

Abstract
Background:
Hepatocellular carcinoma (HCC) is a multistep complex process, caused by many of genetic alteration. Insulin-like growth factors and their receptor have been widely implicated to HCC. Insulin-like growth factor-II (IGF-II) is a mitogenic polypeptide, found in various fetal and neonatal tissues of humans and rats and expresses in HCC. Here we investigated anticancer potential of phosphorothioate antisense oligonucleotides (ASOs) against three coding exons (exon-1/exon-2/exon-3) of IGF-II messenger ribonucleic acid in rat hepatocarcinogenesis model.
Materials and Methods:
During diethylnitrosamine and 2-acetylaminofluorene induced hepatocarcinogenesis, rats were treated with ASOs. Various biochemical and histological studies were conducted.
Results:
About 40% of carcinogen treated rats, which received two oligomers (against exon-1 or-3) did not show any hepatic lesion, hyperplastic nodule or tumor and remaining 60% of those rats showed lesion incidence and had about 59% and 55% reductions in the numbers of hepatic altered foci, respectively. Reductions in the total lesion-area when compared with carcinogen control rats were 64% and 53%, respectively for the animals treated with carcinogen and received the ASOs against exon-1/-3. Fluorescein isothiocyanate-labeled ASO reached in the hepatocytes in 2 h. No predominant IGF-II overexpression was observed in case of rats treated with the two ASOs. Treatment of the antisense IGF-II oligomers in carcinogen treated rats show better hepatocellular integrity along with several preneoplastic/neoplastic marker isoenzyme/enzyme modulations.
Conclusions:
Two of the three antisense oligomer-types effectively controlled IGF-II overexpression, causing the delay of the development and/or progress of hepatic cancer in rats.
Keywords: Antisense oligonucleotides, hepatic altered foci, hepatocellular carcinoma, hyperplastic nodules, insulin-like growth factor
INTRODUCTION
Hepatocellular carcinoma (HCC) that arises from hepatocytes is a common fatal solid malignant tumors world-wide.[1,2] HCC is characterized by multiple causes, multiple stages and multifocal process of tumorigenesis related intimately to the overexpression/suppression of many cellular factors.[3,4] In general, chemical-induced liver cancer (hepatocarcinogenesis) animal models have been widely used for investigating therapeutic and other combating strategies in vivo against HCC. Transformation of hepatocytes to HCC occurs through multiple genetic alterations and protein expressions. One of which is insulin-like growth factor-II (IGF-II). IGF-II is a mitogenic polypeptide structurally and functionally related to insulin.[5] IGF-II, a ligand of the insulin-like growth factor-I receptor (IGF-IR), is believed to exert its effect during cellular proliferation and widely distributed in various fetal and neonatal human and rat tissues, including liver and during HCC.[6] A relationship exists between the IGF/IGF-IR systems in development of various types of cancer, including HCC.[7,8] Report suggests that IGF-II promotes hepatocyte proliferation through a paracrine mechanism in the pre-cancerous stage[9] and when hepatocytes are transformed into malignant cells, they secrete IGF-II and promote malignant cell proliferation by an autocrine mechanism.[9]
Thus, an appropriate silencing of IGF-II gene overexpression by synthetic antisense oligonucleotides/oligomers (ASOs) may be useful in the treatment of liver tumors in which IGF-II has a predominant role, as ASOs are believed to possess great specificity to an intended molecular target.[10]
In the present study, we have investigated the effectiveness of phosphorothioate ASOs targeted against IGF-II messenger ribonucleic acids (mRNAs) of three coding exons (exon-1, exon-2 and exon-3) in controlling the progress of rat hepatocarcinogenesis.
MATERIALS AND METHODS
Oligonucleotides
The 30-base sequence 5’-TCCATGGTTACC CCTAGGGTCACCCCTTCA-3’ targeted against exon-1 partially (ASO-1), 24-base sequence 5’-CGTCCGGAAGTTCGGCACGGTTGG-3’ targeted against exon-2 partially (ASO-2) and 32-base sequence 5’-TGCTGAAGGGGTCTATGGGGCACCCG TTCAAG-3’ targeted against exon-3 partially (ASO-3) synthesized on a 0.05 μmol scale, HPLC purification and phosphorothioate backbone modification (synthesized by Eurofins Genomics India Pvt. Ltd., Bangalore, India) were used in this study. A set of them also contained fluorescein isothiocyanate (FITC)-labeled IGF-II ASOs used for in vivo localization of IGF-II ASOs.
Rat cancer model
Male Sprague-Dawley rats (procured from the Indian Institute of Chemical Biology, Kolkata, India) with body weights approximately 130-150 g were maintained in polypropylene cages and housed at a temperature (25°C ± 1°C) and a relative humidity 55% ±5% with a normal day and night photoperiod, in the university animal house. They have been acclimatized to the facility for 2 weeks. All the animal experiments were conducted upon receiving the approval of the Animal Ethics Committee, Jadavpur University. Rats were divided into eight groups [Figure 1] - (Groups A-H). Group-A (normal control rats) and Group-B (carcinogen control rats), Group-C (carcinogen treated rats received ASO-1), Group-D (carcinogen treated rats received ASO-2), Group-E (carcinogen treated rats received ASO-3), Group-F (normal rats received ASO-1), Group-G (normal rats received ASO-2) and Group-H (normal rats received ASO-3). Each group contained 10 rats.
Figure 1.
