

Abstract
Detergents have several biological applications but present cytotoxicity concerns, since they can solubilize cell membranes. Using the IonFlux 16, an ensemble whole cell planar patch clamp, we observed that anionic sodium dodecyl sulfate (SDS), cationic cetyltrimethylammonium bromide (CTAB), and cationic, fluorescent octadecyl rhodamine B (ORB) increased the membrane permeability of cells substantially within a second of exposure, under superfusion conditions. Increased permeability was irreversible for 15 min. At subsolubilizing detergent concentrations, patched cells showed increased membrane currents that reached a steady state and were intact when imaged using fluorescence microscopy. SDS solubilized cells at concentrations of 2 mM (2× CMC), while CTAB did not solubilize cells even at concentrations of 10 mM (1000× CMC). The relative activity for plasma membrane current induction was 1:20:14 for SDS, CTAB, and ORB, respectively. Under quiescent conditions, the relative ratio of lipid to detergent in cell membranes at the onset of membrane permeability was 1:7:5 for SDS, CTAB, and ORB, respectively. The partition constants (K) for SDS, CTAB, and ORB were 23000, 55000, and 39000 M–1, respectively. Combining the whole cell patch clamp data and XTT viability data, SDS ≤ 0.2 mM and CTAB and ORB ≤ 1 mM induced cell membrane permeability without causing acute toxicity.
Introduction
Detergents are employed for a wide variety of applications in biology including dissolving cell membranes,1−4 isolating membrane components such as proteins,5 transport of agents such as drugs and genes into the cell,6 adjuvants in vaccination,7 and providing aqueous solubility to hydrophobic and/or nanoscale materials.8 In part, this wide variety of roles stems from the ability to vary both the headgroup size and charge of a detergent, as well as the length and shape of the hydrocarbon tail. However, a common detergent such as sodium dodecyl sulfate (SDS) is able to play many of the roles described above by varying the concentration. This great variety of applications indicates the detergent/membrane interaction is multifaceted and has substantial continued promise for biological manipulation. In order to fulfill this promise, increased understanding of the balance between the roles in enhanced transport, membrane permeability, and overall dissolution of the cell membrane is needed.
The theory of detergent interaction with cell plasma membranes has been developed using homogeneous phospholipid bilayer models. The interaction is generally proposed to go through a three-stage equilibrium model (see the Supporting Information, Figure S1) as the detergent/lipid (D/L) ratio increases:1−4
-
(I)
intercalation of non-micellar detergent into the bilayer
-
(II)
equilibrium coexistence of phospholipid-saturated D/L micelles with detergent-saturated phospholipid bilayer
-
(III)
dissolution of detergent-saturated phospholipid bilayer into D/L micelles.
These three stages, although a simplification when considering detergent interaction with a cell plasma membrane containing numerous other components including proteins and cholesterol,5,9 nevertheless provide a starting point for considering the detergent/plasma membrane interaction and the experiments used to probe the biophysical outcomes. There are also three steps considered as important for the kinetics of the detergent/lipid bilayer interaction:4
-
(i)
insertion into the outer leaflet of the bilayer, ranging from milliseconds to seconds
-
(ii)
equilibration between the outer and inner leaflets, ranging from milliseconds to days
-
(iii)
equilibration between the inner leaflet and the inner aqueous compartment occurring over minutes to days
When considering the interaction of detergents with most cell types, both the thermodynamic and kinetic aspects described above fail to take account of active processes, particularly lipid cycling between the outer cell plasma membrane and the inner cellular membranes, which can serve to replenish lipid and other components, as well as transport detergent to the cell interior. For most eukaryotic cell types, the plasma membrane represents less than 5% total membrane composition of the cells.10 In this regard, the structure and dynamics of most eukaryotic cell plasma membranes differ substantially from that of lipid bilayer vesicle models or that of red blood cells (erythrocytes), in which the plasma membrane is the only membrane present. Another significant difference between physiological exposure of cells to detergent as compared to both model bilayer systems and most in vitro cell models is the choice of quiescent versus flowing systems. Most cell-based experiments to date have been performed as quiescent experiments using erythrocytes,11−14 although HeLa15 and B1616 cells have also been employed. The erythrocyte studies generally use hemolysis as a primary assay for membrane permeability, although the ability of this assay to effectively test stage I intercalation events has been questioned.13 Studies testing whether detergent-induced transbilayer lipid motion (flip-flop) was an early stage I event that could be directly related to cell membrane leakage determined that flip-flop and permeability were independent events.
In studies with a variety of positively charged nanomaterials including synthetic mimics of antimicrobial peptides, antimicrobial peptides, proteins, polymers, and particles on eukaryotic cells such as KB, Rat2, HeLa, and HEK293A, we and others noted that cell plasma membranes showed evidence of membrane leakage (lactate dehydrogenase (LDH), propidium iodide (PI), and fluorescein assays; increased membrane current) caused by disruption of the membrane and/or membrane pore formation far below concentrations that induced lysis.17−22 In model membrane systems, we and others have noted the direct introduction of nanoscale holes or pores, membrane thinning, and membrane intercalation.17,18,23−30 Increased membrane current was ascribed to structural membrane disruption or pore formation because the current induction was not cation specific and lacked rectification, as would be observed for ion-channel-based changes in current.19 With these findings in hand, and considering the extensive studies discussed above using detergents that also have a rich nanoscale structure, we were interested in exploring the stage I to II interactions of detergent with eukaryotic cell membranes. In particular, we wanted to take advantage of the sensitivity of electrical measurements using a whole cell patch clamp as a complementary approach to the hemolysis studies most common in the literature.
In this paper, we examine the interaction of SDS, CTAB, and ORB with HEK 293A cells using an automated planar patch clamp (IonFlux 16). The following major conclusions were reached: (1) detergent partitions from solution to cell plasma membrane much faster (seconds) than detergent exchanges between the plasma membrane and internal cell membranes (minutes), (2) detergent-induced cell membrane permeability does not decrease after removal of detergent from the external solution over a time period of >15 min, even with active equilibration with internal membranes, (3) XTT assays indicated ranges of detergent-induced cell plasma membrane permeability that were not acutely toxic, (4) the relative activity of SDS, CTAB, and ORB for the induction of membrane permeability HEK 293A cells was quantified for both superfusion and quiescent conditions, (5) whole cell patch clamp measurement of current induction was employed to obtain partition coefficients for SDS, CTAB, and ORB with the HEK 293A cells.
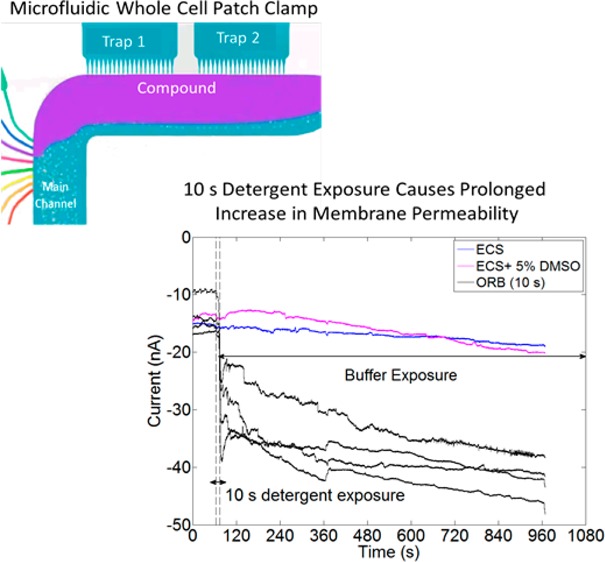
The IonFlux 16 uses the whole cell patch clamp configuration to measure the changes in membrane conductance for 16 groups of 20 cells in 8 independent patterns (320 patched cells per experimental run) (Figure 1). This instrument has a number of advantages/differences as compared to a traditional whole cell patch clamp19 using a single cell/electrode combination including (1) simultaneous ability to run multiple repeats and/or multiple exposure concentrations, (2) subsecond ability to change concentration, (3) continuous superfusion environment, and (4) ready posthoc analysis following electrical characterization of all 320 cells using fluorescence microscopy. Electrical characterization of cell plasma membrane permeability was obtained for all three detergents under dynamic exposure most closely related to thermodynamic stages I and II and kinetic stages i–iii. For SDS exposure, progression to stage III occurred as concentration increased, as evidenced by the observation of open channel currents. For both CTAB and ORB, intercalation did not result in progression to stage III (membrane dissolution), consistent with the elegant physical chemical studies of Seelig et al.31 and Xia and Onyuksel32 on lipid bilayer models that the headgroup dramatically effects tendency toward micellization. Consistent with their results, we observe SDS inducing micellization and complete dissolution of the cells for a 2 mM exposure, whereas the cell plasma membranes remain intact up to 10 and 1.4 mM for CTAB and ORB, respectively. Considering the ability of each detergent to induce membrane permeability prior to micellization occurring, we find that the relative activity as a function of concentration under flow conditions is 1:20:14 for SDS, CTAB, and ORB, respectively.
