

Abstract
Human Leukocyte Antigen-G (HLA-G) contributes to cancer cell immune escape from host antitumor responses. The clinical relevance of HLA-G in several malignancies has been reported. However, the role of HLA-G expression and functions in Acute Myeloid Leukemia (AML) is still controversial. Our group identified a subset of tolerogenic dendritic cells, DC-10 that express HLA-G and secrete IL-10. DC-10 are present in the peripheral blood and are essential in promoting and maintaining tolerance via the induction of adaptive T regulatory (Treg) cells. We investigated HLA-G expression on blasts and the presence of HLA-G-expressing DC-10 and CD4+ T cells in the peripheral blood of AML patients at diagnosis. Moreover, we explored the possible influence of the 3′ untranslated region (3′UTR) of HLA-G, which has been associated with HLA-G expression, on AML susceptibility. Results showed that HLA-G-expressing DC-10 and CD4+ T cells are highly represented in AML patients with HLA-G positive blasts. None of the HLA-G variation sites evaluated was associated with AML susceptibility. This is the first report describing HLA-G-expressing DC-10 and CD4+ T cells in AML patients, suggesting that they may represent a strategy by which leukemic cells escape the host's immune system. Further studies on larger populations are required to verify our findings.
1. Introduction
Human Leukocyte Antigen (HLA)-G is a nonclassical HLA class I molecule, originally described essential for promoting fetus-maternal tolerance [1, 2]. It is now clear that HLA-G is involved in promoting beneficial tolerance in several settings, such as autoimmunity and organ transplantation, and in contributing to detrimental tolerance in viral infections and cancer [3]. HLA-G is expressed in seven different isoforms, four of which are membrane-bound (HLA-G1, G2, G3, and G4) and three are soluble (HLA-G5, G6, and G7) [4, 5]. Among these isoforms, the best characterized are HLA-G1, the most stable membrane-bound isoform, soluble HLA-G5, and shed HLA-G1. HLA-G modulates immune responses through several nonexclusive mechanisms: it inhibits cytolytic activities of NK and CD8+ cytotoxic T cells [6] and proliferation of T cells [7, 8], and it modulates antigen presenting cell (APC) differentiation and function [9]. APCs overexpressing HLA-G are poor stimulators and are able to induce anergic/suppressor CD4+ T cells [9–12]. Our group described a subset of tolerogenic IL-10-producing DC (DC-10) that is present in the peripheral blood [13, 14]. DC-10 are characterized by the expression of membrane-bound HLA-G and by their ability to induce adaptive IL-10-producing T regulatory (Treg) cells [13, 14]. We demonstrated that DC-10 accumulate in human decidua during pregnancy where they contribute in generating a tolerogenic microenvironment limiting immune responses in vivo [15].
Studies on solid tumors revealed that HLA-G could be potentially expressed by all tumors, either as membrane-bound or as soluble isoform [16, 17]. In these contexts, HLA-G acts as a negative regulator of the immune response through different mechanisms, including inhibition of angiogenesis, prevention of antigen recognition and T cell migration, and suppression of T and NK cytotoxicity [16, 18, 19]. HLA-G expression by leukemic cells is still controversial. Analysis performed on blasts from patients with different leukemia, including Acute Myeloid Leukemia (AML), Acute Lymphoid Leukemia (ALL), and Chronic Myeloid Leukemia (CML), demonstrated that neither mRNA for any HLA-G isoforms nor HLA-G antigen was detected [20, 21]. However, more recently it has been shown that the expression of HLA-G by circulating blasts from AML, CML, but not B-ALL, and by B-CLL cells was strongly associated with an unfavorable outcome of the disease [22–24]. In addition, a correlation between soluble HLA-G plasma levels and AML, ALL, and B-CLL was proposed [25, 26].
