

Abstract
Pheromones are substances which are secreted to the outside by an individual and received by a second individual of the same species. Many examples exist in animals but their role in humans remains uncertain since adults have no functioning vomeronasal organ, which processes pheromone signals in animals. Yet pheromones can be detected by the olfactory system although humans under develop and underrate their smelling sense. Pheromones may be present in all bodily secretions but most attention has been geared toward axillary sweat which contains the odorous 16-androstenes. One of these steroidal compounds, androstadienone, is present at much higher concentrations in male sweat and can be detected by women, albeit with wide variation in sensitivity. Upper-lip application of a pharmacological dose of androstadienonein women results in improved mood and heightened focus - particularly to capture emotional information. A positive mood is known to facilitate women’s sexual response, and increased focus improves sexual satisfaction. Indeed, some studies showed a beneficial effect of androstadienone on sexual desire and arousal. However, these effects were dependent on the context of the experiment, for example, on the presence of a male attendant. Pheromones may also play a role in mate selection which is “disassortative” regarding the human leukocyte antigen (HLA)-genotype. Preliminary evidence suggests that exposure to androstadienone in women promotes attractiveness ratings of potential mates. In conclusion, some data indicate that 16-androstene pheromones, in particular androstadienone, play a beneficial role in women’s mood, focus and sexual response, and perhaps also in mate selection.
Keywords: Androstadienone, mate selection, pheromones, sexual response
Introduction: sexuality and the senses
The senses play a pivotal role in the sexual response of women. In Basson’s model, emotional intimacy with a partner may render women sexually aroused through specific stimuli, of which physical contact (the tactile sense) is pre-eminent (Basson, 2001). Bancroft et al. developed the dual-control (excitation/inhibition) model of the sexual response and emphasized its neurophysiologic substrate (Bancroft et al., 2009). In their model, tactile stimuli give excitatory (or inhibitory) input to the spinal cord, thereby mediating the genital arousal response.
Yet the other senses are also important. While tactile interaction outside a one-to-one relationship is frowned upon, no one in our image-obsessed world doubts the role of visual stimuli in the sexual response. Auditory stimuli, if less evident (eg, lounge music?), may equally act as sexual cues. But what about olfactory stimuli? While most people would agree that unpleasant smells can be strong inhibitory signals (in the Bancroft sense), it is unclear whether olfactory cues can act as excitatory sexual input.
Pheromones in animals and humans
The animal kingdom is full of examples of social and sexual chemosignals which have been given the name “pheromones” in 1959. They are defined as “substances which are secreted to the outside by an individual and received by a second individual of the same species, in which they release a specific reaction, for example, a definite behavior or a developmental process”.
Over time, four types of pheromones have been postulated (Table 1). Releaser pheromones often prompt an immediate behavioral reaction, and have received the greatest attention in animals. For example, boars exude androstenone (5α-androst-16-en-3α-one) which elicits lordosis (mating readiness) in sows during their estrus period (Dorries et al., 1997); therefore, androstenone is used to assist artificial insemination in the porcine species.
Table I. Categories of pheromones.
| Name | Effect |
| Releaser | Elicit an often immediate, specific, behavioral response |
| Signaler | Provide information regarding the individual |
| Modulator | Affect mood and emotion |
| Primer | Affect over time endocrine or neuroendocrine systems related to reproductive physiology or development |
Signaler pheromones are mainly social chemosignals: they provide information on the hierarchy and the animal’s place in it, and on the type of food recently consumed by other animals and thus the nearby availability of food. They are also important for mate selection.
Modulator pheromones influence neuropsychological parameters. While these effects are difficult to gauge in animals, they are the focus of most human studies. Finally, primer pheromones affect (neuro)-endocrine parameters, including the menstrual cycle. Since endocrine changes generally modulate neuropsychological parameters, modulator and primer pheromones likely overlap.