Figure 1.

(A) The IonFlux 16 consists of a 96-well plate with eight patterns (colored in gray and black and numbered in orange circles). (B) Each pattern has two trap wells (trap 1, trap 2) that are filled with ICS. Cells are loaded in the “INLET” wells, and waste is collected in the “OUTLET” well. Compounds are loaded in wells C1–C8. (C) A zoomed in view of the trap zones. The cells are suspended in ECS flow in the “main channel”, which flows past the trap zones. When a compound is released, the laminar flow ensures that the compound and ECS streams do not mix. (D) Image showing cells trapped in a trap zone. (Parts A–C were developed by Fluxion Biosciences.)
In order to understand the nature of detergent induced membrane permeability further, we also subjected cells to detergent exposure for 10–300 s and monitored if the membrane currents returned to baseline levels. For SDS (at sub-solubilizing concentrations), CTAB, and ORB, we observed that the increase in membrane permeability caused within 10 s of detergent exposure was not reversible for over 15 min. Further work is necessary to determine if this increased membrane permeability can be chronic and its long-term impact on cellular function in vitro and in vivo.
In order to understand the impact of changes in lipid concentration on the detergent induced membrane permeability, we also performed experiments under quiescent, equilibrium conditions by mixing detergent with varying numbers of cells followed by patching the cells and measuring the resulting membrane currents. Partitioning assays have been widely used in the literature to measure the lipid bilayer composition and partition constant of detergents in the bilayer.1−4,11−13,33−37 The concentration of detergent required to induce a given magnitude of membrane permeability is linearly related to the lipid concentration. At a fixed membrane perturbation level, the amount of total detergent in the system is constant, as presented in eq 1 where Db denotes detergent in bilayer and Dw denotes detergent in water.1−4
| 1 |
Rb is the ratio of detergent to lipid (L) in the bilayer for a given level of membrane disruption, as given in eq 2.1−4 Substituting eq 2 in eq 1 results in eq 3.
| 2 |
| 3 |
The partitioning of detergent into the lipid membrane can be described by a model (eq 4) where the mole fraction of detergent in the bilayer (Xb) is related to the detergent in water (Dw) through a partition constant (K).1−4
| 4 |
The substitution of eqs 2 and 4 into eq 3 results in eq 5.1−4
| 5 |
This experiment thus allows us to readily compare our patch clamp derived data with extensive literature in the field. We have determined the partition constants of SDS, CTAB, and ORB in the membrane to be 23000, 55000, and 39000 M–1, respectively. Fluorescence experiments for ORB indicated that detergent intercalated into the outer leaflet was internalized into the inner cellular membranes over the approximately 20 min time period of these experiments. The combination of the whole cell patch clamp data and XTT viability data suggest there is a range, ≤0.2 mM for SDS and ≤1 mM for CTAB and ORB, where detergent exposure can cause long-term plasma membrane permeability without causing acute toxicity. The results from our experiments indicate that additional work is needed to understand the role of low doses of these classes of materials on inducing inflammatory responses.
Experimental Methods
SDS and CTAB were obtained from Sigma-Aldrich (Sigma-Aldrich; St. Louis, MO). ORB was obtained from Life technologies (Carlsbad, CA). Other reagents were obtained from Fisher Scientific unless mentioned otherwise.
Ensemble Whole-Cell Patch Clamp Using IonFlux 16
Solutions
Extracellular solution (ECS) consisted of 138 mM NaCl, 4 mM KCl, 1.8 mM CaCl2, 1 mM MgCl2, 10 mM HEPES, and 5.6 mM glucose adjusted to pH 7.45 using NaOH. Intracellular solution (ICS) consisted of 100 mM potassium aspartate, 30 mM KCl, 5 mM MgCl2, 5 mM EGTA, 4 mM Tris ATP, and 10 mM HEPES adjusted to pH 7.2 using KOH. SDS and CTAB stock solutions were made by dissolving the detergents in ECS to obtain a final concentration of 10 mM. Other concentrations were obtained by serial dilution of the stock solution in ECS. A 10 mg portion of octadecyl rhodamine B (ORB) was dissolved in 0.5 mL of DMSO (final stock concentration 27.4 mM). A 100 μL portion of stock was added to 1.9 mL of ECS to obtain 1.37 mM ORB in ECS + 5% DMSO. Lower concentrations of ORB were obtained by performing serial dilutions in ECS.
Determining Critical Micelle Concentrations of Detergents in ECS
The fluorimetric method described by Chattopadhyay and London was used to determine the CMC of SDS and CTAB in the ECS.38 Briefly, 10 μL of 1 mM diphenyl-(1,6)-hexatriene (DPH) dissolved in tetrahydrofuran was added to detergents dissolved in ECS. The fluorescence of these samples was measured using a FluoroMax-2 instrument (Horiba Instruments Inc.). The excitation wavelength was 358 nm. Emission was measured from 400 to 500 nm. Above the CMC, when detergents are present as micelles, DPH is incorporated into the micelles and there is a sharp increase in fluorescence.
Cell Culture Materials
Complete media was made by adding 50 mL fetal bovine serum, 5 mL nonessential amino acids, and 5 mL penicillin-streptomycin to 500 mL DMEM high glucose with sodium pyruvate and glutamine (Thermo Scientific). Serum free media (SFMII) for a suspension culture of HEK 293A cells was purchased from Invitrogen. Detachin was purchased from Gelantis Inc. PBS (1X) without Ca2+ and Mg2+ was obtained from Thermo Scientific.
Cell Preparation for Whole Cell Patch Clamp
HEK 293A cells (Cat. No. CRL-1573; ATCC; passages 12–17) were cultured in 175 cm2 flasks in complete media at 37 °C and 5% CO2. The cells were cultured to ∼90% confluency (∼20–25 million cells). The cells were suspended by first washing with 10 mL of PBS and suspended by treatment with 5 mL of detachin at 37 °C for 5 min. A 5 mL portion of compete media was added, and the cells were triturated. The suspension was centrifuged at 1000 rpm for 2 min (220g), and the supernatant was discarded. The cells were suspended in SFM II supplemented with 25 mM HEPES and Pen-Strep, placed in a 25 cm2 suspension flask and shaken at 75 rpm for 5 min. Employing SFMII as opposed to the regular serum free DMEM resulted in a roughly 3-fold increase in cell count and was critical for achieving improved seal resistance in each trapping zone. The cells were triturated and counted using a cytometer at this stage. The suspension was centrifuged at 1000 rpm for 2 min. The cells were suspended in (ECS). The cells were centrifuged, resuspended in ECS to a concentration of 8–12 million cells/mL, and loaded in the IonFlux 16 96-well microfluidic plate.
