

Abstract
Background
Nitric oxide (NO) is produced as part of the host immune response to bacterial infections, including urinary tract infections. The enzyme flavohemoglobin, coded by the hmp gene, is involved in protecting bacterial cells from the toxic effects of NO and represents a potentially interesting target for development of novel treatment concepts against resistant uropathogenic bacteria. The aim of the present study was to investigate if the in vitro antibacterial effects of NO can be enhanced by pharmacological modulation of the enzyme flavohemoglobin.
Results
Four clinical isolates of multidrug-resistant extended-spectrum β-lactamase (ESBL)-producing uropathogenic E. coli were included in the study. It was shown that the NO-donor substance DETA/NO, but not inactivated DETA/NO, caused an initial growth inhibition with regrowth noted after 8 h of exposure. An hmp-deficient strain showed a prolonged growth inhibition in response to DETA/NO compared to the wild type. The imidazole antibiotic miconazole, that has been shown to inhibit bacterial flavohemoglobin activity, prolonged the DETA/NO-evoked growth inhibition. When miconazole was combined with polymyxin B nonapeptide (PMBN), in order to increase the bacterial wall permeability, DETA/NO caused a prolonged bacteriostatic response that lasted for up to 24 h.
Conclusion
An NO-donor in combination with miconazole and PMBN showed enhanced antimicrobial effects and proved effective against multidrug-resistant ESBL-producing uropathogenic E. coli.
Keywords: Uropathogenic E. coli, Extended-spectrum β-lactamase, Nitric oxide, Polymyxin B nonapeptide
Background
Nitric oxide (NO) is formed from an inducible form of nitric oxide synthase (iNOS) as part of the host antimicrobial defence program [1]. Increased iNOS activity has been documented in the bladder and urine in patients suffering from urinary tract infection (UTI) [2,3]. The antimicrobial effects of NO are complex but NO and its autoxidation products interact with a wide variety of targets including iron-sulfur thiols, tyrosine residues, lipids and DNA bases [1,4]. NO inhibits growth of a wide variety of gram-negative and gram-positive bacterial species [1]. Growth inhibitory effects on common urinary pathogens have been reported [5], but uropathogenic E. coli (UPEC) are less sensitive to NO than non-pathogenic strains of E. coli[6,7].
To protect against the damage caused by nitrosative stress, bacteria possess multiple defence mechanisms involving scavengers, detoxifying enzymes and evasion strategies. Flavohemoglobin, encoded by the hmp gene [8], is a protein that has been shown to have NO dioxygenase activity [8] and be involved in protecting bacterial cells from nitrosative stress [9,10]. Flavohemoglobin utilizes O2 for NO detoxification and oxidize NO to harmless nitrate, which protect the bacteria from the toxic effect of NO. The significance of flavohemoglobin in NO-protection has been shown using hmp-deficient mutants that are more sensitive to NO and nitrosative stress [8,11]. We have previously shown that UPEC increase flavohemoglobin expression after exposure to NO and that an hmp-deficient UPEC strain is more sensitive to NO than a wild type strain [6,12]. The importance of NO-protection and flavohemoglobin for UPEC colonization has been addressed in previous studies. UPEC that had been pre-conditioned to nitrosative stress showed facilitated colonization of the mouse urinary tract [7] while an impaired colonization was observed following flavohemoglobin gene depletion [12]. Moreover, elevated hmp expression was found in UPEC isolates isolated from patients with UTI, suggesting that UPEC isolates face host-derived nitrosative stress during human UTI and activate the NO-detoxifying enzyme flavohemoglobin [12].
Pharmacological inhibition of flavohemoglobin might represent a new strategy to combat human infections, including UTI [10]. X-ray structural information has revealed that the flavohemoglobin protein possess large heme pockets capable of sequestering imidazole antibiotics [13]. Antifungal azoles, like miconazole, are able to inhibit the activity of microbial flavohemoglobin, including the NO dioxygenase activity of E. coli[14]. Furthermore, the binding of miconazole to the heme moiety of flavohemoglobin has been demonstrated to increase the intracellular oxidative stress and enhanced the antimicrobial activity against Staphylococcus aureus[15].
During the last decade an increasing prevalence of extended-spectrum β-lactamase (ESBL)-producing E. coli has been detected worldwide. Plasmid-mediated β-lactamase enzymes inactivate β-lactam antibiotics, which results in ineffective compounds and therapy failure. The majority of ESBL-producing bacteria are isolated from urine samples [16,17] and the prevalence of uropathogenic ESBL-producing isolates have increased in community-acquired UTIs the last decade [18]. The CTX-M-type β-lactamases are the dominant ESBLs and the CTX-M family is classified into five major groups based on similarities in their amino acid sequences [19]. In addition to resistance to most β-lactam antibiotics, multidrug resistance (i.e. resistance ≥ 3 different antimicrobial groups) is common [19]. Dissemination of multidrug-resistant ESBL-producing E. coli may in the future change uncomplicated, treatable urinary tract infections into life threatening infections and new therapeutic options are urgent. Microbial defence enzymes that enable the bacteria to resist host-derived factors have emerged as attractive targets for drug development [20]. Inhibition of factors involved in NO-defence may find applications as antimicrobial therapy by disable the bacterial resistance mechanisms and enhance the toxicity of host-derived or exogenously administered NO.
The aim of the present study was to investigate the antibacterial effects of NO in multidrug-resistant ESBL-producing isolates with special focus on inhibition of the NO-consuming enzyme flavohemoglobin.
