

Abstract
Background
This study aimed to develop a biocontrol system for ginseng root rot caused by Fusarium cf. incarnatum.
Methods
In total, 392 bacteria isolated from ginseng roots and various soils were screened for their antifungal activity against the fungal pathogen, and a bacterial isolate (B2-5) was selected as a promising candidate for the biocontrol because of the strong antagonistic activity of the bacterial cell suspension and culture filtrate against pathogen.
Results
The bacterial isolate B2-5 displayed an enhanced inhibitory activity against the pathogen mycelial growth with a temperature increase to 25°C, produced no pectinase (related to root rotting) and no critical rot symptoms at low [106 colony-forming units (CFU)/mL] and high (108 CFU/mL) inoculum concentrations. In pot experiments, pretreatment with the bacterial isolate in the presumed optimal time for disease control reduced disease severity significantly with a higher control efficacy at an inoculum concentration of 106 CFU/mL than at 108 CFU/mL. The establishment and colonization ability of the bacterial isolates on the ginseng rhizosphere appeared to be higher when both the bacterial isolate and the pathogen were coinoculated than when the bacterial isolate was inoculated alone, suggesting its target-oriented biocontrol activity against the pathogen. Scanning electron microscopy showed that the pathogen hyphae were twisted and shriveled by the bacterial treatment, which may be a symptom of direct damage by antifungal substances.
Conclusion
All of these results suggest that the bacterial isolate has good potential as a microbial agent for the biocontrol of the ginseng root rot caused by F. cf. incarnatum.
Keywords: Bacillus species, biological control, Fusarium cf. incarnatum, ginseng root rot, Panax ginseng Meyer
1. Introduction
Korean ginseng (Panax ginseng Meyer) is a perennial herbaceous and half-heliophobus plant in the family Araliaceae. It has been widely used as a highly valued medicinal plant not only for traditional herbal prescriptions for thousands of years [1], but also for the prevention and cure of cardiovascular diseases and chronic metabolic syndromes such as diabetes in modern times [2,3].
Ginseng should be grown in the same field soil for several years to produce quality raw roots of white and red ginseng. However, this cultivation practice makes ginseng vulnerable to attacks by a variety of soil-borne pathogens including fungi, bacteria, and nematodes [4–10]. Fungi are the major pathogens causing ginseng root diseases, among which Cylindrocarpon destructans (Zins.) Sholten (teleomorph: Nectria radicicola Gerlach & L. Nilsson) is one of the most important root-rot causing pathogens and the main cause of replanting problems in ginseng [10–13]. Other major fungal pathogens in ginseng are Fusarium species [14–16]. This was also noted in a survey of Fusarium pathogenicity to ginseng roots, which revealed the distribution of three dominant species (Fusarium solani, Fusarium oxysporum, and Fusarium moniliforme) and other minor species, although only a few were virulent to ginseng roots [5]. Fusarium species inhabit soils worldwide and are responsible for a variety of plant diseases; thus, there may be many other Fusarium species with the potential to induce ginseng root rot [17].
The control of fungal diseases relies mainly on the use of pesticides. However, pesticide use is not recommended for soil-borne diseases because of high costs and low control efficiencies. Furthermore, pesticides may be toxic to humans, animals, and crops, and might lead to the development of fungicide-tolerant pathogen strains [18,19]. The exclusion of toxic substances is particularly important for ginseng roots, which are used for health promotion. Biological control of soil-borne diseases using microorganisms (microbial fungicides) is an important alternative to the chemical control of plant diseases, offering a way to control pathogens efficiently with no or few harmful effects on humans, animals, or the environment [17].
In total, 14 microbial fungicides are commercially registered in Korea. These fungicides mainly contain Bacillus spp. that are primarily plant growth-promoting rhizobacteria [20,21] with demonstrated antifungal activity for controlling root rot in ginseng and other various crops [22,23]. The aim of this study was to evaluate the effectiveness of Bacillus species in the biological control of ginseng root rot caused by a Fusarium species that was newly isolated from cactus with rot symptoms. The results will aid in efforts to protect field-grown ginseng from root rot pathogens using biological control by antagonistic microorganisms.
2. Materials and methods
2.1. Isolation and characterization of the pathogen causing ginseng root rot
2.1.1. Pathogen isolation and inoculum preparation
The fungal pathogen used in this study was isolated from cactus stems with rot symptoms. For the pathogen isolation, cactus stem tissues with rot symptoms were excised and surface-disinfected in 1% NaOCl for 30 s and 70% ethanol for 30 s, and plated on water agar after rinsing in sterile distilled water (SDW). After 3 d of incubation at 25°C, hyphal tips grown out of the stem tissues were transferred to fresh potato–dextrose agar (PDA) and incubated at 25°C for 7 d to form pure fungal colonies. All isolates formed morphologically identical colonies and produced falcate or slightly curved macroconidia with multiple septa and hyaline microconidia, which are typical mycological characteristics of the genus Fusarium [24]. Among these colonies, a Fusarium isolate named CT4-1, which induced the most severe root rot, was selected and used for this study.
To develop the pathogen inoculum for ginseng root discs, Fusarium CT4-1 was cultured on carnation leaf agar (CLA) at 25°C for 10 d, and the macro- and mesoconidia that formed were diluted in SDW to make conidial suspensions at proper concentrations. To develop the pathogen inoculum for whole ginseng roots (pot experiments), the fungal culture was grown on PDA after mixing homogeneously with an oatmeal medium consisting of oatmeal (15 g), sand (300 g), and SDW (60 mL), and incubated at 25°C for 7 d. Prior to use, this inoculum was mixed with sterilized sandy soil, diluting them to the proper concentrations.
2.1.2. Pathogenicity test
Pathogenicity tests of the Fusarium isolate were conducted on root discs and whole 4-yr-old ginseng roots, using the pathogen inocula mentioned above. For the pathogenicity test on ginseng root discs, 20 μL of the conidial suspensions with inoculum concentrations of approximately 104 or 106 conidia/mL were inoculated on the center of 4-yr-old ginseng root discs approximately 0.5 cm thick with nine replications. These inoculated root discs were placed on filter paper soaked with SDW to maintain proper moisture in a plastic container and incubated at 25°C in an incubation chamber. Rot symptom development was examined daily up to 6 d after inoculation. The degree of rotting was scored based on the following disease severity rating system of 0, no rot; 1, 1–10%; 2, 10–30%; 3, 30–50%; 4, 50–70%; and 5, >70% (or fully) rotted, which was modified from the disease severity rating system for whole ginseng roots [25].
For the pathogenicity test of whole ginseng roots, fresh 4-yr-old ginseng roots planted in the oatmeal-sand medium were inoculated with 0%, 0.2%, 1.0%, and 5.0% pathogen inoculum and incubated at 21°C in 10 replicates. Rot symptom development on the ginseng roots was examined visually at 20 d after inoculation based on a disease severity scale of 0–5: 0,= no visible lesions; (1) brown lesions up to <1.0 mm; (2) dark brown lesions of 1.0 to <4.0 mm; (3) black lesions of 4.0 to <7.0 mm; (4) black lesions of ≥7.0 mm in diameter that coalesce with one another; and (5) mostly coalesced black lesions covering more than 70% of the surface (or fully rotted) [25].