Staining with PKH26
In order to determine if cells were present and intact at the patch clamp sites after experimental treatments including exposure to ECS, SDS, and CTAB, we stained cells with membrane dye PKH26 according to the instructions from Sigma-Aldrich. Briefly, ∼20 million cells were suspended in serum free media using detachin as described above. The cells were centrifuged at 1400 rpm (431g) for 1 min. The supernatant was discarded, and the cells were suspended in serum free DMEM. The cells were once again centrifuged at 1400 rpm and suspended in 1 mL of diluent C. A 4 μL portion of PKH26 (1 mM in ethanol) was dissolved in 1 mL of diluent C. The cell suspension was mixed with the PKH26 solution and incubated for 5 min with periodic mixing every 5 min. A 10 mL portion of complete media was added to stop the staining process. The cells were washed with complete media three times and suspended in SFM II. The cells were then counted and suspended in ECS for use in the whole cell patch clamp experiments.
Whole Cell Patch Clamp Using the IonFlux 16
The IonFlux 16 consists of a 96-well microwell plate etched with microfluidic channels at the plate bottom as previously described by Ionescu-Zanetti et al. (Figure 1).39 Solutions flow over patched cells, giving a superfusion environment for detergent exposure. Each plate is divided into eight patterns containing two zones that trap 20 cells each. The current trace obtained by each electrode is the combined current from 20 cells. Each pattern has eight wells for compound addition that are independently controlled pneumatically. Each pattern has an IN well which was loaded with cells suspended in ECS. Before the experiment, the plate was “preprimed” for 2 min. During the preprime step, the IN well was filled with ECS. The trap wells were filled with ICS, and the compound wells had compound solutions. After the preprime step, the IN well was filled with cells suspended in ECS. The experiment consisted of four phases: prime, trap, break, and data acquisition. The timeline and pressure settings for the four phases are provided in the Supporting Information (Figure S2). In the trap phase, the fluid in the main channel flowed in pulses (no flow for 4.2 s, followed by fluid flowing for 0.8 s), which allowed cells to be trapped. The trapped cells were perforated using a rectangular pressure pulse of 4 psi amplitude for 10 s during the break phase. During the data acquisition phase, the main channel and the trap pressures were constant at 0.16 psi and 6 mmHg, respectively. The voltage for data acquisition was set at −70 mV, and the sampling rate was 500 Hz. The data was collected in frames that lasted 30 s followed by a 0.3 s period when no data was collected.
Detergent Exposure after Patching
Once the cells were trapped, they were exposed to ECS for 65 s, following which they are exposed to detergents at different concentrations. Cells were exposed to SDS and CTAB for 900 s and exposed again to ECS for 120 s. Cells were treated with ORB for 600 s and treated with ECS again for 120 s. A schematic of the experimental protocol is provided in the Supporting Information (Figure S2).
Data Analysis
The current vs time trace files were exported and processed using Microsoft Excel and MATLAB. The change in membrane current upon exposure to the detergents for different lengths of time was calculated. The time averages of current from 7.2 to 11.2 s after exposure (72.6–76.6 s) and from 931 to 935 s after exposure (991.6–995.6 s) were subtracted with the time average of current just before detergent exposure (60.6–64.6 s). For cells exposed to ORB, the time average current 10 min after exposure (666.6–670.6 s) was subtracted from the time average current immediately before exposure (60.6–64.6 s). One-way analysis of variance (ANOVA) followed by Tukey’s multicomparison test was performed to determine the statistical significance of the difference in current changes across different treatments.
Quiescent Experiments Using Cells Pretreated with Detergent
Partitioning assays have been widely used to measure the lipid bilayer composition and partition constant of detergents in the bilayer.1−4,11−13,33−37 In this experiment, different numbers of HEK 293A cells were pretreated with detergent followed by whole cell patch clamp measurements to evaluate the detergent partitioning into the membrane as a function of lipid concentration.
A fixed number of cells (e.g., 5 million cells) were suspended in ECS and incubated with various concentrations of detergents for 15 min. The suspension was then centrifuged at 1000 rpm. The cells were resuspended in ECS and patched using the IonFlux 16. The current vs time trace files were exported, and the time averaged current from the first 4 min was used as a measure of membrane permeability. This process was repeated for four different cell counts (1.5 million, 5 million, 10 million, and 15 million cells).
For CTAB alone, the partitioning behavior was also probed using a trypan blue assay. Trypan blue is a dye that is excluded from cells when the cell membrane is not compromised. In this experiment, cells were pretreated with CTAB as described above but trypan blue assay was used as a membrane permeability marker instead of a whole cell patch clamp.
Experimental Design and Data Analysis
In order to compare membrane perturbation induced by SDS, CTAB, and ORB at thermodynamic stage I and determine partition coefficients, the change in membrane behavior was generally measured at the onset of perturbation.12,33 This approach was also taken because the plateau reached after substantial exposure can be the result of differing physical states including complete cell solubilization (SDS) or saturation of the cell plasma membrane with detergent (CTAB, ORB). Figure S4 (Supporting Information) provides an example of this behavior and method of calculation for the trypan blue assay for CTAB.
For the whole cell patch clamp study, it was difficult to identify the point of onset of increase in current due to the variability in the patch quality of cells. Hence, we used the following three-step method to estimate the detergent concentrations required for the onset of increased permeability (Figure S5, Supporting Information) and calculate partition constants:
-
(1)
The rate of increase in current from control levels until a current level of ∼50 nA was measured (Figure S5, Supporting Information). The inverse of this slope provides us with a measure of the amount of detergent necessary to increase the current by 1 nA.
-
(2)
Upon examination of multiple data sets, current changes of ∼15 nA were determined to represent a reliable magnitude of change to indicate an detergent-induced current increase. The detergent necessary to increase the current by 1 nA was multiplied by 15 to obtain an estimate for detergent necessary to induce membrane currents of 15 nA.
-
(3)The detergent concentrations at the onset of increased membrane permeability were then plotted with respect to the lipid concentration and fit with a linear model. The lipid concentration in cells was estimated from the literature to be 109 lipid molecules per cell membrane.10 The slope and intercept of this line were Rb and Dw, respectively. The partition constant K was calculated using eq 6.
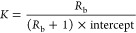
6
In step 1, a current level of 50 nA was used to calculate the slope because the cells treated using SDS, CTAB, and ORB are in a comparable physical state corresponding to thermodynamic stages I and II of the partition model. Higher current levels represent the current after cells have been solubilized by SDS. At a current level of 50 nA, the cells were not solubilized when visualized using PKH26 and ORB and gave a cell count that did not differ from the control.
XTT Assay
XTT kits from Sigma-Aldrich were used for this experiment. HEK293A cells were plated in a 96-well tissue culture plate at a density of 100000 cells per well and incubated overnight at 37 °C and 5% CO2. Cells were incubated with SDS, CTAB, ORB, or ECS for 20 min. The supernatant was then removed, and 50 μL of PBS was added to each well. 50 parts of solution A was mixed with 1 part of solution B to prepare the XTT reagent. A 30 μL portion of this reagent was added to each well and incubated at 37 °C and 5% CO2 for 2 h. The absorbance of the well was then measured at 490 and 690 nm, and the difference in absorbance between the two wavelengths was calculated. A large difference would indicate more viable cells. The percent viability was then calculated with respect to cells treated with ECS.