Methods
Bacteria
Four ESBL-producing E. coli isolates, recovered from urine from standard patient care individuals with indwelling urinary catheters and symptoms of UTI, were obtained from the Department of Microbiology, Örebro University hospital, Sweden. The identity of the patients was anonymized and after that further analysis of the bacterial strains was performed. Antimicrobial susceptibility testing was performed as recommended by the Swedish Reference Group for Antibiotics (http://www.folkhalsomyndigheten.se/raf). The blaCTX-M gene was detected using real time PCR and nucleotide sequencing as previously described [21]. The CTX-M types and the antibiotic susceptibility of the different ESBL-producing E. coli are shown in Table 1.
Table 1.
Summary of the results from the antibiotic susceptibility testing and the molecular characterization of the producing CTX-M-types
| Isolate | CTX-M subgroup | CTX-M type | Antibiotic resistance |
|---|---|---|---|
| ESBL 1 |
CTX-M-9 |
CTX-M-24 |
CTX, CAZ, CTB, TMP, CIP, MEL, GEN |
| ESBL 6 |
CTX-M-1 |
CTX-M-15 |
CTX, CAZ, CTB, TMP, CIP |
| ESBL 7 |
CTX-M-1 |
CTX-M-15 |
CTX, CAZ, TMP, CIP, MEL |
| ESBL 9 | CTX-M-9 | CTX-M-14 | CTX, CAZ, TMP, CIP, MEL |
Abbreviations: CTX cefotaxime, CAZ ceftazidime, CTB ceftibuten, TMP trimethoprim, CIP ciprofloxacin, MEL mecillinam, GEN gentamicin.
UPEC strain J96, a pyelonephritis isolate, and an hmp-deficient mutant of J96 (J96Δhmp) were also used in the study. Deletion mutation of hmp was constructed using homologous recombination and the FLP recombinase as previously described [12]. There was no difference in the growth ability (measurement of OD600 for 24 hours) between the mutant strain (J96Δhmp) and the wild type strain (J96wt) [12]. Bacteria were maintained on tryptic soy agar (TSA) (Becton Dickinson, Le Pont Claix, France).
Antimicrobial agents
All antimicrobial solutions were prepared freshly immediately before use. DETA/NO belongs to the family of diazeniumdiolates that consists of a complex of NO bound to a polyamine parent compound that spontaneously decompose in a pH-dependent, first-order process [22]. Stock solutions of DETA/NO (DETA/NONOate, Alexis Biochemicals, Lausen, Schweiz) were prepared in PBS (Invitrogen; Grand Island N.Y., USA), cefotaxime and polymyxin B nonapeptid (PMBN; Sigma-Aldrich, St. Louis, USA) were prepared in sterile water. Nitrofurantoin (Sigma-Aldrich, St. Louis, USA) was prepared in DMSO (Merck, Hohenbrunn, Germany) and miconazole (Sigma-Aldrich, St. Louis, USA) was dissolved in 25% ethanol (70%) and 75% polyetylenglykol 400 while heating for 1 h at 60°C.
Determination of MIC
MIC (minimum inhibitory concentration) was determined by the broth dilution test. The test substances were inoculated with a bacterial suspension (~ 106 CFU/ml) in Luria-Bertani (LB) broth (Lennox, Becton Dickinson, Franklin Lakes, USA) for 8–24 h at 37°C. All MIC tests were repeated in two isolates and at least twice. The MIC value for cefotaxime was determined in a cephalosporin-sensitive uropathogenic E. coli isolate. The MIC value for DETA/NO was determined after 8 h since bacterial re-growth was seen in all tubes after 24 h. A working concentration of 4 × MIC was used in all experiments.
Evaluation of bacterial viability
An overnight culture grown in LB broth, at 37°C on shake at 200 rpm, were diluted 1/1000 to give a bacterial count of approximately 106 CFU/ml in the tube. Time-zero samples (starting inoculums) were taken and the number of viable colonies determined as described below. Bacterial cultures were combined with antibiotics or relevant vehicle and incubated in darkness at 37°C. Samples were taken at different times after addition of antibiotics or vehicle (2, 4, 8 and 24 h) depending on the experimental protocol. The samples were diluted in PBS and at least three serial dilutions were plated on TSA-plates. Following overnight culture at 37°C, bacterial CFU/ml was determined by using mean from two dilutions. Growth was calculated as the numbers of CFU/ml in treated cultures or controls divided by the number of CFU/ml formed upon the plating of the initial starting inoculums and expressed as log CFU/ml.
Experimental protocol
1) The bacterial viability of DETA/NO (4 × MIC, 4 mM), cefotaxime (4 × MIC, 0.26 μg/ml) and nitrofurantoin (4 × MIC, 32 μg/ml) was compared in all four ESBL-producing isolates after 8 h of treatment. Two isolates (ESBL 1 and ESBL 7) were further analysed at 2 and 4 h of treatment. 2) To confirm that the effect of DETA/NO was caused by NO and not the parent compound, a tube with DETA/NO in solution was inactivated at 37°C for 96 h followed by exposure in open air for 20 h. 3) To examine the involvement of the NO-consuming enzyme flavohemoglobin, an hmp-deficient UPEC strain (J96Δhmp) and the wild type J96 strain were exposed to DETA/NO for 4, 8 and 24 h and the viability evaluated. 4) The effect of the imidazole antibiotic miconazole (50 μM) in combination with DETA/NO was examined in ESBL isolate 7 at 4, 8 and 24 h. 5) PMBN (5 μg/ml), a compound that increases cell membrane permeability [23], was examined in combination with miconazole and DETA/NO and the viability evaluated in all four ESBL isolates and in J96Δhmp.