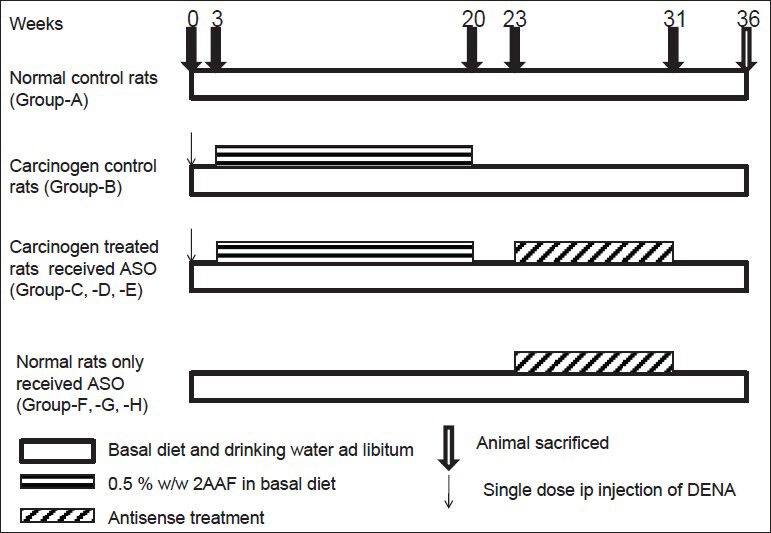
Experimental regimen. ASO: Antisense oligomer, 2AAF: 2-acetylaminfluorine, DENA: Diethylnitrosamine, ip: Intraperitoneal
In Groups B-E animals, carcinogenesis was initiated in liver by diethylnitrosamine (Sigma Aldrich, Bangalore, India) at a dose of 200 mg/kg body weight once intraperitoneally at the start of the experiment, i.e., day 0.[3] The animals were then treated with 0.5% w/w of 2-acetylaminofluorene (Sigma Aldrich, Bangalore, India) in the diet to promote carcinogenesis.[3] 2-Acetylaminofluorene was mixed with a small amount of food and given every morning.[11] After 6 h they were given the basal diet.[11] The rats had free access to water. Carcinogenesis was promoted from the beginning of 3rd week to the end of 20th week, i.e., for a period of 18 weeks. Groups C-E animals received i.v. injection of ASO-1/ASO-2/ASO-3 in normal saline (5 mg/kg body weight) thrice a week, respectively [Figure 1]. IGF-II ASO treatment was started from 23rd week and continued up to 31st week in Groups C-E rats. Groups F-H received the treatment of ASO-1/ASO-2/ASO-3, respectively. At the 36th week, the animals were sacrificed[12] and further experiments were conducted. All animals remained unfed for 12 h before being killed. The livers were removed, sliced (5-10 mm thick) and were snap-frozen in liquid nitrogen. The tissues were stored at −80°C in ultra-low temperature freezer (So-Low, USA) until further use.
Liver morphology study
Liver tissues were subjected to macroscopic examination on the surfaces for visible hyperplastic nodules (HNs). The nodules were identified and differentiated from the reddish brown non-nodular surrounding liver by their grayish-white and darkish red color with clear demarcation.[13]
Histopathological study
The liver tissue samples were undergone serial sections of hepatic tissues (6 μm thick), which were further processed histochemically by periodic acid-Schiff reaction, toluidine blue and hematoxylin-eosin for investigating tissue architecture and detecting different hepatic altered foci (HAF).[13] Liver lesions were identified as HAF[3,14] and were classified as clear endoplasmic reticulum rich clear cell foci (ground glass appearance), mixed cell foci and ribosome rich basophilic foci.[13] Numbers and area of lesions were determined by Zeiss light microscope and Axio Vision software 4.7.1. (Jena, Germany).
Marker enzymes study
Microsomal cytochrome P-450 (cyt.P-450),[15] glutathione S-transferase (GST) activity[16] and uridine diphosphate glucuronyl transferase (UDPGT) activity[17] were determined using the referred methods. Catalase activity using catalase activity assay kit (BioVision, Milpitas, USA), superoxide dismutase (SOD) activity using SOD activity assay kit (BioVision, Mountain View, USA) and protein using modified Lowry protein assay kit (Thermo Scientific, Rockford, USA) were determined.
Detection of IGF-II mRNA by in situ hybridization
In situ hybridization using IGF-II sense and antisense digoxigenin-labeled mRNA obtained by in vitro transcription was conducted on 6 μm cryosections of the liver samples to localize IGF-II gene expression in the cancerous and non-cancerous rat hepatic tissues.[13]
Field emission scanning electron microscopy of liver tissue
Morphology of liver tissue from different experimental groups of rats was performed with the help of FESEM (JSM Electron microscope, JEOL, Tokyo, Japan). Processed tissue samples were placed on to a carbon tape over a stub. The platinum coating was applied on samples using an ion sputtering device, dried in a vacuum and examined using FESEM.
In vivo tissue localization of FITC-labeled IGF-II ASOs by confocal laser scanning microscope
Intravenous injection of FITC – labeled IGF-II ASOs was given in normal rats. Rats were dissected at different time points (after injection) 2 h, 4 h, 8 h and 12 h. Liver samples were collected, washed and fixed with formalin solution and embedded in paraffin blocks by the conventional methods.[18] The unstained tissue sections were observed at ×400 magnification under a confocal laser scanning microscope (LSM 510; Carl Zeiss, Jena, Germany) at an excitation wavelength of 494 nm and emission wavelength of 521 nm to observe the distribution of IGF-II ASOs in liver tissue.
In vivo tissue uptake of FITC-labeled IGF-II ASOs by spectrofluorimeter
FITC-labeled oligomers in liver were quantified using a spectrofluorimeter (Perkin-Elmer LS 55, Walthan, USA) with excitation and emission wavelength at 494 nm and 521 nm respectively, for FITC activity.[19]
Statistical calculations
Statistical calculations were performed with GraphPad Instat version 3.0 (GraphPad Software, Inc., San Diego California). The data were analyzed either by one-way analysis of variance or by Dunnett's t-test to determine statistical level of significance with a P < 0.05.