Despite low degree of genetic variability in the coding region of HLA-G, several polymorphisms are present in the noncoding region of the gene both at 5′ upstream regulatory region (URR) and 3′ untranslated region (UTR), which may influence the HLA-G expression [27]. The most studied polymorphism at the 3′ UTR is the presence (Ins) or absence (Del) of a fragment of 14 base pairs (14 bp Ins/Del) that has been associated with HLA-G mRNA stability [28–30]. In addition, the +3142 C/G single nucleotide polymorphism (SNP) controls the magnitude of mRNA production, since the presence of the G may increase the affinity of this region for miR-148a, miR-148b, and miR-152 [31, 32]. The +3187 A/G SNP has been reported to affect mRNA stability due to its proximity to an AU-rich motif, which mediates the HLA-G mRNA degradation [33]. Beside these polymorphic sites, other less studied SNPs of the 3′UTR are located at positions +3001 T/C, +3003 T/C, +3010 C/G, +3027 C/A, +3035 C/T, and +3196 C/G [34, 35].
The 14 bp Ins/Del has been associated with tolerance in different clinical conditions including autoimmunity [36–38], pathological pregnancy [39–42], recurrent spontaneous abortions [39, 40, 42, 43], and preeclampsia [29, 41, 43–45], although the results on the two latter conditions are contradictory. The presence of the 14 bp Del has been found to be predictive of the incidence of graft versus host disease after unrelated [46] and HLA-identical sibling [47] hematopoietic stem cell transplantation (HSCT) for beta-Thalassemia, suggesting a role for this polymorphism in the establishment of immunological tolerance also in the context of HSCT. The association of HLA-G polymorphisms with malignancies has been studied in a wide range of solid tumors, including breast and cervical cancers [48–52], but, thus far, it has not been evaluated in leukemia.
We investigated the expression of HLA-G on leukemic blasts and tolerogenic immune cells, DC-10 and CD4+ T cells, in the peripheral blood of AML patients at diagnosis. We also determined whether polymorphisms at 3′ UTR of HLA-G locus correlate with AML susceptibility.
2. Materials and Methods
2.1. Patients
All protocols were approved by the institutional review board and samples were collected under written informed consent according to the Declaration of Helsinki. 22 patients affected by AML were included in this retrospective study and analyzed for biological and clinical characteristics. AML diagnosis was based on standard cytological criteria according to the French-American-British (FAB) classification. Patients' diagnosis was subclassified by morphological and immune phenotyping. None of the patients received medical interventions before the study. Patients' characteristics are listed in Tables 1 and 2.
Table 1.
Clinical patients' characteristics.
| Variable | All patients | Blasts in PB Mean % (range) | HLA-G+ blasts n (mean %) | HLA-G− blasts n (mean %) | P value$ |
|---|---|---|---|---|---|
| Number of patients (%) | 22 | 65.6 (13.3–96.4) | 15 (68.2) | 7 (31.8) | |
| Age at diagnosis (year) | 59 (22–83) | ||||
| Male (%) | 8 (36) | 73.9 (13.3–94.5) | 8 (100) | 0 (0) | 0.022 |
| Female (%) | 14 (64) | 60.8 (23.9–96.4) | 7 (50) | 7 (50) | |
| AML | 3 | 58.1 (42.2–85.2) | 2 (66.7) | 1 (33.3) | |
| AML-M0 | 2 | 35.6 (25.4–45.7) | 1 (50) | 1 (50) | |
| AML-M1 | 1 | 68 | / | 1 (100) | |
| AML-M2 | 7 | 62.4 (23.9–94.5) | 5 (71.5) | 2 (28.5) | |
| AML-M3 | 1 | 94.4 | 1 (100) | / | |
| AML-M4 | 6 | 81.5 (40–95.3) | 4 (66.6) | 2 (33.4) | |
| AML-M5a | 1 | 96.4 | 1 (100) | / | |
| AML-M6 | 1 | 13.3 | 1 (100) | / |
$Comparison between HLA-G+ and HLA-G− blasts using Fisher's exact test.
Table 2.