Pheromones in humans may be present in bodily secretions such as urine, semen or vaginal secretions, breast milk and potentially also saliva and breath, yet most attention thus far has been directed toward axillary sweat. The axillary secretions originate from the highly dense eccrine and apocrine (which secrete into hair follicles) sweat glands and sebaceous glands. The secretions are odorless but apocrine sweat acquires odor after interaction with the cutaneous bacterial microflora. Odor dispersal is modulated by several factors including clothing layers, axillary temperature, total hair surface, arm movements and the proximity of one’s nose. The main components of sweat are simple organic acids (eg, E-3-methyl-2-hexenoic acid and 3-methyl-3-hydroxylhexanoic acid) (Hays, 2003).
The putative axillary sweat pheromones are steroid-structure components, in particular the odorous 16-androstenes: androstadienone (4,16-androstadien-3α-one), androstenone (mentioned above) and androstenol (5α-androst-16-en-3-ol) (Fig. 1). Of these steroids, androstadienone appears to have the most robust effects upon both sexes but particularly upon women (Jacob et al., 2001a). The concentration of 16-androstenes is much higher in male compared to female axillary sweat. The sex gradient may be the result of the threefold difference in plasma levels as well as different skin flora. Although these steroids are present at concentrations 50-100 times below those of some organic acids mentioned previously, their effect ultimately depends on the volatility and the perceptive threshold for these compounds as well as the duration of exposure. Women tend to be more sensitive to androstadienone than men, and there appears to be a subpopulation of individuals with a high sensitivity (“super-smellers”) (Lundström et al., 2003b).
Fig. 1. The main putative human pheromones: androstenone, androstenol, androstadienone, and estratetraenol.

A putative pheromone secreted particularly by women is estratetraenol (estra-1,3,5(10),16-tetraen-3-ol, Fig. 1) which was first isolated from the urine of pregnant women (Thysen et al., 1968). This chemosignal is likely present in other secretions such as axillary sweat. In general, as we will see, the effects of estratetraenol are smaller than those of androstadienone but go in the same direction. While in theory a pheromone targeted at women’s partners, estratetraenol also has discernible effects in heterosexual women (Jacob et al., 2001a; Bensafi et al., 2004a).
In small mammals, the presence of a lactating dam within the same cage induces maternal behavior and lengthens the cycle of the non-pregnant females (reviewed in Spencer et al., 2004). Oxytocin is, of course, the first compound that comes to mind. This hormone is important for parent-infant bonding and human attachment in general, and is therefore known as a “pro-social hormone”. Oxytocin is readily detected in plasma but is also present, albeit at low concentrations, in urine and saliva (Feldman et al., 2011). The hormone is released bytactile interaction (suckling but also, for example, massage) and within-family conversation – “auditory massage”, so to speak. Whether oxytocin is released by olfactory interaction is unknown.
Vomeronasal, nasal or transdermal?
Non-human mammals and non-mammal animal species like reptiles and amphibians exhibit a specific tissue that detects most pheromones: the vomeronasal organ (VNO), situated in the rostral (anterior) nasal cavity. The VNO contains bipolar receptor cells, and its axonal nerve projections traverse the nasal septum and cross the cribriform plate to terminate in the accessory olfactory bulb, adjacent to - yet unconnected with- the main olfactory bulb (Fig. 2).
Fig. 2. The putative vomeronasal organ (VNO) in humans. There is no evidence that the VNO is still functioning in human adults. Figure reproduced from Monti-Bloch et al., 1998.