Statistical analysis
Data are expressed as mean ± SEM. Differences between groups were assessed by the unpaired two-tailed Student’s t-test. Results were considered statistically significant at p < 0.05. n = number of independent biological replicates.
Results
Characteristics of the isolates
Antibiotic susceptibility testing revealed that the four isolates were resistant to cefotaxime, ceftazidime, trimethoprim and ciprofloxacin. In addition, resistance to ceftibuten, mecillinam and gentamicin was detected in some isolates (Table 1). Nucleotide sequencing revealed that the included isolates were represented by the CTX-M types CTX-M-15, CTX-M-14 and CTX-M-24 (Table 1).
The antibacterial effect of DETA/NO
The antibacterial effect of the NO-donor DETA/NO on the ESBL-producing isolates was compared with the effect of the established antibiotics cefotaxime and nitrofurantoin after exposure for 8 h. Untreated isolates (controls) showed a growth of 2–3 log units during the 8 h period (Figure 1). As predicted, based on the antibiotic susceptibility tests, all four ESBL-producing isolates were resistant to the β-lactam antibiotic cefotaxime (Figure 1A), but sensitive to nitrofurantoin (Figure 1B). The growth response in DETA/NO-treated isolates was reduced by approximately 1–1.5 log units compared to controls (Figure 1C). The ESBL isolates 1 and 7, representing the most and the least DETA/NO-sensitive isolates were further evaluated by time-course studies after exposure to DETA/NO, cefotaxime and nitrofurantoin for 2, 4, and 8 h (Figure 2). In both isolates, DETA/NO demonstrated a significant growth inhibition compared to controls after 2 and 4 h of exposure (Figure 2). However, after 8 h the growth inhibitory effect was less pronounced and only significant in ESBL isolate 7 (Figure 2). A second dose of DETA/NO, administered after 4 h, did not prolong the growth inhibition at 8 h (data not shown). The effects of nitrofurantoin increased over time and, at least in ESBL isolate 7, the effect was bactericidal (decrease in viability by > 103 log units) after 8 h. Both ESBL isolates were resistant to cefotaxime at all time points tested (Figure 2).
Figure 1.

A comparison of the antibacterial effect of cefotaxim, nitrofurantoin and DETA/NO. The effect of cefotaxim (A), nitrofurantoin (B) and DETA/NO (C) on bacterial viability studied in four different ESBL-producing isolates (ESBL 1, ESBL 6, ESBL 7, ESBL 9) after exposure for 8 h. Growth is shown as log CFU/ml of treated bacteria or controls (untreated) and compared to the initial starting inoculum. Representative data from one experiment is shown and similar results were obtained in at least two independent experiments for each isolate.
Figure 2.

Time-course studies of the antibacterial effect of cefotaxim, nitrofurantoin and DETA/NO. Time-course study of cefotaxim, nitrofurantoin and DETA/NO on bacterial viability studied in (A) ESBL-producing isolate 1 and (B) ESBL-producing isolate 7 after exposure for 2, 4 and 8 h. Growth is shown as log CFU/ml of treated bacteria or controls (untreated) and compared to the initial starting inoculum. Data are expressed as mean ± SEM of three independent experiments. *p < 0.05, **p < 0.01, ***p < 0.001 vs control.
DETA/NO belongs to the family of diazeniumdiolates and is a complex of NO bound to a polyamine parent compound. An inactivated DETA/NO solution (prolonged heating and open air exposure) was found to lack antibacterial effects at 2, 4 and 8 h (data not shown), verifying that the antibacterial effect of DETA/NO is caused by NO.
Inhibition of flavohemoglobin by gene deletion
To examine the role of flavohemoglobin for the antibacterial effect evoked by DETA/NO an hmp-deficient UPEC strain (J96Δhmp) was used. The growth response of untreated J96Δhmp and the wild type strain did not differ during the experimental period of 24 h (Figure 3). However, the J96Δhmp strain showed a more prolonged growth inhibition in response to DETA/NO compared to wild type J96 (p < 0.001 at 8 h; Figure 3). The growth of the hmp-deficient strain recovered after 24 h (Figure 3).
Figure 3.
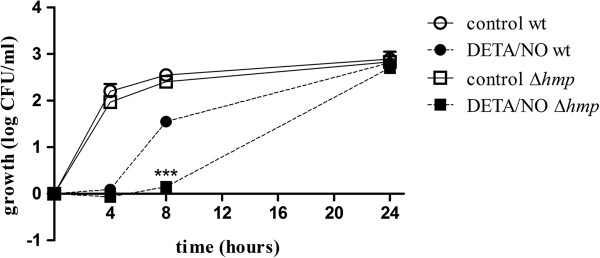
The antibacterial effect of DETA/NO in a mutant strain lacking the flavohemoglobin gene (hmp). The effect of DETA/NO on bacterial viability in an hmp-deficient UPEC strain (Δhmp) and a wild type (wt) UPEC strain after exposure for 4, 8 and 24 h. Growth is shown as log CFU/ml of treated bacteria or controls (untreated) and compared to the initial starting inoculum. Data are expressed as mean ± SEM of three independent experiments. ***p < 0.001 DETA/NO wt vs DETA/NO Δhmp.
