016/j.stem.2013.11.002. [DOI] [PubMed] [Google Scholar]
- 74.Shalem O, et al. Genome-scale CRISPR-Cas9 knockout screening in human cells. Science. 2014;343:84–87. doi: 10.1126/science.1247005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Wang T, et al. Genetic screens in human cells using the CRISPR-Cas9 system. Science. 2014;343:80–84. doi: 10.1126/science.1246981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Li W, et al. Simultaneous generation and germline transmission of multiple gene mutations in rat using CRISPR-Cas systems. Nature biotechnology. 2013;31:684–686. doi: 10.1038/nbt.2652. [DOI] [PubMed] [Google Scholar]
- 77.Li D, et al. Heritable gene targeting in the mouse and rat using a CRISPR-Cas system. Nature biotechnology. 2013;31:681–683. doi: 10.1038/nbt.2661. [DOI] [PubMed] [Google Scholar]
- 78.Yang H, et al. One-step generation of mice carrying reporter and conditional alleles by CRISPR/Cas-mediated genome engineering. Cell. 2013;154:1370–1379. doi: 10.1016/j.cell.2013.08.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Wang H, et al. One-Step Generation of Mice Carrying Mutations in Multiple Genes by CRISPR/Cas-Mediated Genome Engineering. Cell. 2013;153:910–918. doi: 10.1016/j.cell.2013.04.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Wu Y, et al. Correction of a Genetic Disease in Mouse via Use of CRISPR-Cas9. Cell stem cell. 2013;13:659–662. doi: 10.1016/j.stem.2013.10.016. [DOI] [PubMed] [Google Scholar]
- 81.Hwang WY, et al. Efficient genome editing in zebrafish using a CRISPR-Cas system. Nature biotechnology. 2013;31:227–229. doi: 10.1038/nbt.2501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Chang N, et al. Genome editing with RNA-guided Cas9 nuclease in zebrafish embryos. Cell research. 2013;23:465–472. doi: 10.1038/cr.2013.45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Jao LE, et al. Efficient multiplex biallelic zebrafish genome editing using a CRISPR nuclease system. Proceedings of the National Academy of Sciences of the United States of America. 2013;110:13904–13909. doi: 10.1073/pnas.1308335110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Xiao A, et al. Chromosomal deletions and inversions mediated by TALENs and CRISPR/Cas in zebrafish. Nucleic acids research. 2013;41:e141. doi: 10.1093/nar/gkt464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Nakayama T, et al. Simple and efficient CRISPR/Cas9-mediated targeted mutagenesis in Xenopus tropicalis. Genesis. 2013;51:835–843. doi: 10.1002/dvg.22720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Blitz IL, et al. Biallelic genome modification in F0 Xenopus tropicalis embryos using the CRISPR/Cas system. Genesis. 2013;51:827–834. doi: 10.1002/dvg.22719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Shan Q, et al. Targeted genome modification of crop plants using a CRISPR-Cas system. Nature biotechnology. 2013;31:686–688. doi: 10.1038/nbt.2650. [DOI] [PubMed] [Google Scholar]
- 88.Li JF, et al. Multiplex and homologous recombination-mediated genome editing in Arabidopsis and Nicotiana benthamiana using guide RNA and Cas9. Nature biotechnology. 2013;31:688–691. doi: 10.1038/nbt.2654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Feng Z, et al. Efficient genome editing in plants using a CRISPR/Cas system. Cell research. 2013;23:1229–1232. doi: 10.1038/cr.2013.114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Miao J, et al. Targeted mutagenesis in rice using CRISPR-Cas system. Cell research. 2013;23:1233–1236. doi: 10.1038/cr.2013.123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Jiang W, et al. Demonstration of CRISPR/Cas9/sgRNA-mediated targeted gene modification in Arabidopsis, tobacco, sorghum and rice. Nucleic acids research. 2013;41:e188. doi: 10.1093/nar/gkt780. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Nekrasov V, et al. Targeted mutagenesis in the model plant Nicotiana benthamiana using Cas9 RNA-guided endonuclease. Nature biotechnology. 2013;31:691–693. doi: 10.1038/nbt.2655. [DOI] [PubMed] [Google Scholar]
- 93.Upadhyay SK, et al. RNA-Guided Genome Editing for Target Gene Mutations in Wheat. 2013;G33:2233–2238. doi: 10.1534/g3.113.008847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Bassett AR, et al. Highly Efficient Targeted Mutagenesis of Drosophila with the CRISPR/Cas9 System. Cell reports. 2013;4:220–228. doi: 10.1016/j.celrep.2013.06.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Gratz SJ, et al. CRISPR/Cas9-mediated genome engineering and the promise of designer flies on demand. Fly. 2013;7:249–255. doi: 10.4161/fly.26566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Gratz SJ, et al. Genome Engineering of Drosophila with the CRISPR RNA-Guided Cas9 Nuclease. Genetics. 2013;194:1029–1035. doi: 10.1534/genetics.113.152710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Yu Z, et al. Highly efficient genome modifications mediated by CRISPR/Cas9 in Drosophila. Genetics. 2013;195:289–291. doi: 10.1534/genetics.113.153825. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Kondo S, Ueda R. Highly improved gene targeting by germline-specific Cas9 expression in Drosophila. Genetics. 