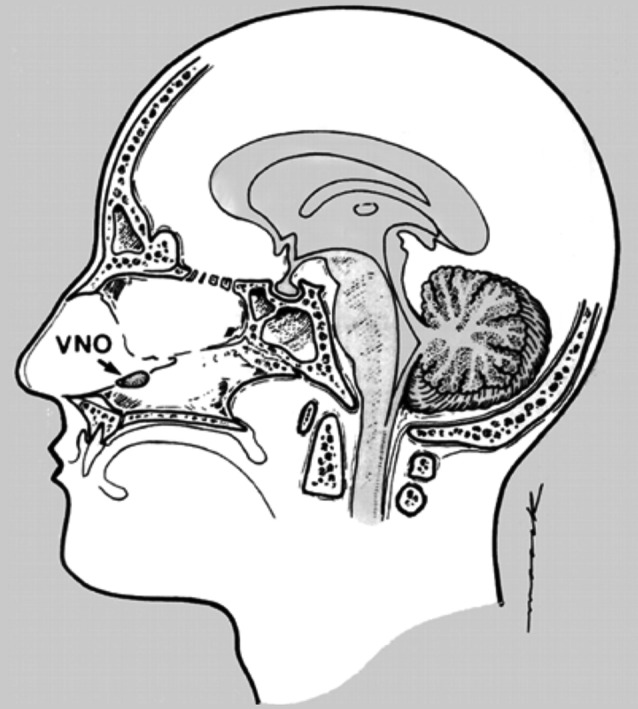
While the human fetus shows a VNO with the typical bipolar cells and axonal projections, most evidence shows that it is functionally inactive after birth. Genes coding for receptor proteins are pseudogenes with no protein expression; bipolar receptor cells are no longer detectable, and the accessory olfactory bulb is absent. Interestingly, GnRH-neurons – pivotal for pubertal development and reproduction – replace the degenerated VNO-nerves as they migrate from the fetal VNO-area to the basal forebrain (Wysocki and Preti, 2004).
However, some pheromones in non-human mammals are detected by the “regular” olfactory system rather than by the VNO. For example, the sow response to boar androstenone, alluded to above, was still present after the VNO was blocked with surgical cement (Dorries et al., 1997). Hence, the absence of a functional VNO in human adults does not preclude pheromone perceptive abilities. Human olfaction is as good as that of other mammals but it is consciously underdeveloped and overwhelmingly rated as the least important sense (Sela and Sobel, 2010) – people shamelessly gawp and overlisten but most balk at sniffing. Pheromone detection can improve with practice, as was shown for androstenone (Wysocki et al., 1989).
Pheromone solutions are often applied to the skin of study participants. Given that they affect physical parameters including the autonomic nervous system, Bensafi et al. (2004b) proposed that pheromones may also act through transdermal diffusion.
Pheromone study methodology and potential biases
The study subjects were usually volunteers recruited by posters etc., which means an overall younger and well-educated sample. In the majority of studies, care was taken to not disclose the true purpose of the study to the participants, who were told it was a study on “odors”. Yet, as some groups built up an extensive research expertise, one might doubt that all campus volunteers were oblivious to the nature of the study; inclusion restrictions (such as women not using contraceptive pills) and sexual preference disclosure might also have raised doubts that it was simply a study on “odors”. In the very best of studies, the study purpose was not revealed at all but the volunteers were “deceived “with some trick and the odor was subliminally introduced (reviewed in Havlicek and Roberts, 2009).
In the majority of studies, the pheromone and control solutions were applied to the skin between the upper lip and nose on a cotton swab, taped gauze or Q-tip, or they were sniffed from jars. Clearly, the concentration of the pheromone in the test solution was pharmacological in all studies, typically 0.25-6.25*10-3mol/l (compared to the physiological 0.44*10-6mol/l in apocrine sweat) for androstadienone. Given the high concentration and the odorous nature of 16-androstenes, a subset of the study participants could identify the musky smell of the pheromone solution, comparing it to “sweat”, “urine”, “a male roommate” or “clothes” (Jacob and McClintock, 2000; Lundström et al., 2003a). Therefore, in subsequent studies, researchers were at pains to mask the pheromone (and control) odor, for example by adding1% clove oil or 1% eugenol to the vehicle solution. However, adding masking odors to the solution may have affected the study results. In addition, masking odors do not eliminate bias: some participants thought the clove solutions smelled like “Christmas” – with possible positive or negative associations (Lundström et al., 2003a); and some still ascribed a particular smell to the masked pheromone solution (Saxton et al., 2008).