Pharmacological modulation of the antibacterial effect of DETA/NO
The imidazole antimicrobial agent miconazole was used to examine whether pharmacological inhibition of flavohemoglobin affects the antibacterial effects of DETA/NO. Miconazole (50 μM) per se had small effects on the growth response when compared to controls (Figure 4A). When DETA/NO and miconazole were combined, the growth inhibition at 8 h was significantly more pronounced (p < 0.001) for the combination treatment than for DETA/NO alone (Figure 4B). However, after 24 h the differences in growth between the treatments were small and overall the viability was not significantly different from the untreated controls (Figure 4B).
Figure 4.

The antibacterial effect of DETA/NO in combination with miconazole and polymyxin B nonapeptide. The effect of DETA/NO on bacterial viability when combined with miconazole and polymyxin B nonapeptide (PMBN) in ESBL-producing isolate 7 after exposure for 4, 8 and 24 h. Growth is shown as log CFU/ml of treated bacteria or controls (untreated) and compared to the initial starting inoculum. (A) The effects of miconazole and PMBN per se are shown. *p < 0.05 control vs miconazole (B) The effects of DETA/NO alone and in combination with miconazole are shown. ***p < 0.001 DETA/NO vs DETA/NO + miconazole (C) The effects of DETA/NO alone and DETA/NO in combination with miconazole and PMBN are shown. **p <0.01, ***p < 0.001 DETA/NO vs DETA/NO + miconazole + PMBN. All data are expressed as mean ± SEM of three-four independent experiments.
Hydrophobic antibiotics, like miconazole, have poor cell membrane permeability in gram-negative bacteria. Polymyxin B antibiotics, like polymyxin B nonapeptide (PMBN), can be used to increase the bacterial cell wall permeability of E. coli[23]. Time-course studies showed that PMBN (5 μg/ml) per se had no effect on the bacterial viability at 4, 8 or 24 h (Figure 4A). In MIC studies, PMBN had no effect on bacterial growth at the highest concentration tested (64 μg/ml). Miconazole and PMBN were used in concentrations that per se had no or minor effect on bacterial viability (Figure 4A). When DETA/NO was combined with miconazole and PMBN a marked and prolonged bacteriostasis for up to 24 h (p < 0.001) was noted in ESBL isolate 7 (Figure 4C). A prolonged bacteriostasis after treatment of DETA/NO in combination with miconazole and PMBN was also noted in ESBL isolates 6 (p < 0.01) and 9 (p < 0.01), while the effect was less pronounced in ESBL isolate 1 (p < 0.05; Figure 5A). PMBN and miconazole in combination slightly reduced the bacterial growth (Figure 5A). DETA/NO and PMBN in combination showed full recovery of growth after 24 h (data not shown).
Figure 5.

The antibacterial effect of DETA/NO in combination with miconazole and polymyxin B nonapeptide in three additional ESBL-producing isolates and in a mutant strain lacking the flavohemoglobin gene (hmp). (A) The effect of DETA/NO on bacterial viability when combined with miconazole and polymyxin B nonapeptide (PMBN) in three additional ESBL-producing isolates (ESBL 1, ESBL 6, ESBL 9) after exposure for 24 h. *p < 0.05, **p < 0.01, ***p < 0.001 vs control, †p <0.05 DETA/NO + miconazole + PMBN vs PMBN + miconazole. (B) The effect of DETA/NO on bacterial viability when combined with miconazole and polymyxin B nonapeptide (PMBN) in an hmp-deficient UPEC strain (Δhmp) after exposure for 8 h. Growth is shown as log CFU/ml of treated bacteria or controls (untreated) and compared to the initial starting inoculum. Data are expressed as mean ± SEM of three independent experiments.
The effect of miconazole in the hmp-deficient UPEC strain
To investigate whether the effect of miconazole can be explained by inhibition of flavohemoglobin, miconazole was further evaluated in the hmp-deficient UPEC strain (J96Δhmp). This strain does not express the flavohemoglobin protein and should therefore be insensitive to pharmacological inhibition of flavohemoglobin. As previously noted (Figure 3), DETA/NO inhibited the growth of the hmp-deficient UPEC during the 8 h study period. The addition of miconazole did not further enhance the growth inhibitory response evoked by DETA/NO (Figure 5B). The inhibitory effect of miconazole and DETA/NO in combination with PMBN was not significantly different from the effect by DETA/NO alone (Figure 5B).
Discussion
The present study investigated the antibacterial effects of NO in multidrug-resistant ESBL-producing isolates with special focus on inhibition of the NO-consuming enzyme flavohemoglobin. Measuring bacterial sensitivity to gaseous NO is difficult because NO per se is stable for only minutes under physiological conditions [24]. Instead, NO-releasing compounds e.g., DETA/NO can be added to bacterial cultures to evaluate the growth response. In E. coli, NO acts in a bacteriostatic fashion and its targets include respiratory enzymes like cytochromes bo and bd and biosynthesis pathways of branched-chain amino acid [25]. In our study, DETA/NO induced a temporary growth inhibition in ESBL-producing UPEC isolates but after 8 hours of DETA/NO exposure a resumed growth was found. A second dose of DETA/NO, administered after 4 hours, did not prolong the growth inhibition (data not shown), suggesting that stress-response factors and NO-defence mechanisms may have been activated by the first dose. All isolates were resistant to cefotaxime but nitrofurantoin showed a time-dependent bactericidal effect. Nitrofurantoin is an antibiotic used for treatment of uncomplicated UTIs, and is so far effective against many isolates of ESBL-producing E. coli[26].
