

Abstract
Source memory represents the origin (source) of information. Recently, we proposed that rats (Rattus norvegicus) remember the source of information. However, an alternative to source memory is the possibility that rats selectively encoded some, but not all, information rather than retrieving an episodic memory. We directly tested this ‘encoding failure’ hypothesis. Here, we show that rats remember the source of information, under conditions that cannot be attributed to encoding failure. Moreover, source memory lasted at least seven days but was no longer present 14 days after studying. Our findings suggest that long-lasting source memory may be modelled in non-humans. Our model should facilitate attempts to elucidate the biological underpinnings of source memory impairments in human memory disorders such as Alzheimer's disease.
Keywords: source memory, episodic memory, encoding failure hypothesis, animal model, retention interval, rat (Rattus norvegicus)
1. Introduction
Source memory refers to memories about the conditions under which information was acquired [1,2]. Episodic memory (i.e. our memories for unique personal past events) typically involves source memory because those memories focus on the origin of mental experiences [3,4]. Source memory is at work when, for example, we tell a favourite joke to friends while avoiding retelling the joke to the person who originally told it to us [5]. Importantly, source memory allows us to differentiate one episodic memory from another because source memory includes features that were present when the memory was formed [1,2].
We recently proposed that rats remember the source of encoded information [6]. In our approach, rats foraged for food in a radial maze (food could be obtained at the distal ends of eight arms radiating from a central chamber). Rats foraged for distinctive flavours of food that replenished or failed to replenish at its recently encountered location according to a source information rule. The source memory of eating chocolate pellets was manipulated by the experimenter placing the rat at the food trough of one arm which dispensed chocolate, whereas the rat encountered chocolate on its own at a food trough on a different arm (these two arms were randomly selected and the rats discovered chow-flavoured pellets at two other randomly selected arms). After a retention interval, the rats discovered chow-flavoured pellets at the four not-yet-visited arms. The arm where the rat discovered chocolate on its own provided additional chocolate at the test (replenishment), whereas the arm where the rat was placed by the experimenter did not provide additional chocolate (non-replenishment) in some experiments; in other experiments, the replenishment contingency was reversed. Rats revisited the replenishment location at a higher rate than the non-replenishment location while avoiding revisits to chow locations (which never replenished).
We proposed that rats monitor and remember the source of encoded information. However, a non-episodic memory alternative explanation exists for our earlier experiments. It may be argued that the rats might have learned that encoding the chocolate location was not required at the non-replenishment arm, which we refer to as the encoding failure hypothesis [7], a form of selective encoding. Because non-replenishment could be predicted at the time the rat initially encountered chocolate, a rat might solve the task by encoding the chocolate location when initially discovering the replenishment location (e.g. when encountering the chocolate on its own), but not encoding the chocolate location when discovering the non-replenishment location (e.g. placement by the experimenter). Importantly, higher revisit rates at the replenishment location compared with the non-replenishment location would occur in this situation without the rat remembering the episode.
Here, we tested the encoding failure hypothesis. Our approach (figure 1) was to provide rats with some information needed to predict replenishment at the time of encoding (location, food flavour, source), but one critical piece of information needed to predict replenishment was not presented until immediately before the memory assessment. We provided a retrieval cue (the presence/absence of additional chocolate pellets in the central chamber, counterbalanced across rats) that was needed to predict replenishment when combined with source memory; chow-flavoured food was available at all other locations but never replenished. Thus, to solve the task, the rats needed to encode location, flavour and source information at both chocolate locations, but decoding of the replenishment location was delayed until immediately before memory assessment. If the encoding failure hypothesis explained the results from our previous experiments, then it would be impossible for the rats to solve the current task. By contrast, if rats used source memory, they should revisit chocolate at a higher rate at replenishment than non-replenishment locations. Next, we examined source memory using up to 14 day retention intervals. Because source memory, and episodic memory more generally, is a form of long-term memory, we expect that source memory will survive relatively long retention-interval challenges.
Figure 1.