HLA-G expression and cytogenetic karyotype.
| Patient number | FAB classification | Sex | Age | HLA-G expression | Karyotype |
|---|---|---|---|---|---|
| 1 | AML-M2 | F | 83 | Positive | 46XX +8[4];[16] |
| 3 | sAML | F | 69 | Negative | 46XX |
| 5 | AML-M4 | F | 54 | Positive | 46XX |
| 9 | AML-M4 | F | 44 | Negative | 46XX |
| 10 | AML-M2 | M | 60 | Positive | 46XY t(8; 21)(q22;q22), del(9)(q?22) |
| 11 | AML-M4 | M | 70 | Positive | 46XY[19]; +21[2]; iso p21[1] |
| 18 | AML-M2 | F | 68 | Negative | 46XX iso 8p |
| 19 | AML-M4 | F | 76 | Positive | 46XX |
| 20 | AML-M0 | F | 83 | Positive | 46XX −7/t(1;7;4;12)[12] |
| 21 | AML-M2 | F | 66 | Positive | 46XX +8[4];[16] |
| 23 | AML-M2 | M | 74 | Positive | 46XY |
| 34 | AML-M0 | F | 72 | Negative | 46XX |
| 36 | AML-M6 | M | 53 | Positive | 46XY complex(del5q,-10 14; 19, 7p-)[20] |
| 37 | AML-M2 | F | 59 | Negative | 46XX |
| 38 | AML | M | 59 | Positive | 46XY |
| 39 | AML-M1 | F | 47 | Negative | 46XX |
| 41 | AML-M4 | F | 41 | Negative | 46XY |
| 44 | AML | M | 36 | Positive | 46XY |
| 45 | AML-M4 | F | 44 | Positive | 46XX |
| 49 | AML-M2 | M | 77 | Positive | 46XY |
| 54 | AML-M3 | M | 22 | Positive | 46XY[6], t(15;17)[5] |
| 55 | AML-M5a | F | 58 | Positive | 46XX |
2.2. Cells Isolation and Serum Collection
Peripheral blood mononuclear cells (PBMCs) from AML patients were isolated by Ficoll density gradient centrifugation and cryopreserved in gas phase of liquid nitrogen to the time of analysis. Serum was obtained from the blood samples of AML patients by centrifugation and cryopreserved in gas phase of liquid nitrogen for ELISA test.
2.3. Cytogenetic Analysis
Cytological analysis was performed using standard G-band karyotyping technique. Results were described according to the International System for Human Cytogenetic Nomenclature [53].
2.4. Flow Cytometry Analysis
Frozen PBMCs were thawed in X-VIVO 15 medium (Lonza, Italy), supplemented with 5% pooled AB human serum (Lonza, Italy) and 100 U/mL penicillin/streptomycin (Lonza, Italy), and washed twice in Phosphate Buffered Saline (PBS) (Sigma, CA, USA) with 2% Fetal Bovine Serum (FBS) (Lonza, Italy). PBMCs were initially incubated for 10 min at room temperature with FcR blocking reagent (Miltenyi Biotech, Germany) and stained for additional 20 min at room temperature in the dark with monoclonal antibodies (mAbs) specific for the following human antigens: CD45 (BioLegend, USA), CD16 (BD Pharmigen, CA, USA), CD4 (Becton Dickinson, CA, USA), CD14 (Becton Dickinson, CA, USA), and HLA-G (MEM-G9, Exbio, Czech Republic). Cells were identified using a multiparametric approach based on the combination of mAbs. Samples were acquired using a FACS Canto II flow cytometer (Becton Dickinson, CA, USA), and data were analyzed with FCS express (De Novo Software, CA, USA). Quadrant markers were set accordingly to unstained controls. Leukemic blasts were identified as CD45dim according to Lacombe et al. [54].
2.5. Detection of Soluble HLA-Gs
Levels of shed HLA-G1 and soluble HLA-G5 were determined by enzyme-linked immunosorbent assay (ELISA), as previously described [55, 56]. To detect sHLA-G (shed HLA-G1 and HLA-G5) plates (Nunc-Immuno Plate PolySorp, Thermoscientific, Denmark) were coated with the mAb G233 (Exbio, Czech Republic), whereas to detect HLA-G5 plates were coated with the mAb 5A6G7 (Exbio, Czech Republic). sHLA-G or HLA-G5 was detected with biotinylated β 2-microglobulin or W6/32 mAbs (Exbio, Czech Republic), respectively. Supernatants from HLA-G transfected LCL721.221 [57] and HeLa HLA-G5 transfected cells (kindly provided by Dr. R. Rizzo, Università di Ferrara) purified by affinity chromatography by using the W6/32 mAb were used for the generation of standard calibration curves for sHLA-G and HLA-G5, respectively. The limit of sensitivity was 0.5 ng/mL.