RESULTS
Cellular uptake of all the three ASOs (ASO-1,-2 and-3) along with their sense controls were studied in vitro in rat hepatoma H4IIEC3 cells (B.D Biosciences, Mumbai, India). All the sense and antisense oligomers were found to be internalized by the cells as detected by using FITC labeled oligomers with the help of confocal microscopy and all the antisense oligomers (ASO-1,-2 and -3) were found to control cellular proliferation investigated by thymidine incorporated deoxyribonucleic acid (DNA) synthesis[12] in cells (data not shown), although ASO-2 had about 30% less activity as compared to ASO-1 and -3 in this regard. All the three sense oligomers had a similar level of DNA synthesis as detected for untreated rat hepatoma H4IIEC3 cells. Thus, the sense treatments remained ineffective with respect to cellular proliferations. Antisense oligomers therefore were administered in carcinogen treated rats and in normal rats to investigate their efficacy in vivo. Interestingly, in normal animals, antisense oligomers had no predominant effect. This may be due to the absence of IGF-II expression in normal rats. Carcinogen control rats (Group-B) had maximum numbers and size of macroscopic hepatic HNs [Figure 2a] in liver. Appearance of HNs varied predominantly [Figure 2b] in carcinogen treated rats treated with or without IGF-II ASOs. Group-B rats had hepatic tumor [Figure 2c]. Carcinogen treated rats received ASO-1 (Group-C) had minimum number of HNs on the liver surfaces and the carcinogen treated rats received ASO-2 (Group-D) had the maximum number of HNs and tumor in liver, among the carcinogen with ASO treated rats. Tumor incidences were not seen in Group-C and Group-E rats. Hepatic HNs, preneoplastic and neoplastic lesions and tumors were not seen in case of normal control (Group-A) rats and rats treated with ASO-1/ASO-2/ASO-3 (Groups F-H, respectively). Group-B and Group-D rats showed 100% incidence of liver lesions and hepatic tumors. However, 40% of carcinogen treated rats which received ASO-1 or ASO-3 did not show any hepatic lesions, HNs or tumors [Table 1].
Figure 2.
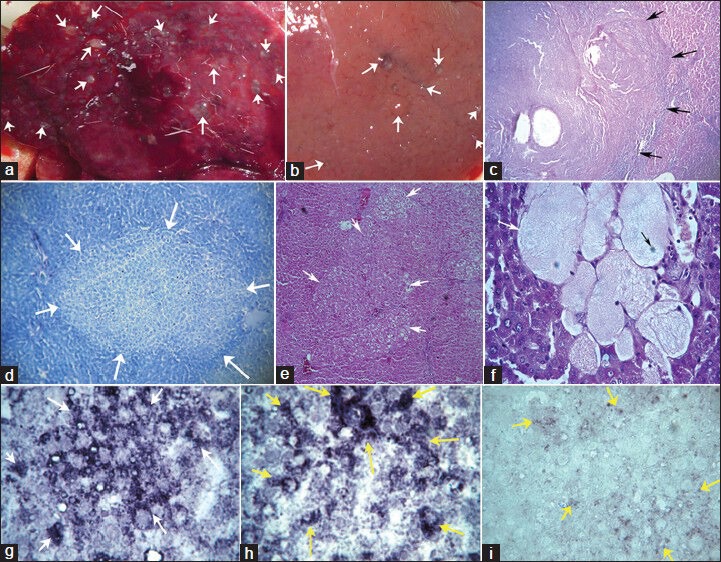
Morphology, histology, insulin-like growth factor-II (IGF-II) gene expression in experimental rats. (a and b) External morphology of liver showing multiple hyperplastic nodules (white arrow) on liver surface of Group-B and Group-C rats, respectively. (c) Section of hepatic tumor (shown by black arrow) in hepatocellular carcinoma (Group-B); using periodic acid-Schiff reaction ×100. (d) A prominent hepatic mixed cell lesion (shown by white arrow) in Group-B rats, using toluidine blue ×100. (e) Highly differentiated HCC composed of a mixed population of clear (glycogenetic) cells, acidophilic cells (ground-glass hepatocytes) and some glycogen-poor, basophilic cells with PAS in Group-B rats ×100. (f) Section of experimental rat hepatic tissue showing spongiosis hepatis (white arrow) with nucleus (black arrow) with PAS in Group-B rats ×400. (g and h) IGF-II overexpressed lesions in rat liver shown by white arrow and yellow arrow of Group-B and Group-D rats respectively ×400. (i) Shows IGF-II expression in scattered hepatocytes in Group-C rats, shown by yellow arrow ×400
Table 1.
Effect of antisense oligomers on number of HAF lesions and area of lesions

In Group-B and Groups C-E rats, numbers of HAF were counted/unit area (/cm2) of hepatic tissue [Table 1]. Maximum count for HAF was observed in Group-B rats, followed by Group-D rats. The values were numerically close to each other. However, 60% of Group-C and Group-E rats showed lesion incidence and had about 59% and 55% reduction in the numbers of HAF, respectively. Reductions in the total lesion area compared to Group-B rats were 64% and 53%, respectively for Group-C and Group-E rats. Group-D rats had total lesion area even slightly more (statistically non-significant) compared to Group-B rats. Clear cell lesions (ground glass appearance),[13] basophilic lesions and mixed cell lesions [Figure 2d and e] were predominantly scattered in hepatic tissues. Those lesions were significantly (P < 0.05) less in numbers, size and area in Group-C and Group-E animals [Table 1]. Group-B and Group-D rats had some spongioblast cells [Figure 2f].
IGF-II overexpression as focal lesions and in individual hepatocytes was observed in Group-B and Group-D rats [Figure 2g and h]. No predominant IGF-II overexpression was observed in rats treated with ASO-1/-3 (for Group-C/Group-E rats). However, scattered IGF-II expression in few hepatocytes was seen in some of those rats [Figure 2i].
