

Abstract
Objectives
Dengue represents one of the most serious life-threatening vector-borne infectious diseases that afflicts approximately 50 million people across the globe annually. Whilst symptomatic infections are frequently reported, asymptomatic dengue remains largely unnoticed. Therefore, we sought to investigate the immune correlates conferring protection to individuals that remain clinically asymptomatic.
Methods
We determined the levels of neutralizing antibodies (nAbs) and gene expression profiles of host immune factors in individuals with asymptomatic infections, and whose cognate household members showed symptoms consistent to clinical dengue infection.
Results
We observed broad down-regulation of host defense response (innate, adaptive and matrix metalloprotease) genes in asymptomatic individuals as against symptomatic patients, with selective up-regulation of distinct genes that have been associated with protection. Selected down-regulated genes include: TNF α (TNF), IL8, C1S, factor B (CFB), IL2, IL3, IL4, IL5, IL8, IL9, IL10 and IL13, CD80, CD28, and IL18, MMP8, MMP10, MMP12, MMP15, MMP16, and MMP24. Selected up-regulated genes include: RANTES (CCL5), MIP-1α (CCL3L1/CCL3L3), MIP-1β (CCL4L1), TGFβ (TGFB), and TIMP1.
Conclusion
Our findings highlight the potential association of certain host genes conferring protection against clinical dengue. These data are valuable to better explore the mysteries behind the hitherto poorly understood immunopathogenesis of subclinical dengue infection.
Introduction
Dengue represents one of the world's most dreadful vector-borne flavivirus infections with its increasing incidence, making it a major disease burden in the tropics and subtropics. Global estimates show that ∼3.6 billion individuals, representing ∼55% of the world's population, are at increasing risk for contracting dengue virus (DENV) infection. Annual estimates show that the number of dengue cases reported is ∼390 million, of which ∼96 million represent dengue hemorrhagic fever (DHF) or dengue shock syndrome (DSS), and ∼300 million others representing mild or asymptomatic cases [1]. DENV is a member of the family Flaviviridae and genus Flavivirus, a positive-stranded RNA virus transmitted by Aedes aegypti (A.aegypti) and A. albopictus mosquitoes. There are four serotypes namely DENV 1, 2, 3 and 4, which can all elicit the complete spectrum of disease severity, from the most common asymptomatic subclinical infection to severe plasma leakage, shock, haemorrhage and, in some cases, death. The viruses are maintained in an A. aegypti-human-A. aegypti cycle, where humans acquire infection following the bite of a DENV-infected female mosquito. The mosquito reportedly feeds on multiple individuals over a given period of time. It is, thus, common that the same mosquito could infect several members of the same household [2]–[4].
The recent WHO classification has categorized the disease as dengue without warning signs (DWOS), dengue with warning signs (DWWS) and severe dengue (SD) [5]. A DWOS DENV infection could manifest as asymptomatic or a ‘flu-like syndrome’, while DWWS could be characterized by sudden onset of fever, usually accompanied by nonspecific signs and symptoms, such as headache, back pain, stiffness, and flushed facial skin [6]. In severe dengue infections, for instance DHF (DWWS), plasma leakage and thrombocytopenia can be life threatening, especially following hypovolemic shock in severe dengue. Few studies of asymptomatic dengue have been performed and thus knowledge on the full burden of dengue infection is limited [7]. Primary DENV infections are often asymptomatic and will generate immunity to the homologous strain. However, ∼90% of DWWS (DHF) reportedly occurs following second exposure to a heterologous strain of DENV [8]. This will greatly increase the future risk of onset of SD following asymptomatic infections as the previous DENV infection had gone undetected. Therefore, subclinical infections provide ample opportunities for researchers to explore host immune factors that confer protection against clinical DENV infections.
Viral virulence [9], host genetic background [10], T-cell activation [11], viral burden [12], antibody dependent enhancement [13] and autoantibodies [14] are reportedly implicated in disease pathogenesis. Host immune factors [15]–[20] have also been reported as contributing to onset of DENV infection. It is hypothesized that DENV infection of monocytes/macrophages increases T-cell activation leading to release of cytokines and chemical mediators resulting in increased vascular permeability, plasma leakage, shock, and malfunction of the coagulatory system, culminating in hemorrhage and shock. Evidence that implicates immune factors in dengue severity is derived from chemical mediators, such as tumor necrosis factors (TNF), interleukin-1 (IL1), IL2, IL6, platelet-activating factor (PAF), complement components C3a and C5a, and histamine [21]. CD4+ T cells produce a plethora of cytokines, which recruit numerous other cytokines and chemical mediators that further increase vascular permeability [22]. Nonetheless, these studies have all concentrated on the apparent clinical dengue infection but not on asymptomatic cases, which therefore, remains a grey area of investigation. Here, we sought to investigate the molecular mechanisms underlying asymptomatic DENV infection by determining neutralizing antibody (nAb) levels and analyzing immunological genes expression profiles.
Materials and Methods
Specimens
Blood specimens were collected from clinical dengue cases admitted to University Malaya Medical Center (UMMC), Ampang Hospital and Klang Tengku Ampuan Rahimah Hospital, Malaysia. Blood specimens were also collected from household members of individuals presenting with clinical dengue for the investigation. The study protocols were approved by the institutional review board of the University of Malaya Medical Center (FPU-DOF-BK-012-05-R01) and from both Ampang and Klang Hospitals (Ethics no. NMRR-10-683-6420). Written informed consent from patients and asymptomatic donors was obtained, and the study was conducted in accordance with the Declaration of Helsinki. At least 1 household member's blood was collected per dengue patient. Clinical information and medical history for each patient and the accompanying household member was documented. Blood samples of dengue patients were collected during both the acute and convalescence stage. In contrast, only one time point of blood sampling was done for the suspected asymptomatic household member.
Peripheral blood mononuclear cells
All blood samples were collected in BD Vacutainer (BD, Franklin Lakes, NJ, USA) tubes for extraction of serum. For the isolation of peripheral blood mononuclear cells (PBMCs), blood samples were collected in Vacutainer Plus (Plastic) Sterile Evacuated K2 EDTA spray dried Blood Collection Tubes (BD, Plymouth, UK). Isolation of PBMC was done using the Ficoll-Hypaque (Lymphoprep; Axis-Shield, Oslo, Norway) density gradient centrifugation [23]. Briefly, the blood was centrifuged at 2500 rpm for 15 min at 4°C; plasma was separated from the whole blood and stored at −20°C. Blood cells were washed with RPMI1640 (Roswell Park Memorial Institute) media and were gently layered over Ficoll in 50 ml centrifuge tubes. This was followed by centrifugation at 2000 rpm for 20 min without brake to separate the buffy coat from red cells. The buffy coat containing PMBCs was collected and washed with RPMI and the total number of PMBCs was determined using a hemocytometer. The isolated PBMCs were stored in liquid nitrogen until further use.
Diagnosis of dengue
Sera from symptomatic cases and their corresponding household members were collected and subjected to in-house IgM-Capture ELISA [24] for IgM detection, hemagglutination inhibition (HI) test [25] for total dengue antibody detection, and quantitative real-time (qRT-PCR) [26] for detection of viral RNA. Samples were considered as confirmed dengue positive based on the criteria that any of the diagnostic assays carried out showed positivity, that is: 1) dengue nucleic acid detection through PCR, 2) IgM detection during both acute and convalescence phases, 3) sero-conversion, or 4) 4-fold increase in HI titre from acute to convalescence phase. As for the presumptive dengue positive cases, samples that showed IgM detection only during the acute phase, HI titres of more than 1280 in a single serum, or >1∶640 nAb titres of 50% reduction in neutralization by the plaque reduction neutralization test (PRNT) were considered [5]. PRNT was performed to determine the levels of neutralizing antibodies (nAb) against each of the dengue virus serotypes, following a protocol slightly modified than others [27]. Briefly, porcine kidney epithelial cells (PS cells) were seeded in 24-well plates and incubated overnight. Serum samples were diluted to 1∶10 dilution and heat-inactivated at 56°C for 30 min, followed by four-fold serial dilution using L-15 media containing 1% heat-inactivated fetal bovine serum (FBS). Equal volumes of virus with PFU of 15–30 per well were added to the diluted sera, and incubated at 37°C for 1 hour. Prototype strains of dengue viruses were used and these included Den1-Hawaiian, Den2-New Guinea C, Den3-H87 and Den4-H241. Subsequently, the virus-antibody mixture was added onto the cell monolayers in the 24-well plates, and incubated at 37°C for 3 hours. An overlay medium of 3% carboxymethyl-cellulose (CMC) was then added to the monolayers and incubated at 37°C without CO2 for 7 days, after which cells were washed and stained with 1% naphthalene black. The viral plaques that formed were enumerated and plaque neutralization titers were determined accordingly.
Sample selection
Twenty-nine pairs of patients and their accompanying asymptomatic household members were recruited for gene expression investigations based on positive results for DENV infection (see the diagnosis section). Notably, among the 29 pairs, there were four that corresponded to two distinct patients paired with two asymptomatic household members each. In further microarray experiments, only the acute blood samples of the 29 patient and accompanying household member pairs were tested.
Microarray hybridization
The stored PBMCs of patients and asymptomatic subjects were subjected to microarray hybridization (Miltenyi Biotec GmbH) as described in the manufacturer's instructions. Briefly, RNA was isolated using standard RNA extraction protocols (NucleoSpin RNA II, Macherey-Nagel). RNA samples quality was determined by the Agilent 2100 Bioanalyzer platform (Agilent Technologies). The results were visualized in a gel image and electropherogram using the Agilent 2100 Bioanalyzer expert software. The RNA integrity number (RIN) and the overall quality of total RNA were determined using the same software. RNA with RIN value >6 was subjected to linear T7-based RNA amplification to obtain sufficient antisense RNA. Amplified RNA (aRNA) was again examined on the Agilent 2100 Bioanalyzer platform, and the samples subjected to fluorescent labeling according to PIQOR user manual. The fluorescent-labeled samples were hybridized overnight to human antisense topic-defined PIQOR Immunology Microarrays (http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GPL17653) using the a-Hyb Hybridization Station. Each dengue patient-asymptomatic contact pair's RNA samples were hybridized with each other. Fluorescence signals of the hybridized PIQOR Microarrays were detected using a laser scanner (Agilent Technologies). ImaGene software (BioDiscovery, Hawthorne CA, USA) was used to obtain signal and local background intensities for each spot of the microarray images. These data are available in NCBI's Gene Expression Omnibus (GEO) database through GEO accession ID GSE50634 (http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE50634).