2.1.3. Pathogen identification
The Fusarium isolate pathogenic to ginseng roots was grown on CLA and PDA and identified based on the mycological characteristics referred to the descriptions of the Fusarium Laboratory Manual [24]. For molecular identification of the Fusarium isolate, genomic DNA was extracted from the mycelia of the pure fungal culture obtained by single spore isolation using PrepMan Ultra Sample Preparation Reagent (Applied Biosystems, Foster City, CA, USA) [26]. The translation elongation factor-1α gene (EF-1α) was amplified through polymerase chain reaction using primers EF1/EF2, and nucleotide sequences were generated using BigDye terminator version 3.1 cycle sequencing kits (Applied Biosystems) and registered in GenBank as GenBank Accession No. KC478361. Molecular identification of the pathogen was accomplished by BLAST analysis of the gene sequences by comparing sequence similarities to others registered in GenBank.
2.2. Antifungal characteristics of bacterial isolates for the biocontrol of ginseng root rot
2.2.1. In vitro screening of antifungal bacteria against the Fusarium pathogen
To select antifungal bacteria against the Fusarium pathogen causing ginseng root rot, 392 bacteria were isolated from diseased ginseng roots and from mountain-, wetland-, and field-soils of various crops. For the dual culture tests, bacteria were grown in nutrient broth for 2 d, and 10 μL bacterial suspensions were spotted on three sections of the PDA. A mycelial plug (5 mm diameter) of the pathogen culture taken with a 5-mm-diameter cork-borer from the margin of a 7-d-old colony on the PDA was placed in the center of another PDA spotted with bacterial suspensions. After 1 wk of incubation, the pathogen mycelial growth of bacterial colonies (relative to the untreated control) was measured to determine the antifungal activity of the bacterial isolates. Three replications were used for each treatment.
2.2.2. Identification of antifungal bacterium selected by in vitro screening
One bacterial isolate (isolate B2-5) out of 392 that showed a strong antifungal activity was selected and identified based on Gram staining, bacterial morphology, carbon source assimilation, and 16S ribosomal RNA (rRNA) gene sequencing analysis. Gram staining of the bacterial cells was conducted following the Laboratory Guide for Identification of Plant Pathogenic Bacteria [27]. The bacterial morphology was examined under a transmission electron microscope (JEM-1010, JEOL Ltd., Tokyo, Japan) operating at an accelerating voltage of 80 kV after negative staining with 1.0% uranyl acetate. Carbon source assimilation of the bacterial isolate was examined in the Biolog GN test kit (Biolog Inc., Hayward, CA, USA). For 16S rRNA gene sequencing analysis, the bacterial isolate was cultured on BHI agar at 28°C for 2 d, and its genomic DNA was extracted from the colony using a FastDNA spin kit (MP Biomedicals, Santa Ana, CA, USA). To amplify the 16S rRNA gene, polymerase chain reaction was performed with the primers 27mF (5′-AGAGTTTGATCMTGGCTCAG-3′) and 1492mR (5′-GGYTACCTTGTTACGACTT-3′) [28,29]. The 16S rRNA gene sequences registered as GenBank Accession No. KC478362 were confirmed by a similarity search of GenBank using the Basic Local Alignment Search Tool (BLAST).
2.2.3. Antifungal activity of selected bacteria at different incubation temperatures
The fungal pathogen was cultured on PDA for 7 d, and 5-mm mycelial plugs were placed on the center of the PDA plates. Following this, 10 μL of the bacterial suspension grown in brain heart infusion (BHI) broth (CONDA, Madrid, Spain) at 28°C for 2 d was spotted 3 cm apart from the mycelial plugs on the media. These agar plates were incubated at different temperatures of 15°C, 18°C, 21°C, 25°C, and 28°C and the antifungal activity of the bacterial isolates was examined after 1 wk of incubation. SDW was used as an untreated control, and three replications were used for each treatment.
2.2.4. Effect of the bacterial suspension and cell-free culture filtrate on the conidial germination of the pathogen
The bacterial isolate was cultured in BHI broth at 28°C for 2 d. The bacterial culture was adjusted to concentrations of 106 colony-forming units (CFU)/mL and 108 CFU/mL for treatment. To obtain a cell-free culture filtrate, the bacterial culture was centrifuged at 5,162 g for 20 min and the supernatant was passed through a 0.22 μm Millipore filter (Millipore Corp., Cork, Ireland). Sterile paper discs (8 mm in diameter) soaked with 40 μL of bacterial suspension or culture filtrate were placed on PDA with approximately 106 conidia/mL plated and incubated at 25°C. After 2 d of incubation, the sizes of clear halos formed around the paper discs were measured to determine the inhibition of conidial germination.
To verify the germination rate of conidia, 1 mL of bacterial suspension at low and high concentrations (106 CFU/mL and 108 CFU/mL, respectively) was mixed with 1 mL of conidial suspension containing approximately 106 conidia/mL. Conidial germination was examined at intervals of 6 h and considered positive when the germ-tube length was longer than the nongerminated conidia. Germ-tube lengths were measured randomly up to 100 conidia under a compound light microscope with three replications.
2.2.5. Effect of inoculum concentrations on root rot development at different temperatures
The bacterial isolate selected in our study was grown in BHI broth and incubated at 28°C with 200 rpm in a shaking incubator. After incubation for 2 d, bacterial cell suspensions were adjusted to 106 CFU/mL or 108 CFU/mL. Three-yr-old ginseng roots were surface-disinfected with 70% ethanol and 1% sodium hypochlorite for 5 min each and rinsed twice with SDW. These roots were cut into discs of 0.5 cm in thickness and placed on filter paper soaked with SDW in 9-cm petri dishes with three replicates. Cell suspensions (20 μL) were spotted on the ginseng discs. Pure BHI broth was used as a control. Root discs placed on the dishes were incubated at temperatures of 18°C, 21°C, 25°C, and 28°C. Root discs were then examined for symptom development after 2 d of incubation, following an assessment of disease severity based on the rating system of 0, no rot; 1, 1–10%; 2, 10–30%; 3, 30–50%; 4, 50–70%; and 5, >70% (or fully) rotted as mentioned above.