2.6. Amplification and Sequencing of 3′UTR of the HLA-G Gene
Genomic DNA was extracted from PBMCs using a commercial kit (QIAamp, QIAGEN, Italy) according to the manufacturer's instructions. Briefly, 100 ng of genomic DNA was amplified in a 25 μL reaction containing 1X polymerase chain reaction (PCR) buffer (Roche, USA), 0.2 mM dNTP mix (Roche, USA), 1.5 mM MgCl2 (Roche, USA), 0.8 U Taq Polymerase (Roche, USA), and 1 μM of each primer (For.: 5′ TCACCCCTCACTGTGACTGA 3′; Rev.: 5′ TTCTCATGTCTTCCATTTATTTTGTC 3′). The initial denaturation step was carried out at 95°C for 3 min, followed by 30 cycles at 93°C for 60 s, 58°C for 60 s, and 72°C for 60 s and by a final extension step at 72°C for 10 min. The amplification product was evaluated using a 2.5% agarose gel, purified using a commercial kit (Wizard SV Gel and PCR Clean-Up System, Promega, WI, USA) according to the manufacturer's instructions, and subjected to direct sequencing on both strands. All polymorphic sites observed at the 3′UTR were individually annotated and named according to previous reports [35].
2.7. Statistical Analysis
All results are presented as mean values ± SD. Comparison of parameters between subgroups of patients was performed using the nonparametric Mann-Whitney U test for continuous variable and Fisher's exact test for categorical data. Differences were regarded as significant at P < 0.05. The results were analyzed using GraphPad Prism 3.0 (GraphPad Software, USA).
3. Results and Discussion
We investigated the expression of HLA-G on leukemic blasts from 22 patients affected by Acute Myeloid Leukemia (AML), referring to the San Raffaele Hematology and Bone Marrow Transplantation Program (Table 1). The cohort of patients analyzed was characteristic of AML, with a median age of 59 years, both male and female (36% and 64%, resp.), with AML-M2 and AML-M4 subtype predominance. The mean percentage of blasts in the peripheral blood of patients was 65.6 ± 27.97% (mean ± SD, n = 22; range 13.3–96.4%). Patients' PBMCs were analyzed by flow cytometry and the expression of HLA-G was determined on blasts identified as CD45dim cells with MEM-G/9 mAb, which detects the full-length HLA-G1 isoform. HLA-G expression on AML blasts less than 1% was considered negative. Results showed that HLA-G was expressed in 15 out of 22 (68.2%) AML patients, and percentage of HLA-G+ blasts varied from 1.5% to 59% (Figure 1). Although our study has been performed in a small cohort of Caucasian AML patients, it confirmed previous analyses in Chinese AML patients [58, 59], and indicate that membrane-bound HLA-G can be expressed on AML blasts. The analysis of the association between the expression of HLA-G on blasts and clinical parameters, including patient age, gender, subtype of AML, and percentage of blasts at diagnosis, revealed that HLA-G expression is independent of all the abovementioned variables except for gender. In our cohort of AML patients, all males showed HLA-G+ blasts (Table 1). Nevertheless, these results are partially in accordance with previous analyses demonstrating that the HLA-G expression on leukemic blasts is not associated with specific patients' characteristics [58].
Figure 1.
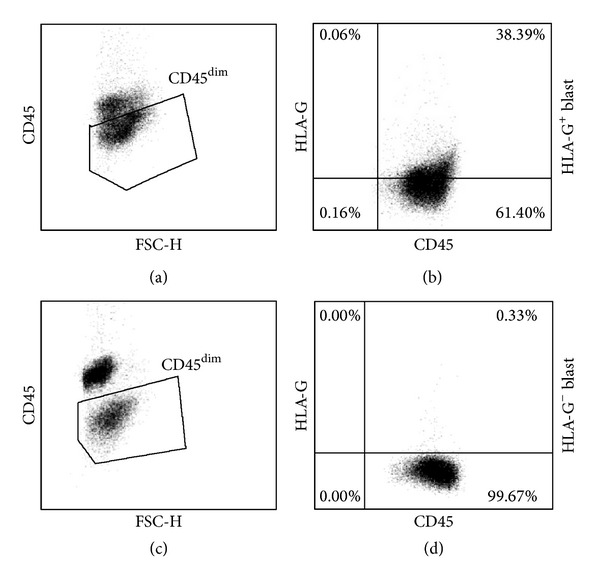
HLA-G expression on leukemic blasts. PBMCs of AML patients were analyzed by flow cytometry to determine the expression of HLA-G on blasts. An anti-human CD45 mAb was used to discriminate leukemic cells from normal mononuclear cells, being blasts CD45dim. Representative plots from a patient with HLA-G+ blast ((a) and (b)) or with HLA-G− blast ((c) and (d)) are shown.