Microarray data processing and statistical analysis
The median signal and median local background intensity values for each spot of the microarray images (obtained from ImaGene) were imported into Partek Genomics Suite (Partek Inc., St Louis MO, USA) for statistical analysis. Background subtraction was performed using the median intensities of the arrays' four replicate spots for each gene. These signal values were normalized by shifting their minimum to 2 and then converted to log2 notation for statistical analysis. Quality control of the results, using principal component analysis (PCA) and supervised hierarchical clustering (data not shown), revealed two samples to be extreme outliers so they and their paired household members' were excluded from further analysis. The paired sample t-test was used to analyze the remaining 27 household pairs, comparing the asymptomatic member's gene expression to that of the patient's.
The t-test's fold change and statistical p-values were then imported into Spotfire DecisionSite with Functional Genomics (TIBCO Spotfire Boston MA, USA) for further analysis and graphical representation. The volcano plot in Figure 1 depicts the paired sample t-test results, showing each gene's asymptomatic to symptomatic fold changes and that comparisons' statistical significance. The heat map in Figure 2 depicts a supervised clustering of all 27 samples' mean-subtracted log2 signal values for genes that were differentially expressed at more than 2 and less than −2 linear fold changes (p<0.01).
Figure 1. Asymptomatic versus patient paired volcano plot to determine differentially expressed genes.

The x-axis shows the paired-sample t-test log2 fold change of asymptomatic versus symptomatic patients, whereas the y-axis shows the −log10 of the paired t-test p-values of asymptomatic versus symptomatic patients. Each dot represents a gene and the dot color depicts the gene's expression as shown by the standard deviation (SD) of the paired t-test log2 fold change, e.g. shades of blue for less than or equal to −1 SD, grey for between −1 SD and +1 SD, and red for greater than or equal to +1 SD. A total of 345 genes were differentially expressed at more than 2 and less than −2 linear fold changes (p<0.01) and were considered statistically significant.
Figure 2. Supervised hierarchical correlation clustering of 27 sample pairs.
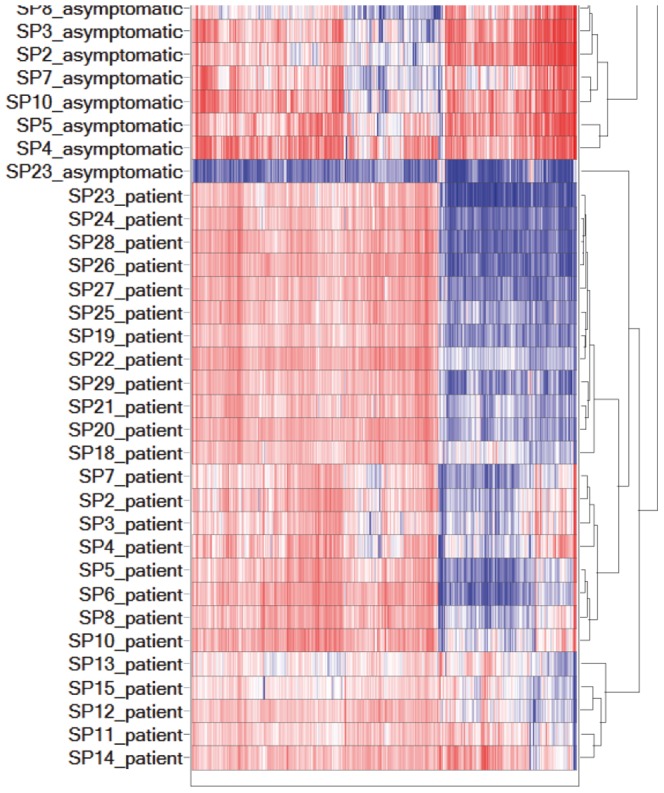
The heat map shows the gene expression of samples in columns, with a dendogram representing their similarity based on correlation. The samples' mean-subtracted log2 signal values were clustered using complete linkage (maximum) method and the genes' expression is shown with blue representing down-regulation, white for no change (zero), and red for up-regulation.
Pathway enrichment analysis
The Spotfire DecisionSite-processed data was uploaded to Ingenuity Pathway Analysis (IPA) software for pathway enrichment analysis in order to understand their biological meaning. Ingenuity Knowledge Base (Genes Only) served as reference set for p-value calculation and Fisher's exact test was used to calculate the p-values for the canonical pathways. Only human genes and relationships were used from the database, and only experimentally observed or high confidence (predicted) observations were considered in the analysis. In addition, genes which were differentially expressed at more than 2 and less than −2 linear fold changes (p<0.01) were selected, resulting in 334 analysis-ready genes. The analysis produced a total number of 337 canonical pathways (data not shown). Among the canonical pathways, those specific to other diseases, not directly known to be relevant to dengue immune-pathogenesis/-protection, or general/unrelated signalling pathways were eliminated, leaving 53 canonical pathways for detailed analysis. The 53 selected canonical pathways were then classified into three groups, namely. innate immune response, adaptive immune response and matrix metalloproteases. Individual pathways of the innate immune response and adaptive immune response groups were studied and combined into a single mega pathway for each, whereas the matrix metalloproteases group pathway was used as-is as it was the only pathway in the group. Both the innate and adaptive mega pathways included numerous cytokines and chemokines, which are important for mediating communication among immune cells, and thus were also studied as a separate mega pathway.
Results
Patient characteristics
As shown in Table 1, we obtained 30 confirmed dengue positive cases of which 29 were dengue patients while 1 individual was an asymptomatic household member. There were 28 presumptive positive dengue samples of which all are asymptomatic household members. From the summary of diagnostics in Table 2, the confirmed dengue patients showed only 2 (7%) positive results for viral RNA detection. However, we found one (3%) asymptomatic household member with positive detection in PCR for viral RNA which confirmed dengue positivity. Our results showed 28 (97%) patients with positive paired IgM or seroconversion and 13 (45%) patients with HI titer >1∶1280. For presumptive positive dengue, we observed 18 (62%) asymptomatic household members with positive IgM in a single sample and 10 (34%) of these individuals had HI titre >1280. Additionally, for the PRNT that was performed, our investigations showed 9 (31%) patients and 11 (37.9%) asymptomatic individuals had nAbs against one DENV serotype as shown in Table 3. We observed dengue patients had a higher percentage (65.5%) of nAbs for polytypic infection in comparison with asymptomatic individuals (48.3%). We also observed that almost half of the asymptomatic individuals exhibited polytypic nAbs towards DENV infection.
Table 1. Diagnostic results of dengue patients and their accompanying household members.
| Family | Identification | Infection status | Patient's sample | Presumptive positive dengue | Neutralizing antibodies | ||||||
| Confirmed dengue positive | Presumptive positive dengue | PCR | IgM | HI | Dengue 1 | Dengue 2 | Dengue 3 | Dengue 4 | |||
| 1 | P1 | √ | P1a | Negative | Negative | 160 | 80 | <10 | <10 | <10 | |
| P1c | Positive | 640 | 320 | <10 | <10 | <10 | |||||
| H1 | √ | H1 | Negative | Positive | 640 | <10 | 320 | <10 | <10 | ||
| 2 | P2 | √ | P2a | Negative | Positive | 320 | 160 | 20 | <10 | <10 | |
| P2c | Positive | 640 | 320 | <10 | <10 | <10 | |||||
| H2 | √ | H2 | Negative | Negative | 640 | <10 | 80 | 320 | 2560 | ||
| 3 | P3 | √ | P3a | Negative | Positive | 640 | 80 | 20 | <10 | <10 | |
| P3c | Positive | 1280 | 160 | 40 | <10 | <10 | |||||
| H3 | √ | H3 | Negative | Negative | 640 | >640 | <10 | <10 | <10 | ||
| 4 | P4 | √ | P4a | Negative | Negative | 40 | <10 | <10 | <10 | <10 | |
| P4c | Positive | 160 | <10 | 20 | <10 | <10 | |||||
| H4 | √ | H4 | Negative | Negative | 2560 | 640 | <10 | <10 | <10 | ||
| 5 | P5 | √ | P5a | Negative | Positive | 640 | 160 | 20 | <10 | <10 | |
| P5c | Positive | 2560 | 1280 | <10 | <10 | <10 | |||||
| H5 | √ | H5 | Negative | Positive | 5120 | 320 | <10 | <10 | <10 | ||
| 6 | P6 | √ | P6a | Negative | Positive | 320 | 640 | <10 | <10 | <10 | |
| P6c | Positive | >10240 | 5120 | <10 | 20 | <10 | |||||
| H6 | √ | H6 | Negative | Positive | 640 | 640 | <10 | <10 | <10 | ||
| 7 | P7 | √ | P7a | Negative | Negative | 2560 | 640 | 40 | <10 | <10 | |
| P7c | Negative | >10240 | >10240 | 80 | <10 | <10 | |||||
| H7 | √ | H7 | Negative | Positive | 640 | 320 | <10 | 40 | <10 | ||
| 8 | P8 | √ | P8a | Negative | Negative | 2560 | 2560 | 40 | >10240 | 20 | |
