

Abstract
It has long been presumed that activation of the apoptosis-initiating Death Receptor 5, as well as other structurally homologous members of the TNF-receptor superfamily, relies on ligand-stabilized trimerization of noninteracting receptor monomers. We and others have proposed an alternate model in which the TNF-receptor dimer—sitting at the vertices of a large supramolecular receptor network of ligand-bound receptor trimers—undergoes a closed-to-open transition, propagated through a scissorslike conformational change in a tightly bundled transmembrane (TM) domain dimer. Here we have combined electron paramagnetic resonance spectroscopy and potential-of-mean force calculations on the isolated TM domain of the long isoform of DR5. The experiments and calculations both independently validate that the opening transition is intrinsic to the physical character of the TM domain dimer, with a significant energy barrier separating the open and closed states.
Death receptor 5 (DR5) is a member of the tumor necrosis factor receptor (TNFR) superfamily that mediates apoptosis when bound by its cognate ligand, TNF-related apoptosis-inducing ligand (1). Upregulated in cancer cells, DR5 is among the most actively pursued anticancer targets (2). TNF-related apoptosis-inducing ligand binds to preassembled DR5 trimers at their extracellular domains, causing the formation of oligomeric ligand-receptor networks that are held together by receptor dimers (3). In the long-isoform of DR5, this dimer is crosslinked via ligand-induced disulfide bond formation between two transmembrane (TM) domain α-helices at Cys-209, and is further stabilized by a GxxxG motif one helix-turn downstream (3).
Our recent study of the structurally homologous TNFR1 showed that receptor activation involves a conformational change that propagates from the extracellular domain to the cytosolic domain through a separation (or opening) of the TM domains of the dimer (4). We have therefore hypothesized that the activation of DR5, and indeed all structurally homologous TNF-receptors, involves a scissorslike opening of the TM domain dimer (Fig. 1).
Figure 1.

Activation model of the DR5-L TM dimer. The sequence and positions of the disulfide bond and TOAC spin label (top), along with our previously published model (bottom, left) are shown. We propose an activation model (bottom, right) in which the transmembrane dimer pivots at its disulfide bond to reach an active open conformation.
Using electron paramagnetic resonance (EPR) spectroscopy, a technique that has been used previously to study TM helix architecture and dynamics (5,6), and potential-of-mean force (PMF) calculations (7,8), this study addresses the question of whether the isolated disulfide-linked DR5-L TM domain dimer occupies distinct open and closed states (Fig. 1), and how its dynamic behavior contributes to the free-energy landscape of the opening transition of the full-length receptor.
The DR5-L TM domain was synthesized with TOAC, an amino acid with a nitroxide spin label rigidly fixed to the α-carbon (9), incorporated at position 32 (Fig. 1), with some minor modification to facilitate EPR measurements. Previous work confirmed that this peptide forms disulfide-linked dimers (e.g., via comparison to 2-ME treated sample) and a negligible population of higher-order oligomers (further supported by model fitting of the EPR data below). For peptide work, residues were renumbered such that Thr-204 corresponds to Thr-1, and so on. The cytosolic Cys-29 (which we previously showed does not participate in a disulfide bond in cells) was replaced with serine to prevent the formation of antiparallel disulfide-linked dimers, and Trp-34 was replaced with tyrosine to prevent intrinsic fluorescence in fluorescence studies (not published). Continuous-wave (CW) dipolar EPR (sensitive only to spin-spin distances <25 Å) was used to measure TOAC-TOAC distances within the TM dimers and revealed an ordered Gaussian distribution centered at 16 Å (full width half-maximum (FWHM) = 4 Å), corresponding to a closed state (Fig. 2 A). Double electron-electron resonance (DEER) (sensitive to spin-spin distances from 15 to 60 Å) also detected a short distance consistent with the dipolar EPR data, along with a longer, disordered component (32.9 Å, FWHM = 28 Å) (Fig. 2 B). Together, these measurements indicate the presence of a compact, ordered closed state and a broader, disordered open state. EPR on oriented membranes also indicated two structural states. Global fitting revealed two populations of spin-label tilt angles (orientation of the nitroxide principal axis relative to the membrane normal): a narrow conformation (24°, FWHM = 20°), and a disordered conformation (50°, FWHM = 48°) (Fig. 2 C). This bimodal orientational distribution (Fig. 2 C) is remarkably consistent with the bimodal distance distribution (Fig. 2 B).
Figure 2.