2.2.6. Enzyme activities of antifungal bacteria
The antifungal bacteria were grown in 3 mL of BHI broth for 2 d at 28°C in a shaking incubator (with 200 rpm). The bacterial suspensions (106 CFU/mL and 108 CFU/mL) were spotted onto agar plates prepared as follows (/L): for starch hydrolysis: 0.6 g beef extract, 1 g peptone, 2 g starch azure and 15 g agar; for cellulase: 0.5 g NH4SO4, 0.5 g L-asparagine, 1 g KH2PO4, 0.2 g crystalline MgSO4, 0.1 g CaCl2, 0.5 g yeast extract, 10 g carboxyl methyl cellulose, and 20 g agar; for hemicellulase: 5 g gum guar, 5 g yeast extract, 4 g K2HPO4, 10 g casein, 0.0015 g crystal violet, and 18 g agar; and for pectinase: 10 g pectin, 2 g NaNO3, 0.5 g KCl, 1 g K2HPO4, 0.5 g MgSO4∙7H2O, 0.01 g FeSO4, and 20 g agar [30]. After 2 d of incubation at the different temperatures of 21°C, 25°C, and 28°C, the plates were stained according to the following: Gram's iodine solution for starch, 0.1% Congo red for cellulose, and saturated copper acetate for pectin [30]. The hemicellulose staining used crystal violet that was included in the medium during its preparation. The sizes of halos that formed around bacterial spots were measured for enzymatic activities after 2 d of incubation.
2.3. Biological control of F. cf. incarnatum on ginseng
2.3.1. Optimum application time for treatment of the antagonistic bacterium in suppressing root rot on ginseng root discs
Treatments were applied at three times for the control of root rot caused by the Fusarium isolate on 4-yr-old ginseng root discs: pretreatment (2 d prior to inoculation of the fungal pathogen), simultaneous with treatment, and post-treatment (2 d after inoculation). The antagonistic bacterium was cultured in BHI broth at 28°C for 48 h in a shaking incubator with 200 rpm and adjusted to the concentrations of 106 CFU/mL and 108 CFU/mL, respectively. The fungal pathogen was grown on CLA for 10 d and conidia were harvested by flooding 10-d-old cultures with SDW. The suspensions were centrifuged at 3,123 g for 10 min, the supernatant was discarded, and 2 mL of SDW were added to each conidial pellet. This process was repeated three times for washing, and the concentration of conidial suspensions was adjusted to about 106 conidia/mL by a hemacytometer. Ginseng root discs were treated with 100 μL of bacterial suspensions at the three timings: 2 d before (pretreatment), simultaneously (with treatment), and 2 d after (post-treatment) inoculation. For each treatment, 20 μL of conidial suspension were also inoculated following spotting of the discs with bacterial treatment, after which the discs were dried for 30 min on a clean bench. Inoculated ginseng discs were placed on water-soaked filter paper and incubated at 25°C. Rot development was measured daily up to 5 d after inoculation with the conidial suspension, based on the disease severity rating system mentioned above.
2.3.2. Biocontrol of ginseng root rot by the antifungal bacterium
The antifungal bacterium was grown in 250 mL BHI broth and incubated at 28°C in a shaking incubator. After incubation for 2 d, bacterial suspensions were adjusted to concentrations of 106 CFU/mL or 108 CFU/mL. Three-yr-old ginseng roots were surface-sterilized with 70% ethanol for 5 min and 1% NaOCl for 5 min. The roots were then rinsed twice with SDW. The sterilized ginseng roots were dipped in bacterial suspensions (106 CFU/mL and 108 CFU/mL) for 40 min and dried for 1 h on a clean bench [31]. The roots were transplanted into artificially infested soil in plastic pots with concentrations of 5% oatmeal-culture fungal inoculum and incubated at 25°C. Root rot symptoms were examined visually 10 d following inoculation.
2.3.3. Changes in population densities of the bacterium species on ginseng roots at different inoculum concentrations
Two concentrations of Bacillus broth cultures (106 CFU/mL and 108 CFU/mL) were used as treatment. Ginseng root discs were treated with 20 μL of the bacterial suspensions and were placed on moistened filter paper inside petri dishes and incubated at 25°C. There were three replicates of root discs for each treatment, and the experiment was performed twice. To measure cell population changes, the whole root discs treated and inoculated were ground in a blender and suspended in 10 mL SDW. The solution was then diluted with SDW, spread on BHI agar, and incubated at 28°C. After incubation of 20 h, the number of colonies formed on the agar plates was counted with the naked eye for the total bacterial population on the root discs. These were examined daily up to 7 d after incubation [32].
2.3.4. Scanning electron microscopic observation of root tissues
To prepare the samples for scanning electron microscopy (SEM), the bacterial isolates grown in BHI broth for 2 d were mixed with the Fusarium isolate and incubated on PDA at 25°C. One d after incubation, mycelial discs were fixed with modified Karnovsky's fixative [2% paraformaldehyde and 2% glutaraldehyde in 0.05M sodium cacodylate buffer (pH 7.2)] for 12 h at 4°C [33]. The fixed specimens were washed with 0.05M sodium cacodylate buffer three times for 10 min each. These were postfixed in 1% OsO4 at 4°C for 2 h, and briefly washed with distilled water. The specimens were then dehydrated in an ethanol series of 30%, 50%, 70%, 80%, and 90% for 10 min each, and in 100% ethanol three times for 10 min each [33]. Using hexamethyldisilazane (Electron Microscopy Sciences, Hatfield, PA, USA), specimens were dried and coated with gold using a sputter coater (MSC-101, JEOL). The specimens were observed under a field emission SEM (Auriga, Zeiss, Berlin, Germany) at an acceleration voltage of 5.0 kV.
3. Results
3.1. Pathogen isolation and identification
The fungal isolate C4-1 obtained from the rotten cactus stem had all the same mycological characteristics of Fusarium species and formed multicellular falcate macroconidia. Morphological characteristics of the fungal isolate were as follows: extensive and cotton-like mycelia with a colony color of pale orange or yellowish brown on PDA; macroconidia produced from polyphialides on CLA, slightly curved, frequently 3–5 septate, with a curved and tapering apical cell and a foot-shaped basal cell, measuring 37.9 ± 4.3 μm × 4.2 ± 0.5 μm; mesoconidia, which were fusoid, 1–5 septate, measuring 20.2 ± 4.3 μm × 3.7 ± 0.7 μm; intercalary chlamydospores; and absent microconidia (Table 1, Fig. 1), indicating that they are matched well with the F. cf. incarnatum previously reported by Akino and Kondo [34]. For molecular analysis, the DNA sequences of the translation elongation factor-1α (EF-1α), which amplified using primers EF1/EF2 (GenBank Accession No. KC478361), also had 100% sequence identity to F. cf. incarnatum strains (GenBank Accession No. JF270205 and GQ339786) (data not shown), confirming it to be F. cf. incarnatum as shown by the above mycological characteristics.
Table 1.
Mycological Characteristics of the Present Fusarium Isolate Causing Ginseng Root Rot
| Characteristics |
Present isolate |
Fusarium cf. incarnatum1) |
|---|---|---|
| Colony color | Orange, yellowish brown | Deep olive-buff to tan |
| Macroconidia | ||
| Shape | Slightly curved | Straight to slightly curved |
| Apical | Curved and tapering to a point | Curved |
| Basal | Foot shaped | Foot shaped or notched |
| Septum | 3–5 | 3–5, mostly 3 |
| Size (μm) | 37.9 ± 4.3 × 4.2 ± 0.5 | 31.5 ± 3.3 × 5.5 ± 0.3 |
| Mesoconidia | ||
| Shape | Fusoid | Fusoid |
| Septum | 1–5 | 1–2 |
| Size (μm) | 20.2 ± 4.3 × 3.7 ± 0.7 | 18.1 ± 3.4 × 3.0 ± 3.0 |
Akino and Kondo (2012) [34].