Upon diffusing into the bacteria, NO may react with Fe-S clusters, undergo autoxidation or be consumed directly through enzymatic detoxification [4]. Under aerobic conditions the vast majority of intracellular NO in E. coli is consumed through flavohemoglobin detoxification [4,9,10]. The flavohemoglobin enzyme is not constitutively expressed and needs to be induced by gene transcription [9,10]. We have previously shown that uropathogenic E. coli increase gene and protein expression of flavohemoglobin after exposure to DETA/NO [6,12]. Thus, induction of the flavohemoglobin enzyme and a fast consumption of NO to submicromolar intracellular NO concentrations may explain the temporary growth inhibition with a subsequent growth recovery in our experiments. Indeed, a mutant strain lacking the flavohemoglobin enzyme (J96Δhmp) showed prolonged inhibition of growth, but after 24 hours this mutant also showed resumed growth. It has previously been verified that the hmp-deficient mutant used in the present study does not express the flavohemoglobin gene or protein when exposed to DETA/NO [12]. In the absence of functional flavohemoglobin NO is predicted to be metabolized mainly through autoxidation, enzymatic reduction by NorV and NrfA and by Fe-S nitrosylation [4]. Inhibition of flavohemoglobin by gene deletion was performed in a non ESBL-producing UPEC strain (J96). Gene deletion in ESBL-producing isolates is hampered by the obvious difficulties to find selection antibiotics in these multidrug-resistant isolates. However, we have confirmed a marked increase in hmp expression in an ESBL-producing isolate by real time RT-PCR when exposed to DETA/NO (data not shown), confirming that hmp is induced by DETA/NO also in ESBL-producing isolates.
Miconazole is known to interfere with synthesis of fungal and bacterial lipid membranes as it restrains the ergosterol synthesis [27], but recent studies also suggest that azole antibiotics target the flavohemoglobin enzyme [14,15]. Miconazole in combination with DETA/NO prolonged the DETA/NO-induced growth inhibition in ESBL-producing UPEC isolates. Notably, the pattern of DETA/NO-evoked growth inhibition achieved by addition of miconazole was similar to the pattern noted after hmp-deletion. Furthermore, the fact that miconazole did not increase the DETA/NO-induced growth inhibition in an hmp-mutant strain, support that inhibition of flavohemoglobin [14] contributes to the antibacterial effect of miconazole in our experiments. Thus, the prolonged growth inhibition evoked by DETA/NO and miconazole in combination may be a result of interactions of miconazole with flavohemoglobin, causing both inhibition of NO dioxygenase activity and oxidative stress following high levels of cytotoxic superoxide production [14,15]. In agreement with our results, intracellular survival studies in activated NO-producing macrophages demonstrated decreased survival of miconazole-treated S. aureus compared to untreated bacteria [15]. DETA/NO and miconazole have a synergistic antifungal effect in Candida species [28], and the present study demonstrates that these two compounds also caused an enhanced antibacterial effect against multidrug-resistant ESBL-producing E. coli isolates.
It is noteworthy that inhibition of flavohemoglobin activity by miconazole is more pronounced in purified enzyme than in intact E. coli[14], in line with the poor membrane permeability of E. coli to imidazole antibiotics [29]. Polymyxin B antibiotics may be used to sensitize the outer membrane of gram-negative bacteria to hydrophobic antibiotics [23]. We used polymyxin B nonapeptide (PMBN), a compound that increases the cell permeability in E. coli without affecting the bacterial viability [23,30], to avoid that polymyxin B mask the antibacterial effects of NO. PMBN per se had no antibacterial effect, while miconazole at the concentration used showed a minor inhibitory effect on UPEC growth. An in vitro synergism of miconazole and polymyxin B has been reported in E. coli, related predominantly to the ability of polymyxin B to increase the penetration of miconazole to the intracellular space [31]. In our experiments, miconazole and PMBN in combination caused a significant inhibition of growth compared to untreated controls. Interestingly, when DETA/NO was added to miconazole and PMBN a prolonged bacteriostatic response that persisted for 24 hours was observed. It is not likely that the underlying mechanism is a more effective inhibition of NO-detoxification by flavohemoglobin since the hmp-mutant showed recovered growth after 24 hours. However, a better access of miconazole to intracellular targets like flavohemoglobin, when combined with PMBN, may cause enhanced antibacterial activity through magnification of intracellular oxidative stress responses [15]. Furthermore, increased formation of toxic peroxynitrite (ONOO−), a potent oxidant formed from NO and superoxide radical [1], could conceivably contribute to the spectrum of potential antibacterial mechanisms of the combination treatment. The bacterial cell membrane is another possible target and miconazole is known to affect the integrity of the lipid membrane [27,32]. Morphological analysis have revealed cell membrane deteriorations with widespread structural deformations as a consequence of NO exposure in E. coli[33], and a synergistic effect of all three substances on the cell membrane is possible. However, the exact mechanisms for the prolonged bacteriostasis evoked by DETA/NO in combination with miconazole and PMBN need to be further studied. Interestingly, impaired adhesion to host renal epithelial cells and broken fimbriae has been reported after NO exposure in E. coli[33,34]. This suggests that the antibacterial effects of NO may be widespread and that NO not only has growth inhibitory effects but also may affect bacterial virulence properties and host activating mechanisms.