Results
The critical micelle concentrations (CMCs) of SDS and CTAB in ECS were determined to be 0.8–1 and 0.03–0.06 mM, respectively. The CMC values in water were also obtained giving values of 6–8 and 1–2 mM, respectively, consistent with the published data in the literature.38 Line drawings of the detergent structures are provided in Figure S3 (Supporting Information).
The change in current as a function of time measured for patched cells exposed to 0.004–2 mM SDS is presented in Figure 2. Each trace is the total current measured across 20 cells. Cells exposed to 0.2 mM SDS (number of traces, n = 11) and 2 mM SDS (n = 8) exhibited an initial sharp increase in current when compared to the ECS controls. On average, the current increase started between 2–3 and 1 s, respectively, for cells treated with 0.2 and 2 mM SDS. For cells treated with 0.2 mM SDS, the initial change in current lasted 3–4 s. The current then increased slowly for the next 67 ± 25 s. This was followed by a faster increase in current, which reached ∼50 nA at the end of 15 min. The currents for cells exposed to 2 mM SDS eventually reached open channel current levels. Except for one trace that took 871 s after exposure to reach open channel current level, other traces reached open channel current in 120 ± 61 s.
Figure 2.

(A) Exposure of HEK293A cells to 0.004 and 0.04 mM SDS does not result in an increase in conductivity when compared to the ECS controls. Exposure to 0.2 and 2 mM SDS caused a large and sudden increase in conductivity within 1–3 s. The CMC of SDS is ∼0.8 mM. Thus, SDS causes increased cell permeability even below the CMC. (B) The differences between current before exposure to detergents, 7–11 s after detergent release and 931–935 s after detergent release, were calculated. One-way ANOVA combined with Tukey test indicates that the current changes caused by 0.2 and 2 mM SDS are significantly different from the current changes caused by ECS, 0.04 mM SDS, and 0.004 mM SDS. (C) Fluorescent image of HEK293A cell exposed to 0.04 mM SDS shows intact cells. This is consistent with the absence of an increase in current. (D) Cells treated with 0.2 mM SDS are still intact after exposure for 10 min. This implies that the observed increase in current is due to the increased permeability of the membrane. (E) Cells treated with 2 mM SDS are not present in the trap region. This is consistent with current levels similar to open channel current levels. (F) Cells treated with just the ECS are intact after 10 min.
The differences between the pre-exposure current and the currents at 7.2–11.2 and 931–935 s after exposure are presented in Figure 2B. One-way analysis of variance indicated the change in current in the first 7.2–11.2 s after compound release was significantly different between the treatments (p < 0.001). Tukey’s multiple comparison analysis indicated that the current increases caused by 0.2 and 2 mM SDS were significantly different from the control and from each other (α = 0.05). Similarly, the mean change in current 931–935 s after exposure to detergents was significantly different for the different treatments (p < 0.001).
The change in current as a function of time measured for patched cells exposed to 0.001–10 mM CTAB is presented in Figure 3. The introduction of CTAB at 0.0001 mM (n = 4) and 0.001 mM (n = 4) did not cause a change in conductance as compared to the controls. The introduction of CTAB at 0.01 mM (n = 8), which is still below the CMC, caused a slow increase in current that eventually reached ∼1/3 of the open channel current. The exposure of cells to CTAB at 0.1 mM (n = 10), 1 mM (n = 12), and 10 mM (n = 4) caused a rapid increase in membrane current that reached a steady state between −40 and −70 nA. The times of onset for the change in current after compound release were 2–3 s for 0.1 mM CTAB and 1 s for both 1 and 10 mM CTAB.
Figure 3.

(A) Exposure of HEK293A cells to ECS and 0.0001 and 0.001 mM CTAB did not result in an increase in conductivity. Exposure to 0.01 mM CTAB caused a delayed increase in conductivity. The exposure of cells to 0.1 mM CTAB causes a large and sudden increase in conductivity within 1–3 s after exposure. The CMC of CTAB is ∼0.06 mM. Thus, CTAB caused increased cell permeability even below the CMC. (B) The differences between current before exposure to detergents, 7–11 s after detergent release and 931–935 s after detergent release, were calculated. One-way ANOVA combined with Tukey test indicates that the current changes caused by 0.01–10 mM CTAB are significantly different from the current changes caused by ECS, 0.0001 mM CTAB, and 0.001 mM CTAB. (C, D) Fluorescent images of HEK293A cells treated with 0.001 and 0.01 mM CTAB show that the cells are intact. This is consistent with the absence of an increase in current. (E–G) Cells treated with 0.1, 1, and 10 mM CTAB are still intact after exposure for 10 min. This implies that the observed increase in current is due to the increased permeability of the membrane. (H) Cells treated with just ECS are intact after 10 min.
ANOVA indicated that the mean current increases at 7.2–11.2 and 931–935 s after detergent exposure between all the treatments were significantly different (p < 10–5). At the 7.2–11.2 s time point, Tukey’s multiple comparison test indicates that the current changes produced by 0.1, 1, and 10 mM CTAB were significantly different from the ECS controls, 0.0001 and 0.001 mM CTAB (α = 0.05). At the 931–935 s time point, the current increases produced by 0.01, 0.1, 1, and 10 mM CTAB were significantly different from the ECS controls, 0.0001 and 0.001 mM CTAB (α = 0.05). Unlike SDS, a CTAB concentration as much as 200 times higher than the CMC did not result in open channel current levels.
The exposure of cells to detergents caused significant changes in the membrane currents observed by a whole cell patch clamp. The currents in some traps treated with 2 mM SDS reached open channel levels; however, currents in traps treated with 0.2 mM SDS and 0.1, 1, and 10 mM CTAB remained steady at approximately half the open channel levels. These steady state current levels could be either due to 20 intact but leaky cells or a mixture of intact cells and dissolved cells, leading to open channels in some fraction of the 20 trapping sites per zone. Fluorescence microscopy was employed to distinguish between these two possibilities. In order to determine if cells were intact after treatment with the various concentrations of SDS and CTAB in the microfluidic plates, we stained cells with PKH26. PKH26 is a dye that localizes to the cell membrane. The cells were then patched and treated with detergents, as described in the previous section. Figures 2C–F and 3C–H show the fluorescent images of cells stained with PKH26 after treatment with detergents for 15 min. Cells treated with ECS, 0.04 mM SDS, and 0.001 mM CTAB were intact, as indicated by their fluorescent perimeter. This is consistent with the observation that cells do not display an increase in permeability when treated with detergents at these concentrations. Cells treated with 0.2 mM SDS and 0.1–10 mM CTAB were also intact even though they display substantially increased membrane currents. The patterns treated with 2 mM SDS do not show any intact cells, consistent with the measured open channel current levels.
As an alternative approach to employing PKH26, we also employed octadecyl rhodamine B (ORB), a positively charged, amphiphilic, fluorescent detergent. This approach provided the opportunity to explore the relationship between intercalation and current increase at the lowest concentration exposures. The change in current as a function of time measured for patched cells exposed to 0.035–1.4 mM ORB is presented in Figure 4. ORB at 0.035, 0.14, and 1.4 mM increased the membrane current significantly when compared with the controls. Current induced by 0.014 mM ORB is significantly different from the ECS control but not 0.007 mM ORB. Fluorescence microscopy images (Figure 4C–F) reveal that the cells are fluorescent and intact after exposure to ORB at all three concentrations for 10 min.
Figure 4.