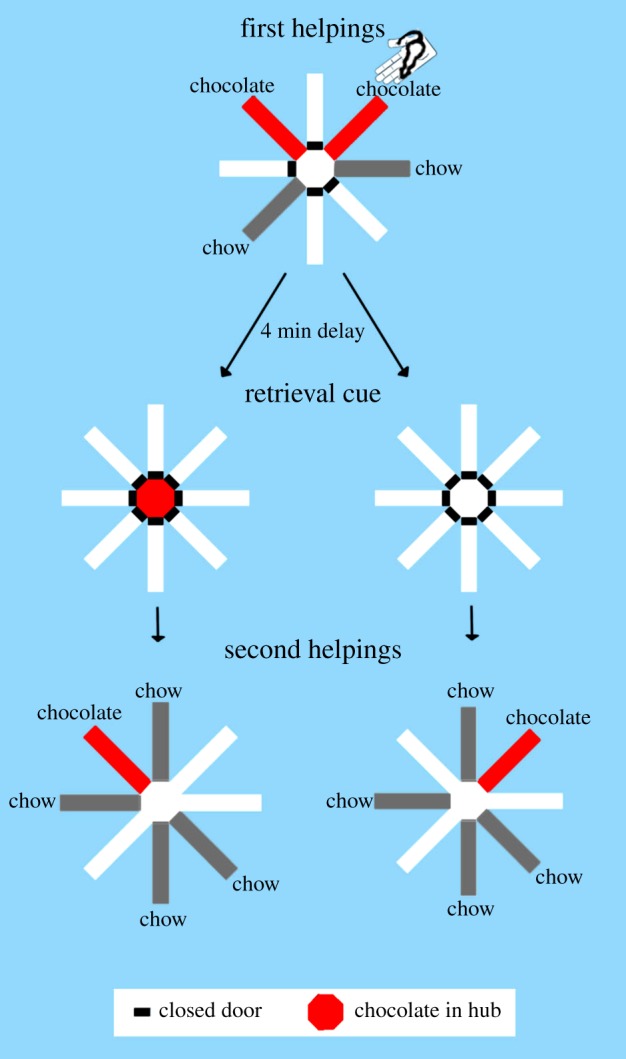
Schematic of experimental design showing topographical views of the maze. For half of the rats, hub baiting with 15 chocolate pellets signalled that only the self-located chocolate arm would be replenished, whereas hub baiting with zero chocolate pellets signalled that only the arm where the experimenter had placed the rat would be replenished; these contingencies were reversed (not shown) for the remaining rats.
2. Material and methods
(a). Subjects
Sixteen male Long-Evans rats (Rattus norvegicus; Harlan; 61 days and 325 g) were individually housed with light onset and offset at 07.00 and 19.00, respectively; one rat was inadvertently euthanized by a different experimenter, and so the dataset had 15 rats. They received 45 mg chow and chocolate pellets (F0165 and F0299; Bio-Serv) during sessions followed by 15 g d−1 of 5012-Rat-Diet (PMI Nutrition International). Water was ad libitum, except in the maze.
(b). Apparatus
A radial maze (central chamber and eight arms, each equipped with a guillotine door, food trough, 45 mg pellet dispenser and photobeams) was used (see [6,8]).
(c). Experimental procedure
Two experiments were conducted: encoding failure and retention-interval experiments.
(i). Encoding failure
Pretraining and initial training are described in the Detailed Methodology section of the electronic supplementary material.