The pharmacological concentration of the pheromone is not in itself an issue – provided the experiments yield clear and consistent results with a convincing P level. Another crucial issue in research regarding social/psychosexual chemosignals is the setting of the experiment and the gender (and one might add the looks, voice and smell) of the study attendant who distributes the solutions and supervises the tests.
One study examined pheromones released by breastfeeding women using within-brassiere pads (likely containing spilled milk, apocrine sweat, baby saliva, etc.) and axillary pads obtained from donors, subsequently “equilibrated” by buffer solutions and cut into four. Recipient women needed to wipe one specimen of both type of pads under their noses, each day for 3 months (Spencer et al., 2004). Such experiments might be considered ethically borderline these days.
Modulator effects of pheromones in women
Several studies with slightly different methodology demonstrated that female participants showed increased positive-stimulated mood (affect) and reduced negative mood after exposure to either male axillary extracts (Preti et al., 2003) or purified androstadienone (but not androstenol or vehicle) (Jacob and McClintock, 2000; Jacob et al., 2002; Wyart et al., 2007). The androstadienone-induced mood uplift was dependent on its dose (Bensafi et al., 2004b) and on the pre-exposure mood engendered by a “sad” or “happy” video (Bensafi et al., 2004a), and was abolished when a pain stimulus was given (Villemure and Bushnell, 2007). A robust reduction in feelings of negative affect and negative character was also found when androstadienone was administered by a vapor pulse directly to the putative VNO (Grosser et al., 2000).
In addition, female participants also felt more focused (physically more aroused) after exposure to androstadienone (Lundström et al., 2003a; Wyart et al., 2007).
The improvement in mood and focus, induced by androstadienone, is potentially important for women’s sexuality. Indeed, studies in a laboratory setting showed that adoption of a positive mood (ter Kuile et al., 2010) or a positive sexual schema (set of cognitive views) (Kuffel and Heiman, 2006) facilitated subsequent mental and genital arousal, also in women with low baseline mood. In addition, cognitive distraction during sexual activity is an established predictor of low sexual satisfaction (Dove and Wiederman, 2000). Perhaps through these psychomechanistic pathways, androstadienone (as well as estratetraenol) were found to enhance sexual arousal induced by an erotic video (Bensafi et al., 2004a). In addition, the application of breast and axillary pads from breastfeeding women over 3 months robustly increased sexual desire and sexual fantasies in recipient women (Spencer et al., 2004); although it remains unclear whether putative pheromones from sweat or nursing secretions were responsible for this observation.
Androstadienone also affected biological parameters compatible with increased physical arousal. Activation of the autonomic nervous system produced sympathetic effects in women, cooling their hand palms and increasing their digit skin conductance (Jacob et al., 2001a; Bensafi et al., 2003), particularly at high concentrations (Bensafi et al., 2004b). Moreover, salivary cortisol levels were higher after androstadienone exposure compared with vehicle (Wyart et al., 2007) - perhaps reflecting a weak “cortisol awakening response”, well characterized following awakening in the morning (Elder et al., 2013).
The brain areas activated by androstadienone are still unclear, with inconsistent results across studies (Jacob et al., 2001b; Gulyás et al., 2004; Berglund et al., 2006). Preliminary data suggest that the brain processing of androstadienone and estratetraenol is different in heterosexual and lesbian women (Berglund et al., 2006).
Importantly, the positive effects of androstadienone on mood and skin conductance (Jacob et al., 2001a) as well as attention (Lundström and Olsson, 2005) depended on the study attendant’s gender, being present only with a male attendant. One might argue that, in Freudian terms, the modulator effect of the pheromone was “transferred” onto the study attendant. In contemporary language, the pheromone was interpreted within a given ecological context. Nonetheless, androstadienone also enhanced attention to emotional information in a more recent study which was performed largely without an attendant (Hummer and McClintock, 2009).