The ESBL-producing UPEC isolates used in the present study were obtained from patients with catheter-associated UTI. Indwelling medical devices, including urinary catheters and biofilm formation increase the risk of bacterial infection and result in considerable antimicrobial use. When these infections are caused by multidrug-resistant bacteria commonly used empirical antimicrobial therapy are not effective [35]. Administration of NO directly into the bladder through a silicone balloon catheter represents a local delivery system for NO-based therapy and has been suggested as one strategy to prevent catheter-associated infections [5]. Urinary catheters impregnated with NO have been shown to inhibit both biofilm formation and planktonic E. coli growth [36]. A limitation of the present study is that the number of clinical isolates used is small. However, three out of four isolates responded identical and with a prolonged bacteriostasis to the triple combination. These clinical isolates represented different CTX-M enzymes but it is, however, not possible to draw any conclusions on possible correlations between susceptibility to treatment and the CTX-M enzyme based on this material. Importantly, two of the isolates belonged to the CTX-M-15 ESBL type and the sequence type 131 (data not shown) which represent the dominating worldwide emerging CTX-M type and clone in community-acquired UTIs [37,38].
Colistin, a polymyxin antibiotic, has regained interest for its activity against multidrug-resistant gram negative pathogens, including those harbouring carbapenamases [39]. Thus, the emergency of multi-resistant pathogens encourages rediscovery of older antibiotics with activity against these resistant bacteria and in new combinations. Our data suggest that two existing antibiotics, an azole antifungal and a polymyxin B compound, are able to enhance the antimicrobial effects of exogenously administered NO. In contrast to antibiotics that are specific to one or few bacterial species, the antibacterial effects of NO are broad and both gram-positive and gram-negative pathogens, including antibiotic-resistant isolates show sensitivity [40,41]. Development of resistance mechanisms to NO may be limited by its multiple biochemical targets, but metabolic adaptions to nitrosative stress including induction of flavohemoglobin [10] and lactate dehydrogenase in S. aureus[42] decrease the antimicrobial action of NO. Therefore, as highlighted in the present study, a combination treatment where exogenous NO is combined with an inhibitor of NO-protective mechanisms appears to be an attractive approach to improve the antimicrobial effects of NO.
Conclusion
This work shows that the antibacterial effect of NO against multidrug-resistant ESBL-producing uropathogenic E. coli is improved by combination with miconazole and polymyxin B nonapeptide.
Competing interests
The authors declared that they have no competing interests.
Authors’ contributions
Conception and design of the study: KP. Laboratory work: CSB, AK, MK, AÖ. Data analysis and interpretation: CSB, AK, MK, KP. Manuscript writing: CSB, BS, KP. All authors read and approved the final manuscript.
Contributor Information
Charlotte Sahlberg Bang, Email: charlotte.sahlberg-bang@oru.se.
Annica Kinnunen, Email: ann_ica_@hotmail.com.
Marie Karlsson, Email: Marie.A.Karlsson@apoteket.se.
Anna Önnberg, Email: anna.onnberg@orebroll.se.
Bo Söderquist, Email: bo.soderquist@orebroll.se.
Katarina Persson, Email: katarina.persson@oru.se.
Acknowledgement
This project was supported by the Swedish Council for Working Life and Social Research (FAS), Nyckelfonden at Örebro University Hospital and Örebro University.
References
- Fang FC. Antimicrobial reactive oxygen and nitrogen species: concepts and controversies. Nat Rev Microbiol. 2004;2(10):820–832. doi: 10.1038/nrmicro1004. [DOI] [PubMed] [Google Scholar]
- Smith SD, Wheeler MA, Weiss RM. Nitric oxide synthase: an endogenous source of elevated nitrite in infected urine. Kidney Int. 1994;45(2):586–591. doi: 10.1038/ki.1994.76. [DOI] [PubMed] [Google Scholar]
- Wheeler MA, Smith SD, Garcia-Cardena G, Nathan CF, Weiss RM, Sessa WC. Bacterial infection induces nitric oxide synthase in human neutrophils. J Clin Invest. 1997;99(1):110–116. doi: 10.1172/JCI119121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robinson JL, Brynildsen MP. A kinetic platform to determine the fate of nitric oxide in Escherichia coli. PLoS Comput Biol. 2013;9(5):e1003049. doi: 10.1371/journal.pcbi.1003049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carlsson S, Weitzberg E, Wiklund P, Lundberg JO. Intravesical nitric oxide delivery for prevention of catheter-associated urinary tract infections. Antimicrob Agents Chemother. 2005;49(6):2352–2355. doi: 10.1128/AAC.49.6.2352-2355.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Svensson L, Marklund BI, Poljakovic M, Persson K. Uropathogenic Escherichia coli and tolerance to nitric oxide: the role of flavohemoglobin. J Urol. 2006;175(2):749–753. doi: 10.1016/S0022-5347(05)00144-8. [DOI] [PubMed] [Google Scholar]