(A) Exposure of HEK293A to 0.035–1.4 mM ORB results in an increase in conductivity within 2–50 s that is not seen in the 5% DMSO and ECS controls. (B) The differences between current before exposure to detergents and 931–935 s after detergent release were calculated. One-way ANOVA combined with Tukey test indicates that the current changes caused by 0.035, 0.14, and 1.4 mM ORB are significantly different from the current changes caused by ECS, 0.007 mM ORB, and 0.014 mM ORB. Current induced by 0.014 mM ORB is significantly different from ECS control but not 0.007 mM ORB. (C) The fluorescent image of HEK293A cells treated with 1.37 mM ORB indicates that cells are intact after 10 min of exposure even though they exhibit a change in conductivity. (D) Cells treated with 0.137 mM ORB are still intact after exposure for 10 min. This implies that the observed increase in current for the 0.137 mM ORB is due to the increased permeability of the membrane. (E) Cells treated with 0.0137 mM ORB are fluorescent but do not exhibit a change in membrane conductivity (F). Bright field image of cells treated with just 5% DMSO in ECS that are intact after 10 min.
Figure 5 presents the current induced after cells are exposed to SDS, CTAB, and ORB for 900 s. SDS and ORB induce increased membrane permeability at 0.2 and 0.14 mM, respectively, while CTAB induces membrane permeability at 0.01 mM. CTAB induces permeability at concentrations 20 times less than SDS and 14 times less than ORB. The relative activity of SDS:CTAB:ORB is thus 1:20:14 as measured by this method.
Figure 5.
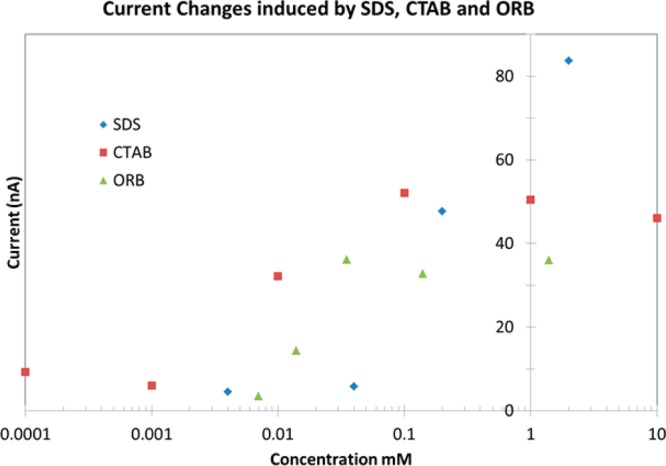
Figure showing the change in current 900 s after exposure to SDS, CTAB, and ORB dissolved in ECS. SDS and ORB induce increased membrane permeability at 0.2 mM, respectively, while CTAB and ORB induce membrane permeability at 0.01 and 0.014 mM, respectively. CTAB induces permeability at concentrations 20 times less than SDS and 14 times less than ORB. The relative activity of SDS:CTAB:ORB is thus 1:20:14 as measured by this method.
The metabolic activity of cells exposed to SDS, CTAB, and ORB as measured by the XTT assay is presented in Figure 6. Cells exposed to 0.004–0.2 mM SDS, 0.001–0.1 mM CTAB, and 0.0137–0.137 mM ORB do not show a loss in metabolic activity. Cells exposed to 2 mM SDS, 1 and 10 mM CTAB, and 1.37 mM ORB show reduced metabolic activity.
Figure 6.
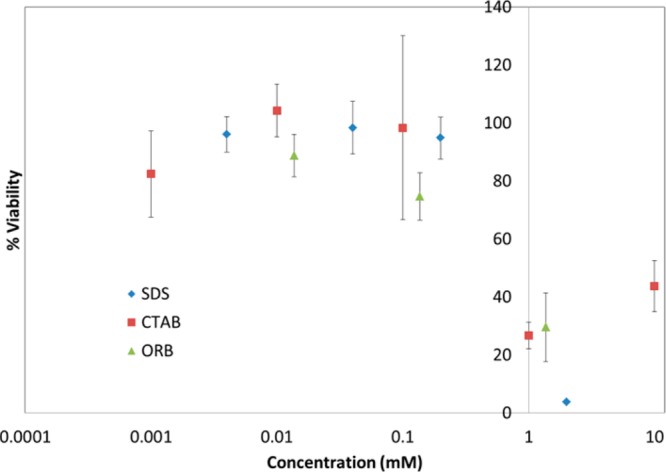
Figure showing the percent of metabolically active cells after exposure to SDS, CTAB, and ORB dissolved in ECS compared to cells exposed to ECS alone. All three compounds cause reduced cell viability at concentrations greater than 1 mM.
Detergents can increase membrane permeability either by removing lipids from the membrane or by forming stable pores in the membrane. In order to further probe the reversibility of the current induction in the plasma membrane, we exposed cells to detergents at sub-solubilization concentrations for 10, 30, or 300 s followed by ECS for 900 s (Figure 7). Cells were exposed to 0.2 mM SDS, 0.1 mM CTAB, and 0.137 mM ORB. The exposure of cells to SDS, CTAB, and ORB for 10 s is sufficient to cause increased membrane permeability. The increased permeability induced within 10 s is not reversible, even 15 min after exposure for all three materials. This result is once again consistent with the hypothesis that the increased permeability is caused by the intercalation of detergents in the cell membrane.
Figure 7.

(A) The increased conductivity induced in HEK293 A by 0.2 mM SDS after 30 s of exposure is not reversed at the end of the experiment (600 s). (B) The increased conductivity induced in HEK293 A by 0.1 mM CTAB after 10 s of exposure is not reversed at the end of the experiment (600 s). (C) The increased conductivity induced in HEK293 A by 0.137 mM ORB after 10 s of exposure is not reversed at the end of the experiment (600 s). (D) Exemplar figure showing current traces of cells exposed to ORB for 10 s in comparison to control current traces from cells exposed to only ECS or ECS + 5% DMSO. The dotted lines marked with the double arrow indicated the time when cells were exposed to ORB.
The experiments discussed so far have all been for detergent exposure under superfusion conditions. The vast majority of experiments in the literature have examined detergent exposure under quiescent conditions.11−13,33−37 In order to compare our superfusion-based data to quiescent data, we treated cells with varying amounts of detergent and then patched the cells to make the current measurements (Figure 8). The amount of detergent necessary to induce membrane conductivity increased with the cell number. Cells treated with the highest concentrations of SDS reached the open channel current plateau of the IonFlux 16 instrument, since the cells were fully solubilized. Cells exposed to CTAB and ORB remained intact even at the highest concentrations, and their current plateaus arise from detergent saturation of plasma membrane.
Figure 8.

Increasing numbers of HEK 293 A cells require increasing concentrations of (A) SDS, (B) CTAB, and (C) ORB to induce the same amount of conductivity.
The detergent concentrations at the onset of increased membrane permeability were calculated, plotted with respect to the lipid concentration, and fit using a linear model (Figure 9). The values of Rb and K were calculated as in eq 5 (Table 1). The ratio of detergent to lipid in the bilayer at the onset of increased permeability was 2.0 for SDS, 0.3 for CTAB, and 0.4 for ORB. The partition constants (K) for SDS, CTAB, and ORB are 23000, 55000, and 39000 M–1, respectively. The partition constant of CTAB measured using the trypan blue assay is 48000 M–1.
Figure 9.
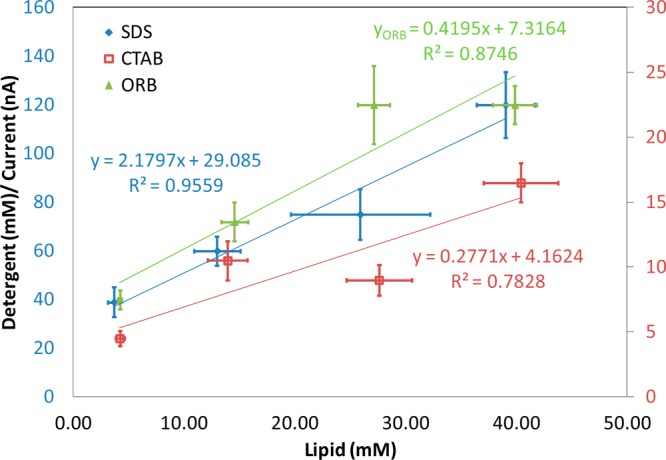
Detergent concentrations at the onset of increased permeability in current increase with increasing cell count and then further fit with a line.