| P8c | Positive | >10240 | >10240 | 80 | >10240 | 20 | |||||
| H8 | √ | H8 | Negative | Negative | 2560 | 320 | 20 | <10 | <10 | ||
| 9 | P9 | √ | P9a | Negative | Positive | >10240 | 2560 | <10 | <10 | <10 | |
| P9c | Positive | >10240 | 5120 | 20 | <10 | <10 | |||||
| H9 | √ | H9 | Negative | Negative | 5120 | <10 | <10 | <10 | 160 | ||
| 10 | P10 | √ | P10a | Negative | Negative | 1280 | 160 | <10 | <10 | <10 | |
| P10c | Positive | >10240 | 640 | <10 | <10 | <10 | |||||
| H10 | √ | H10 | Negative | Negative | 2560 | <10 | <10 | <10 | 80 | ||
| 11 | P11 | √ | P11a | Dengue 1 | Negative | <10 | 160 | 640 | 640 | 160 | |
| P11c | Positive | 1280 | 160 | >2560 | >2560 | >2560 | |||||
| H11 | √ | H11 | Negative | Positive | >10240 | >2560 | >2560 | >2560 | >2560 | ||
| 12 | P12 | √ | P12a | Negative | Positive | 160 | 160 | >2560 | >2560 | 2560 | |
| P12c | Positive | >10240 | 160 | >2560 | >2560 | >2560 | |||||
| H12 | √ | H12 | Negative | Positive | <10 | 40 | 160 | 10 | <10 | ||
| 13 | P13 | √ | P13a | Negative | Positive | <10 | 160 | 40 | 2560 | 2560 | |
| P13c | Positive | <10 | 640 | 640 | >2560 | >2560 | |||||
| H13 | √ | H13 | Negative | Positive | <10 | 160 | <10 | <10 | <10 | ||
| 14 | P14 | √ | P14a | Negative | Positive | 20 | 640 | 160 | 40 | 160 | |
| P14c | Positive | >10240 | >2560 | >2560 | 160 | 2560 | |||||
| H14 | √ | H14 | Negative | Negative | 10 | 640 | <10 | 40 | <10 | ||
| 15 | P15 | √ | P15a | Negative | Positive | 20 | 640 | 160 | 40 | 160 | |
| P15c | Positive | >10240 | >2560 | >2560 | 160 | 2560 | |||||
| H15 | √ | H15 | Negative | Negative | 80 | <10 | <10 | <10 | <10 | ||
| 16 | P16 | √ | P16a | Negative | Positive | 1280 | <10 | <10 | 40 | <10 | |
| P16c | Positive | >10240 | <10 | <10 | 2560 | <10 | |||||
| H16 | √ | H16 | Negative | Positive | 20 | <10 | <10 | <10 | <10 | ||
| 17 | P17 | √ | P17a | Negative | Positive | 1280 | <10 | 640 | <10 | <10 | |
| P17c | Positive | 2560 | <10 | 1280 | <10 | <10 | |||||
| H17 | √ | H17 | Negative | Positive | 10 | <10 | <10 | <10 | <10 | ||
| 18 | P18 | √ | P18a | Negative | Positive | 10 | <10 | <10 | <10 | <10 | |
| P18c | Positive | 160 | 160 | <10 | <10 | <10 | |||||
| H18 | √ | H18 | Negative | Positive | 10 | <10 | <10 | <10 | <10 | ||
| 19 | P19 | √ | P19a | Negative | Positive | >10240 | >2560 | 160 | 640 | <10 | |
| P19c | Positive | >10240 | >2560 | >2560 | 2560 | 160 | |||||
| H19 | √ | H19 | Negative | Positive | 1280 | 40 | 40 | 160 | 40 | ||
| 20 | P20 | √ | P20a | Negative | Positive | 640 | 160 | 640 | <10 | <10 | |
| P20c | Positive | 2560 | 160 | 640 | <10 | <10 | |||||
| H20 | √ | H20 | Negative | Positive | 640 | <10 | 160 | <10 | <10 | ||
| 21 | P21 | √ | P21a | Negative | Positive | 2560 | 640 | 160 | 40 | 640 | |
| P21c | Positive | 2560 | 640 | 160 | 40 | 640 | |||||
| H21 | √ | H21 | Dengue 3 | Negative | 320 | 160 | 160 | 640 | 160 | ||
| 22 | P22 | √ | P22a | Negative | Positive | 2560 | 640 | 160 | 40 | 640 | |
| P22c | Positive | 2560 | 640 | 160 | 40 | 640 | |||||
| H22 | √ | H22 | Negative | Positive | 10 | <10 | <10 | 40 | <10 | ||
| 23 | P23 | √ | P23a | Negative | Positive | 5120 | 40 | 2560 | 10 | 40 | |
| P23c | Positive | >10240 | 640 | 2560 | 10 | 40 | |||||
| H23 | √ | H23 | Negative | Equivocal | 10240 | 10 | 160 | 10 | 10 | ||
| 24 | P24 | √ | P24a | Negative | Positive | 20 | <10 | 10 | <10 | <10 | |
| P24c | Positive | 40 | <10 | 40 | <10 | <10 | |||||
| H24 | √ | H24 | Negative | Positive | 320 | 40 | 40 | 40 | 10 | ||
| 25 | P25 | √ | P25a | Negative | Positive | 2560 | <10 | 640 | 640 | <10 | |
| P25c | Positive | >10240 | <10 | 640 | 2560 | <10 | |||||
| H25 | √ | H25 | Negative | Negative | 160 | <10 | 40 | <10 | <10 | ||
| 26 | P26 | √ | P26a | Dengue 3 | Positive | 160 | <10 | <10 | 160 | <10 | |
| P26c | Positive | 1280 | <10 | <10 | 640 | <10 | |||||
| H26 | √ | H26 | Negative | Positive | 5120 | <10 | 2560 | 1280 | <10 | ||
| 27 | P27 | √ | P27a | Negative | Positive | 1280 | 40 | 1280 | 10 | <10 | |
| P27c | Positive | 10240 | 2560 | 1280 | 10 | <10 | |||||
| H27 | √ | H27 | Negative | Negative | 640 | <10 | 40 | 10 | <10 | ||
| 28 | P28 | √ | P28a | Negative | Positive | 1280 | 40 | 640 | 160 | 40 | |
| P28c | Positive | 5120 | 2560 | 640 | 160 | 40 | |||||
| H28 | √ | H28 | Negative | Equivocal | 80 | 10 | 10 | 10 | 10 | ||
| 29 | P29 | √ | P29a | Negative | Negative | <10 | <10 | 10 | 10 | 10 | |
| P29c | Positive | 10 | 160 | 10 | 10 | 10 | |||||
| H29 | √ | H29 | Negative | Positive | 2560 | 40 | 1280 | 640 | 40 | ||
*P indicates dengue patient while H indicate household member.
**a indicates acute samples while c indicates convalescence sample.
Table 2. Summary of dengue diagnostic results obtained from patients and their corresponding/accompanying household members.
| Diagnostic test | Patients (%) | Household members (%) |
| Real time RT-PCR | 2 (7) | 1 (3) |
| Positive IgM | 28 (97) | 18 (62) |
| HI>1280 | 13 (45) | 10 (34) |
Table 3. Summary of plaque reduction neutralization test (PRNT) results obtained from patients and their corresponding/accompanying household members.
| Category | Monotypic Infection | Polytypic Infection | No neutralizing antibody detected | Total |
| Dengue patient acute sera | 9 (31.0) | 19 (65.5) | 1 (3.4) | 29 |
| Asymptomatic dengue individuals | 11 (37.9) | 14 (48.3) | 4 (13.8) | 29 |
The numbers within parentheses indicate percentage.
Differentially expressed genes determined from paired-sample t-test
The volcano plot, Figure 1 displays relative gene expression, showing the standard deviation (SD) of the paired t-test log2 fold change against −log10 p-value of the paired t-test. The high statistical significance of genes as depicted in this plot provided confidence in reliability of the expression values. A total of 345 genes were differentially expressed at more than 2 and less than −2 linear fold changes (p<0.01) and were considered statistically significant.
Supervised hierarchical clustering discriminates symptomatic patients from asymptomatic individuals
The hierarchical supervised correlation clustering of 27 sample pairs (SPs), Figure 2, shows that a majority of the patients clustered together as they had similar gene expression patterns, except for SP16 and SP17 patients who separated the asymptomatic household members into two clusters. This separation, however, was not considered significant as it was attributed to batch effect as suggested by the PCA plot (data not shown). SP23_asymptomatic, which clustered by itself in a single group, was not deemed as an outlier according to the PCA plot and thus was retained for further analysis. Based on the overall clustering, it is clear that the gene expression patterns of the asymptomatic household members' were different from the patients'.
Gene profiling for biological significance
Pathway enrichment analysis of the 27 paired sample data was performed to glean the biological significance of the differential expression pattern observed between asymptomatic individuals and symptomatic patients. The resulting 53 selected pathways with their respective expressed genes shown in Table 4 (see Table S1 for more details) were grouped according to the two broad categories of reported dengue host defence mechanisms, namely innate immune response (Figure S1) and adaptive immune response (Figure S2). Additionally, the global role of the cytokines and chemokines, which mediate communication between the immune cells, was studied through a separate pathway in Figure S3. The matrix metalloproteases pathway was studied as well and shown in Figure S4.