- Bower JM, Gordon-Raagas HB, Mulvey MA. Conditioning of uropathogenic Escherichia coli for enhanced colonization of host. Infect Immun. 2009;77(5):2104–2112. doi: 10.1128/IAI.01200-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gardner PR, Gardner AM, Martin LA, Salzman AL. Nitric oxide dioxygenase: an enzymic function for flavohemoglobin. Proc Natl Acad Sci U S A. 1998;95(18):10378–10383. doi: 10.1073/pnas.95.18.10378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poole RK, Hughes MN. New functions for the ancient globin family: bacterial responses to nitric oxide and nitrosative stress. Mol Microbiol. 2000;36(4):775–783. doi: 10.1046/j.1365-2958.2000.01889.x. [DOI] [PubMed] [Google Scholar]
- Forrester MT, Foster MW. Protection from nitrosative stress: a central role for microbial flavohemoglobin. Free Radic Biol Med. 2012;52(9):1620–1633. doi: 10.1016/j.freeradbiomed.2012.01.028. [DOI] [PubMed] [Google Scholar]
- Membrillo-Hernandez J, Coopamah MD, Anjum MF, Stevanin TM, Kelly A, Hughes MN, Poole RK. The flavohemoglobin of Escherichia coli confers resistance to a nitrosating agent, a "Nitric oxide Releaser," and paraquat and is essential for transcriptional responses to oxidative stress. J Biol Chem. 1999;274(2):748–754. doi: 10.1074/jbc.274.2.748. [DOI] [PubMed] [Google Scholar]
- Svensson L, Poljakovic M, Save S, Gilberthorpe N, Schon T, Strid S, Corker H, Poole RK, Persson K. Role of flavohemoglobin in combating nitrosative stress in uropathogenic Escherichia coli–implications for urinary tract infection. Microb Pathog. 2010;49(3):59–66. doi: 10.1016/j.micpath.2010.04.001. [DOI] [PubMed] [Google Scholar]
- Ilari A, Bonamore A, Farina A, Johnson KA, Boffi A. The X-ray structure of ferric Escherichia coli flavohemoglobin reveals an unexpected geometry of the distal heme pocket. J Biol Chem. 2002;277(26):23725–23732. doi: 10.1074/jbc.M202228200. [DOI] [PubMed] [Google Scholar]
- Helmick RA, Fletcher AE, Gardner AM, Gessner CR, Hvitved AN, Gustin MC, Gardner PR. Imidazole antibiotics inhibit the nitric oxide dioxygenase function of microbial flavohemoglobin. Antimicrob Agents Chemother. 2005;49(5):1837–1843. doi: 10.1128/AAC.49.5.1837-1843.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nobre LS, Todorovic S, Tavares AF, Oldfield E, Hildebrandt P, Teixeira M, Saraiva LM. Binding of azole antibiotics to Staphylococcus aureus flavohemoglobin increases intracellular oxidative stress. J Bacteriol. 2010;192(6):1527–1533. doi: 10.1128/JB.01378-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pitout JD, Laupland KB. Extended-spectrum beta-lactamase-producing Enterobacteriaceae: an emerging public-health concern. Lancet Infect Dis. 2008;8(3):159–166. doi: 10.1016/S1473-3099(08)70041-0. [DOI] [PubMed] [Google Scholar]
- Khanfar HS, Bindayna KM, Senok AC, Botta GA. Extended spectrum beta-lactamases (ESBL) in Escherichia coli and Klebsiella pneumoniae: trends in the hospital and community settings. J Infect Dev Ctries. 2009;3(4):295–299. doi: 10.3855/jidc.127. [DOI] [PubMed] [Google Scholar]
- Hoban DJ, Lascols C, Nicolle LE, Badal R, Bouchillon S, Hackel M, Hawser S. Antimicrobial susceptibility of Enterobacteriaceae, including molecular characterization of extended-spectrum beta-lactamase-producing species, in urinary tract isolates from hospitalized patients in North America and Europe: results from the SMART study 2009–2010. Diagn Microbiol Infect Dis. 2012;74(1):62–67. doi: 10.1016/j.diagmicrobio.2012.05.024. [DOI] [PubMed] [Google Scholar]
- Bonnet R. Growing group of extended-spectrum beta-lactamases: the CTX-M enzymes. Antimicrob Agents Chemother. 2004;48(1):1–14. doi: 10.1128/AAC.48.1.1-14.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nathan C. Antibiotics at the crossroads. Nature. 2004;431(7011):899–902. doi: 10.1038/431899a. [DOI] [PubMed] [Google Scholar]
- Onnberg A, Molling P, Zimmermann J, Soderquist B. Molecular and phenotypic characterization of Escherichia coli and Klebsiella pneumoniae producing extended-spectrum beta-lactamases with focus on CTX-M in a low-endemic area in Sweden. APMIS. 2011;119(4–5):287–295. doi: 10.1111/j.1600-0463.2011.02730.x. [DOI] [PubMed] [Google Scholar]
- Keefer LK, Nims RW, Davies KM, Wink DA. "NONOates" (1-substituted diazen-1-ium-1,2-diolates) as nitric oxide donors: convenient nitric oxide dosage forms. Methods Enzymol. 1996;268:281–293. doi: 10.1016/s0076-6879(96)68030-6. [DOI] [PubMed] [Google Scholar]
- Vaara M. Agents that increase the permeability of the outer membrane. Microbiol Rev. 1992;56(3):395–411. doi: 10.1128/mr.56.3.395-411.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pullan ST, Monk CE, Lee L, Poole RK. Microbial responses to nitric oxide and nitrosative stress: growth, "omic," and physiological methods. Methods Enzymol. 2008;437:499–519. doi: 10.1016/S0076-6879(07)37025-0. [DOI] [PubMed] [Google Scholar]