Table 1. Rb and K Values for SDS, CTAB, and ORB.
| material | line equation | R2 | Rb ± standard error | Dw ± standard error | K (average) [lower bound, upper bound] |
|---|---|---|---|---|---|
| SDS | 2 X + 28.7 | 0.96 | 2 ± 0.4 | 29 ± 10 | 23000, [16300, 39200] |
| CTAB | 0.3 X + 4.2 | 0.79 | 0.3 ± 0.1 | 4.2 ± 2.8 | 55000, [29000, 476000] |
| ORB | 0.4 X + 7.3 | 0.87 | 0.4 ± 0.1 | 7.3 ± 3.1 | 39000, [24000, 99000] |
The error associated with the estimates of Rb and the intercept obtained using linear regression of the data in Figure 9 can be used to estimate the error in K (full details provided in Supporting Information). Employing the error in Rb and the intercept, a range of values for the partition constant (K) for SDS [16300–39200], CTAB [29000–476000], and ORB [24000–99000] were obtained. The partition constant of CTAB measured using the trypan blue assay ranged from 37000 to 67000 M–1. Ranges are provided rather than the standard deviation because K is not a linear function of R and the mean estimate of K calculated from R and Dw is not the midpoint of the distribution of K. Literature values for K for liposomes and erythrocytes reported to date gave no error term.12,33
Discussion
The changes in transmembrane currents observed in cells treated with different concentrations of SDS can be categorized into three groups (numbered 1–3 in Figure 2A): (1) traces that show no change in membrane conductivity compared to controls; (2) traces that show a rapid change in conductivity within 2 s of exposure to detergents and then reach a steady state current that is less than the open channel current, and (3) traces that show a rapid increase in current that reaches open channel levels over a few minutes. Traces from cells exposed to 2 mM SDS are in group 3. Visualization of these patch sites for PKH 26 stained cells indicates complete solubilization of cells (Figure 2E). By way of contrast, CTAB did not solubilize cells even at 200 times the CMC. Current traces of cells exposed to 0.2 mM SDS and 0.1–10 mM CTAB fall into group 2, and PKH26 stained cells are readily visible in the patch sites (Figure 3). Current traces from cells exposed to ECS, 0.004–0.04 mM SDS, and 0.0001–0.001 mM CTAB belong to group 1 and also exhibit readily visualized PKH26 stained cells. Cells treated with 0.01 mM CTAB exhibited behavior midway between groups I and II with both a slower rate of current increase and final current values that did not reach the saturation limit.
In group 2, all 20 PKH26 stained cells in a trap were intact after exposure to detergents for 10 min. Thus, the steady state current is not caused by the incomplete solubilization of cells causing some fraction of the trapping sites to exhibit open channel current values. This observation is consistent with the hypothesis that the increased membrane permeability is caused by detergent intercalation in the membrane. This hypothesis is further supported by the fact that ORB, which is known to localize in the membrane, also induces increased membrane permeability (Figure 4).
It has been suggested that detergent micelles are required for the solubilization of lipid membranes,9 or that solubilization occurs near the CMC.5,16 Evidence has also been presented that micelles are not necessary for solubilization.32 In our experiments, SDS solubilized cells above the CMC. By way of contrast, CTAB does not solubilize cells even at concentrations 200 times above the CMC. Thus, the presence of detergent micelles alone is not a sufficient criterion for the complete solubilization of cell membranes. Moreover, the effect of 0.2 mM SDS (below the CMC) on membrane current and cell viability is similar to the effect of 0.1 mM CTAB (above the CMC) in spite of the different aggregation states. This suggests that the aggregation state of free detergent in solution does not influence membrane solubilization. Rather, our cell-level studies agree with the physical chemical studies of Seelig et al.31 and Xia and Onyuksel32 on lipid bilayer models that it is the degree of incorporation into the membrane, headgroup shape, and charge of the detergent that determine the efficiency of cell membrane solubilization by detergents. This process may also be affected by the cycling of lipid from the membrane within the cell. From our study, it is seen that the detergent partitioning occurs in the scale of a few seconds as compared to the several minutes needed for lipid recycling.
Several models have been proposed for the perturbation of lipid membranes by detergents. Detergents can increase conductivity by intercalating in the membrane and disrupting normal lipid packing, forming stable detergent lined pores, or forming lipid-lined pores due to the formation of mixed micelles. In our whole cell patch clamp experiments under superfusion conditions, the increased membrane conductivity induced by these detergents is not reversible. The fact that the pores induced by SDS, CTAB, and ORB are not reversible over 15 min indicates the detergent is intercalated in the membrane and/or stabilizing any pores formed. The detergent could be intercalated in the membrane and cause membrane bending or thinning, as in a carpet model,40 or it could form stable detergent-lined pores, as has been reported for barrel-stave41,42 and toroidal pore models.43,44 The data reported here cannot distinguish between these two possibilities, in part because the amplifier response is averaged over 20 patched cells. This prevents clear resolution of single pore formation and the type of comparison we could make with X-ray structural data when performing single cell patch clamp studies.19,45 By way of comparison, apparently unstabilized and detergent-free, pores formed by sonoporation are reversible over 5–20 s46 and unstabilized pores in giant unilamellar vesicles close in about 100 ms.47 One possible factor that could confound the IonFlux 16 reversibility experiment is the adsorption of detergent in the walls of the microfluidic channel during the 10–300 s of exposure and subsequent release when cells are exposed to ECS alone (Figure 7D). To address this concern, we modeled the exponential release of detergent from a monolayer adsorbed in the channel wall between the compound release site and trap site where cells are located. Results presented in Figure S6 (Supporting Information) show that the cells are exposed to concentrations much lower than the concentrations necessary to induce membrane permeability.
The results in Table 1 show that the ratio of detergent to lipid in the bilayer at the onset of increased membrane permeability is 7 and 5 times greater for SDS when compared to CTAB and ORB, respectively. The partition constant for SDS in POPC and POPC/POPG vesicles has been determined by Beck and Seelig et al. to be 33000 and 37000 M–1, respectively.31 Another study by Tan and Seelig et al. reported the partition constant of SDS in POPC and POPC/POPG small and large unilamellar vesicles to range from 12500 to 70000 M–1 depending on the size and composition of the vesicles as well as the temperature.37 Our observation of 23000 M–1 is thus within the range predicted from studies of vesicle models. Beck and Seelig et al. also report a K value of 350000 M–1 for CTAB in POPC vesicles. Our partition constants of 55000 M–1 (patch clamp) and 48000 M–1 (trypan blue) measured for HEK 293A cells are substantially smaller and differ substantially from the value determined using a vesicle model.
During the estimation of partition constants for charged detergents into well-defined lipid systems such as liposomes, the electrostatic interaction of the detergent with the membrane is also taken into account. This is because the concentration of a charged detergent near the surface of the charged cell membrane need not be equal to the bulk detergent concentration in the aqueous phase.4,31,37 The local concentration of the SDS, CTAB, and ORB near the membrane surface is affected by the membrane surface charge density, buffer composition, etc.4,31,37 Thus, K is a function of Rb, Dw, and the surface potential (ψ0), as described by eq 3. The overall partition constant K is related to the partition constant that accounts for electrostatic interactions as described by eq 3.4,31,37 In eq 7, z is the charge of the detergent headgroup, F0 is Faraday’s constant, R is the gas constant, T is the temperature, and ψ0 is the membrane surface potential.
| 7 |
In our experiment, the membrane surface potential of cells was not readily available, since it could be affected by a number of biological factors such as the presence of additional proteins, sugar moieties on the surface, the physiological state of the cell, etc. In addition, the lipid membrane is cycled through the cell and it is unclear how the surface potential would be affected by that process. Hence, the binding constants calculated in this work are overall binding constants that do not take into account electrostatic interactions.