Table 4. 53 of the selected canonical pathways studied, with genes up- and down-regulated in the subclinical/asymptomatic dengue are indicated.
| Group | Canonical Pathway | Differential Expression | ||
| No. | Name | Up-regulation | Down-regulation | |
| Innate | 1 | Acute phase response Signaling | AKT1, FOS, IKBKG, NFKB1, NFKBIA, TRADD | C1S, CFB, CRP, IL1A, IL1RAP, MAP2K7, SOCS3, TNF |
| 2 | CCR5 signaling in macrophages | CALM1, CCL5, CD247, CD3D, CD3E, FOS, PRKCB | FASLG, PLCG1, PRKCE | |
| 3 | CD27 signaling in lymphocytes | FOS, IKBKG, NFKB1, NFKBIA | MAP2K7, MAP3K2, MAP3K4 | |
| 4 | CD40 signaling | FOS, IKBKG, NFKB1, NFKBIA | ICAM1, LTA, MAP2K7, PIK3C2A, PIK3C2G | |
| 5 | Complement system | - | C1S, CFB | |
| 6 | Crosstalk between dendritic cells and natural killer cells | ACTB, ICAM3, IL2RG, KLRC4-KLRK1/KLRK1, KLRD1, LTB, NFKB1, PRF1 | CD28, CD80, CSF2, CSF2RB, FASLG, IL2, IL2RB, IL3, IL4, LTA, LTBR, TNF | |
| 7 | Dendritic cell maturation | AKT1, B2M, CD1D, FCGR3A, HLA-DQA1, IKBKG, LTB, NFKB1, NFKBIA | CD1A, CD1B, CD80, COL11A2, COL1A1, COL1A2, COL2A1, COL3A1, CSF2, ICAM1, IL10, IL1A, LTA, LTBR, LY75, PIK3C2A, PIK3C2G, PLCB1, PLCB4, PLCD1, PLCD3, PLCE1, PLCG1, TNF | |
| 8 | iNOS signaling | CALM1, FOS, IKBKG, IRF1, NFKB1, NFKBIA | - | |
| 9 | Mechanisms of viral exit from host cells | ACTB, PRKCB | NEDD4, PRKCE | |
| 10 | MIF regulation of innate immunity | CD74, FOS, NFKB1, NFKBIA | PLA2G12B, PLA2G3, PLA2G4A | |
| 11 | Natural killer cell signaling | AKT1, CD247, FCGR3A, FYN, KLRC4-KLRK1/KLRK1, KLRD1, LCK, PRKCB | PIK3C2A, PIK3C2G, PLCG1, PRKCE, VAV3 | |
| 12 | Production of Nitric Oxide and reactive Oxygen species in macrophages | AKT1, CLU, FOS, IKBKG, IRF1, NFKB1, NFKBIA, PRKCB, RHOA | IL4, MAP2K7, MAP3K2, MAP3K4, PIK3C2A, PIK3C2G, PLCG1, PPARA, PRKCE, TNF | |
| 13 | Role of pattern recognition receptors in recognition of bacteria and viruses | CASP1, CCL5, NFKB1, PRKCB | IL10, IL2, PIK3C2A, PIK3C2G, PRKCE, TLR6, TNF | |
| 14 | Role of PKR in interferon induction and antiviral response | AKT1, IKBKG, IRF1, NFKB1, NFKBIA | TNF | |
| 15 | Role of RIG1-like receptors in antiviral innate immunity | IKBKG, NFKB1, NFKBIA | CASP10, IFNA2 | |
| 16 | Toll-like receptor signaling | FOS, IKBKG, NFKB1, NFKBIA | PPARA, TLR6 | |
| 17 | TWEAK signaling | IKBKG, NFKB1, NFKBIA, TNFRSF25, TRADD | CASP7 | |
| Adaptive | 18 | B cell development | HLA-DQA1, IL7R, PTPRC | CD80, RAG1 |
| 19 | B cell receptor signaling | AKT1, CALM1, CSK, IKBKG, NFATC3, NFKB1, NFKBIA, PRKCB, PTEN, PTPRC | EGR1, MAP2K7, MAP3K2, MAP3K4, PIK3C2A, PIK3C2G, VAV3 | |
| 20 | B cell activating factor signaling | FOS, IKBKG, NFATC3, NFKB1, NFKBIA | MAP2K7 | |
| 21 | CD28 signaling in T helper cells | AKT1, CALM1, CD247, CD3D, CD3E, CSK, FOS, FYN, HLA-DQA1, IKBKG, ITK, LCK, NFATC3, NFKB1, NFKBIA, PTPRC | CD28, CD80, IL2, PIK3C2A, PIK3C2G, PLCG1 | |
| 22 | CTLA4 signaling in cytotoxic T lymphocytes | AKT1, B2M, CD247, CD3D, CD3E, FYN, LCK | CD28, CD80, PIK3C2A, PIK3C2G, PLCG1 | |
| 23 | Cytotoxic T lymphocytes-mediated apoptosis of target cells | B2M, CD247, CD3D, CD3E, HLA-DQA1, PRF1 | BCL2, CASP7, FASLG | |
| 24 | Differential regulation of cytokine production in intestinal epithelial cells by IL17A and IL18 | CCL5 | CSF2, CXCL1, IL3, IL9, IL10, IL13, IL1A, TNF | |
| 25 | Differential regulation of cytokine production in macrophages and T helper cells by IL17A and IL18 | CCL5 | CSF2, CXCL1, IL3, IL9, IL10, IL13, TNF | |
| 26 | Fcγ Receptor-mediated Phagocytosis in Macrophages and Monocytes | ACTB, AKT1, FCGR3A, FGR, FYN, PRKCB, PTEN | CBL, CSF2, PIK3C2G, PLCG1, PRKCE, VAV3, YES1 | |
| 27 | FcγRIIB Signaling in B Lymphocytes | AKT1 | PIK3C2A, PIK3C2G | |
| 28 | NF-kB activation by viruses | AKT1, IKBKG, ITGB1, ITGB2, LCK, NFKB1, NFKBIA, PRKCB | ITGA2, ITGAV, PIK3C2A, PIK3C2G, PRKCE | |
| 29 | T cell receptor signaling | CALM1, CD247, CD3D, CD3E, CSK, FOS, FYN, IKBKG, ITK, LCK, NFATC3, NFKB1, NFKBIA, PTPRC | CBL, CD28, PIK3C2A, PIK3C2G, PLCG1, VAV3 | |
| 30 | T helper cell differentiation | HLA-DQA1, IL10RA, IL2RG, TBX21, TGFB1 | CD28, CD80, FOXP3, IL10, IL13, IL18R1, IL2, IL4, IL5, TGFBR1, TNF | |
| Cytokine | 31 | Chemokine signaling | CALM1, CCL5, CXCR4, FOS, MPRIP, PRKCB, RHOA | PIK3C2G, PLCB1, PLCB4, PLCG1 |
| 32 | GM-CSF signaling | AKT1, PRKCB | CCND1, CSF2, CSF2RA, CSF2RB, PIK3C2A, PIK3C2G | |
| 33 | IL1 signaling | FOS, IKBKG, NFKB1, NFKBIA | ADCY8, IL1A, IL1RAP, MAP2K7 | |
| 34 | IL10 signaling | FOS, IKBKG, IL10RA, NFKB1, NFKBIA | IL10, IL1A, IL1RAP, IL1RAPL1, IL1RAPL2, SOCS3, TNF | |
| 35 | IL15 production | IRF1, NFKB1 | MST1R | |
| 36 | IL15 signaling | AKT1, IL2RG, LCK, NFKB1, STAT5A | BCL2, CSF2, IL2RB, IL4, PIK3C2A, PIK3C2G, PLCG1 | |
| 37 | IL2 signaling | AKT1, FOS, IL2RG, LCK, STAT5A | IL2, IL2RB, PIK3C2A, PIK3C2G | |
| 38 | IL22 signaling | AKT1, STAT5A | IL22RA1, SOCS3 | |
| 39 | IL3 signaling | AKT1, FOS, PRKCB, STAT5A | CRKL, CSF2RB, IL3, PIK3C2A, PIK3C2G, PRKCE | |
| 40 | IL4 signaling | AKT1, HLA-DQA1, IL2RG, NFATC3 | IL4, PIK3C2A, PIK3C2G | |
| 41 | IL6 signaling | AKT1, FOS, IKBKG, NFKB1, NFKBIA | COL1A1, CRP, IL1A, IL1RAP, IL1RAPL1, IL1RAPL2, IL8, MAP2K7, PIK3C2A, PIK3C2G, SOCS3, SRF, TNF | |
| 42 | IL8 signaling | AKT1, FOS, IKBKG, ITGAX, ITGB2, NFKB1, PRKCB, RHOA | BCL2, CCND1, CXCL1, FIGF, ICAM1, IL8, IL9, ITGAV, KDR, PGF, PIK3C2A, PIK3C2G, PRKCE, VEGFC | |
| 43 | IL9 signaling | IL2RG, NFKB1, STAT5A | IL9, PIK3C2A, PIK3C2G, SOCS3, TNF | |
| MMP | 44 | Inhibition of matrix metalloproteases | TIMP1 | MMP10, MMP12, MMP15, MMP16, MMP24, MMP8 |
| Interaction between innate, cytokines and adaptive | 45 | TREM1 signaling | AKT1, CASP1, ITGAX, ITGB1, NFKB1, STAT5A | CSF2, CXCL3, ICAM1, IL10, IL8, PLCG1, TLR6, TNF |
| 46 | Interferon signaling | IRF1, PTPN2, TAP1 | BCL2, IFNAR2 | |
| 47 | IL17 signaling | AKT1, IL17RA, NFKB1, TIMP1 | CRP, CXCL1, IL19, IL8, PIK3C2A, PIK3C2G | |
| 48 | IL17A signaling in fibroblasts | FOS, IKBKG, IL17RA, NFKB1, NFKBIA | - | |
| 49 | Regulation of IL2 expression in activated and anergic T lymphocytes | CALM1, CD247, CD3D, CD3E, FOS, FYN, IKBKG, NFATC3, NFKB1, NFKBIA, TGFB1 | CD28, CD80, IL2, MAP2K7, PLCG1, TGFB2, TGFBR1, VAV3 | |
| 50 | TGF-β Signaling | BMP4, FOS, TGFB1 | BCL2, BMP2, BMP7, FOXH1, PIAS4, TGFB2, TGFBR1 | |
| 51 | Communication between innate and adaptive immune cells | B2M, CCL3L1/CCL3L3, CCL5 | CD28, CD80, CSF2, IL10, IL1A, IL2, IL3, IL4, IL5, IL8, TLR6, TNF | |
| 52 | IL12 signaling and production in macrophages | AKT1, CLU, FOS, IKBKG, IRF1, NFKB1, PRKCB, TGFB1 | IFNA2, IL10, IL4, MST1R, PIK3C2A, PIK3C2G, PRKCE, TGFB2, TNF | |
| 53 | Role of cytokines in mediating communication between immune cells | TGFB1 | CSF2, IFNA2, IL10, IL13, IL1A, IL2, IL3, IL4, IL5, IL8, TNF | |
The pathways have been grouped according to the broad categories of reported dengue host defence mechanisms. The gene symbols are based on HUGO Gene Nomenclature Committee (HGNC) symbols. See Table S1 for more details as it also includes the list of genes that showed no change for each of the pathways listed and provides linear fold change for genes discussed in the manuscript.