Fig. 1.
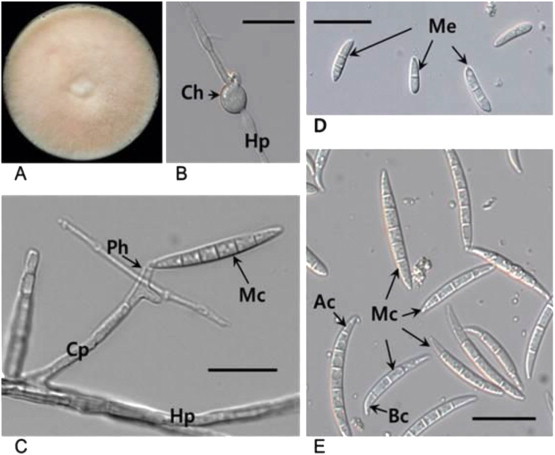
Phenotypic characteristics of Fusarium cf. incarnatum. (A) The cotton-like fungal colony formed on PDA. (B) An intercalary chlamydospore (Ch) in hypha (Hp). (C) Conidiophore (CP) and polyphialide (Ph) producing macroconidium (Mc). (D) Fusoid mesoconidia (Me), and macroconidia (Mc) slightly curved, frequently 3–5 septate, with a curved and tapering apical cell (Ac) and a foot-shaped basal cell (Bc). Bars, 20 μm.
3.2. Pathogenicity of the fungal isolate on ginseng
In the pathogenicity tests with different conidial inoculum concentrations of the Fusarium isolate obtained from diseased cactus, the initial rot symptoms appeared on the root discs inoculated with 106 conidia/mL after 2 d. After 6 d of incubation, rot symptoms developed on whole root discs at a high conidial concentration (106 conidia/mL) of the fungal isolate CT4-1 (Fig. 2). The root discs inoculated with 104 conidia/mL of the fungal isolate rarely showed rot symptom development, only slight discoloration during 6 d of incubation, and no symptoms were observed in the non-inoculated control. In the pot experiment, severe root rot also developed in ginseng roots inoculated with the fungal isolate C4-1 at inoculum concentrations of 1% and 5%; however, only mild and no rot symptoms were induced by the fungus with 0.2% inoculum concentration and the noninoculated control, respectively (Fig. 2).
Fig. 2.
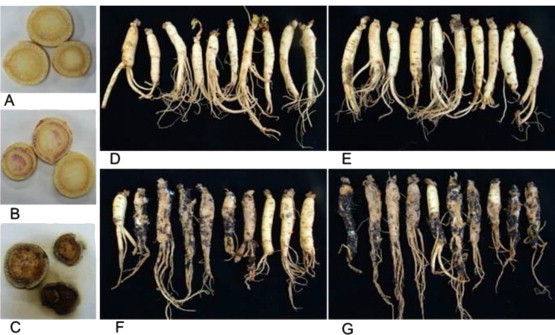
Rot symptom development on root discs. (A) Control (no rot). (B) Inoculated with 104 conidia/mL (slight discoloration). (C) Inoculated with 106 conidia/mL (severe rot) of the fungus after 6 d of incubation, and (D) on whole roots in the control (no rot). Inoculated with the fungus at inoculum concentrations of (E) 0.2% (mild rot symptoms), (F) 1%, and (G) 5% (severe rot symptoms) in the pot experiment.
3.3. Isolation of antifungal bacteria against F. cf. incarnatum
In total, 392 microbial isolates obtained from various areas including rotten ginseng roots, crop fields, and mountain areas were screened for antifungal activity against F. cf. incarnatum C4-1, among which 10 bacterial isolates were selected as potential antagonists. These antagonistic bacteria and two additional bacterial isolates with no antifungal activity (for comparison) were screened again for antifungal activity against the fungal pathogen using a dual culture method. Among the tested bacterial isolates, B2-5 and B8 most inhibited the pathogen's mycelial growth (Fig. 3). The isolate B2-5 was selected and used for further biocontrol studies because B8 had a phytotoxic effect on the ginseng root tissues (data not shown).
Fig. 3.
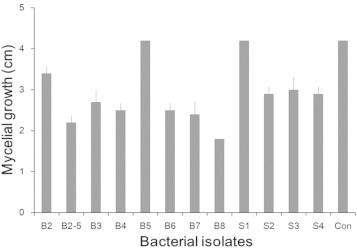
Mycelial growth of Fusarium cf. incarnatum after 7 d of incubation with bacterial isolates, showing that B2-5 and B8 had the strongest antifungal activity among the 12 bacterial isolates examined. Con: no treatment control. Bars and vertical lines are averages and standard deviations of three replications.
3.4. Identification of the antagonistic bacterial isolate
The bacterial isolate B2-5 was Gram-positive, rod-shaped, and bacillus-like with peritrichous flagella (Fig. 4), showing the typical characteristics of Bacillus species as in a previous study [33]. Biological analysis showed that the isolate B2-5 utilized 24 carbon sources including sorbitol, but did not utilize 25 carbon sources including D-arabinose, revealing a 96.6% similarity to Bacillus subtilis and Bacillus amyloliquefaciens (data not shown). The 16S rRNA gene sequences of B2-5 (GenBank Accession No. KC478362) were found to have the highest similarity to B. amyloliquefaciens subsp. plantarum (NCBI Accession No. CP000560) of 99.80% (data not shown). Therefore, the bacterial isolate B2-5 was identified as B. amyloliquefaciens subsp. plantarum.
Fig. 4.
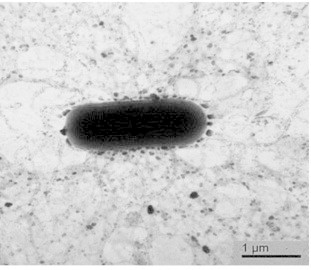
Electron micrograph of the bacterial isolate B2-5, showing rod-shaped morphology with numerous peritrichous flagella. Bar, 1 μm.
3.5. Antifungal activity of the bacterial isolate B2-5 at different incubation temperatures
The effects of the bacterial isolate on antifungal activity against the pathogen were tested at temperatures of 15°C, 18°C, 21°C, 25°C, and 28°C. Mycelial growth increased with temperature increase up to 25°C and decreased slightly at 28°C. The antifungal activity of isolate B2-5 was the highest at 25°C with an inhibition rate of 85.5%, which was slightly lowered at 28°C with an inhibition rate of 67.6%, and decreased with the decrease of the incubation temperature (Table 2).
Table 2.