In conclusion, the upper-lip application of androstadienone (and to a lesser extent estratetraenol) was associated with a positive mood and greater focus – in particular to capture emotional information – in women. However, these effects were dependent on the socioexperimental context.
Primer effects on neuro-endocrine systems in women
Preliminary studies have suggested some degree of menstrual synchrony after pheromone exposure. For example, in a double-blind experiment, recipient women who daily applied onto their upper lip a donor axillary pad extract, reduced their cycle-day difference with donor women from a mean of 8.3 days to a mean of 3.9 days over 3 cycles. In contrast, recipients who applied a blank solution had on average 6.2 cycle-days difference with these donor women at the start and 7.6 days after 3 cycles (Preti et al., 1986). The effect may be mediated by alterations in hypothalamic-pituitary regulation, since exposure to male axillary extracts shortened the LH-pulse interval by (a modest) 20% (Preti et al., 2003).
Yet Strassmann (1999) has criticized some studies that purportedly show a role for pheromones in menstrual synchrony (Stern and McClintock, 1998). She notes that no anthropological evidence for synchrony could be obtained in African communities living close to one another. Evolutionary speaking, amenorrhea was (is) the “gold standard” with most women in their reproductive years either pregnant or lactating in times of plenty, and in (hypogonadal) amenorrhea to save micronutrients in times of famine or war. Menstrual synchrony is no priority when menses are few (Kung women in the Kalahari only had a mean of 48 menstrual periods during their lifetime). In addition, a significant fraction of menses were (are) the result of early pregnancy failures; and age >35 years adds intra- and inter-individual variation in cycle length owing to (non-synchronizable) ovarian follicle depletion.
In conclusion, any potential effects of pheromones on the menstrual cycle need further validation. At the present time, no firm conclusions can be drawn.
Signaler effects of pheromones
Axillary sweat is believed to provide a “chemical signature”. Adult humans can identify their own worn T-shirts out of 100 identical shirts worn by others (Lord and Kaszprak, 1989). Family members can also identify T-shirts belonging to their partners, children and siblings (Porter and Moore, 1981; Wysocki and Preti, 2004). Even breastfed infants can identify the axillary pad belonging to their mother – assessed by the duration of orientation of their heads toward the pad (Cernoch and Porter, 1985).
Such signaler effects may be crucial for partner selection. In several species, mating is “disassortative” regarding the major histocompatibility complex (MHC) genotype, also known as the human leukocyte antigen (HLA A, B, C, DR, DQ) system in humans: i.e., individuals prefer a mate with a dissimilar MHC-genotype, probably in an attempt to protect their offspring from homozygosity-induced illness. MHC-heterozygosity is more frequent than expected by chance. For dissimilar mate selection, the senses are critical: the visual sense for face and body scanning and attributing “attractiveness” to them, the auditory sense for vocal characteristics, and the olfactory (and VNO in non-humans) sense for body odors. Although the research is still ongoing, there is no evidence to date that facial preference is MHC-disassortative, in fact one study showed a preference for HLA-similar faces (reviewed in Havlicek and Roberts, 2009). On the other hand, most although not all studies confirmed that body odor preference is indeed HLA-disassortative (Havlicek and Roberts, 2009).
If body odor scanning is important for mate selection, do the 16-androstenes – the most important pheromones identified at this time – play a mediating role? Saxton et al. (2008) used a real-world approach to examine the effect of androstadienone: speed-dating events. They set up 3 different experiments with 12-25 women and 19-25 men, and 3-minute interactions between them; women scored the attractiveness of the potential date while being exposed to a cotton wool pad under their noses containing either androstadienone in 1% clove oil, 1% clove oil, or water. In two of the 3 experiments, the attractiveness ratings were better with androstedienone than water, and in 1 experiment better with androstedienone than clove oil only. In another study, there was a correlation between women’s preference for masculine faces for a long-term relationship and their pleasantness rating of androstadienone (but not androstenone) (Cornwell et al., 2004).