Daily sessions consisted of one trial, but they were divided across study and test phases as follows: a study phase (first helpings of food), a retention interval, a retrieval cue (pellets or no-pellets in the hub) and a test phase (second helpings of food). In the study phase (first helpings), doors to four out of eight arms (randomly chosen for each rat each day) were opened and closed individually by the experimenter so the rat would enter a predetermined (randomly selected) sequence of arms. Controlled access also allowed the experimenter to place the rat in one of the designated chocolate arms (randomly selected); the rat was placed at the distal end of the arm near the food trough and facing the trough and dispenser. Visits to two of these study arms were reinforced with three chocolate pellets and visits to the other two study arms were reinforced with one chow pellet. Pellet(s) were delivered to accessible troughs with the interruption of the trough photobeam. The study phase ended when food had been dispensed at each accessible location and then the rat was removed. After a delay of approximately 4 min, the animal was replaced in the hub for presentation of retrieval cues; on a daily basis, half the rats (randomly selected) received 15 chocolate pellets in the central hub and the other half zero pellets. Hub baiting with 15 or zero chocolate pellets was followed by a test phase (second helpings) with doors to all eight arms opened. In the test phase, one chow pellet was available in each arm that was not accessible during the study phase (first helpings); in addition, one of the two study-phase chocolate locations provided three chocolate pellets per visit (second helpings) during up to five visits per chocolate location. The particular hub-baiting retrieval cue signalled which study-phase chocolate location would be replenished as diagramed in figure 1. For half of the rats, hub baiting with 15 chocolate pellets signalled that only the self-located chocolate arm would be replenished, whereas hub baiting with zero chocolate pellets signalled that only the arm where the experimenter had placed the rat would be replenished; these contingencies were reversed for the remaining rats. Initially, the type of hub baiting was presented in alternating blocks of 4–34 sessions, for a total of 134 sessions. Finally, in mixed testing, the type of hub baiting was randomly selected each session for a total of 24 sessions with restriction that there be three of each type in each six-session block and no more than three consecutive sessions of the same type. All mixed-testing sessions were analysed.
Detailed methodology is provided in the electronic supplementary material.
(ii). Retention interval
The procedure (23 sessions) was identical to mixed testing described above, except the retention interval was approximately 4 min, 3 h, 1, 4, 7 or 14 days. Short (4 min) and longer retention-intervals were mixed across days. We initially tested 4 min to 7 day retention intervals, which was the range we used previously [6]. Because we did not observe any evidence for source memory decay in this range, we added the 14 day retention-interval condition to verify that source memory eventually decays. All sessions were analysed.
3. Results
(a). Encoding failure experiment
When replenishment was decoded by a retrieval cue immediately before memory assessment, rats preferentially revisited the chocolate location when it was about to replenish (figure 2a; F1,14 = 21.8, p < 0.001). The type of retrieval cue (food versus no-food in hub) did not influence revisit rates (F1,14 = 2.0, p > 0.05) nor did it interact with the replenishment variable (F1,4 = 1.5, p > 0.05). Differential rates of revisiting chocolate locations were accomplished while rats accurately avoided revisits to depleted chow locations (table 1).
Figure 2.
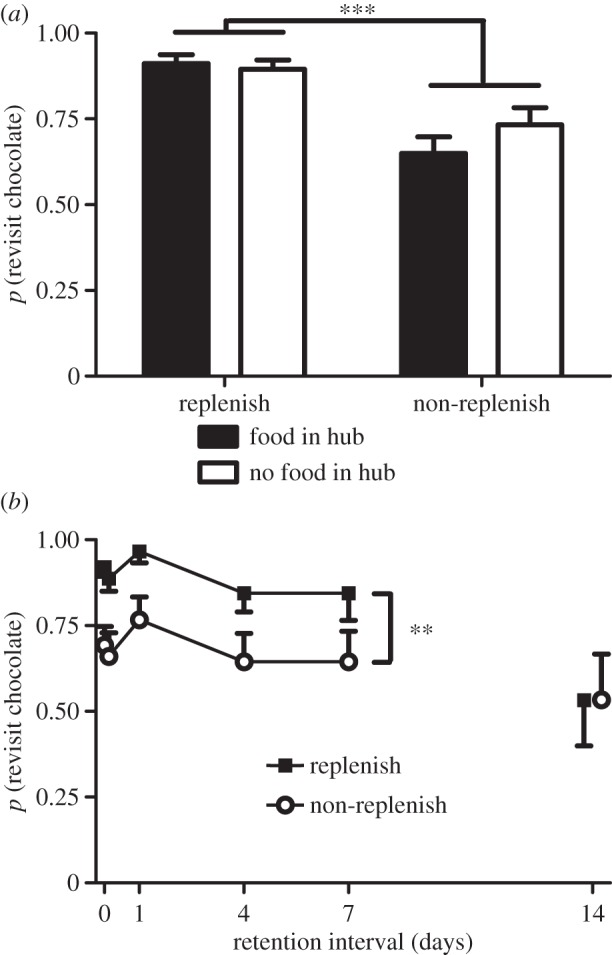
(a,b) Rats preferentially revisit the chocolate location when it is about to replenish in (a) encoding failure and (b) retention-interval experiments. The data are mean with 1 s.e.m.; the probability of at least one revisit to the chocolate location was calculated from the first five choices in memory-assessment phases (1 if at least one revisit occurred; 0 otherwise). Replenishment is different from non-replenishment: **p < 0.01, ***p < 0.001. The probability expected by chance is 0.487 (calculated with a geometric distribution). (a) All values are above chance (p < 0.001). (b) Mean across retention intervals less than 14 days are above chance in replenishment (p < 0.001) and non-replenishment (p < 0.05); performance is not different from chance at 14 day delay.