Effect of Incubation Temperature on the Mycelial Growth of F. cf. Incarnatum Affected by the Bacterial Treatment
| Treatment | Mycelial growth (mm) of F. cf. incarnatum |
||||
|---|---|---|---|---|---|
| Incubation temperatures (°C) | |||||
| 15 | 18 | 21 | 25 | 28 | |
| Control | 18.3 ± 0.6 | 23.0 ± 0.0 | 32.0 ± 1.0 | 43.0 ± 0.0 | 41.0 ± 1.0 |
| Treatment (B2-5) | 13.3 ± 1.9 (27.3)1) | 13.0 ± 2.5 (43.5) | 8.4 ± 2.0 (73.6) | 6.2 ± 1.1 (85.5) | 13.3 ± 1.6 (67.6) |
Inhibition rates (%) of mycelial growth compared to untreated controls at the same temperature.
3.6. Effect of bacterial suspensions and cell-free culture filtrates on conidial germination
Treatment of the bacterial culture filtrate and cell suspensions with inoculum concentrations of 106 CFU/mL and 108 CFU/mL inhibited the conidial germination significantly to form clear inhibition zones around the treated paper discs, showing smaller inhibition zones with the culture filtrate than with the bacterial suspensions and no significant difference between the two inoculum concentrations (Fig. 5A). The conidial germination rates were significantly reduced in the bacterial treatments, compared to the untreated control, and by a smaller degree in the culture filtrate than in the bacterial suspensions. No significant difference was found between low and high inoculum concentrations of the bacterial cell suspension (Fig. 5B).
Fig. 5.
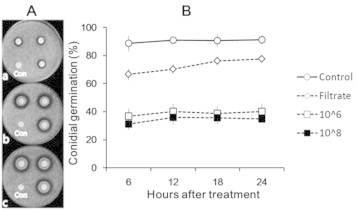
Effects of the bacterial isolate B2-5 on the inhibition of conidial germination (A) on solid medium and (B) in liquid medium, showing clear inhibition zones around the paper discs treated with (a) cell-free culture filtrate (Filtrate) and bacterial cell suspensions of (b) 106 CFU/mL (10^6) and (c) 108 CFU/mL (10^8), respectively, in (A), and conidial germination rates in bacterial treatments in (B). Marks and vertical lines in (B) are averages and standard deviations of three replications. Con, control.
3.7. Effect of bacterial inoculum concentrations on root rot development at different temperatures
No rot symptoms were produced on ginseng root discs inoculated with bacterial cell suspensions at two different inoculum concentrations of 106 CFU/mL and 108 CFU/mL at 18°C and 21°C, or at the inoculum concentration of 106 CFU/mL in the untreated control at 25°C and 28°C (Fig. 6). However, brownish discoloration or mild rot symptoms were produced on ginseng root discs treated with the high inoculum concentration of 108 CFU/mL and at the higher temperatures of 25°C and 28°C.
Fig. 6.
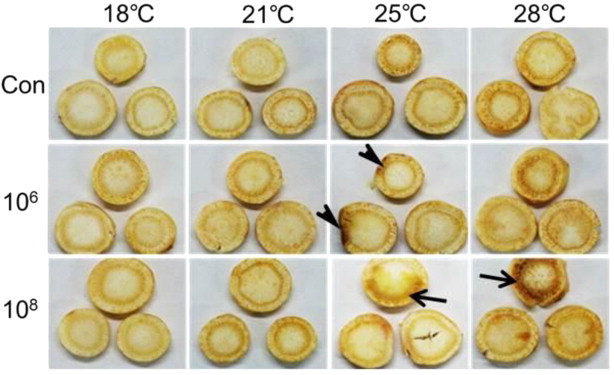
Effects of the bacterial isolate B2-5 with different inoculum concentrations [0 (Con), 106 CFU/mL (10ˆ6)] and temperatures on rot symptom development on ginseng root discs, for which the degrees of rotting are 0, no rot; 1, 1–10%; 2, 10–30%; 3, 30–50%; 4, 50–70%; and 5, >70% (or fully) rotted. Mild rot symptoms (arrows) developed only at a high inoculum concentration (108 CFU/mL) and at 25°C and 28°C. Arrowheads denote nonspecific decay by wounding.
3.8. Enzyme activities of antifungal bacteria
Pectinase activity responsible for bacterial soft rots was not detected in the bacterial isolate B2-5 at any of the temperature conditions or inoculum concentrations used, whereas other enzyme activities such as starch hydrolysis, cellulase, and hemicellulase were detected in the bacterial isolate with no significant differences in the degree of enzyme activity among incubation temperatures and inoculum concentrations (Table 3) [30]. The degrees of hemicellulase activity were higher than those of other enzymes, regardless of incubation temperature and inoculum concentration.
Table 3.
Effect of Incubation Temperature and Inoculum Concentrations of the Bacterial Isolate B2-5 on Selected Enzyme Activities
| Enzyme | Inoculum conc. (CFU/mL) | Degree of enzyme activity1) at |
||
|---|---|---|---|---|
| 21°C | 25°C | 28°C | ||
| Starch hydrolysis | 106 | + | + | + |
| 108 | ± | + | + | |
| Cellulase | 106 | ± | ± | ± |
| 108 | ± | ± | ± | |
| Hemicellulase | 106 | ++ | ++ | ++ |
| 108 | ++ | ++ | ++ | |
| Pectinase | 106 | − | − | − |
| 108 | − | − | − | |
Note. Data from “Involvement of growth-promoting rhizobacterium Paenibacillus polymyxa in root rot of stored Korean ginseng,” by Y.H. Jeon, S.P. Chang, I. Hwang, and Y.H. Kim, 2003, J Microbiol Biotechnol, 13, p. 881–91. Copyright 2003, Metadata. Reprinted with permission.
Size of halos formed around bacterial colonies on agar (degree of enzyme activity): ++, larger than 5 mm (highly active); +, 3–5 mm (medially active); ±, 1–3 mm (weakly active); and −, no halo formed (not active) [30].
3.9. Biological control of F. cf. incarnatum on ginseng
3.9.1. Optimal treatment time of bacterial isolate B2-5 for the inhibition of root rot on ginseng root discs
Among the three application times (pre-, simultaneous-, and post-treatment), the pretreatment (2 d prior to pathogen inoculation) was the most effective for the treatment of bacterial isolate B2-5, showing the appearance of only mild rot symptoms with a disease severity index of <2.0 for both inoculum concentrations with inhibition rates over 50%, whereas the simultaneous- and post-treatments showed lowered inhibition rates below 40% and 32%, respectively (Fig. 7). For all treatment times, the bacterial isolate B2-5 with low inoculum concentration of 106 CFU/mL inhibited rot symptom development somewhat more than with a high inoculum concentration of 108 CFU/mL.
Fig. 7.
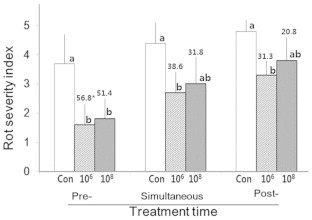
Effects of treatment times (pre-, simultaneous, and post-treatment) of the bacterial isolate B2-5 with different inoculum concentrations [0 (Con), 106 CFU/mL (106), and 108 CFU/mL (108)] on the inhibition of rot symptom development on ginseng root discs inoculated with Fusarium cf. incarnatum at 5 d after inoculation. Bars and vertical lines are means and standard deviations of nine replications. * Figures are inhibition rates (%) of rot symptom development relative to the control at the same treatment time. The same letters above the bars denote no significant difference within the same treatment time at p < 0.05 by the least significant different (LSD) test.