On the basis of the XTT assay, the detergent treated cells in group 2 can be further differentiated as cells with increased membrane currents that do not exhibit reduced metabolic activity (2a) and cells with increased membrane current that show decreased metabolic activity (2b). Group 2a is represented by cells exposed to 0.2 mM SDS and 0.1 mM CTAB, while 2b consists of cells exposed to 1 and 10 mM CTAB. Thus, at least at certain concentrations, detergents increase membrane permeability to ion flow without reducing metabolic activity and without damaging, to the limit of our optical resolution, the structure of the cell. Moreover, the cells treated with 0.137 mM ORB are fluorescent, show increased membrane currents, and are metabolically active. This shows that an amphiphilic molecule with a charged headgroup can intercalate in cell membranes and cause increased membrane conductivity but not reduce the metabolic activity of the cells.
Conclusion
Detergents increased the transmembrane conductivity of cells very rapidly, on the order of a few seconds. SDS and CTAB increased the membrane conductivity of cells even when their bulk concentration was below the CMC. The experiments support previous conclusions that detergent headgroup charge and shape affect cell membrane solubilization. Fluorescence microscopy images indicated that cells with increased conductivity were intact. Patch clamp experiments combined with microscopy also showed that ORB both intercalates in the membrane and increases membrane conductivity significantly. The increased membrane conductivity induced by detergents at sub-solubilizing concentrations does not decrease once compound exposure is stopped. These results suggest intercalated detergent remains in the plasma membrane for at least 15 min. Moreover, our studies provide partition constants for SDS, CTAB, and ORB in non-erythrocytic mammalian cell membranes.
Acknowledgments
This project has been funded in part with Federal funds from the National Institutes of Health, National Institute of Biomedical Imaging and Bioengineering, under Award EB005028.
Supporting Information Available
Schematic of detergent–cell membrane interaction (S1); description of IonFlux 16; line diagrams of detergents used in study (S3); membrane partitioning of CTAB using trypan blue assay (S4); estimation of detergent concentration at the onset of membrane permeability (S5); amount of detergent released from the microfluidic channel wall (S6); estimation of error in partition constant estimate. This material is available free of charge via the Internet at http://pubs.acs.org.
The authors declare no competing financial interest.
Funding Statement
National Institutes of Health, United States
Supplementary Material
References
- Heerklotz H.; Seelig J. Correlation of Membrane/Water Partition Coefficients of Detergents with the Critical Micelle Concentration. Biophys. J. 2000, 78, 2435–2440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lichtenberg D.; Ahyayauch H.; Alonso A.; Goni F. M. Detergent Solubilization of Lipid Bilayers: A Balance of Driving Forces. Trends Biochem. Sci. 2013, 38, 85–93. [DOI] [PubMed] [Google Scholar]
- Lasch J. Interaction of Detergents with Lipid Vesicles. Biochim. Biophys. Acta 1995, 1241, 269–292. [DOI] [PubMed] [Google Scholar]
- Heerklotz H. Interactions of Surfactants with Lipid Membranes. Q. Rev. Biophys. 2008, 41, 205–264. [DOI] [PubMed] [Google Scholar]
- Kragh-Hansen U.; le Maire M.; Møller J. V. The Mechanism of Detergent Solubilization of Liposomes and Protein-Containing Membranes. Biophys. J. 1998, 75, 2932–2946. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van der Woude I.; Wagenaar A.; Meekel A. A.; ter Beest M. B.; Ruiters M. H.; Engberts J. B.; Hoekstra D. Novel Pyridinium Surfactants for Efficient, Nontoxic in Vitro Gene Delivery. Proc. Natl. Acad. Sci. U.S.A. 1997, 94, 1160–1165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hamouda T.; Myc A.; Donovan B.; Shih A. Y.; Reuter J. D.; Baker J. R. Jr. A Novel Surfactant Nanoemulsion with a Unique Non-Irritant Topical Antimicrobial Activity Against Bacteria, Enveloped Viruses and Fungi. Microbiol. Res. 2001, 156, 1–7. [DOI] [PubMed] [Google Scholar]
- Sau T. K.; Murphy C. J. Seeded High Yield Synthesis of Short Au Nanorods in Aqueous Solution. Langmuir 2004, 20, 6414–6420. [DOI] [PubMed] [Google Scholar]
- Deo N.; Somasundaran P. Disintegration of Liposomes by Surfactants: Mechanism of Protein and Cholesterol Effects. Langmuir 2003, 19, 2007–2012. [Google Scholar]
- Alberts B.Molecular biology of the cell; Garland Science: New York, 2002. [Google Scholar]
- Preté P. S. C.; Malheiros S. V. P.; Meirelles N. C.; de Paula E. Quantitative Assessment of Human Erythrocyte Membrane Solubilization by Triton X-100. Biophys. Chem. 2002, 97, 1–5. [DOI] [PubMed] [Google Scholar]
- Preté P. S. C.; Gomes K.; Malheiros S. V. P.; Meirelles N. C.; de Paula E. Solubilization of Human Erythrocyte Membranes by Non-Ionic Surfactants of the Polyoxyethylene Alkyl Ethers Series. Biophys. Chem. 2002, 97, 45. [DOI] [PubMed] [Google Scholar]
- Preté P. S. C.; Domingues C. C.; Meirelles N. C.; Malheiros S. V. P.; Goñi F. M.; de Paula E.; Schreier S. Multiple Stages of Detergent-Erythrocyte Membrane interaction—A Spin Label Study. Biochim. Biophys. Acta 2011, 1808, 164–170. [DOI] [PubMed] [Google Scholar]
- Goñi F. M.; Alonso A. Spectroscopic Techniques in the Study of Membrane Solubilization, Reconstitution and Permeabilization by Detergents. Biochim. Biophys. Acta 2000, 1508, 51–68. [DOI] [PubMed] [Google Scholar]
- Koley D.; Bard A. J. Triton X-100 Concentration Effects on Membrane Permeability of a Single HeLa Cell by Scanning Electrochemical Microscopy (SECM). Proc. Natl. Acad. Sci. U.S.A. 2010, 107, 16783–16787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Partearroyo M. A.; Ostolaza H.; Goñi F. M.; Barberá-Guillem E. Surfactant-Induced Cell Toxicity and Cell Lysis. Biochem. Pharmacol. 1990, 40, 1323–1328. [DOI] [PubMed] [Google Scholar]
- Hong S.; Bielinska A. U.; Mecke A.; Keszler B.; Beals J. L.; Shi X.; Balogh L.; Orr B. G.; Baker J. R. Jr.; Banaszak Holl M. M. Interaction of Poly(Amidoamine) Dendrimers with Supported Lipid Bilayers and Cells: Hole Formation and the Relation to Transport. Bioconjugate Chem. 2004, 15, 774–782. [DOI] [PubMed] [Google Scholar]