We further investigated whether the experimental IGF-II ASOs (FITC-labeled) reached liver after their intravenous administration. Livers were eventually rich in FITC fluorescence with the increasing period of time (until 12 h in this case) [Figure 3a]. This was also quantified spectrofluorometrically. FITC labeled IGF-II ASO was about 0.25 nM/g of hepatic tissue after 12 h of administration. The value was about 13 times more than that at 2 h, with an increasing trend of values through 4-8th h of assessment [Figure 3b].
Figure 3.
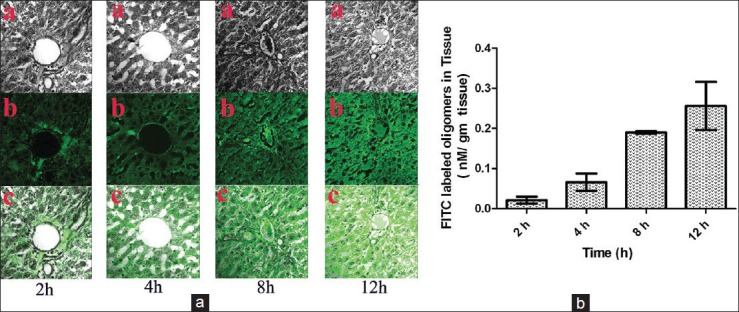
Hepatic localization of fluorescein isothiocyanate-labeled insulin-like growth factor-II (IGF-II) antisense oligomers and quantification of FITC-labeled oligomers in experimental rats. (a) Confocal microscopic image of liver section of rats treated with FITC-labeled IGF-II antisense oligomer ×400. a: Tissue alone, b: FITC-oligomer alone, c: FITC-oligomer in liver tissue. (b) Tissue content of FITC-labeled oligomer in liver tissue of experimental animals
Another interesting study with the experimental livers using a simple FESEM technique provides some significant findings related to the structure of the hepatocytes and the tissues. In many cases, adjacent cell walls were not connected in Group-A [Figure 4a and b] and Groups F-H rats (data not shown). The cellular structures appear well preserved with nuclei and on occasions, the spaces between hepatocytes were occupied by non-cellular materials [Figure 4b]. Carcinogen control livers had cellular structures without much space between the hepatocytes compared with Group-A and Groups F-H rat livers. Nuclei were comparatively positioned centrally in the cells and more number of binucleated cells often existed in Group-B rats [Figure 4c and d]. Less fine granular cytoplasm with less structural compactness was seen in Group-B rats when compared to Group-A and Groups F-H rats. Treatment of IGF-II ASO in carcinogen treated rats (Group-C) show better hepatocellular integrity [Figure 4e and f] compared with that of Group-B rats. Group-E rats had similar results (data not shown). Group-D rats had hepatocellular architecture similar (data not shown) to that of Group-B rats.
Figure 4.
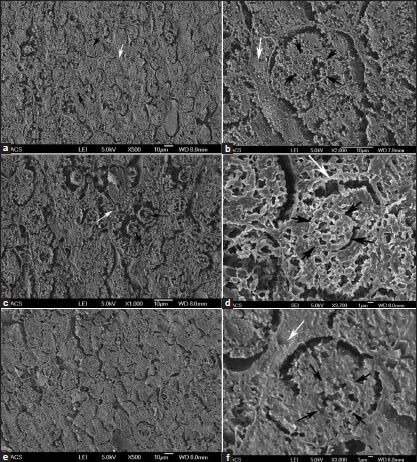
Field emission scanning electron microscopic images of experimental hepatic tissues. (a and b) Normal control rat liver tissues (Group-A). (c and d) Carcinogen control rat liver tissue (Group-B). (e and f) Carcinogen treated rats received antisense oligomer-1 (Group-C) rat liver tissue. White arrows indicate space between the hepatocytes and black arrows indicate nucleus of the cell
Variation in activities or levels of some important marker isoenzyme/enzymes known for their role in hepatocellular cancer development, were observed in the livers of the experimental rats.[3]
In Group-B rats, the content of cyt.P-450 was reduced compared to normal control animals (Group-A) [Figure 5a]. Upon comparison of cyt.P-450 content of Group-B animals with those values of Groups C-E rats, cyt.P-450 contents were found to increase significantly (P < 0.05) in Group-C and Group-E rats, whereas in Group-D rats the isoenzyme content was almost similar to that of Group-B rats. Groups F-H and Group-A rats had significantly (P < 0.05) higher values of cyt.P-450 content than that of Group-B rats. Cyt.P-450 content was about double (P < 0.05) in Group-F rats when compared to Group-A rats.
Figure 5.