3.9.2. Biological control of ginseng root rot by the antifungal bacterium
In the pot experiment under greenhouse conditions, the ginseng roots inoculated with F. cf. incarnatum alone were severely rotten with a severity index of 4.8, whereas treatment with the bacterial isolate B2-5 significantly reduced root rot disease development on the ginseng roots with degrees of control efficacy of 70.0% and 55.8% for the inoculum concentrations of 106 CFU/mL and 108 CFU/mL, respectively (Table 4 [25], Fig. 8).
Table 4.
Effect of Bacterial Treatments with Different Inoculum Concentrations on the Control of Ginseng Root Rot Caused by F. cf. Incarnatum
| Treatment | Inoculum conc. (CFU/mL) | Rot severity1) | Control efficacy (%)2) |
|---|---|---|---|
| Control | 4.8 ± 0.6* | 0.0 | |
| Treatment (B2-5) | 106 | 2.3 ± 1.6** | 70.0 |
| 108 | 2.9 ± 1.4** | 55.8 |
*,** The same asterisks in a column denote no significant difference at p < 0.05 by the least significant difference test.
Rot severity was examined 10 d after inoculation by scoring indices as 0, no symptoms; 1, 1–10%; 2, 10–30%; 3, 30–50%; 4, 50–70%; 5, >70% [25].
Control efficacy (%) = (rot severity of control − rot severity of treatment)/(rot severity of control) × 100.
Fig. 8.

Effects of bacterial treatment with inoculum on rot symptom development in ginseng roots caused by Fusarium cf. incarnatum at 10 d after pathogen inoculation. (A) Pathogen inoculation only with no bacterial treatment. Concentrations of (B) 106 CFU/mL and (C) 108 CFU/mL. (D) no bacterial treatment and pathogen inoculation.
3.9.3. Bacterial population changes on ginseng root discs at different initial inoculum concentrations
When the bacterial isolate B2-5 was given alone to ginseng root discs with no pathogen inoculation, the bacterial population densities from a high inoculum concentration of 108 CFU/mL decreased slowly, maintaining more than half of the initial population density until 7 d after inoculation, whereas those from the low initial inoculum concentration of 106 CFU/mL decreased rapidly to be nondetectable after 5 d following inoculation (Fig. 9). By contrast, the bacterial population densities on ginseng root discs inoculated with F. cf. incarnatum increased for 4–5 d after inoculation, regardless of the initial inoculum concentrations, maintaining the initial inoculum concentration of 108 CFU/mL when treated with high inoculum concentration, but decreased thereafter to be eventually nondetectable when treated with low inoculum concentration (Fig. 9).
Fig. 9.
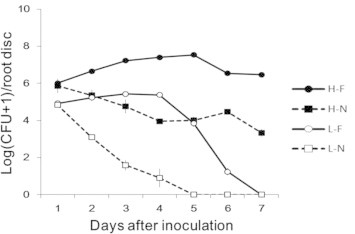
Population changes of the bacterial isolate B2-5 from low (L-; 106 CFU/mL) and high (H-; 108 CFU/mL) initial inoculum on ginseng root discs with (-F) and without (-N) inoculation of Fusarium cf. incarnatum. Marks and vertical lines are averages and standard deviations of nine replications.
3.9.4. SEM observation
SEM observations of Fusarium cf. incarnatum treated with the bacterial isolate B2-5 at inoculum concentrations of 106 CFU/mL and 108 CFU/mL showed the pathogen hyphae to be wrinkled, distorted, and shrunken (Fig. 10). Hyphae had bacterial cells adhering on some portions to varying degrees, which increased in number in the treatments with the higher inoculum concentration of 108 CFU/mL. Conversely, in the untreated control, pathogen hyphae looked intact with a smooth surface, sometimes showing a contour of the septum with no bacterial cells present in the untreated control (Fig. 10).
Fig. 10.

Scanning electron micrographs of Fusarium cf. incarnatum with no bacterial treatment (A), and treated with the bacterial isolate B2-5 at inoculum concentrations of (B) 106 CFU/mL and (C) 108 CFU/mL, showing wrinkled, distorted, and shrunken pathogen hyphae (Hy) adhered with bacterial cells (Ba), compared to intact hyphae (Hy) with smooth surface showing a contour of septum (St) in (A) the untreated control. Bars, 20 μm.
4. Discussion
Fusarium species are ubiquitous in soil, and these unspecialized parasites have a wide host range and can cause diseases in plants, humans, and domesticated animals [17,24]. Fusarium species such as F. solani, Fusarium equiseti, and Fusarium avenaceum have previously been reported as causal pathogens of ginseng diseases including root rots, seedling rots, and decayed seed [14,16].
F. cf. incarnatum, also known by the synonyms F. pallidoroseum and F. semitectum, is often regarded as a secondary colonizer of plant tissues and causes several plant diseases including pod and collar rot in soybeans [35], soybean root rot [36], and postharvest fruit rot in oriental melon [37]. It produces apicidins phytotoxic to seedlings and 2-wk-old plants of diverse species [38] and is one of 11 pathogenic Fusarium species listed as quarantine pests in Korea [39]. In addition, it has also been isolated from rotten ginseng roots [5]. Therefore, F. cf. incarnatum may be a potential cause of ginseng root rot of its strong pathogenicity for ginseng root rots as shown in this study.
In our study, in vitro and in vivo experiments showed that disease severity increased with an increase in the amount of inoculum tested. The same fungus was re-isolated from the diseased root tissues, confirming that the present Fusarium species is the true pathogen of the disease according to Koch’s postulates [17]. Considering its disease-inducing nature and capacity, F. cf. incarnatum may have potentials to become an important causal agent of ginseng root rot.
Bacillus species are usually found in diverse natural environments of soil, water, and air and have antifungal effects against several kinds of plant fungal pathogens [21,23,40]. They also show controlling capacities for root rots and Phytophthora blight of ginseng caused by Cylindrocarpon destructans and Phytophthora cactorum, respectively [22,33]. In our study, a bacterial isolate identified as B. amyloliquefaciens B2-5 had a strong antagonistic activity against the causal pathogen of ginseng root rot, F. cf. incarnatum, showing strong inhibitory activity against mycelial growth and conidial germination that play important roles in the infection cycle of the pathogen [17]. These attributes may make the bacterium useful for controlling the ginseng root rot caused by this fungal pathogen. The bacterial isolate B2-5 had the highest control efficiency of ginseng root rot caused by F. cf. incarnatum when it was applied 2 d prior to pathogen inoculation (by pretreatment); significantly lowered control efficacies were observed in the simultaneous treatment and post-treatment. This suggests that the proper application time of the bacterial isolate may be any time prior to the disease occurrence as Bacillus spp. are durable in harsh environments due to endospore formation [41], which may be an advantage for easy formulation of the bacterial isolate for the commercialization of microbial fungicidal products.