In conclusion, preliminary studies suggest that pheromones play a role in disassortative mate selection through the attribution of attractiveness.
General conclusions
Pheromones continue to be an exciting area of research. Currently, the 16-androstenes, and in particular androstadienone is fairly well characterized as a pheromone for women. They are present in male axillary sweat and possibly in other bodily secretions. Androstadienone improves mood and focus and modulates biological endpoints as well. A positive mood and heightened focus are important for women’s sexual response and sexual satisfaction. However, further study is required to validate these conclusions before any commercial claim of the “arousing” activity of androstadienone can be endorsed scientifically. Preliminary evidence suggests that androstadienone also plays a role in mate selection, perhaps through the attribution of attractiveness to facial and bodily features of potential mates.
As the pheromone field matures, so will study methodology. The ideal study does not disclose the purpose of the study at all and does not involve skin application of compounds with masking odors, but introduces the experimental pheromone subliminally and with no additive to the participants (in the room air) while strongly controlling the psychosocial setting of the experiment.
References
- Bancroft J, Graham CA, Janssen E, et al. The dual control model: current status and future directions. J Sex Res. 2009;46:121–142. doi: 10.1080/00224490902747222. [DOI] [PubMed] [Google Scholar]
- Basson R. Human sex-response cycles. J Sex Marit Ther. 2001;27:33–43. doi: 10.1080/00926230152035831. [DOI] [PubMed] [Google Scholar]
- Bensafi M, Brown WM, Tsutsui T, et al. Sex-steroid compounds induce sex-specific effects on autonomic nervous system function in humans. Behav Neurosc. 2003;117:1125–1134. doi: 10.1037/0735-7044.117.6.1125. [DOI] [PubMed] [Google Scholar]
- Bensafi M, Brown WM, Khan R, et al. Sniffing human sex-steroid derived compounds modulates mood, memory and autonomic nervous system function in specific behavioral contexts. Behav Brain Res. 2004;152:11–22. doi: 10.1016/j.bbr.2003.09.009. [DOI] [PubMed] [Google Scholar]
- Bensafi M, Tsutsui T, Khan R, et al. Sniffing a human sex-steroid derived compound affects mood and autonomic arousal in a dose-dependent manner. Psychoneuroendocrinology. 2004;29:1290–1299. doi: 10.1016/j.psyneuen.2004.03.007. [DOI] [PubMed] [Google Scholar]
- Berglund H, Lindström P, Savic I. Brain response to putative pheromones in lesbian women. Proc Natl Acad Sci USA. 2006;21:8269–8274. doi: 10.1073/pnas.0600331103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cernoch JM, Porter RH. Recognition of maternal axillary odors by infants. Child Developm. 1985;56:1593–1598. [PubMed] [Google Scholar]
- Cornwell RE, Boothroyd L, Burt DM, et al. Concordant preferences for opposite-sex signals? Human pheromones and facial characteristics. Proc Biol Sci. 2004;271:635–640. doi: 10.1098/rspb.2003.2649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dorries KM, Regan E, Halpern BP. Sensitivity and behavioral responses to the pheromone androstenone are not mediated by the vomeronasal organ in domestic pigs. Brain Behav Evol. 1997;49:53–62. doi: 10.1159/000112981. [DOI] [PubMed] [Google Scholar]