Innate immune response
The expression of Toll-like receptors (TLRs), which are critical for pathogen recognition, was observed (Table 4; Figure S1, and S5 for pathway legends). The TLR complex indicated in the figure includes several genes (TLR2, TLR3, TLR4, TLR7 and TLR9) that remained unchanged between asymptomatic household members and symptomatic patients, and one (TLR6) that was down-regulated. We also observed expression of CD40, CD83 and CD86 but at no significant level change. However, CD80 expression was significantly down-regulated. Besides that, we observed a no change in CD32A (FCGR2B), but an up-regulation in CD16A (FCGR3A). Regarding the complement system, we observed down regulation of C1S and complement factor B (CFB). C1S is a protein component that cleaves C2 and C4 into C2a, C2b, C4a and C4b in the classical pathway, while CFB is a component of the alternative pathway.
Adaptive immune response
The expression of both T-cell antigen presentation molecules MHC class 1, found on all nucleated cells, and MHC class II, expressed on B-cells, macrophages and dendritic cells, were up-regulated. However, there was no change in the expression levels of both CD8 and CD4 complexes. Meanwhile, the results showed a down-regulation of CD28 (Table 4; Figure S2, S5), which is expressed on T cells to provide co-stimulatory signal for T-cell activation upon binding to CD80, which was also down-regulated (Table 4; Figure S2). We also observed an up-regulation in asymptomatic siblings for T-bet (TBX21) (Table 4; Figure S2), which is a critical regulator for T helper cell 1 (Th1) differentiation and an inducer for the production of IFNγ (IFNG) [28]. It simultaneously inhibits the opposing Th2 and Th17 differentiation program. Additionally, FOXP3, a transcription factor involved in regulatory T cell (Treg) development was seen to be down-regulated.
Cytokines and chemokines
Cytokines and chemokines are important inflammatory mediators of both innate and adaptive immune responses that are stimulated in response to pathogens. In our study, we found a down regulation of IL1α (IL1A), IL2, IL3, IL4, IL5, IL8, IL9, IL10, IL13, IL18R (IL18R1), IL22R (IL22RA1), TNFα (TNF), TGFβR (TGFBR1), GM-CSF (CSF2) and MIP-2 (CXCL3) expression as shown in Table 4 and Figure S3. Observed up-regulated genes included MIP1α (CCL3L1/CCL3L3), MIP1β (CCL4L1), RANTES (CCL5), TGFβ1 (TGFB1), IL4R IL10R (IL10RA) and IL7R. Genes with no significant change in expression level between asymptomatic siblings and symptomatic patients included IL1β (IL1B), IL6, IL7, IL12 (IL12B), IL15, IL17A, IL17F, IL18, IL21, IL24, IFNα (IFNA1/IFNA13), IFNβ (IFNB1), IFNγ (IFNG), CCL2 and CCR5.
Matrix metalloproteases (MMPs)
MMPs are a family of proteins that cleave most extracellular matrix constituents. We observed the reduced expression of MT-MMP (includes MMP15, MMP16, and MMP24, among others) and Extracellular MMP (includes MMP8, MMP10, and MMP12, among others) complexes (Figure S4, S5). The genes MMP8, MMP10, MMP12, MMP15, MMP16 and MMP24 were down-regulated in asymptomatic siblings, while MMP2 and MMP9 were detected with similar expression level in both groups of subjects (Table 4; Figure S4, S5). TIMP1, a factor that inhibits activity of MMPs was up-regulated.
Discussion
The differences in dengue clinical manifestations are widely being investigated to better understand the disease pathogenesis. Herein, we investigated the protective mechanisms contributing to the apparent lack of clinical manifestations in individuals that remain asymptomatic when compared to clinical dengue cases.
We note that selection criteria for gene expression study herein are based solely on the presumptive positive IgM levels and HI titres of single asymptomatic samples as there were difficulties in obtaining second voluntary blood samples from the household members. From the diagnostic results, only three subjects were PCR positive. Virus titre could not be determined through PCR for most of the samples due to the duration of infection. The samples collected for PCR were mostly from days 5 to 6, by which time viral titres may have declined, with increasing antibody levels. Thus, PRNT, a gold standard test used widely to determine and quantify nAbs against DENV infection [29], was carried out to mitigate these limitations. We observed that asymptomatic individuals had high concentration of nAbs against DENV infection. Despite having polytypic infections, half of these asymptomatic individuals are protected against clinical dengue. It has been reported that antibodies could play a greater role than immune cells in heterologous DENV infection [30]. This entails that individuals infected with DENV manifest clinical symptoms differently with some presenting with more severe symptoms than others due to the presence of pre-existing nAbs in the asymptomatic individuals. However, apart from nAbs, protection from DENV infection in asymptomatic individuals could also be attributed to other factors viz., host genetic factors [31], which comprise a complex network of genes that are expressed differentially in the asymptomatic individuals.
Inflammation involves the activation of immune cells and recruitment of specific immune cells to the site of inflammation [32], eventually contributing to nonspecific clearance of DENV, albeit causing mild symptoms. However, in some cases, exaggerated inflammation can culminate in detrimental clinical manifestations, pathognomonic of DWWS/SD. The mutual synergism between the innate and adaptive immune responses is the key to responding to infectious microorganisms. Here, we hypothesize that dengue asymptomatic individuals downplay the factors responsible for inflammation and regulate the associated immune factors to a level that is merely required to facilitate virus clearance.
Genes of the innate immune response
The innate host response to DENV is mediated by dendritic cells (DCs), phagocytes (macrophages) and natural killer lymphocytes which sense viral proteins or nucleic acids through Toll-like receptors (TLRs). In our study, we observed no change in the expressions of TLR3 and TLR7 between the asymptomatic individuals and symptomatic patients. TLR 3 has been shown to play an important role in restricting DENV infection in synergy with RIG-I and MDA5 [33]. We could not assess this relationship due to the absence of RIG-I and MDA5 from the array used. Besides, each TLR recognizes distinct microbial components and activates different signaling pathways by selective utilization of adaptor molecules. Since dengue is a single stranded RNA virus, TLR3 may not respond as it is known to recognize viral double stranded RNA [34]. However, in support of TLR3 as a protective factor, recent studies have demonstrated that its expression induces type I interferon and inhibits the replication of DENV in different cell lines [35], [36]. In contrast, TLR7 receptor has been shown to recognize the virus, with significantly higher expression levels in DHF patients than in DF [37], [38]. Although this does not explain the role of TLR7 in the protective mechanism(s) preventing the development of clinical symptoms, a no change observation in the gene may relatively reduce the effect of more severe clinical symptoms that could have resulted from an up-regulation of TLR7.
FcγR is critical in binding to the DENV immune complex, enhancing virus uptake by DCs and macrophages [39]. We observed a no change in CD32A (FCGR2B), but an up-regulation of CD16A (FCGR3A). These Fcγ receptors connect the innate and the adaptive immune responses by transmitting activating signals to natural killer lymphocytes and myeloid cell upon recognition of Fc of IgG [40]. CD32A is expressed in all myeloid cells, platelets, and endothelial cells, whereas CD16A is present on monocytes, macrophages, NK cells and γ/δ T cells [41]. The up-regulation of CD16A in our study suggests the activation of antibody response by the NK cells, which may be a protective factor in dengue clinical manifestation. This is supported in other studies that CD16A is the only Fc receptor expressed on NK cells and is responsible for IgG-initiated antibody dependent cell-mediated cytotoxicity (ADCC) [42], which may reduce the incidence of antibody-dependent enhancement (ADE), one of the hallmarks of dengue pathogenesis.
The complement system is an important arm of innate immunity. Earlier studies suggest that complement activation plays a role in the pathogenesis of DHF [43], [44]. We observed a down regulation of C1S and factor B complement expression in asymptomatic siblings. An in vitro study provided evidence that complements promote the uptake of DENV into myeloid cells through CR3, augmenting the infection [45]. Several studies have shown that DENV non-structural protein 1 (NS1) binds to C4 and C1S, regulating complement activation and hence could be involved in DHF pathogenesis [44], [46]. Therefore, with the decreased expression of C1S in our study, the activation of complement system is reduced, suppressing severe dengue pathogenesis and allowing only subclinical symptoms to be manifested.
Genes of the adaptive immune response
The adaptive immune system includes antibody-secreting B cells and cytotoxic T cells that specifically and efficiently target the pathogen and infected cells. Activated DENV-infected antigen presenting cells (APCs), such as DCs, macrophages or B-cells, acquire an activation profile with elevated expression of CD40, CD80, CD83 and CD86 [47], [48]. These surface proteins assist the communication between DENV-infected cells and T cells. In the present study, we observed a down regulation of CD80 expression in asymptomatic siblings, and detected no expression change for CD40, CD83 and CD86. An in vitro study demonstrated that defect in co-stimulatory proteins failed to provide adequate signaling for T cell activation and proliferation, resulting in impaired cell-mediated response [49]. This suggests an ineffective interaction between DENV-infected immune cells and T cells, despite up-regulation of MHC I (Beta-2-microglobulin, B2M) and MHC II (HLA-DQA1) complex members' expressions. Nonetheless, there was no change in the expression levels of CD8 and CD4 T-cell complexes between both groups. This suggests that the host immune response may have reduced disease severity that is caused by T-cell-mediated tissue damage. It was reported in a study that the magnitude of T-cell responses correlates with dengue disease severity due to cross reactive T cells [50]. Moreover, the down regulation of CD28, required to provide the co-stimulatory signal for T-cell activation, and cytokines produced by T cells, namely IL2, IL4, IL5, IL10 and IL18, further reinforces the broad suppression of cell-mediated immune response in asymptomatic siblings.
Naïve T helper cells can differentiate into Th1, Th2, Th17, follicular T helper cells (TFH) and T regulatory cells (Tregs) [51]. We observed an up regulation of T-bet in asymptomatic siblings. This is suggestive of a partial Th1 response, which plays a role in viral clearance but not in causing clinical symptoms. Furthermore, it has been shown that T-bet inhibits production of Th2 and Th17 cytokines [52], and with a reduced expression of IL4 in asymptomatic siblings, it is reasonable to infer that Th2 responses are suppressed by T-bet.