The mycelial growth of F. cf. incarnatum increased with temperature increase; however, the antagonistic activity of the bacterial isolate to the pathogen was enhanced much more than the fungal growth increase with a temperature increase up to 25°C, at which temperature the growth of the pathogen treated with antagonistic bacterium was reduced the most. This suggests that the antagonistic bacterium may exert its full disease-control capacity at a range of optimum temperatures in controlling the growths of the fungal pathogen and the half-heliophobus ginseng plant, and accordingly may lead to improved efficacy for the control of the root rot caused by F. cf. incarnatum. The inhibition of the conidial germination by the bacterial culture filtrate and the hyphal damages with no noticeable parasitism following the bacterial treatment as viewed by microscopy, suggest that bacterial antibiotics and other toxic compounds present in bacterial metabolites or a direct interaction might be responsible for the inhibition of the pathogen growth, for which antibiosis is the major action mode that exhibits instant disease control effects [42].
One of the important constraints in the biological control of plant diseases using beneficial microbes is the need for massive introduction of biological control agents into field soils, which may be achieved by increasing their concentrations and/or volumes (masses) [43]. In our study, however, B. amyloliquefaciens B2-5 reduced rot symptom development at the lower inoculum concentration (106 CFU/mL) with somewhat more prominent control efficacies than at the higher one (108 CFU/mL; Fig. 7). This finding may be derived from there being no difference in the inhibition of the fungal conidial germination and equivalent fungal damages, as viewed in microscopy, between the inoculum concentrations and phytotoxicity to ginseng root tissues at the higher inoculum concentration. Also the bacterial population increased initially and was maintained for a certain period of time on the ginseng root tissues inoculated with the pathogen in spite of its rapid decrease on the root tissues with no pathogen inoculation. These aspects suggest higher efficacy of the disease control at the lower inoculum concentrations than at higher ones, which may make the effective control of the disease possible by bacterial treatment with a relatively low inoculum concentration.
Bacillus amyloliquefaciens B2-5 produced no pectinase at any temperature or at high inoculum concentrations in our study, even though it is the major enzyme responsible for tissue rots (or soft rots) in various crops caused by pectinase-producing bacteria such as Pectobacterium carotovorum subsp. carotovorum [17]; this indicates that this bacterium is not a true root-rotting pathogen. The phytotoxicity of the bacterial isolate B2-5 to ginseng roots appearsed to be lower than that of previously studied Bacillus (Paenibacillus) species, although it induced definite rot symptoms on ginseng root tissues at high inoculum concentration (108 CFU/mL) and all species produced starch hydrolytic enzyme associated with ginseng root rot to some extent [33,41]. Bacillus and relatives are plant growth-promoting rhizobacteria that can have beneficial effects on plant growth [44], as proven by their control of a complex disease caused by the root-knot nematode and fusarium wilt fungus [45]. The results of this study indicate that Bacillus amyloliquefaciens B2-5 has great potential as an efficient biocontrol agent for managing ginseng root rot caused by F. cf incarnatum.
Footnotes
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
References
- 1.Park H.J., Kim D.H., Park S.J., Kim J.M., Ryu J.H. Ginseng in traditional herbal prescriptions. J Ginseng Res. 2012;36:225–241. doi: 10.5142/jgr.2012.36.3.225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Kim J.H. Cardiovascular diseases and Panax ginseng: a review on molecular mechanisms and medical applications. J Ginseng Res. 2012;36:16–26. doi: 10.5142/jgr.2012.36.1.16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Yuan H.D., Kim J.T., Kim S.H., Chung S.H. Ginseng and diabetes: the evidences from in vitro, animal and human studies. J Ginseng Res. 2012;36:27–39. doi: 10.5142/jgr.2012.36.1.27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Kim J.H., Jeon Y.H., Park H., Lee B.D., Cho D.H., Park B.Y., Khan Z., Kim Y.H. The root-lesion nematode, Pratylenchus subpenetrans, on ginseng (Panax ginseng) in Korea. Nematology. 2006;8:637–639. [Google Scholar]
- 5.Lee S.K. Fusarium species associated with ginseng (Panax ginseng) and their role in the root-rot of ginseng plants. Res Plant Dis. 2004;10:248–259. [Google Scholar]
- 6.Lee J.H., Yu Y.H., Kim Y.H., Ohh S.H., Park W.M. Morphological characteristics and pathogenicity of Alternaria isolates causing leaf and stem blights and black root rot of Korea ginseng. Kor J Plant Pathol. 1990;6:13–20. [Google Scholar]
- 7.Jeon Y.H., Park H., Lee B.D., Yu Y.H., Chang S.P., Kim S.G., Hwang I., Kim Y.H. First description of crown gall disease on ginseng. Plant Pathol J. 2008;24:207–210. [Google Scholar]
- 8.Ohh S.H., Lee S.K., Lee J.H., Han S.C. New root rot disease of Panax ginseng due to Ditylenchus destructor Thorne. Kor J Plant Prot. 1983;22:181–185. [Google Scholar]
- 9.Ohh S.H., Yu Y.H., Cho D.H., Lee J.H., Kim Y.H. Effect of chemical treatments on population changes of Ditylenchus destructor and responses of Panax ginseng. Kor J Plant Prot. 1986;25:169–173. [Google Scholar]
- 10.Yu Y.H., Ohh S.H. Research on ginseng diseases in Korea. Kor J Ginseng Sci. 1993;17:61–68. [Google Scholar]
- 11.Chung H.S. Studies on Cylindrocarpon destructans (Zins.) Scholten causing root rot of ginseng. Rep Tottori Mycol Inst. 1975;12:127–138. [Google Scholar]
- 12.Park K.J. Fitness analysis of the forecasting model for root rot progress of ginseng based on bioassay and soil environmental factors. Res Plant Dis. 2001;7:20–24. [Google Scholar]
- 13.Yu Y.H. Root rot diseases of Panax ginseng and their control in Korea. Kor J Plant Pathol. 1987;3:318–319. [Google Scholar]
- 14.Punja Z.K. Fungal pathogens of American ginseng (Panax quinquefolium) in British Columbia. Can J Plant Pathol. 1997;19:301–306. [Google Scholar]