- Dove NL, Wiederman MW. Cognitive distraction and women’s sexual functioning. J Sex Marit Ther. 2000;26:67–78. doi: 10.1080/009262300278650. [DOI] [PubMed] [Google Scholar]
- Elder GJ, Wetherell MA, Barclay NL, et al. The cortisol awakening response – applications and implications for sleep medicine. Sleep Med Rev. 2013;(in press) doi: 10.1016/j.smrv.2013.05.001. [DOI] [PubMed] [Google Scholar]
- Feldman R, Gordon I, Sharon O. Maternal and paternal plasma, salivary, and urinary oxytocin and parent-infant synchrony: considering stress and affiliation components of human bonding. Dev Sci. 2011;14:752–761. doi: 10.1111/j.1467-7687.2010.01021.x. [DOI] [PubMed] [Google Scholar]
- Grosser BI, Bloch L, White C, et al. Behavioral and electrophysiological effects of androstadienone, a human pheromone. Psychoneuroendocrinology. 2000;25:289–299. doi: 10.1016/s0306-4530(99)00056-6. [DOI] [PubMed] [Google Scholar]
- Gulyás B, Kéri S, O'Sullivan BT, et al. The putative pheromone androstadienone activates cortical fields in the human brain related to social cognition. Neurochem Int. 2004;44:595–600. doi: 10.1016/j.neuint.2003.10.003. [DOI] [PubMed] [Google Scholar]
- Havlicek J, Roberts SC. MHC-correlated mate choice in humans: a review. Psychoneuroendocrinology. 2009;34:497–512. doi: 10.1016/j.psyneuen.2008.10.007. [DOI] [PubMed] [Google Scholar]
- Hays WST. Human pheromones: have they been demonstrated? Behav Ecol Sociobiol. 2003;54:89–97. [Google Scholar]
- Hummer TA, McClintock MK. Putative human pheromone androstadienone attunes the mind specifically to emotional information. Horm Behav. 2009;55:548–559. doi: 10.1016/j.yhbeh.2009.01.002. [DOI] [PubMed] [Google Scholar]
- Jacob S, McClintock MK. Psychological state and mood effects of steroidal chemosignals in women and men. Horm Behav. . 2000;37:57–78. doi: 10.1006/hbeh.1999.1559. [DOI] [PubMed] [Google Scholar]
- Jacob S, Hayreh DJS, McClintock MK. Context-dependent effects of steroid chemosignals on human physiology and mood. Physiol Behav. 2001;74:15–27. doi: 10.1016/s0031-9384(01)00537-6. [DOI] [PubMed] [Google Scholar]
- Jacob S, Kinnunen LH, Metz J, et al. Sustained human chemosignal unconsciously alters brain function. Neuro Report. 2001;12:2391–2394. doi: 10.1097/00001756-200108080-00021. [DOI] [PubMed] [Google Scholar]
- Jacob S, Garcia S, Hayreh D, et al. Psychological effects of musky compounds: comparison of androstadienone with androstenol and muscone. Horm Behav. 2002;42:274–283. doi: 10.1006/hbeh.2002.1826. [DOI] [PubMed] [Google Scholar]
- Kuffel SW, Heiman JR. Effects of depressive symptoms and experimentally adopted schemas on sexual arousal and affect in sexually healthy women. Arch Sex Behav. 2006;35:163–177. doi: 10.1007/s10508-005-9015-1. [DOI] [PubMed] [Google Scholar]
- Lord T, Kasprzak M. Identification of self through olfaction. Percept Mot Skills. 1989;69:267–277. doi: 10.2466/pms.1989.69.1.219. [DOI] [PubMed] [Google Scholar]
- Lundström JN, Gonçalves M, Esteves F, et al. Psychological effects of subthreshold exposure to the putative human pheromone 4,16-androstadien-3-one. Horm Behav. 2003;44:395–401. doi: 10.1016/j.yhbeh.2003.06.004. [DOI] [PubMed] [Google Scholar]
- Lundström JN, Hummel T, Olsson MJ. Individual differences in sensitivity to the odor of 4,16-androstadien-3-one. Chem Senses. 2003;28:643–650. doi: 10.1093/chemse/bjg057. [DOI] [PubMed] [Google Scholar]