We next investigated whether HLA-G expression on blasts can be associated with cytogenetic karyotype abnormalities. Results indicated that abnormalities were present in 7 out of 15 AML patients with HLA-G+ blasts and only in 1 patient with HLA-G− blasts (Table 2). These results are in contrast to previous data in which a marked difference in the frequency of cytogenetic abnormalities between HLA-G+ and HLA-G− AML blasts was observed [24]. It cannot be excluded that this discrepancy can be due to the different ethnic population analyzed. Nevertheless, it remains to define whether a correlation between cytogenetic abnormalities and HLA-G expression exists by enlarging the cohort of patients.
One of the strategies by which tumors escape the host's immune surveillance is the upregulation of HLA-G expression on both cancer and non-tumor cells, such as mononuclear cells [17]. Our group described DC-10, which constitutively expressed HLA-G and are involved in promoting tolerance [13–15]. We postulated that DC-10 might play a role in favoring tumor escape in AML patients. The population of cells containing DC-10 was identified by the concomitant expression of CD14 and CD16 on CD45bright cells. Results showed a significantly higher frequency of DC-10 in the peripheral blood of AML patients compared to that observed in healthy donors (1.5 ± 2.13%, n = 18, mean ± SD versus 0.19 ± 0.13%, n = 14, mean ± SD; P = 0.027; Figure 2(a)). The percentage of human DC-10 in AML patients with HLA-G+ and HLA-G− blasts was similar (1.4 ± 2.5, n = 12, mean ± SD versus 1.6 ± 1.26%, n = 6, mean ± SD, resp.; data not shown). Previous reports indicated that HLA-G itself promotes the expression of HLA-G on T and myeloid cells [10]; thus, we compared the frequency of HLA-G-expressing DC-10 in patients with HLA-G+ or HLA-G− blasts. Results showed a significantly higher frequency of HLA-G+ DC-10 in patients with HLA-G+ blasts as compared to those with HLA-G− blasts (41.3 ± 29.25%, mean ± SD, n = 12 versus 10.05 ± 12.08%, mean ± SD, n = 6; P = 0.013, Figures 2(b) and 2(c)). In parallel, the presence of naturally occurring CD4+ Treg cells constitutively expressing HLA-G (HLA-G+ CD4+ Treg cells), which have been identified in the peripheral blood of healthy donors and patients [15, 60, 61], was analyzed. The proportion of CD4+ T cells in the peripheral blood of leukemic patients and of healthy donors was similar (29.6 ± 18.59%, mean ± SD, n = 21 versus 31.43 ± 8.57%, mean ± SD, n = 16; Figure 3(a)). Interestingly, as for DC-10, the percentage of HLA-G+ CD4+ Treg cells was significantly higher in patients with HLA-G+ blasts than in those with HLA-G− blasts (4.2 ± 5.79%, mean ± SD, n = 14 versus 0.44 ± 1.01%, mean ± SD, n = 6; P = 0.0072 Figures 3(b) and 3(c)). This is the first report demonstrating the presence of HLA-G-expressing DC-10 and CD4+ T cells in the peripheral blood of leukemic patients. Our findings indicate that the frequency of regulatory cells, DC-10 and HLA-G+ CD4+ T cells, is increased in patients with HLA-G-expressing blasts, supporting the hypothesis that the expression of HLA-G on blasts may be a strategy by which leukemia promotes a tolerogenic microenvironment limiting anti-tumor responses. This mechanism of immune escape has been previously proposed for solid tumor where both infiltrating cells and tumor cells can express HLA-G [16]. It remains to be defined whether HLA-G-expressing tolerogenic cells are present in the bone marrow of AML patients where leukemic blasts reside and proliferate before emerging in the periphery. Moreover, correlation studies between HLA-G expression on blasts and the frequency of DC-10 and HLA-G+ CD4+ T cells will elucidate whether the microenvironment enriched in immunomodulatory factors allows the recruitment or the induction of tolerogenic cells inhibiting the antileukemic effects.