- 15.Punja Z.K., Wan A., Goswami R.S., Verma N., Rahman M., Barasubiye T., Seifert K.A., Lévesque C.A. Diversity of Fusarium species associated with discolored ginseng roots in British Columbia. Can J Plant Pathol. 2007;29:340–353. [Google Scholar]
- 16.Punja Z.K., Wan A., Goswami R. Root rot and distortion of ginseng seedling roots caused by Fusarium oxysporum. Can J Plant Pathol. 2008;30:565–574. [Google Scholar]
- 17.Agrios G.N. 5th ed. Elsevier Academic Press; San Diego: 2005. Plant pathology. [Google Scholar]
- 18.Haas D., Défago G. Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat Rev Microbiol. 2005;3:307–319. doi: 10.1038/nrmicro1129. [DOI] [PubMed] [Google Scholar]
- 19.Handelsman J., Stabb E.V. Biocontrol of soilborne plant pathogens. Plant Cell. 1996;8:1855–1869. doi: 10.1105/tpc.8.10.1855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kim S.G., Jang Y., Kim H.Y., Koh Y.J., Kim Y.H. Comparison of microbial fungicides in antagonistic activities related to the biological control of phytophthora blight in chili pepper caused by Phytophthora capsici. Plant Pathol J. 2010;26:340–345. [Google Scholar]
- 21.Kloepper J.W., Ryu C.M., Zhang S. Induced systemic resistance and promotion of plant growth by Bacillus spp. Phytopathology. 2004;94:1259–1266. doi: 10.1094/PHYTO.2004.94.11.1259. [DOI] [PubMed] [Google Scholar]
- 22.Kim C.H. Bacillus spp. as biocontrol agents of root rot and phytophthora blight on ginseng. Plant Pathol J. 2004;20:63–66. [Google Scholar]
- 23.Ryder M.H., Yan Z., Terrace T.E., Rovira A.D., Tang W., Correll R.L. Use of strains of Bacillus isolated in China to suppress take-all and Rhizoctonia root rot, and promote seedling growth of glasshouse-grown wheat in Australian soils. Soil Biol Biochem. 1998;31:19–29. [Google Scholar]
- 24.Leslie J.F., Summerell B.A. Blackwell Publishing Professional; Ames: 2006. The Fusarium laboratory manual. [Google Scholar]
- 25.Rahman M., Punja Z.K. Biochemistry of ginseng root tissues affected by rust root symptoms. Plant Physiol Biochem. 2005;43:1103–1114. doi: 10.1016/j.plaphy.2005.09.004. [DOI] [PubMed] [Google Scholar]
- 26.Yun H.Y., Rossman A.Y. Tubakia seoraksanensis, a new species from Korea. Mycotaxon. 2011;115:369–373. [Google Scholar]
- 27.Schaad N.W., Jones J.B., Chun W. 3rd ed. APS Press; St Paul: 2001. Laboratory guide for identification of plant pathogenic bacteria. [Google Scholar]
- 28.Brosius J., Palmet M.L., Kennedy P.J., Noller H.F. Complete nucleotide sequence of a 16S ribosomal RNA gene from Escherichia coli. Proc Natl Acad Sci USA. 1978;75:4801–4805. doi: 10.1073/pnas.75.10.4801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Weisburg W., Barns S.M., Pelletier D.A., Lane D.J. 16S ribosomal DNA amplification for phylogenetic study. J Bacteriol. 1991;173:697–703. doi: 10.1128/jb.173.2.697-703.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Jeon Y.H., Chang S.P., Hwang I., Kim Y.H. Involvement of growth-promoting rhizobacterium Paenibacillus polymyxa in root rot of stored Korean ginseng. J Microbiol Biotechnol. 2003;13:881–891. [Google Scholar]
- 31.Jang Y., Kim S.G., Kim Y.H. Biocontrol efficacies of Bacillus species against Cylindrocarpon destructans causing ginseng root rot. Plant Pathol J. 2011;27:333–341. [Google Scholar]
- 32.Jeon Y.H., Kim Y.H. Differential structural responses of ginseng root tissues to different initial inoculum levels of Paenibacillus polymyxa GBR-1. Plant Pathol J. 2008;24:352–356. [Google Scholar]
- 33.Karnovsky M.J. A formaldehyde-glutaraldehyde fixative of high osmolarity for use in electron microscopy. J Cell Biol. 1965;27:137A. [Google Scholar]
- 34.Akino S., Kondo N. Common spear rot of oil palm in Indonesia. Res Plant Dis. 2012;96:537–543. doi: 10.1094/PDIS-08-10-0569. [DOI] [PubMed] [Google Scholar]
- 35.Sinclari J.B., editor. Compendium of soybean diseases. APS Press; St Paul: 1982. [Google Scholar]
- 36.Wang D., Kurle J.E., Estevez De J.C., Percich J.A. Radiometric assessment of tillage and seed treatment effect on soybean root rot caused by Fusarium spp. in central Minnesota. Plant Soil. 2004;258:319–331. [Google Scholar]
- 37.Kim J.W., Kim H.J. Fusarium fruit rot of postharvest oriental melon (Cucumis melo L. var. makuwa Mak.) caused by Fusarium spp. Res Plant Dis. 2004;10:260–267. [Google Scholar]
- 38.Jin J., Baek S.R., Lee K.R., Lee J., Yun S.H., Kang S., Lee Y.W. Purification and phytotoxicity of apicidins by the Fusarium semitectum KCTT16676. Plant Pathol J. 2008;24:417–422. [Google Scholar]
- 39.Hong W.J., Kang M.R., Cho E.J., Kim H.K., Yun S.H. Specific PCR detection of four quarantine Fusarium species in Korea. Plant Pathol J. 2010;26:409–416. [Google Scholar]
- 40.Zhang J., Xue A., Tambong J. Evaluation of seed and soil treatments with novel Bacillus subtilis strains for control of soybean root rot caused by Fusarium oxysporum and F. graminearum. Plant Dis. 2009;93:1317–1323. doi: 10.1094/PDIS-93-12-1317. [DOI] [PubMed] [Google Scholar]
- 41.Jeon Y.H., Kim S.G., Hwang I., Kim Y.H. Effects of initial inoculation concentration of Paenibacillus polymyxa on colony formation and starch-hydrolytic activity in relation to root rot in ginseng. J Appl Microbiol. 2010;109:461–470. doi: 10.1111/j.1365-2672.2010.04674.x. [DOI] [PubMed] [Google Scholar]
- 42.Kim S.G., Zakaullah K., Jeon Y.H., Kim Y.H. Inhibitory effect of Paenibacillus polymyxa GBR-462 on Phytophthora capsici causing phytophthora blight in chili pepper. J Phytopathol. 2009;157:329–337. [Google Scholar]
- 43.Upadhyay R.S., Rai B. Biocontrol agents of plant pathogens: their use and practical constraints. In: Mukerji K.G., Garg K.L., editors. vol. 1. CRC Press Inc; Boca Raton: 1988. pp. 15–36. (Biocontrol of plant diseases). [Google Scholar]
- 44.Chanway C.P. Plant growth promotion by Bacillus and relatives. In: Berkeley R., Heyndricks M., Logan N., De Vos P., editors. Applications and systematics of Bacillus and relatives. Blackwell Science Ltd; Malden: 2002. pp. 219–235. [Google Scholar]
- 45.Son S.H., Khan Z., Kim S.G., Kim Y.H. Plant growth-promoting rhizobacteria, Paenibacillus polymyxa and Paenibacillus lentimorbus suppress disease complex caused by root-knot nematode and fusarium wilt fungus. J Appl Microbiol. 2009;107:524–532. doi: 10.1111/j.1365-2672.2009.04238.x. [DOI] [PubMed] [Google Scholar]