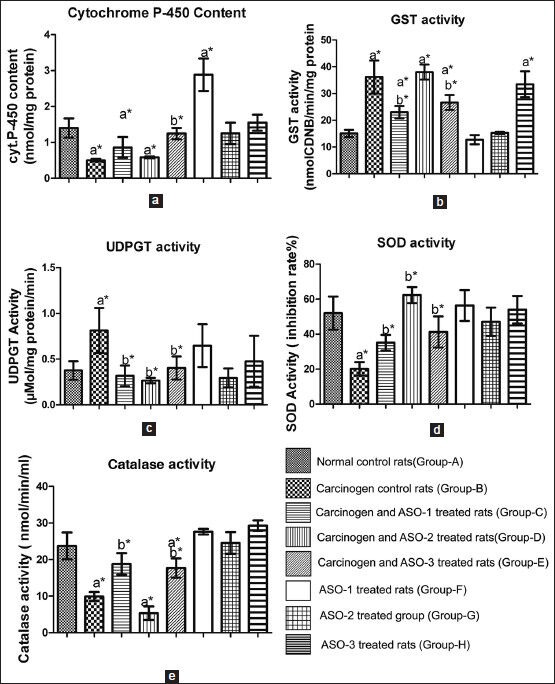
Various enzyme activities, iso-enzyme level in hepatic tissues of the experimental rats, (a) cytochrome P-450 content, (b) glutathione S-transferase activity, (c) uridine diphosphate glucuronyl transferase activity, (d) superoxide dismutase activity, (e) catalase activity. Values represent mean ± standard deviation (n = 10), * P < 0.05, a s tatistical l evel o f s ignificance (using D unnett's t-test) when all groups were compared with Group-A, b statistical level of significance (using Dunnett's t-test) when Group-C, Group-D, and Group-E were compared with Group-B
In Group-B animals, the activities of phase-II biotransforming enzymes such as GST and UDPGT were significantly (P < 0.05) higher [Figure 5b and c compared to Group-A rats. When compared to Group-B rats, Group-C and Group-E rats had significantly decreased (P < 0.05) GST and UDPGT activities. In Group-D animals, GST activity was similar to that found in Ground-B rats. When Groups F-H rats were compared with Group-A rats, there was no significant change of GST activity in Group-F and Group-G rats. However administration of ASOs in Group-H animals enhanced GST activity twice more than that value in Group-A rats. UDPGT activity was decreased to near normal values in Groups C-E rats when compared to Group-B rats which had about 2 times higher UDPGT activity than that in Group-A rats. Injection of ASOs in Group-F and-H rats caused enhancement of UDPGT activity compared with Group-A rats, although the data were not statistically significant. Hepatic SOD markedly decreased (about 2.5 times) in Group-B rats compared to Group-A animals [Figure 5d]. SOD activity was significantly (P < 0.05) higher in Groups C-E rats when compared to Group-B rats, upon ASOs treatment. However, except for Group-D animals, the values were less than those in Group-A rats. Rats belonged to Groups F-H did not alter SOD activities significantly as compared to Group-A rats. Similar trend was found in case of catalase activity in carcinogen treated rats which received ASOs [Figure 5e], except in Group-D rats. Catalase activity was found to decrease significantly (P < 0.05) in Group-B rats compared to Group-A rats. Upon ASOs administration (except ASO-2 which had even less activity of catalase than the carcinogen control animals) in Groups-C and-E rats, the activity was found to enhance toward the normal value. No predominant statistically significant difference in the activity of catalase was observed in Groups F-H rats compared to Group-A rats.
DISCUSSION
All the carcinogen control animals showed 100% incidences of HAF and tumors along with HNs scattered predominantly on liver surface. ASO-1/ASO-3 treatment in carcinogen treated rats showed no HAF and tumor in 40% of the rats and remaining 60% of the rats of those groups showed HAF without any tumor incidence and reddish HNs much smaller in size and less in number compared to grey-white, larger HNs of Group-B rats. ASO-1/ASO-3 thus successfully either ceased or reduced the liver cancer incidence in rats. This might be by modulating some genetic pathways in which IGF-II gene has a predominant role in cell signaling for HCC development.[20] Application of ASO-2 in carcinogen-treated rats was found to be ineffective to control HCC development as the group of rats had 100% incidence of HAF and tumors in liver.
Overexpression of IGF-II has been found in HCC and in hepatic lesions.[13,21] In Group-B and Group-D rats, overexpression of IGF-II gene was observed in focal lesions and in some scattered hepatocytes. Hence ASO-2 did not successfully control IGF-II gene overexpression. Rats of Group-C and Group-E (the rats of those two groups in which HAF were seen) had very few scattered IGF-II gene-expressed hepatocytes, suggesting silencing of IGF-II gene by ASO-1/ASO-3 in those rats to a great extent.
Further confocal microscopic images of the liver sections showed that ASOs reached the hepatocytes within 2 h and the quantity of oligomers increased with time (at least up to 12 h).
FESEM data depict that there was a loss of structural integrity in the livers of carcinogen control rats. More dividing cells were observed. Unlike normal control rat livers, cytoplasm had more space with less finely granular appearance. ASO-1/ASO-3 treatment in carcinogen treated rats show hepatocytes with more compact cytoplasm with finer granular structure. Nuclei were positioned more centrally in the hepatocytes as in carcinogen control animals, but space between the hepatocytes was more similar to those in normal control rats. Thus, ASO-1/-3 treatment might partially control the structural changes of hepatocytes during the transformation of normal cells to neoplastic cells.[12,22]
Various enzyme activities and isoenzyme level have long been considered as the hepatic preneoplastic and/or neoplastic markers and many studies have attested to this claim by the findings.[3,12] Cyt.P-450 is an isoenzyme of a collection of cyt.P-450 monooxygenases, which catalyze different types of the oxidation reaction.[23] This enzyme family plays an important role in phase-I metabolism of many drugs. The enzymes are present in many tissues but the highest levels are found in liver.[24] Many earlier findings also showed decreased cyt.P-450 content in rat liver carcinogenesis.[25] Our data suggest that oxidizing activities through cyt.P-450 enzymes might be suppressed due to the presence of carcinogen and unlike ASO-1/-3 treatment, ASO-2 administration had no marked effect to improve cyt.P-450 enzyme activity in carcinogen treated rats. A significant 100% enhancement of cyt.P-450 content in Group-F animals (i.e., ASO-1 control rats) might be due to the use of cyt.P-450 enzymes much more in livers of those rats during the degradation of the phosphorothioate oligomer or its products compared with the rats of Groups G and H which showed no predominant change in cyt.P-450 contents. GST and UDPGT (two important phase II enzymes) are known to play a key role in detoxification of both xenobiotics and endogenous compounds and in reduction of reactive oxygen species (ROS) and DNA adduct formation.