Figure 2.
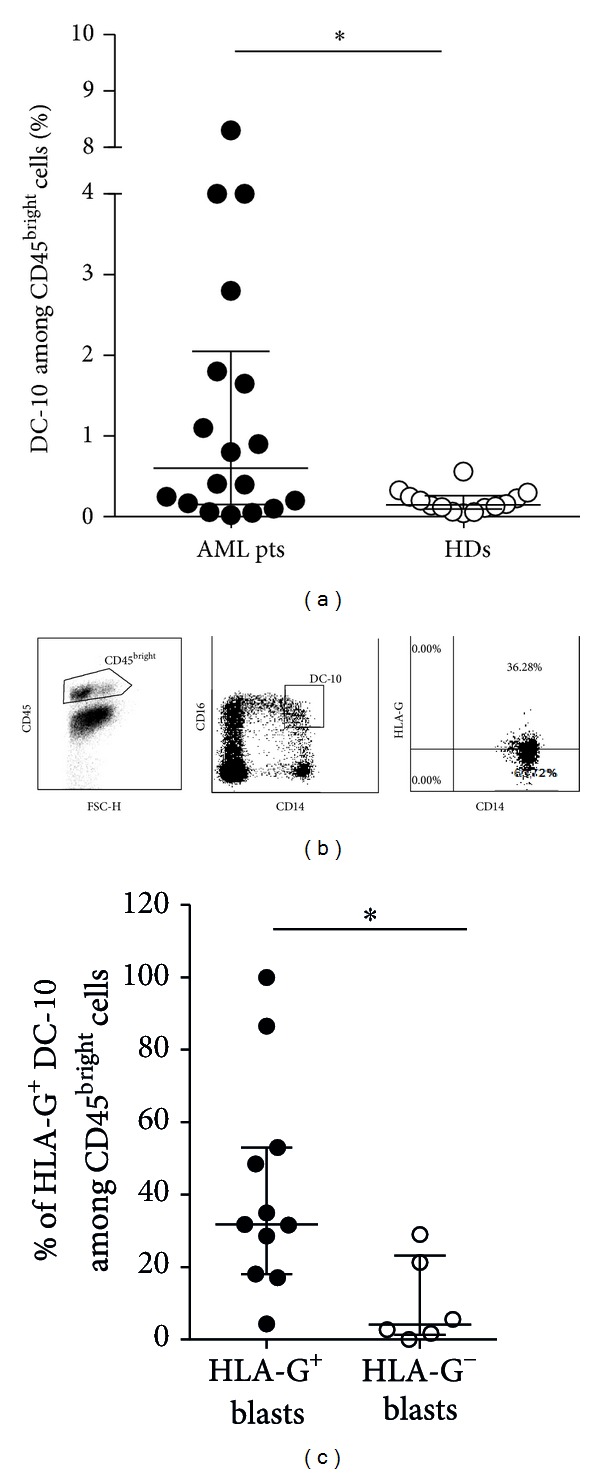
Flow cytometric analysis of cells containing DC-10 in the peripheral blood of AML patients. (a) Percentages of DC-10, identified among the CD45bright cells according to the co-expression of CD14 and CD16, in the peripheral blood of AML patients (AML pts) and healthy donors (HDs) are shown. Each dot represents single AML patient or HD. Lines indicate median and interquartile range of positive cells detected in all patients and donors analyzed. *P < 0.05. (b) Representative dot plots of HLA-G+ DC-10 are depicted. (c) Percentages of HLA-G+ DC-10 in patients with HLA-G+ or HLA-G− blasts are shown. Each dot represents single AML patient. Lines indicate the median and interquartile range of HLA-G+ DC-10. *P < 0.05.
Figure 3.