Table 1.
Accuracy in avoiding revisits to depleted chow-flavoured locations. Accuracy was measured as the proportion correct in the first four choices excluding the chocolate locations in a test phase. This analysis of chow accuracy was restricted to the six non-chocolate arms. The Bayesian expected value by chance is 0.518. The shortest delay by which chow accuracy dropped below the 4-min baseline occurred at 4 days (t14 = 4.3, p < 0.001).
| procedure | retention interval | mean ± s.e.m. |
|---|---|---|
| encoding failure experiment | 4 min | 0.728*** ± 0.029 |
| retention-interval experiment | 4 min | 0.709*** ± 0.027 |
| 3 h | 0.660** ± 0.047 | |
| 1 day | 0.681* ± 0.059 | |
| 4 days | 0.576 ± 0.040 | |
| 7 days | 0.628 ± 0.060 | |
| 14 days | 0.550 ± 0.061 |
*p < 0.05, **p < 0.01, ***p < 0.001.
(b). Retention-interval experiment
We initially examined retention intervals between 4 min and 7 days, which corresponds to the range previously investigated [6]. Revisits to the replenishment-chocolate location was higher than to the non-replenishment-chocolate location (figure 2b; F1,14 = 9.4, p < 0.017). Revisits did not vary across retention intervals up to 7 days (F4,56 = 1.7, p > 0.05) and retention interval did not interact with the replenishment variable (F4,56 < 1, p > 0.05). In contrast to the approximately consistent source memory performance across retention intervals up to 7 days, chow accuracy declined as a function of these retention intervals (F4,56 = 3.2, p < 0.05; table 1). Previously, we observed the introduction of source memory errors with a 7 day retention interval, yet in the present experiment, there was no evidence for source memory decay even after a 7 day delay. Thus, we added a 14 day retention-interval condition, which documented the eventual elimination of source memory (no difference between replenishment and non-replenishment revisits after a 14 day delay, t14 < 1).
4. Discussion
Our findings suggest that rats monitor and remember the source of encoded information in conditions which preclude encoding failure. Moreover, source memory is remarkably long lasting. The rats remembered the chocolate-baited locations and the source by which the chocolate had been obtained (experimenter-generated versus self-generated features of the earlier episode) for at least 7 days without any apparent decline in performance; after a 14 day delay, source memory is apparently eliminated under the present conditions.
Source memory is dissociated from location memory by different forgetting rates because generic location memory decayed over the course of the long retention intervals. Preferential revisits to the replenishment-chocolate location cannot be explained by the relative familiarity of study-phase features (e.g. placement or non-placement feedings, chocolate or chow flavours) because the interval between study and test was constant. Consequently, our data suggest that the rats remembered a specific past episode rather than general information about remoteness [9–13]. Rats apparently remember the source by which information was acquired, in addition to other features of the episode (location, flavour and retrieval cue). Elsewhere we showed that source memory is hippocampal dependent [6] and that retrieval of episodic information can occur when encoding is incidental and memory assessment is unexpected [14,15].