[26] GST and UDPGT are known to be important preneoplastic and neoplastic determinants both in the susceptibility to mutagenic effects of chemical carcinogenesis and in the response of liver tumors to chemotherapy.[27,28] An increase in GST activity is therefore an elevation in the mechanism that protects against the noxious effects of xenobiotics, including carcinogens.[29] Unlike carcinogen control rats, the rats treated with ASO-1/ASO-3 decreased GST activity significantly toward the normal value. However, carcinogen treated animals received ASO-2 (Group-D) had GST activity similar to that of carcinogen control animals, indicating GST activity was not altered by the administration of ASO-2. ASO-1/-2 treatment in normal rats had no predominant effect on GST activity. ASO-3 treatment in normal rats enhanced GST activity by almost 2.5 times of that of normal control rats. GST might have a direct role to degrade ASO-3 or its products by transferase activity or ASO-3 has a direct role to induce GST activity. The synthesis of glucuronides by liver microsomal UDPGT is a major pathway by which the body inactivates both endogenous and foreign compounds.[30] A significantly (P < 0.05) higher UDPGT activity in carcinogen control rats than normal control rats was reduced to near normal value upon application of ASO-1/-3, suggesting that ASO-1/-3 influence the enzyme to inactivate xenobiotics in carcinogen treated rats. ASO-1/ASO-3 (although statistically non-significant) enhanced UDPGT activity in normal rats. However, ASO-2 did not alter UDPGT activity in ASO-2 control rats. Carcinogen administration produces ROS, which can induce several kinds of DNA damage, activation of transcription factors and proto-oncogenes, genomic instability, invasion or metastasis etc.[31] Many antioxidant enzymes such as SOD, catalase, glutathione peroxidase and reductase etc., prevent or repair the damage caused by ROS. SOD and catalase thus protect liver against carcinogen induce hepatotoxicity. It is reported that during hepatocarcinogenesis both SOD and catalase levels decrease, except some cancer cells which process high level of SOD expression and activity.[32] Catalase activity was not significantly different between normal control rats and IGF-II ASO control rats. In carcinogen control animals, the value was reduced significantly compared with normal animals. Carcinogen treated animals received ASO-1/ASO-3 enhanced catalase activities toward the normal control value compared to carcinogen control animals. However, the application of ASO-2 in carcinogen treated animals did not show any enhancement, rather reduced the catalase activity in carcinogen treated rats. Results indicate that treatment of ASO-1/ASO-3 significantly improved the antioxidant defense enzyme system, in particular catalase activity. The differential toxicity due to ROS generated by carcinogen in hepatocytes as compared to normal animals may be brought about by the lack of SOD in preneoplastic and neoplastic cells, as loss of SOD (or increased ROS) occurs in all cancerous cells.[33] Thus loss of SOD from the normal cells causes the cells to appear transformed in some ways or makes the cells more susceptible to transformation. This explains the lower SOD level in the carcinogen control animals. SOD activity did not vary significantly in ASO treated rats (ASO control) compared to normal animals, indicating that in normal rats ASO-1/-2/-3 had no effect on SOD. The activity was reduced to 2.5 times in carcinogen control animals. In Groups C-E, the activities were enhanced to near normal value. The findings suggest that administration of ASO-1/-2/-3 had induced SOD activity directly or indirectly by triggering some other enzyme systems in carcinogen treated rats.
CONCLUSIONS
Out of the three ASOs used here to block IGF-II gene overexpression, ASO-1 was found to control most significantly the development and/or progress of hepatic cancer. ASO-3 was found to be almost equipotent. However, ASO-2 was found to be ineffective. Function of antisense oligomers in vivo depends on many facts such as mRNA-antisense oligomer duplex formation, rate of binding of oligomers with mRNA, stability of the formed duplex in vivo etc.[34,35] In the present study, production of mRNA in vivo might be much faster and distinctly more in an amount than the mRNA-ASO-2 duplex formation. This has been supported by the findings of overexpression of IGF II in ASO-2 administered carcinogen treated rat livers. Occurrence of quicker degradation and/or elimination of mRNA-ASO-2 duplex in vivo should not be ignored, too. ASO-2 could show non-antisense effects as a report suggests that the presence of certain oligonucleotide sequences, such as CG (which were maximum in numbers in ASO-2), have sequence-dependent, non-antisense effects.[36] Besides, ASO-2 might have some non-specific binding with some other macromolecules. Thus, ASO-2 found to be ineffective in vivo may be because of any one of the mentioned reasons or a combine effect of some of the mentioned reasons or due to a cumulative effect of them. However, further studies are warranted in the area.
AUTHOR'S PROFILE
Mr. Miltu Kumar Ghosh, Department of Pharmaceutical Technology, Jadavpur University, Kolkata - 700 032, West Bengal, India and Georg-August-Universität Göttingen, Department of Human Genetics, Heinrich-Düker-Weg 12, 37073 Göttingen, Germany.
Mr. Falguni Patra, Department of Pharmaceutical Technology, Jadavpur University, Kolkata - 700 032, West Bengal, India.
Dr. Shampa Ghosh, Department of Pharmaceutical Technology, Jadavpur University, Kolkata - 700 032, West Bengal, India.
Mr. Chowdhury Mobaswar Hossain, Department of Pharmaceutical Technology, Jadavpur University, Kolkata - 700 032, West Bengal, India.
Prof. (Dr.) Biswajit Mukherjee, Department of Pharmaceutical Technology, Jadavpur University, Kolkata - 700 032, West Bengal, India.
ACKNOWLEDGEMENTS
The work has been carried out with the fund granted to Dr. Biswajit Mukherjee as a major research project from Indian Council of Medical Research, Govt. of India (grant no. 58/7/2009-BMS). We also acknowledge Bose Institute, Kolkata and Indian Association for the Cultivation of Science, Kolkata for providing confocal microscopy and FESEM facilities, respectively.