2013;195:715–721. doi: 10.1534/genetics.113.156737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Sebo ZL, et al. A simplified and efficient germline-specific CRISPR/Cas9 system for Drosophila genomic engineering. Fly. 2013;8:1–6. doi: 10.4161/fly.26828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Friedland AE, et al. Heritable genome editing in C. elegans via a CRISPR-Cas9 system. Nature methods. 2013;10:741–743. doi: 10.1038/nmeth.2532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Lo TW, et al. Precise and heritable genome editing in evolutionarily diverse nematodes using TALENs and CRISPR/Cas9 to engineer insertions and deletions. Genetics. 2013;195:331–348. doi: 10.1534/genetics.113.155382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Waaijers S, et al. CRISPR/Cas9-Targeted Mutagenesis in Caenorhabditis elegans. Genetics. 2013;195:1187–1191. doi: 10.1534/genetics.113.156299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Tzur YB, et al. Heritable custom genomic modifications in Caenorhabditis elegans via a CRISPR-Cas9 system. Genetics. 2013;195:1181–1185. doi: 10.1534/genetics.113.156075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Katic I, Grosshans H. Targeted heritable mutation and gene conversion by Cas9-CRISPR in Caenorhabditis elegans. Genetics. 2013;195:1173–1176. doi: 10.1534/genetics.113.155754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Chiu H, et al. Transgene-free genome editing in Caenorhabditis elegans using CRISPR-Cas. Genetics. 2013;195:1167–1171. doi: 10.1534/genetics.113.155879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Dickinson DJ, et al. Engineering the Caenorhabditis elegans genome using Cas9-triggered homologous recombination. Nature methods. 2013;10:1028–1034. doi: 10.1038/nmeth.2641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Chen C, et al. Efficient genome editing in Caenorhabditis elegans by CRISPR-targeted homologous recombination. Nucleic acids research. 2013;41:e193. doi: 10.1093/nar/gkt805. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Wang Y, et al. The CRISPR/Cas System mediates efficient genome engineering in Bombyx mori. Cell research. 2013;23:1414–1416. doi: 10.1038/cr.2013.146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.DiCarlo JE, et al. Genome engineering in Saccharomyces cerevisiae using CRISPR-Cas systems. Nucleic acids research. 2013;41:4336–4343. doi: 10.1093/nar/gkt135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Jiang W, et al. RNA-guided editing of bacterial genomes using CRISPR-Cas systems. Nature biotechnology. 2013;31:233–239. doi: 10.1038/nbt.2508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Mali P, et al. CAS9 transcriptional activators for target specificity screening and paired nickases for cooperative genome engineering. Nature biotechnology. 2013;31:833–838. doi: 10.1038/nbt.2675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Deveau H, et al. Phage response to CRISPR-encoded resistance in Streptococcus thermophilus. Journal of bacteriology. 2008;190:1390–1400. doi: 10.1128/JB.01412-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Esvelt KM, et al. Orthogonal Cas9 proteins for RNA-guided gene regulation and editing. Nature methods. 2013;10:1116–1121. doi: 10.1038/nmeth.2681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Qi LS, et al. Repurposing CRISPR as an RNA-Guided Platform for Sequence-Specific Control of Gene Expression. Cell. 2013;152:1173–1183. doi: 10.1016/j.cell.2013.02.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Cheng AW, et al. Multiplexed activation of endogenous genes by CRISPR-on, an RNA-guided transcriptional activator system. Cell research. 2013;23:1163–1171. doi: 10.1038/cr.2013.122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Farzadfard F, et al. Tunable and Multifunctional Eukaryotic Transcription Factors Based on CRISPR/Cas. ACS synthetic biology. 2013;13:604–613. doi: 10.1021/sb400081r. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Gilbert LA, et al. CRISPR-Mediated Modular RNA-Guided Regulation of Transcription in Eukaryotes. Cell. 2013;154:442–451. doi: 10.1016/j.cell.2013.06.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Bikard D, et al. Programmable repression and activation of bacterial gene expression using an engineered CRISPR-Cas system. Nucleic acids research. 2013;41:7429–7437. doi: 10.1093/nar/gkt520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Pattanayak V, et al. High-throughput profiling of off-target DNA cleavage reveals RNA-programmed Cas9 nuclease specificity. Nature biotechnology. 2013;31:839–843. doi: 10.1038/nbt.2673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Qi L, et al. RNA processing enables predictable programming of gene expression. Nature biotechnology. 2012;30:1002–1006. doi: 10.1038/nbt.2355. [DOI] [PubMed] [Google Scholar]
- 121.Lee HY, et al. RNA-protein analysis using a conditional CRISPR nuclease. Proceedings of the National Academy of Sciences of the United States of America. 2013;110:5416–5421. doi: 10.1073/pnas.1302807110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Salvail-Lacoste A, et al. Affinity purification of T7 RNA transcripts with homogeneous ends using ARiBo and CRISPR tags. RNA. 2013;19:1003–1014. doi: 10.1261/rna.037432.112. [DOI] [PMC free article] [PubMed] [Google Scholar]