Cytokines and chemokines role in protection
Cytokines and chemokines are important inflammatory mediators that are stimulated in response to pathogens and certain cytokine profiles have been associated with dengue disease severity [53]–[58]. It has been suggested that the pathogenesis of DF/DHF involves amplified cytokine production, also known as “cytokine storm” [59], [60]. This ultimately leads to excessive immune activation that increases vascular permeability [58] and causes plasma leakage and shock. Our results showed a general down regulation in expression of cytokines and chemokines, specifically for IL2, IL3, IL4, IL5, IL8, IL9, IL10 and IL13. However, elevated cytokines including IL2, IL4, IL6, IL8, IL10, IL13, IL18, TNFα (TNFA), IFNγ (IFNG), TGFβ (TGFB), CCL2 and CCL3 have been reported in patients with DHF [53], [54], [57], [61]–[64]. This suggests a limited inflammatory response trigger, sufficient for viral clearance yet below the level that can lead to clinical symptoms. Similarly, other studies have shown some of these cytokines to serve as predictive markers for progression to dengue with warning signs when cytokine kinetics and profiles of dengue patients at different phases of illness were investigated [65].
We observed down regulation of TNFα herein, suggesting an outcome similar to healthy controls. Studies have reported a significantly higher level of TNFα in dengue-infected patients compared to healthy controls, and the elevated TNFα is associated with disease severity [66], [67]. Hence, the down regulation seen in our study's asymptomatic individuals may have been a protective factor in controlling the severity of the disease. In addition, TNFα308 allele polymorphism was observed among the DHF patients and they expressed higher levels of TNFα [68]. We also observed down regulation of IL1α, which may point to the possible role of TNFα and IL1α in dengue pathogenesis. This is supported by another study that has demonstrated TNFα and IL1α to increase vascular permeability in vitro [69].
DENV-infected DCs or macrophages can produce chemokines including IL8, RANTES (CCL5), MCP-1, MIP1 and IP-10. IL8, a chemokine produced by stimulated monocytes is down-regulated in our study. This is consistent with a previous study that showed association of higher IL8 level with disease severity [70]. It has been shown that IL8 has a chemoattractant and degranulation ability for neutrophils [71]. It can also increase the permeability of the endothelial cell monolayer, as shown in vitro [72]. Increased endothelial cell permeability may lead to the development of plasma leakage, a clinical manifestation of severe dengue patients. Therefore, the reduced expression of IL8 in asymptomatic siblings could contribute to their protective mechanisms in preventing clinical manifestations. RANTES is a chemokine that recruits T cells, eosinophils and basophils to the site of infection. Our results showed an up-regulation of RANTES in asymptomatic siblings. We hypothesize that high RANTES levels may play an important role in mitigating clinical dengue symptoms in infected individuals. This is because a previous study has reported that plasma RANTES levels is lower in acute dengue infants compared to infants with other febrile infections [73]. In another study, an association was observed between decreased RANTES levels in patients with acute dengue fever and lower platelet number or thrombocytopenia [74]. Comparing our results with these earlier studies, higher RANTES expression appears to be advantageous in mitigating clinical disease.
In this study, we observed an up-regulation of MIP-1β. MIP-1β has been reported to have an association with good prognosis factors identified in different disease models [75], [76]. In one study, MIP-1β levels were higher in patients with mild dengue fever compared to patients with severe clinical manifestation [77]. The authors suggested that the relationship of MIP-1β and NK cells may play a role in dengue protective mechanisms. Consistent with these findings, our results of up-regulated MIP-1β in asymptomatic siblings further reinforces the possible protective role of MIP-1β.
TGFβ is a multifunctional cytokine that can act as a proinflammatory or anti-inflammatory cytokine depending on its concentration. Our results showed a higher TGFβ expression level in asymptomatic patients, which suggests a protective role for TGFβ in dengue disease. This is supported by a study that showed a TGFβ polymorphism that demonstrated high TGFβ production was associated with protection and mild clinical manifestations [78]. However, on the contrary, there are other studies that report a significantly higher plasma level of TGFβ in DHF patients as compared to DF patients [54], [79]. Given these effects, it remains to be ascertained whether TGFβ expression contributes to a pathogenic or protective role in dengue infection.
MMPs role in dengue infection
MMPs are a family that cleave most extracellular matrix constituents. In our study, we detected no change in MMP9 and MMP2 expressions between patients and asymptomatic subjects, but did observe a broad down-regulation of MMP8, MMP10, MMP12, MMP15, MMP16 and MMP24 expressions. Meanwhile, the inhibitor of MMPs, TIMP1 was up-regulated, which may explain the broad down-regulation of extracellular MMPs that we observed. These findings indicated an overall suppression of MMPs, which may contribute to the protective mechanism in dengue infection. This is reasonable given a study that pointed to the role of MMP in triggering plasma leakage in DHF patients [80]. Activation of IL6, TNFα, IL8, and TGFβ can drive MMP production, which is correlated to the vascular leakage characteristics of DHF [54], [71], [81], [82]. Additionally, overexpression of MMP9 and to a lesser extent MMP2 were observed to have a role in enhancing vascular permeability [20]. This study suggests the role of MMP9 and MMP2 in dengue pathogenesis.
In summary, DENV infection among asymptomatic individuals, as compared to clinical dengue patients, provides a wealth of information associating gene expression and immune correlates. Overall, we observed broad down-regulation of host defense response (innate, adaptive, cytokines and matrix metalloprotease) genes in asymptomatic individuals against symptomatic patients, with selective up-regulation of distinct genes that have been associated with protection. Several of the genes examined herein deserve further assessment to correlate expression with conferring protection against clinical DENV infection. Given the emergence of a wider network of immune molecules in subclinical DENV infection, additional studies may be warranted to investigate the molecular targets associated with improved clinical manifestations and increase understanding of the pathogenesis of subclinical DENV infection.
Supporting Information
Innate immune response mega canonical pathway created using Ingenuity Pathway Analysis. TLR complex includes TLR6 (down-regulated in asymptomatic individuals), TLR3 (no change in expression between the two groups) and TLR7 (no change), among others. Please refer to Figure S5 for a legend explaining Pathway molecule symbols. For additional information, visit IPA legend help page at http://ingenuity.force.com/ipa/articles/Feature_Description/Legend
(TIF)
Adaptive immune response mega canonical pathway created using Ingenuity Pathway Analysis. Please refer to Figure S5 for a legend explaining Pathway molecule symbols. For additional information, visit IPA legend help page at http://ingenuity.force.com/ipa/articles/Feature_Description/Legend
(TIF)
Role of cytokines in mediating communication between immune cell canonical pathways from Ingenuity Pathway Analysis. Please refer to Figure S5 for a legend explaining Pathway molecule symbols. For additional information, visit IPA legend help page at http://ingenuity.force.com/ipa/articles/Feature_Description/Legend
(TIF)
Inhibition of matrix metalloproteases canonical pathway from Ingenuity Pathway Analysis. MT-MMP complex includes MMP15, MMP16, and MMP24, among others, and these three were down-regulated in asymptomatic individuals. Extracellular MMP complex includes MMP8, MMP10, and MMP12, among others, and these three were also down-regulated. Please refer to Figure S5 for a legend explaining Pathway molecule symbols. For additional information, visit IPA legend help page at http://ingenuity.force.com/ipa/articles/Feature_Description/Legend
(TIF)
Pathway molecule symbols legend from Ingenuity Pathway Analysis. The color scheme used to indicate up- and down-regulation is different from that used by IPA and the intensity varies according to fold change. For additional information, visit IPA legend help page at http://ingenuity.force.com/ipa/articles/Feature_Description/Legend
(TIF)
53 of the selected canonical pathways studied, with genes up- and down-regulated in the subclinical/asymptomatic dengue are indicated. Linear fold change (asymptomatic versus symptomatic) values are shown in bracket for those genes discussed in the manuscript. The pathways have been grouped according to the broad categories of reported dengue host defence mechanisms. The gene symbols are based on HUGO Gene Nomenclature Committee (HGNC) symbols.
(DOC)
Acknowledgments
We thank the clinicians and the nurses at the University Malaya Medical Centre (UMMC), Ampang Hospital and Klang Tengku Ampuan Rahimah Hospital, Malaysia who assisted in collecting blood samples, and the patients and families for their participation in this study.