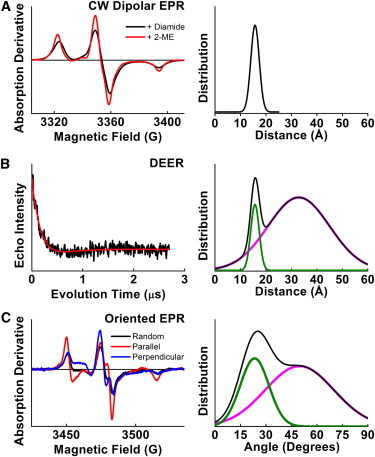
EPR spectra (left) of 32-TOAC-DR5 in lipid, and resulting structural distributions (right). (A) CW dipolar EPR spectra (left) of dimer (1 mM diamide) and monomer (1 mM 2-mercaptoethanol). Best-fit spin-spin distance distribution was a single Gaussian centered at 16 ± 2 Å (right). (B) The DEER waveform (left) of 32-TOAC-DR5 dimer was best fit (right) to a two-Gaussian distribution. The short distance was constrained to agree with the CW data, because DEER has poor sensitivity for distances <20 Å. The long-distance distribution is centered at 32.9 Å and is much broader. (C) CW EPR spectra (left) of 32-TOAC-DR5, with the membrane-normal oriented parallel (red) and perpendicular (blue) to the field. Simultaneous (global) fitting of these spectra reveals narrow and broad components (right). (In panels B and C, the overall distribution is plotted as black, while the closed and open components are plotted as green and magenta, respectively.)
We subsequently conducted a PMF calculation (10) using the DR5-L TM dimer starting configuration developed by our group previously (3), embedded in a DMPC bilayer, with the Leu-32/Leu-32 Cα distance as the reaction coordinate. Three calculations were run from independent starting configurations, each using 50 windows spaced in 0.5° increments, and run for 20 ns at each window (totaling 3 μs). Each of the calculations yielded a similar result, and the averaged free energy curve (Fig. 3 A) agrees remarkably well with our EPR measurements: a narrow distribution at the closed conformation (∼16 Å, Fig. 3 B) separated by an ∼3 kcal/mol energy barrier from a broad distribution of accessible open conformations at ∼27 Å, (Fig. 3 C). Each of the three individual PMF plots can be found in Fig. S1 in the Supporting Material.
Figure 3.

(A) PMF calculation of the DR5 TM domain dimer along the Leu-32/Leu-32 distance reaction coordinate. The PMF calculation reveals a narrow closed state and a broader open state separated by a free energy barrier. Representative snapshots of the (B) closed state and (C) open state.
In the closed state, the helices are tightly packed at the GxxxG interfacial motif and all the way down the juxtaposed helix faces at residues Ala-18, Leu-22, Ala-25, and Val-26. The tight packing is aided by kinking and twisting of the two helices around their common axis, increasing the interacting surface area. In the open conformations, the Ala-18, Leu-22, Ala-25, and Val-26 pairs are dissociated and, interestingly, the GxxxG motif at Gly-10 and Gly-14 remains tightly packed. The open state energy well is only slightly less favorable than the closed state (by ∼2 kcal/mol), and its free energy profile is relatively broad and flat. The increased crossing angle in the open state is facilitated by straightening of the helix kink and is not accommodated by a change in bilayer thickness (see Fig. S3, A and B).
The observed change in helix-helix distance (11 Å between the two minima in the PMF) is extremely close to that observed previously in live-cell FRET studies of a constitutively active form of TNFR1 (∼8 Å change between states using large fluorescence probes at the cytosolic domains) (4). The change observed in the EPR data (17 Å) may be an overestimate because the measurement is made between TOAC spin labels that likely protrude from the two helices, depending on rotational orientation. These results collectively show that activation of these receptors requires a small, but clearly significant conformational opening of the TM domains. One important note is that our EPR experiments recapitulate the equilibrium distribution of the two states despite there being no driving force to traverse the barrier between them (∼3 kcal/mol in the closed-to-open transition and ∼1 kcal/mol in the open-to-closed transition, Fig. 3). We do not interpret the results to mean that the dimer necessarily traverses these barriers at 4°C. Rather, there likely exist multiple reaction paths for dimerization of the abstracted TM domains. Finally, in the context of the full-length receptor, how the ligand induces a conformational change capable of overcoming the closed-to-open barrier remains an important question.
Whether the observed structural transition in the TM domain dimer of the long-isoform of DR5 is a ubiquitous conformational switch that acts over the entire TNFR superfamily remains unknown. Vilar et al. (11) first proposed a similar scissors-model for activation of p75 neurotrophin receptor, which has a cysteine at the center of its TM helix. The short isoform of DR5 lacks a TM domain cysteine, but does form noncovalent dimers in cells, with likely TM domain dimer contacts (3). Among the other closely related and structurally homologous members of the TNFR superfamily, TNFR1 contains a cysteine at the center of the TM domain, but lacks any discernible small residue motifs (e.g., GxxxG). TNFR2 lacks a TM cysteine on the extracellular side, but does have a GxxxG motif positioned similarly to that of DR5. On the other hand, Death Receptor 4, whose functional distinction from DR5 has remained somewhat elusive, lacks both a cysteine and any recognizable small-residue hydrophobic motif.
In summary, we have extended recent findings that point to the TM domain of DR5 as an essential structural component in the conformational change associated with activation. Our findings that the DR5-L TM domain occupies distinct open and closed states, separated by a substantial energy barrier, points the way to further studies across the TNF-receptor superfamily.
Acknowledgments
Simulations were carried out using resources at the University of Minnesota Supercomputing Institute. Electron paramagnetic resonance experiments were performed at the Biophysical Spectroscopy Center, University of Minnesota. D.D.T. was supported by the National Institutes of Health grant No. GM27906.