Our findings suggest that source memory is evolutionarily quite old. Moreover, our findings support the view that rodents may be used to model fundamental aspects of human memory. This view opens the door to combining a deep understanding of biological mechanisms with animal models of human cognition to advance translational research that may ultimately foster the development of therapeutic approaches to severe human memory disorders [16].
Data accessibility
The data supporting this article are included in dryad: doi:10.5061/dryad.jq470.
Funding statement
Supported by National Institute of Mental Health R01MH080052 and R01MH098985 to J.D.C.
References
- 1.Johnson MK, Hashtroudi S, Lindsay DS. 1993. Source monitoring. Psychol. Bull. 114, 3–28 (doi:10.1037/0033-2909.114.1.3) [DOI] [PubMed] [Google Scholar]
- 2.Mitchell KJ, Johnson MK. 2009. Source monitoring 15 years later: what have we learned from fMRI about the neural mechanisms of source memory? Psychol. Bull. 135, 638–677 (doi:10.1037/a0015849) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Johnson MK. 2005. The relation between source memory and episodic memory: comment on Siedlecki et al. (2005). Psychol. Aging 20, 529–531 (doi:10.1037/0882-7974.20.3.529) [DOI] [PubMed] [Google Scholar]
- 4.McDuff SGR, Frankel HC, Norman KA. 2009. Multivoxel pattern analysis reveals increased memory targeting and reduced use of retrieved details during single-agenda source monitoring. J. Neurosci. 29, 508–516 (doi:10.1523/JNEUROSCI.3587-08.2009) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Wright AA. 2013. episodic memory: a rat model of source memory. Curr. Biol. 23, R198–R200 (doi:10.1016/j.cub.2013.01.055) [DOI] [PubMed] [Google Scholar]
- 6.Crystal JD, Alford WT, Zhou W, Hohmann AG. 2013. Source memory in the rat. Curr. Biol. 23, 387–391 (doi:10.1016/j.cub.2013.01.023) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Zhou W, Crystal JD. 2011. Validation of a rodent model of episodic memory. Anim. Cogn. 14, 325–340 (doi:10.1007/s10071-010-0367-0) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Babb SJ, Crystal JD. 2005. Discrimination of what, when, and where: implications for episodic-like memory in rats. Learn. Motiv. 36, 177–189 (doi:10.1016/j.lmot.2005.02.009) [Google Scholar]
- 9.Clayton NS, Dickinson A. 1998. Episodic-like memory during cache recovery by scrub jays. Nature 395, 272–274 (doi:10.1038/26216) [DOI] [PubMed] [Google Scholar]
- 10.Eacott MJ, Easton A, Zinkivskay A. 2005. Recollection in an episodic-like memory task in the rat. Learn. Mem. 12, 221–223 (doi:10.1101/lm.92505) [DOI] [PubMed] [Google Scholar]
- 11.Fortin NJ, Wright SP, Eichenbaum H. 2004. Recollection-like memory retrieval in rats is dependent on the hippocampus. Nature 431, 188–191 (doi:10.1038/nature02853) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Roberts WA, Feeney MC, MacPherson K, Petter M, McMillan N, Musolino E. 2008. Episodic-like memory in rats: is it based on when or how long ago? Science 320, 113–115 (doi:10.1126/science.1152709) [DOI] [PubMed] [Google Scholar]
- 13.Zhou W, Crystal JD. 2009. Evidence for remembering when events occurred in a rodent model of episodic memory. Proc. Natl Acad. Sci. USA 106, 9525–9529 (doi:10.1073/pnas.0904360106) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Zhou W, Hohmann AG, Crystal JD. 2012. Rats answer an unexpected question after incidental encoding. Curr. Biol. 22, 1149–1153 (doi:10.1016/j.cub.2012.04.040) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Crystal JD. 2013. Remembering the past and planning for the future in rats. Behav. Process 93, 39–49 (doi:10.1016/j.beproc.2012.11.014) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Crystal JD, Glanzman DL. 2013. A biological perspective on memory. Curr. Biol. 23, R728–R731 (doi:10.1016/j.cub.2013.07.082) [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The data supporting this article are included in dryad: doi:10.5061/dryad.jq470.