- Lundström JN, Olsson MJ. Subthreshold amounts of social odorant affect mood, but not behavior, in heterosexual women when tested by a male, but not a female, experimenter. Biol Psychol. 2005;70:197–204. doi: 10.1016/j.biopsycho.2005.01.008. [DOI] [PubMed] [Google Scholar]
- Monti-Bloch L, White C, Berliner DL. The human vomeronasal system: a review. Ann NY Acad Sci. 1998;855:373–389. doi: 10.1111/j.1749-6632.1998.tb10595.x. [DOI] [PubMed] [Google Scholar]
- Porter RH, Moore JD. Human kin recognition by olfactory cues. Physiol Behav. 1981;27:493–495. doi: 10.1016/0031-9384(81)90337-1. [DOI] [PubMed] [Google Scholar]
- Preti G, Cutler WB, Garcia CR, et al. Human axillary secretions influence women’s menstrual cycles: the role of donor extract of females. Horm Behav. 1986;20:474–482. doi: 10.1016/0018-506x(86)90009-7. [DOI] [PubMed] [Google Scholar]
- Preti G, Wysocki CJ, Barnhart KT, et al. Male axillary extracts contain pheromones that affect pulsatile secretion of luteinizing hormone and mood in women recipients. Biol Reprod. 2003;68:2107–2113. doi: 10.1095/biolreprod.102.008268. [DOI] [PubMed] [Google Scholar]
- Saxton TK, Lyndon A, Little AC, et al. Evidence that androstadienone, a putative human chemosignal, modulates women’s attributions of men’s attractiveness. Horm Behav. 2008;54:597–601. doi: 10.1016/j.yhbeh.2008.06.001. [DOI] [PubMed] [Google Scholar]
- Sela L, Sobel N. Human olfaction: a constant state of change-blindness. Exp Brain Res. 2010;205:13–29. doi: 10.1007/s00221-010-2348-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spencer NA, McClintock MK, Sellergren SA, et al. Social chemosignals from breastfeeding women increase sexual motivation. Horm Behav. 2004;46:362–370. doi: 10.1016/j.yhbeh.2004.06.002. [DOI] [PubMed] [Google Scholar]
- Stern K, McClintock MK. Regulation of ovulation by human pheromones. Nature. 1998;392:177–179. doi: 10.1038/32408. [DOI] [PubMed] [Google Scholar]
- Strassmann BI. Menstrual synchrony pheromones: cause for doubt. Hum Reprod. 1999;14:579–580. doi: 10.1093/humrep/14.3.579. [DOI] [PubMed] [Google Scholar]
- ter Kuile MM, Both S, van Uden J. The effects of experimentally-induced sad and happy mood on sexual arousal in sexually healthy women. J Sex Med. 2010;7:1177–1184. doi: 10.1111/j.1743-6109.2009.01632.x. [DOI] [PubMed] [Google Scholar]
- Thysen B, Elliott WH, Katzman PA. Identification ofestra-1,3,5(10),16-tetraen-3-ol (estratetraenol) from the urine of pregnant women. Steroids. 1968;11:73–87. doi: 10.1016/s0039-128x(68)80052-2. [DOI] [PubMed] [Google Scholar]
- Villemure C, Bushnell MC. The effects of the steroid androstadienone and pleasant odorants on the mood and pain perception of men and women. Eur J Pain. 2007;11:181–191. doi: 10.1016/j.ejpain.2006.02.005. [DOI] [PubMed] [Google Scholar]
- Wyart C, Webster WW, Chen JH, et al. Smelling a single component of male sweat alters levels of cortisol in women. J Neurosci. 2007;27:1261–1265. doi: 10.1523/JNEUROSCI.4430-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wysocki CJ, Dorries KM, Beauchamp GK. Ability to perceive androstenone can be acquired by ostensibly anosmic people. Proc Natl Acad Sci USA. 1989;86:7976–7978. doi: 10.1073/pnas.86.20.7976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wysocki CJ, Preti G. Facts, fallacies, fears, and frustrations with human pheromones. Anat Record Part A. 2004;281A:1201–1211. doi: 10.1002/ar.a.20125. [DOI] [PubMed] [Google Scholar]