Funding Statement
This study was supported by High Impact Research Grant UM-MOHE; UM.C/625/1/HIR/MOHE/H-20001-00-E000053, Science Fund (No. 12-02-03-2049) of the Ministry of Science, Technology and Innovation, Malaysia, and UM Postgraduate Research Fund (PV039/2012A). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Bhatt S, Gething PW, Brady OJ, Messina JP, Farlow AW, et al. (2013) The global distribution and burden of dengue. Nature [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Gubler DJ, Rosen L (1976) Variation among geographic strains of Aedes albopictus in susceptibility to infection with dengue viruses. The American journal of tropical medicine and hygiene 25: 318. [DOI] [PubMed] [Google Scholar]
- 3. Platt KB, Linthicum KJ, Myint K, Innis BL, Lerdthusnee K, et al. (1997) Impact of dengue virus infection on feeding behavior of Aedes aegypti. The American journal of tropical medicine and hygiene 57: 119–125. [DOI] [PubMed] [Google Scholar]
- 4. Putnam JL, Scott TW (1995) Blood-feeding behavior of dengue-2 virus-infected Aedes aegypti. The American journal of tropical medicine and hygiene 52: 225–227. [DOI] [PubMed] [Google Scholar]
- 5.Dengue W (2009) Guidelines for diagnosis, treatment, prevention and control. Geneva: World Health Organization. [PubMed] [Google Scholar]
- 6. Siler JF, Hall MW, Hitchens AP (1925) REsults obtained in the transmission of dengue fever. Journal of the American Medical Association 84: 1163–1172. [Google Scholar]
- 7. Endy TP, Anderson KB, Nisalak A, Yoon I-K, Green S, et al. (2011) Determinants of inapparent and symptomatic dengue infection in a prospective study of primary school children in Kamphaeng Phet, Thailand. PLoS neglected tropical diseases 5: e975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Mathew A, Rothman AL (2008) Understanding the contribution of cellular immunity to dengue disease pathogenesis. Immunological reviews 225: 300–313. [DOI] [PubMed] [Google Scholar]
- 9. Gubler DJ, Reed D, Rosen L, Hitchcock J Jr (1978) Epidemiologic, clinical, and virologic observations on dengue in the Kingdom of Tonga. The American journal of tropical medicine and hygiene 27: 581–589. [DOI] [PubMed] [Google Scholar]
- 10. Bravo J, Guzman M, Kouri G (1987) Why dengue haemorrhagic fever in Cuba? I. Individual risk factors for dengue haemorrhagic fever/dengue shock syndrome (DHF/DSS). Transactions of the Royal Society of Tropical Medicine and Hygiene 81: 816–820. [DOI] [PubMed] [Google Scholar]
- 11. Green S, Vaughn DW, Kalayanarooj S, Nimmannitya S, Suntayakorn S, et al. (1999) Early immune activation in acute dengue illness is related to development of plasma leakage and disease severity. Journal of Infectious Diseases 179: 755–762. [DOI] [PubMed] [Google Scholar]
- 12. Vaughn DW, Green S, Kalayanarooj S, Innis BL, Nimmannitya S, et al. (2000) Dengue viremia titer, antibody response pattern, and virus serotype correlate with disease severity. Journal of Infectious Diseases 181: 2–9. [DOI] [PubMed] [Google Scholar]
- 13. Morens DM, Halstead S (1990) Measurement of antibody-dependent infection enhancement of four dengue virus serotypes by monoclonal and polyclonal antibodies. Journal of general virology 71: 2909–2914. [DOI] [PubMed] [Google Scholar]
- 14. Lei H-Y, Yeh T-M, Liu H-S, Lin Y-S, Chen S-H, et al. (2001) Immunopathogenesis of dengue virus infection. Journal of biomedical science 8: 377–388. [DOI] [PubMed] [Google Scholar]
- 15. Diamond MS (2003) Evasion of innate and adaptive immunity by flaviviruses. Immunology and cell biology 81: 196–206. [DOI] [PubMed] [Google Scholar]
- 17. Shresta S (2012) Role of complement in dengue virus infection: protection or pathogenesis? mBio 3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Tisoncik JR, Korth MJ, Simmons CP, Farrar J, Martin TR, et al. (2012) Into the eye of the cytokine storm. Microbiology and Molecular Biology Reviews 76: 16–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Fink J, Gu F, Vasudevan SG (2006) Role of T cells, cytokines and antibody in dengue fever and dengue haemorrhagic fever. Reviews in medical virology 16: 263–275. [DOI] [PubMed] [Google Scholar]
- 20. Luplertlop N, Luplerdlop N, Missé D, Bray D, Deleuze V, et al. (2006) Dengue-virus-infected dendritic cells trigger vascular leakage through metalloproteinase overproduction. EMBO reports 7: 1176–1181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Chaturvedi U, Agarwal R, Elbishbishi E, Mustafa A (2000) Cytokine cascade in dengue hemorrhagic fever: implications for pathogenesis. FEMS Immunology & Medical Microbiology 28: 183–188. [DOI] [PubMed] [Google Scholar]
- 22.Kurane I, Ennis FA. Cytokines in dengue virus infections: role of cytokines in the pathogenesis of dengue hemorrhagic fever; 1994. Elsevier. pp. 443–448. [Google Scholar]
- 23. Harris R, Ukaejiofo E (1969) Rapid preparation of lymphocytes for tissue-typing. The Lancet 294: 327. [DOI] [PubMed] [Google Scholar]
- 24. Lam S, Devi S, Pang T (1987) Detection of specific IgM in dengue infection. Southeast Asian journal of tropical medicine and public health 18: 532–538. [PubMed] [Google Scholar]
- 25. Clarke DH, Casals J (1958) Techniques for hemagglutination and hemagglutination-inhibition with arthropod-borne viruses. The American Journal of Tropical Medicine and Hygiene 7: 561–573. [DOI] [PubMed] [Google Scholar]
- 26. Kong YY, Thay CH, Tin TC, Devi S (2006) Rapid detection, serotyping and quantitation of dengue viruses by TaqMan real-time one-step RT-PCR. Journal of virological methods 138: 123–130. [DOI] [PubMed] [Google Scholar]
- 27. Russell PK, Nisalak A, Sukhavachana P, Vivona S (1967) A plaque reduction test for dengue virus neutralizing antibodies. The Journal of Immunology 99: 285–290. [PubMed] [Google Scholar]
- 28. Vanja L, Laurie HG (2011) T-bet in disease. Nature Immunology 12: 597–606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Thomas SJ, Hombach J, Barrett A (2009) Scientific consultation on cell mediated immunity (CMI) in dengue and dengue vaccine development. Vaccine 27: 355–368. [DOI] [PubMed] [Google Scholar]
- 30. Kyle JL, Balsitis SJ, Zhang L, Beatty PR, Harris E (2008) Antibodies play a greater role than immune cells in heterologous protection against secondary dengue virus infection in a mouse model. Virology 380: 296–303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. García G, Sierra B, Pérez AB, Aguirre E, Rosado I, et al. (2010) Asymptomatic dengue infection in a Cuban population confirms the protective role of the RR variant of the FcγRIIa polymorphism. The American journal of tropical medicine and hygiene 82: 1153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Medzhitov R (2008) Origin and physiological roles of inflammation. Nature 454: 428–435. [DOI] [PubMed] [Google Scholar]
- 33. Nasirudeen A, Wong HH, Thien P, Xu S, Lam K-P, et al. (2011) RIG-I, MDA5 and TLR3 synergistically play an important role in restriction of dengue virus infection. PLoS neglected tropical diseases 5: e926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Matsumoto M, Seya T (2008) TLR3: interferon induction by double-stranded RNA including poly (I: C). Advanced drug delivery reviews 60: 805–812. [DOI] [PubMed] [Google Scholar]
- 35. Liang Z, Wu S, Li Y, He L, Wu M, et al. (2011) Activation of Toll-like receptor 3 impairs the dengue virus serotype 2 replication through induction of IFN-β in cultured hepatoma cells. PloS one 6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Tsai Y-T, Chang S-Y, Lee C-N, Kao C-L (2009) Human TLR3 recognizes dengue virus and modulates viral replication in vitro. Cellular microbiology 11: 604–615. [DOI] [PubMed] [Google Scholar]
- 37. Ubol S, Masrinoul P, Chaijaruwanich J, Kalayanarooj S, Charoensirisuthikul T, et al. (2008) Differences in global gene expression in peripheral blood mononuclear cells indicate a significant role of the innate responses in progression of dengue fever but not dengue hemorrhagic fever. Journal of Infectious Diseases 197: 1459–1467. [DOI] [PubMed] [Google Scholar]
- 38. Gomes ALV, Wee LJ, Khan AM, Gil LH, Marques ET Jr, et al. (2010) Classification of dengue fever patients based on gene expression data using support vector machines. PloS one 5: e11267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Guzman MG, Vazquez S (2010) The complexity of antibody-dependent enhancement of dengue virus infection. Viruses 2: 2649–2662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Van Sorge N, Van Der Pol WL, Van de Winkel J (2003) FcγR polymorphisms: implications for function, disease susceptibility and immunotherapy. Tissue antigens 61: 189–202. [DOI] [PubMed] [Google Scholar]
- 41.Zola H (2007) Leukocyte and stromal cell molecules: The CD markers: Wiley. com.