Footnotes
Source and Support: The work has been carried out with the fund granted to Dr. Biswajit Mukherjee as a major research project from Indian Council of Medical Research (ICMR), Govt. of India (grant no. 58/7/2009-BMS). We also acknowledge Bose Institute, Kolkata and Indian Association for the Cultivation of Science, Kolkata for providing confocal microscopy and FESEM facilities, respectively.
Conflict of Interest: None declared.
REFERENCES
- 1.Blum HE. Hepatocellular carcinoma: HCC. Hepat Mon. 2011;11:69–70. [PMC free article] [PubMed] [Google Scholar]
- 2.Bosch FX, Ribes J, Díaz M, Cléries R. Primary liver cancer: Worldwide incidence and trends. Gastroenterology. 2004;127:S5–16. doi: 10.1053/j.gastro.2004.09.011. [DOI] [PubMed] [Google Scholar]
- 3.Mukherjee B, Das T, Ghosh S, Datta S. Changes in the antioxidant defense and hepatic drug metabolizing enzyme and isoenzyme levels, 8-hydroxydeoxyguanosine formation and expressions of c-raf. 1 and insulin-like growth factor II genes during the stages of development of hepatocellular carcinoma in rats. Eur J Cancer Prev. 2007;16:363–71. doi: 10.1097/01.cej.0000236254.01608.33. [DOI] [PubMed] [Google Scholar]
- 4.Ong TP, Cardozo MT, de Conti A, Moreno FS. Chemoprevention of hepatocarcinogenesis with dietary isoprenic derivatives: Cellular and molecular aspects. Curr Cancer Drug Targets. 2012;12:1173–90. doi: 10.2174/156800912803987986. [DOI] [PubMed] [Google Scholar]
- 5.Chao W, D’Amore PA. IGF2: Epigenetic regulation and role in development and disease. Cytokine Growth Factor Rev. 2008;19:111–20. doi: 10.1016/j.cytogfr.2008.01.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Maki RG. Small is beautiful: Insulin-like growth factors and their role in growth, development, and cancer. J Clin Oncol. 2010;28:4985–95. doi: 10.1200/JCO.2009.27.5040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Samani AA, Yakar S, LeRoith D, Brodt P. The role of the IGF system in cancer growth and metastasis: Overview and recent insights. Endocr Rev. 2007;28:20–47. doi: 10.1210/er.2006-0001. [DOI] [PubMed] [Google Scholar]
- 8.LeRoith D, Roberts CT., Jr The insulin-like growth factor system and cancer. Cancer Lett. 2003;195:127–37. doi: 10.1016/s0304-3835(03)00159-9. [DOI] [PubMed] [Google Scholar]
- 9.Wang Z, Ruan YB, Guan Y, Liu SH. Expression of IGF-II in early experimental hepatocellular carcinomas and its significance in early diagnosis. World J Gastroenterol. 2003;9:267–70. doi: 10.3748/wjg.v9.i2.267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Stahel RA, Zangemeister-Wittke U. Antisense oligonucleotides for cancer therapy-an overview. Lung Cancer. 2003;41(Suppl 1):S81–8. doi: 10.1016/s0169-5002(03)00147-8. [DOI] [PubMed] [Google Scholar]
- 11.Mukherjee B, Basu M, Chatterjee M. Effect of selenomethionine on N-methylnitronitrosoguanidine-induced colonic aberrant crypt foci in rats. Eur J Cancer Prev. 2001;10:347–55. doi: 10.1097/00008469-200108000-00008. [DOI] [PubMed] [Google Scholar]
- 12.Das T, Patra F, Mukherjee B. Effect of antisense oligomer in controlling c-raf. 1 overexpression during diethylnitrosamine-induced hepatocarcinogenesis in rat. Cancer Chemother Pharmacol. 2010;65:309–18. doi: 10.1007/s00280-009-1035-4. [DOI] [PubMed] [Google Scholar]
- 13.Mukherjee B, Ghosh S, Das T, Doloi M. Characterization of insulin-like-growth factor II (IGF II) mRNA positive hepatic altered foci and IGF II expression in hepatocellular carcinoma during diethylnitrosamine- induced hepatocarcino genesis in rats. J Carcinog. 2005;4:12. doi: 10.1186/1477-3163-4-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Bannasch P. Glycogenotic hepatocellular carcinoma with glycogen-ground-glass hepatocytes: A heuristically highly relevant phenotype. World J Gastroenterol. 2012;18:6701–8. doi: 10.3748/wjg.v18.i46.6701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Omura T, Sato R. The carbon monoxide-binding pigment of liver microsomes. I. Evidence for its hemoprotein nature. J Biol Chem. 1964;239:2370–8. [PubMed] [Google Scholar]
- 16.Habig WH, Pabst MJ, Jakoby WB. Glutathione S-transferases. The first enzymatic step in mercapturic acid formation. J Biol Chem. 1974;249:7130–9. [PubMed] [Google Scholar]
- 17.Bock KW, Fröhling W, Remmer H, Rexer B. Effects of phenobarbital and 3-methylcholanthrene on substrate specificity of rat liver microsomal UDP-glucuronyltransferase. Biochim Biophys Acta. 1973;327:46–56. doi: 10.1016/0005-2744(73)90102-2. [DOI] [PubMed] [Google Scholar]