- Hong S.; Leroueil P. R.; Janus E. K.; Peters J. L.; Kober M. M.; Islam M. T.; Orr B. G.; Baker J. R. Jr.; Banaszak Holl M. M. Interaction of Polycationic Polymers with Supported Lipid Bilayers and Cells: Nanoscale Hole Formation and Enhanced Membrane Permeability. Bioconjugate Chem. 2006, 17, 728–734. [DOI] [PubMed] [Google Scholar]
- Chen J.; Hessler J. A.; Putchakayala K.; Panama B. K.; Khan D. P.; Hong S.; Mullen D. G.; Dimaggio S. C.; Som A.; Tew G. N.; et al. Cationic Nanoparticles Induce Nanoscale Disruption in Living Cell Plasma Membranes. J. Phys. Chem. B 2009, 113, 11179–11185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leroueil P. R.; Hong S.; Mecke A.; Baker J. R. Jr.; Orr B. G.; Banaszak Holl M. M. Nanoparticle Interaction with Biological Membranes: Does Nanotechnology Present a Janus Face?. Acc. Chem. Res. 2007, 40, 335–342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carmona-Ribeiro A. M.; de Melo Carrasco L. D. Cationic Antimicrobial Polymers and their Assemblies. Int. J. Mol. Sci. 2013, 14, 9906–9946. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sgolastra F.; deRonde B. M.; Sarapas J. M.; Som A.; Tew G. N. Designing Mimics of Membrane Active Proteins. Acc. Chem. Res. 2013, 46, 2977–2987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mecke A.; Lee D.; Ramamoorthy A.; Orr B. G.; Banaszak Holl M. M. Membrane Thinning due to Antimicrobial Peptide Binding: An Atomic Force Microscopy Study of MSI-78 in Lipid Bilayers. Biophys. J. 2005, 89, 4043–4050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mecke A.; Majoros I. J.; Patri A. K.; Baker J. R.; Banaszak Holl M. M.; Orr B. G. Lipid Bilayer Disruption by Polycationic Polymers: The Roles of Size and Chemical Functional Group. Langmuir 2005, 21, 10348–10354. [DOI] [PubMed] [Google Scholar]
- Li S.; Malmstadt N. Deformation and Poration of Lipid Bilayer Membranes by Cationic Nanoparticles. Soft Matter 2013, 9, 4969–4976. [Google Scholar]
- Schulz M.; Olubummo A.; Binder W. H. Beyond the Lipid-Bilayer: Interaction of Polymers and Nanoparticles with Membranes. Soft Matter 2012, 8, 4849–4864. [Google Scholar]
- Lin J.; Alexander-Katz A. Cell Membranes Open “Doors” for Cationic Nanoparticles/Biomolecules: Insights into Uptake Kinetics. ACS Nano 2013, 7, 10799–10808. [DOI] [PubMed] [Google Scholar]
- Carney R. P.; Astier Y.; Carney T. M.; Voïtchovsky K.; Jacob Silva P. H.; Stellacci F. Electrical Method to Quantify Nanoparticle Interaction with Lipid Bilayers. ACS Nano 2013, 7, 932–942. [DOI] [PubMed] [Google Scholar]
- Mishra A.; Lai G. H.; Schmidt N. W.; Sun V. Z.; Rodriguez A. R.; Tong R.; Tang L.; Cheng J.; Deming T. J.; Kamei D. T. Translocation of HIV TAT Peptide and Analogues Induced by Multiplexed Membrane and Cytoskeletal Interactions. Proc. Natl. Acad. Sci. U.S.A. 2011, 108, 16883–16888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leroueil P. R.; Berry S. A.; Duthie K.; Han G.; Rotello V. M.; McNerny D. Q.; Baker J. R. Jr.; Orr B. G.; Holl M. M. Wide Varieties of Cationic Nanoparticles Induce Defects in Supported Lipid Bilayers. Nano Lett. 2008, 8, 420–424. [DOI] [PubMed] [Google Scholar]
- Beck A.; Li-Blatter X.; Seelig A.; Seelig J. On the Interaction of Ionic Detergents with Lipid Membranes Thermodynamic Comparison of n-Alkyl-N+(CH3)3 and n-Alkyl-SO4–. J. Phys. Chem. B 2010, 114, 15862–15871. [DOI] [PubMed] [Google Scholar]
- Xia W. J.; Onyuksel H. Mechanistic Studies on Surfactant-Induced Membrane Permeability Enhancement. Pharm. Res. 2000, 17, 612–618. [DOI] [PubMed] [Google Scholar]
- Heerklotz H.; Seelig J. Leakage and Lysis of Lipid Membranes Induced by the Lipopeptide Surfactin. Eur. Biophys. J. 2007, 36, 305–314. [DOI] [PubMed] [Google Scholar]
- Heerklotz H.; Seelig J. Titration Calorimetry of Surfactant-Membrane Partitioning and Membrane Solubilization. Biochim. Biophys. Acta 2000, 1508, 69–85. [DOI] [PubMed] [Google Scholar]
- de la Maza A.; Parra J. L. Vesicle-Micelle Structural Transition of Phosphatidylcholine Bilayers and Triton X-100. Biochem. J. 1994, 303, 907–914. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de la Maza A.; Coderch L.; Gonzalez P.; Parra J. L. Subsolubilizing Alterations Caused by Alkyl Glucosides in Phosphatidylcholine Liposomes. J. Controlled Release 1998, 52, 159–168. [DOI] [PubMed] [Google Scholar]
- Tan A.; Ziegler A.; Steinbauer B.; Seelig J. Thermodynamics of Sodium Dodecyl Sulfate Partitioning into Lipid Membranes. Biophys. J. 2002, 83, 1547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chattopadhyay A.; London E. Fluorimetric Determination of Critical Micelle Concentration Avoiding Interference from Detergent Charge. Anal. Biochem. 1984, 139, 408–412. [DOI] [PubMed] [Google Scholar]
- Spencer C. I.; Li N.; Chen Q.; Johnson J.; Nevill T.; Kammonen J.; Ionescu-Zanetti C. Ion Channel Pharmacology Under Flow: Automation Via Well-Plate Microfluidics. Assay Drug Dev. Technol. 2012, 10, 313–324. [DOI] [PubMed] [Google Scholar]
- Pouny Y.; Rapaport D.; Mor A.; Nicolas P.; Shai Y. Interaction of Antimicrobial Dermaseptin and its Fluorescently Labeled Analogs with Phospholipid Membranes. Biochemistry 1992, 31, 12416–12423. [DOI] [PubMed] [Google Scholar]
- Baumann G.; Mueller P. A. Molecular Model of Membrane Excitability. J. Supramol. Struct. 1974, 2, 538–557. [DOI] [PubMed] [Google Scholar]
- He K.; Ludtke S. J.; Worcester D. L.; Huang H. W. Neutron Scattering in the Plane of Membranes: Structure of Alamethicin Pores. Biophys. J. 1996, 70, 2659–2666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matsuzaki K. Magainins as Paradigm for the Mode of Action of Pore Forming Polypeptides. Biochim. Biophys. Acta 1998, 1376, 391–400. [DOI] [PubMed] [Google Scholar]
- Ludtke S. J.; He K.; Heller W. T.; Harroun T. A.; Yang L.; Huang H. W. Membrane Pores Induced by Magainin. Biochemistry 1996, 35, 13723–13728. [DOI] [PubMed] [Google Scholar]
- Yang L.; Gordon V. D.; Trinkle D. R.; Schmidt N. W.; Davis M. A.; DeVries C.; Som A.; Cronan J. E.; Tew G. N.; Wong G. C. Mechanism of a Prototypical Synthetic Membrane-Active Antimicrobial: Efficient Hole-Punching Via Interaction with Negative Intrinsic Curvature Lipids. Proc. Natl. Acad. Sci. U.S.A. 2008, 105, 20595–20600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou Y.; Kumon R. E.; Cui J.; Deng C. X. The Size of Sonoporation Pores on the Cell Membrane. Ultrasound Med. Biol. 2009, 35, 1756–1760. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sandre O.; Moreaux L.; Brochard-Wyart F. Dynamics of Transient Pores in Stretched Vesicles. Proc. Natl. Acad. Sci. U.S.A. 1999, 96, 10591–10596. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.