- 42. Rogers KA, Scinicariello F, Attanasio R (2006) IgG Fc receptor III homologues in nonhuman primate species: genetic characterization and ligand interactions. The Journal of Immunology 177: 3848–3856. [DOI] [PubMed] [Google Scholar]
- 43. Malasit P (1987) Complement and dengue haemorrhagic fever/shock syndrome. The Southeast Asian journal of tropical medicine and public health 18: 316–320. [PubMed] [Google Scholar]
- 44. Avirutnan P, Punyadee N, Noisakran S, Komoltri C, Thiemmeca S, et al. (2006) Vascular leakage in severe dengue virus infections: a potential role for the nonstructural viral protein NS1 and complement. The Journal of infectious diseases 193: 1078–1088. [DOI] [PubMed] [Google Scholar]
- 45. Cardosa M, Porterfield J, Gordon S (1983) Complement receptor mediates enhanced flavivirus replication in macrophages. The Journal of experimental medicine 158: 258–263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Avirutnan P, Fuchs A, Hauhart R, Somnuke P, Youn S, et al. (2010) Antagonism of the complement component C4 by flavivirus nonstructural protein NS1. The Journal of experimental medicine 207: 793–806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Libraty D, Pichyangkul S, Ajariyakhajorn C, Endy T, Ennis F (2001) Human dendritic cells are activated by dengue virus infection: enhancement by gamma interferon and implications for disease pathogenesis. Journal of virology 75: 3501–3508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Lechmann M, Berchtold S, Steinkasserer A, Hauber J (2002) CD83 on dendritic cells: more than just a marker for maturation. Trends in immunology 23: 273–275. [DOI] [PubMed] [Google Scholar]
- 49. Mathew A, Kurane I, Green S, Vaughn D, Kalayanarooj S, et al. (1999) Impaired T cell proliferation in acute dengue infection. Journal of immunology (Baltimore, Md : 1950) 162: 5609–5615. [PubMed] [Google Scholar]
- 50. Ramapraba A, Tan Lian H, Lucy Lum Chai S, Phoay Lay T, Jamuna V, et al. (2007) Cross-reactive T-cell responses to the nonstructural regions of dengue viruses among dengue fever and dengue hemorrhagic fever patients in Malaysia. Clin Vaccine Immunol 14: 969–977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Zhu J, Paul WE (2010) Peripheral CD4+ T-cell differentiation regulated by networks of cytokines and transcription factors. Immunological reviews 238: 247–262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Lazarevic V, Glimcher LH (2011) T-bet in disease. Nature immunology 12: 597–606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Chaturvedi U, Raghupathy R, Pacsa A, Elbishbishi E, Agarwal R, et al. (1999) Shift from a Th1-type response to Th2-type in dengue haemorrhagic fever. Current science 76: 63–69. [Google Scholar]
- 54. Agarwal R, Elbishbishi E, Chaturvedi U, Nagar R, Mustafa A (1999) Profile of transforming growth factor-beta 1 in patients with dengue haemorrhagic fever. International journal of experimental pathology 80: 143–150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Jain A, Pandey N, Garg R, Kumar R (2013) IL-17 level in patients with Dengue virus infection & its association with severity of illness. Journal of clinical immunology 33: 613–618. [DOI] [PubMed] [Google Scholar]
- 56. Raghupathy R, Chaturvedi U, Al-Sayer H, Elbishbishi E, Agarwal R, et al. (1998) Elevated levels of IL-8 in dengue hemorrhagic fever. Journal of medical virology 56: 280–285. [DOI] [PubMed] [Google Scholar]
- 57. Mustafa A, Elbishbishi E, Agarwal R, Chaturvedi U (2001) Elevated levels of interleukin-13 and IL-18 in patients with dengue hemorrhagic fever. FEMS immunology and medical microbiology 30: 229–233. [DOI] [PubMed] [Google Scholar]
- 58. Appanna R, Wang SM, Ponnampalavanar SA, Lum LCS, Devi Sekaran S (2012) Cytokine factors present in dengue patient sera induces alterations of junctional proteins in human endothelial cells. American Journal of Tropical Medicine and Hygiene 87: 936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Mathew A, Rothman A (2008) Understanding the contribution of cellular immunity to dengue disease pathogenesis. Immunological reviews 225: 300–313. [DOI] [PubMed] [Google Scholar]
- 60. Rothman A (2011) Immunity to dengue virus: a tale of original antigenic sin and tropical cytokine storms. Nature reviews Immunology 11: 532–543. [DOI] [PubMed] [Google Scholar]
- 61. Chaturvedi U, Agarwal R, Elbishbishi E, Mustafa A (2000) Cytokine cascade in dengue hemorrhagic fever: implications for pathogenesis. FEMS immunology and medical microbiology 28: 183–188. [DOI] [PubMed] [Google Scholar]
- 62. Chaturvedi U, Elbishbishi E, Agarwal R, Raghupathy R, Nagar R, et al. (1999) Sequential production of cytokines by dengue virus-infected human peripheral blood leukocyte cultures. Journal of medical virology 59: 335–340. [DOI] [PubMed] [Google Scholar]
- 63. Hober D, Poli L, Roblin B, Gestas P, Chungue E, et al. (1993) Serum levels of tumor necrosis factor-alpha (TNF-alpha), interleukin-6 (IL-6), and interleukin-1 beta (IL-1 beta) in dengue-infected patients. The American journal of tropical medicine and hygiene 48: 324–331. [DOI] [PubMed] [Google Scholar]
- 64. Kurane I, Innis B, Nimmannitya S, Nisalak A, Meager A, et al. (1991) Activation of T lymphocytes in dengue virus infections. High levels of soluble interleukin 2 receptor, soluble CD4, soluble CD8, interleukin 2, and interferon-gamma in sera of children with dengue. The Journal of clinical investigation 88: 1473–1480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Rathakrishnan A, Wang SM, Hu Y, Khan AM, Ponnampalavanar S, et al. (2012) Cytokine Expression Profile of Dengue Patients at Different Phases of Illness. PloS one 7: e52215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Chakravarti A, Kumaria R (2006) Circulating levels of tumour necrosis factor-alpha & interferon-gamma in patients with dengue & dengue haemorrhagic fever during an outbreak. Indian Journal of Medical Research 123: 25. [PubMed] [Google Scholar]
- 67. Braga E, Moura P, Pinto L, Ignácio S, Oliveira M, et al. (2001) Detection of circulant tumor necrosis factor-alpha, soluble tumor necrosis factor p75 and interferon-gamma in Brazilian patients with dengue fever and dengue hemorrhagic fever. Memórias do Instituto Oswaldo Cruz 96: 229–232. [DOI] [PubMed] [Google Scholar]
- 68. Fernández-Mestre M, Gendzekhadze K, Rivas-Vetencourt P, Layrisse Z (2004) TNF-alpha-308A allele, a possible severity risk factor of hemorrhagic manifestation in dengue fever patients. Tissue antigens 64: 469–472. [DOI] [PubMed] [Google Scholar]
- 69. Royall J, Berkow R, Beckman J, Cunningham M, Matalon S, et al. (1989) Tumor necrosis factor and interleukin 1 alpha increase vascular endothelial permeability. The American journal of physiology 257: 410. [DOI] [PubMed] [Google Scholar]
- 70. Raghupathy R, Chaturvedi U, Al-Sayer H, Elbishbishi E, Agarwal R, et al. (1998) Elevated levels of IL-8 in dengue hemorrhagic fever. Journal of medical virology 56: 280–285. [DOI] [PubMed] [Google Scholar]
- 71. Juffrie M, van Der Meer G, Hack C, Haasnoot K, Sutaryo, et al. (2000) Inflammatory mediators in dengue virus infection in children: interleukin-8 and its relationship to neutrophil degranulation. Infection and immunity 68: 702–707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Talavera D, Castillo A, Dominguez M, Gutierrez A, Meza I (2004) IL8 release, tight junction and cytoskeleton dynamic reorganization conducive to permeability increase are induced by dengue virus infection of microvascular endothelial monolayers. The Journal of general virology 85: 1801–1813. [DOI] [PubMed] [Google Scholar]
- 73. Chau T, Quyen N, Thuy T, Tuan N, Hoang D, et al. (2008) Dengue in Vietnamese infants–results of infection-enhancement assays correlate with age-related disease epidemiology, and cellular immune responses correlate with disease severity. The Journal of infectious diseases 198: 516–524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. de-Oliveira-Pinto L, Marinho C, Povoa T, de Azeredo E, de Souza L, et al. (2012) Regulation of inflammatory chemokine receptors on blood T cells associated to the circulating versus liver chemokines in dengue fever. PloS one 7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75. Bigger C, Guerra B, Brasky K, Hubbard G, Beard M, et al. (2004) Intrahepatic gene expression during chronic hepatitis C virus infection in chimpanzees. Journal of virology 78: 13779–13792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. Wright H, Alex P, Nguyen T, Bader T, Gurakar A, et al. (2005) Multiplex cytokine profiling of initial therapeutic response in patients with chronic hepatitis C virus infection. Digestive diseases and sciences 50: 1793–1803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77. Bozza F, Cruz O, Zagne S, Azeredo E, Nogueira R, et al. (2008) Multiplex cytokine profile from dengue patients: MIP-1beta and IFN-gamma as predictive factors for severity. BMC infectious diseases 8: 86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78. Perez AB, Sierra B, Garcia G, Aguirre E, Babel N, et al. (2010) Tumor necrosis factor–alpha, transforming growth factor– 1, and interleukin-10 gene polymorphisms: implication in protection or susceptibility to dengue hemorrhagic fever. Human immunology 71: 1135–1140. [DOI] [PubMed] [Google Scholar]
- 79. Laur F, Murgue B, Deparis X, Roche C, Cassar O, et al. (1998) Plasma levels of tumour necrosis factor alpha and transforming growth factor beta-1 in children with dengue 2 virus infection in French Polynesia. Transactions of the Royal Society of Tropical Medicine and Hygiene 92: 654–656. [DOI] [PubMed] [Google Scholar]
- 80. Luplertlop N, Missé D (2008) MMP cellular responses to dengue virus infection-induced vascular leakage. Jpn J Infect Dis 61: 298–301. [PubMed] [Google Scholar]
- 81. Nguyen T, Lei H-Y, Nguyen T, Lin Y-S, Huang K-J, et al. (2004) Dengue hemorrhagic fever in infants: a study of clinical and cytokine profiles. The Journal of infectious diseases 189: 221–232. [DOI] [PubMed] [Google Scholar]
- 82. Tseng C-S, Lo H-W, Teng H-C, Lo W-C, Ker C-G (2005) Elevated levels of plasma VEGF in patients with dengue hemorrhagic fever. FEMS immunology and medical microbiology 43: 99–102. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Innate immune response mega canonical pathway created using Ingenuity Pathway Analysis. TLR complex includes TLR6 (down-regulated in asymptomatic individuals), TLR3 (no change in expression between the two groups) and TLR7 (no change), among others. Please refer to Figure S5 for a legend explaining Pathway molecule symbols. For additional information, visit IPA legend help page at http://ingenuity.force.com/ipa/articles/Feature_Description/Legend
(TIF)
Adaptive immune response mega canonical pathway created using Ingenuity Pathway Analysis. Please refer to Figure S5 for a legend explaining Pathway molecule symbols. For additional information, visit IPA legend help page at http://ingenuity.force.com/ipa/articles/Feature_Description/Legend
(TIF)
Role of cytokines in mediating communication between immune cell canonical pathways from Ingenuity Pathway Analysis. Please refer to Figure S5 for a legend explaining Pathway molecule symbols. For additional information, visit IPA legend help page at http://ingenuity.force.com/ipa/articles/Feature_Description/Legend
(TIF)
Inhibition of matrix metalloproteases canonical pathway from Ingenuity Pathway Analysis. MT-MMP complex includes MMP15, MMP16, and MMP24, among others, and these three were down-regulated in asymptomatic individuals. Extracellular MMP complex includes MMP8, MMP10, and MMP12, among others, and these three were also down-regulated. Please refer to Figure S5 for a legend explaining Pathway molecule symbols. For additional information, visit IPA legend help page at http://ingenuity.force.com/ipa/articles/Feature_Description/Legend
(TIF)
Pathway molecule symbols legend from Ingenuity Pathway Analysis. The color scheme used to indicate up- and down-regulation is different from that used by IPA and the intensity varies according to fold change. For additional information, visit IPA legend help page at http://ingenuity.force.com/ipa/articles/Feature_Description/Legend
(TIF)
53 of the selected canonical pathways studied, with genes up- and down-regulated in the subclinical/asymptomatic dengue are indicated. Linear fold change (asymptomatic versus symptomatic) values are shown in bracket for those genes discussed in the manuscript. The pathways have been grouped according to the broad categories of reported dengue host defence mechanisms. The gene symbols are based on HUGO Gene Nomenclature Committee (HGNC) symbols.
(DOC)