Supporting Material
References and Footnotes
- 1.Walczak H., Degli-Esposti M.A., Rauch C.T. TRAIL-R2: a novel apoptosis-mediating receptor for TRAIL. EMBO J. 1997;16:5386–5397. doi: 10.1093/emboj/16.17.5386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Ashkenazi A. Targeting death and decoy receptors of the tumor-necrosis factor superfamily. Nat. Rev. Cancer. 2002;2:420–430. doi: 10.1038/nrc821. [DOI] [PubMed] [Google Scholar]
- 3.Valley C.C., Lewis A.K., Sachs J.N. Tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) induces death receptor 5 networks that are highly organized. J. Biol. Chem. 2012;287:21265–21278. doi: 10.1074/jbc.M111.306480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Lewis A.K., Valley C.C., Sachs J.N. TNFR1 signaling is associated with backbone conformational changes of receptor dimers consistent with overactivation in the R92Q TRAPS mutant. Biochemistry. 2012;51 doi: 10.1021/bi3006626. 6545–6455. [DOI] [PubMed] [Google Scholar]
- 5.Inbaraj J.J., Laryukhin M., Lorigan G.A. Determining the helical tilt angle of a transmembrane helix in mechanically aligned lipid bilayers using EPR spectroscopy. J. Am. Chem. Soc. 2007;129:7710–7711. doi: 10.1021/ja071587l. [DOI] [PubMed] [Google Scholar]
- 6.Karim C.B., Kirby T.L., Thomas D.D. Phospholamban structural dynamics in lipid bilayers probed by a spin label rigidly coupled to the peptide backbone. Proc Natl Acad Sci U S A. 2004;101:14437–14442. doi: 10.1073/pnas.0402801101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Kim T., Im W. Revisiting hydrophobic mismatch with free energy simulation studies of transmembrane helix tilt and rotation. Biophys. J. 2010;99:175–183. doi: 10.1016/j.bpj.2010.04.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Castillo N., Monticelli L., Tieleman D.P. Free energy of WALP23 dimer association in DMPC, DPPC, and DOPC bilayers. Chem. Phys. Lipids. 2013;169:95–105. doi: 10.1016/j.chemphyslip.2013.02.001. [DOI] [PubMed] [Google Scholar]
- 9.Karim C.B., Zhang Z., Thomas D.D. Synthesis of TOAC spin-labeled proteins and reconstitution in lipid membranes. Nat. Protoc. 2007;2:42–49. doi: 10.1038/nprot.2007.2. [DOI] [PubMed] [Google Scholar]
- 10.Grossfield, A. WHAM: The Weighted Histogram Analysis Method, Ver. 2.0.7, http://membrane.urmc.rochester.edu/content/wham.
- 11.Vilar M., Charalampopoulos I., Ibáñez C.F. Activation of the p75 neurotrophin receptor through conformational rearrangement of disulphide-linked receptor dimers. Neuron. 2009;62:72–83. doi: 10.1016/j.neuron.2009.02.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Cho H.S., Dominick J.L., Spence M.M. Lipid domains in bicelles containing unsaturated lipids and cholesterol. J. Phys. Chem. B. 2010;114:9238–9245. doi: 10.1021/jp100276u. [DOI] [PubMed] [Google Scholar]
- 13.De Angelis A.A., Jones D.H., Opella S.J. NMR experiments on aligned samples of membrane proteins. Methods Enzymol. 2005;394:350–382. doi: 10.1016/S0076-6879(05)94014-7. [DOI] [PubMed] [Google Scholar]
- 14.Klein J.C., Burr A.R., Thomas D.D. Actin-binding cleft closure in myosin II probed by site-directed spin labeling and pulsed EPR. Proc. Natl. Acad. Sci. USA. 2008;105:12867–12872. doi: 10.1073/pnas.0802286105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Pannier M., Veit S., Spiess H.W. Dead-time free measurement of dipole-dipole interactions between electron spins. J. Magn. Reson. 2000;142:331–340. doi: 10.1006/jmre.1999.1944. [DOI] [PubMed] [Google Scholar]
- 16.Jeschke G., Chechik V., Jung H. DEERANALYSIS2006—a comprehensive software package for analyzing pulsed ELDOR data. Appl. Magn. Reson. 2006;30:473–498. [Google Scholar]
- 17.Jeschke G. Interpretation of dipolar EPR data in terms of protein structure. In: Mingos D.M.P., editor. Structure and Bonding. Springer; Berlin, Germany: 2012. [Google Scholar]
- 18.Jo S., Kim T., Im W. CHARMM-GUI: a web-based graphical user interface for CHARMM. J. Comput. Chem. 2008;29:1859–1865. doi: 10.1002/jcc.20945. [DOI] [PubMed] [Google Scholar]
- 19.Romo T.D., Grossfield A. LOOS: an extensible platform for the structural analysis of simulations. Conf. Proc. IEEE Eng. Med. Biol. Soc. 2009;2009:2332–2335. doi: 10.1109/IEMBS.2009.5335065. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.