- 18.Sinha B, Mukherjee B, Pattnaik G. Poly-lactide-co-glycolide nanoparticles containing voriconazole for pulmonary delivery: In vitro and in vivo study. Nanomedicine. 2013;9:94–104. doi: 10.1016/j.nano.2012.04.005. [DOI] [PubMed] [Google Scholar]
- 19.Villa AE, Guzman LA, Poptic EJ, Labhasetwar V, D’Souza S, Farrell CL, et al. Effects of antisense c-myb oligonucleotides on vascular smooth muscle cell proliferation and response to vessel wall injury. Circ Res. 1995;76:505–13. doi: 10.1161/01.res.76.4.505. [DOI] [PubMed] [Google Scholar]
- 20.Nussbaum T, Samarin J, Schirmacher P, Breuhahn K. Insulin-like growth factor (IGF)-signalling pathway components are potential therapeutic targets in the treatment of human hepatocellular carcinoma. Verh Dtsch Ges Pathol. 2007;91:278–85. [PubMed] [Google Scholar]
- 21.Su Q, Liu YF, Zhang JF, Zhang SX, Li DF, Yang JJ. Expression of insulin-like growth factor II in hepatitis B, cirrhosis and hepatocellular carcinoma: Its relationship with hepatitis B virus antigen expression. Hepatology. 1994;20:788–99. doi: 10.1002/hep.1840200404. [DOI] [PubMed] [Google Scholar]
- 22.Bannasch P, Mayer D, Hacker HJ. Hepatocellular glycogenosis and hepatocarcinogenesis. Biochim Biophys Acta. 1980;605:217–45. doi: 10.1016/0304-419x(80)90005-0. [DOI] [PubMed] [Google Scholar]
- 23.Millonig G, Wang Y, Homann N, Bernhardt F, Qin H, Mueller S, et al. Ethanol-mediated carcinogenesis in the human esophagus implicates CYP2E1 induction and the generation of carcinogenic DNA-lesions. Int J Cancer. 2011;128:533–40. doi: 10.1002/ijc.25604. [DOI] [PubMed] [Google Scholar]
- 24.Matsubara T, Koike M, Touchi A, Tochino Y, Sugeno K. Quantitative determination of cytochrome P-450 in rat liver homogenate. Anal Biochem. 1976;75:596–603. doi: 10.1016/0003-2697(76)90114-7. [DOI] [PubMed] [Google Scholar]
- 25.Habib SL, Srikanth NS, Scappaticci FA, Faletto MB, Maccubbin A, Farber E, et al. Altered expression of cytochrome P450 mRNA during chemical-induced hepatocarcinogenesis and following partial hepatectomy. Toxicol Appl Pharmacol. 1994;124:139–48. doi: 10.1006/taap.1994.1017. [DOI] [PubMed] [Google Scholar]
- 26.Rowland A, Miners JO, Mackenzie PI. The UDP-glucuronosyltransferases: Their role in drug metabolism and detoxification. Int J Biochem Cell Biol. 2013;45:1121–32. doi: 10.1016/j.biocel.2013.02.019. [DOI] [PubMed] [Google Scholar]
- 27.Satoh K, Kitahara A, Soma Y, Inaba Y, Hatayama I, Sato K. Purification, induction, and distribution of placental glutathione transferase: A new marker enzyme for preneoplastic cells in the rat chemical hepatocarcinogenesis. Proc Natl Acad Sci U S A. 1985;82:3964–8. doi: 10.1073/pnas.82.12.3964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Bock KW, Lilienblum W, Pfeil H, Eriksson LC. Increased uridine diphosphate-glucuronyltransferase activity in preneoplastic liver nodules and Morris hepatomas. Cancer Res. 1982;42:3747–52. [PubMed] [Google Scholar]
- 29.Tatematsu M, Mera Y, Ito N, Satoh K, Sato K. Relative merits of immunohistochemical demonstrations of placental, A, B and C forms of glutathione S-transferase and histochemical demonstration of gamma-glutamyl transferase as markers of altered foci during liver carcinogenesis in rats. Carcinogenesis. 1985;6:1621–6. doi: 10.1093/carcin/6.11.1621. [DOI] [PubMed] [Google Scholar]
- 30.Hanioka N, Oka H, Nagaoka K, Ikushiro S, Narimatsu S. Effect of UDP-glucuronosyltransferase 2B15 polymorphism on bisphenol A glucuronidation. Arch Toxicol. 2011;85:1373–81. doi: 10.1007/s00204-011-0690-5. [DOI] [PubMed] [Google Scholar]
- 31.Klaunig JE, Wang Z, Pu X, Zhou S. Oxidative stress and oxidative damage in chemical carcinogenesis. Toxicol Appl Pharmacol. 2011;254:86–99. doi: 10.1016/j.taap.2009.11.028. [DOI] [PubMed] [Google Scholar]
- 32.Oberley TD, Oberley LW. Antioxidant enzyme levels in cancer. Histol Histopathol. 1997;12:525–35. [PubMed] [Google Scholar]
- 33.Yang LY, Chen WL, Lin JW, Lee SF, Lee CC, Hung TI, et al. Differential expression of antioxidant enzymes in various hepatocellular carcinoma cell lines. J Cell Biochem. 2005;96:622–31. doi: 10.1002/jcb.20541. [DOI] [PubMed] [Google Scholar]
- 34.Chan JH, Lim S, Wong WS. Antisense oligonucleotides: From design to therapeutic application. Clin Exp Pharmacol Physiol. 2006;33:533–40. doi: 10.1111/j.1440-1681.2006.04403.x. [DOI] [PubMed] [Google Scholar]
- 35.Dias N, Stein CA. Antisense oligonucleotides: Basic concepts and mechanisms. Mol Cancer Ther. 2002;1:347–55. [PubMed] [Google Scholar]
- 36.Agrawal S, Kandimalla ER. Antisense therapeutics: Is it as simple as complementary base recognition? Mol Med Today. 2000;6:72–81. doi: 10.1016/s1357-4310(99)01638-x. [DOI] [PubMed] [Google Scholar]