- Hyduke DR, Jarboe LR, Tran LM, Chou KJ, Liao JC. Integrated network analysis identifies nitric oxide response networks and dihydroxyacid dehydratase as a crucial target in Escherichia coli. Proc Natl Acad Sci U S A. 2007;104(20):8484–8489. doi: 10.1073/pnas.0610888104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grandesso S, Sapino B, Mazzucato S, Alessandrini R, Solinas M, Gion M. Study on in-vitro susceptibility of ESBL-positive Escherichia coli isolated from urine specimens. Infez Med. 2010;18(3):162–168. [PubMed] [Google Scholar]
- Vanden Bossche H, Marichal P, Gorrens J, Coene MC, Willemsens G, Bellens D, Roels I, Moereels H, Janssen PA. Biochemical approaches to selective antifungal activity. Focus on azole antifungals. Mycoses. 1989;32(Suppl 1):35–52. doi: 10.1111/j.1439-0507.1989.tb02293.x. [DOI] [PubMed] [Google Scholar]
- McElhaney-Feser GE, Raulli RE, Cihlar RL. Synergy of nitric oxide and azoles against Candida species in vitro. Antimicrob Agents Chemother. 1998;42(9):2342–2346. doi: 10.1128/aac.42.9.2342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cornelissen F, Van den Bossche H. Synergism of the antimicrobial agents miconazole, bacitracin and polymyxin B. Chemotherapy. 1983;29(6):419–427. doi: 10.1159/000238230. [DOI] [PubMed] [Google Scholar]
- Sahalan AZ, Dixon RA. Role of the cell envelope in the antibacterial activities of polymyxin B and polymyxin B nonapeptide against Escherichia coli. Int J Antimicrob Agents. 2008;31(3):224–227. doi: 10.1016/j.ijantimicag.2007.10.005. [DOI] [PubMed] [Google Scholar]
- Pietschmann S, Hoffmann K, Voget M, Pison U. Synergistic effects of miconazole and polymyxin B on microbial pathogens. Vet Res Commun. 2009;33(6):489–505. doi: 10.1007/s11259-008-9194-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thevissen K, Ayscough KR, Aerts AM, Du W, De Brucker K, Meert EM, Ausma J, Borgers M, Cammue BP, Francois IE. Miconazole induces changes in actin cytoskeleton prior to reactive oxygen species induction in yeast. J Biol Chem. 2007;282(30):21592–21597. doi: 10.1074/jbc.M608505200. [DOI] [PubMed] [Google Scholar]
- Deupree SM, Schoenfisch MH. Morphological analysis of the antimicrobial action of nitric oxide on gram-negative pathogens using atomic force microscopy. Acta Biomater. 2009;5(5):1405–1415. doi: 10.1016/j.actbio.2009.01.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Svensson L, Save S, Persson K. The effect of nitric oxide on adherence of P-fimbriated uropathogenic Escherichia coli to human renal epithelial cells. BJU Int. 2010;105(12):1726–1731. doi: 10.1111/j.1464-410X.2009.08986.x. [DOI] [PubMed] [Google Scholar]
- Milan PB, Ivan IM. Catheter-associated and nosocomial urinary tract infections: antibiotic resistance and influence on commonly used antimicrobial therapy. Int Urol Nephrol. 2009;41(3):461–464. doi: 10.1007/s11255-008-9468-y. [DOI] [PubMed] [Google Scholar]
- Regev-Shoshani G, Ko M, Miller C, Av-Gay Y. Slow release of nitric oxide from charged catheters and its effect on biofilm formation by Escherichia coli. Antimicrob Agents Chemother. 2010;54(1):273–279. doi: 10.1128/AAC.00511-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rogers BA, Sidjabat HE, Paterson DL. Escherichia coli O25b-ST131: a pandemic, multiresistant, community-associated strain. J Antimicrob Chemother. 2011;66(1):1–14. doi: 10.1093/jac/dkq415. [DOI] [PubMed] [Google Scholar]
- Karfunkel D, Carmeli Y, Chmelnitsky I, Kotlovsky T, Navon-Venezia S. The emergence and dissemination of CTX-M-producing Escherichia coli sequence type 131 causing community-onset bacteremia in Israel. Eur J Clin Microbiol Infect Dis. 2013;32(4):513–521. doi: 10.1007/s10096-012-1765-9. [DOI] [PubMed] [Google Scholar]
- Yahav D, Farbman L, Leibovici L, Paul M. Colistin: new lessons on an old antibiotic. Clin Microbiol Infect. 2012;18(1):18–29. doi: 10.1111/j.1469-0691.2011.03734.x. [DOI] [PubMed] [Google Scholar]
- Ghaffari A, Miller CC, McMullin B, Ghahary A. Potential application of gaseous nitric oxide as a topical antimicrobial agent. Nitric Oxide. 2006;14(1):21–29. doi: 10.1016/j.niox.2005.08.003. [DOI] [PubMed] [Google Scholar]
- Friedman A, Blecher K, Sanchez D, Tuckman-Vernon C, Gialanella P, Friedman JM, Martinez LR, Nosanchuk JD. Susceptibility of Gram-positive and -negative bacteria to novel nitric oxide-releasing nanoparticle technology. Virulence. 2011;2(3):217–221. doi: 10.4161/viru.2.3.16161. [DOI] [PubMed] [Google Scholar]
- Richardson AR, Libby SJ, Fang FC. A nitric oxide-inducible lactate dehydrogenase enables Staphylococcus aureus to resist innate immunity. Science. 2008;319(5870):1672–1676. doi: 10.1126/science.1155207. [DOI] [PubMed] [Google Scholar]