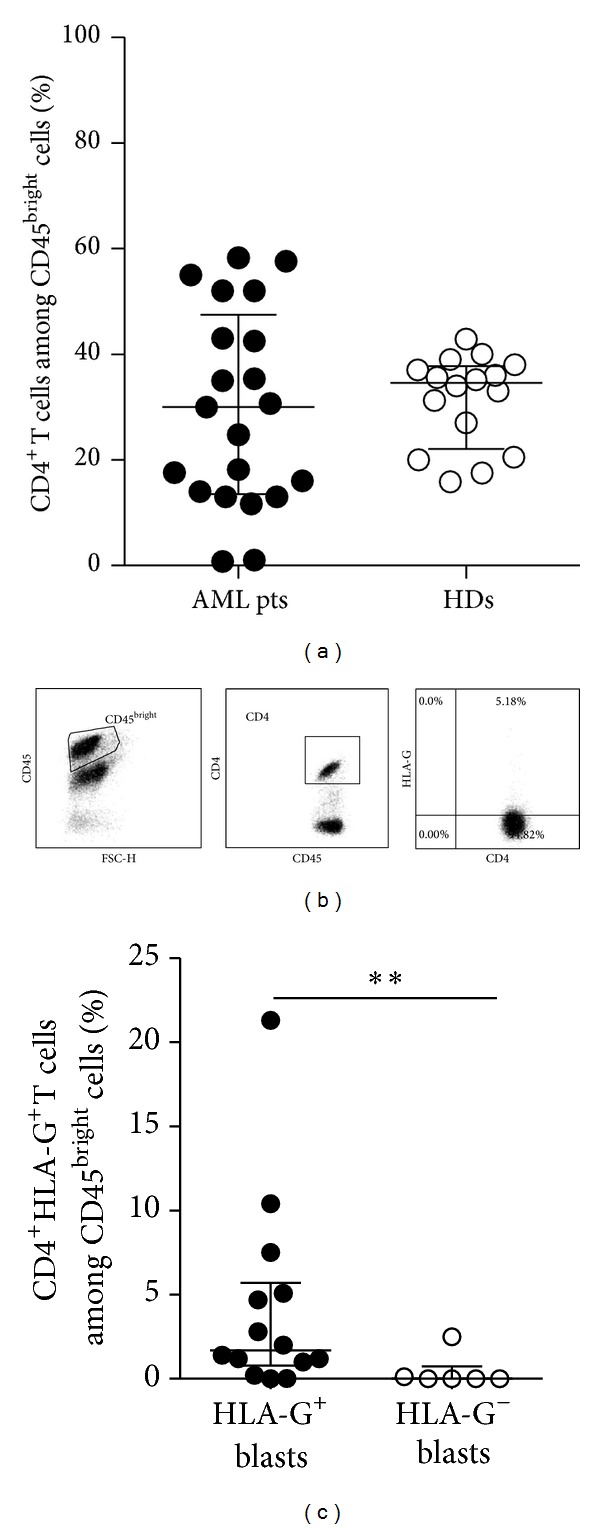
Flow cytometric analysis of CD4+ T cells in the peripheral blood of AML patients. (a) Percentages of CD4+ T cells among the CD45bright cells in the peripheral blood of AML patients (AML pts) and healthy donors (HDs) are shown. Each dot represents single AML patient or HD. Lines indicate the median and interquartile range of positive cells detected in all patients and donors analyzed. (b) Representative dot plots of HLA-G+ CD4+ T cells are depicted. (c) Percentages of HLA-G+ CD4+ T cells in patients with HLA-G+ or HLA-G− blasts are shown. Each dot represents single AML patient. Lines indicate the median and interquartile range of HLA-G+ CD4+ T cells. **P < 0.01.
Thus far, the presence of increased levels of soluble HLA-G has been associated with malignancies, including AML [25, 26]. We therefore sought to evaluate the amounts of soluble HLA-G in the serum of AML patients that were previously analyzed for HLA-G expression. sHLA-G (shed HLA-G1 and HLA-G5) was detected in 15 out of 18 patients with a mean value of positive samples of 1.78 ± 1.28 ng/mL (mean ± SD) and no differences were obtained between sera from patients with HLA-G+ and HLA-G− blasts (1.79 ± 1.29 ng/mL, n = 10 versus 1.75 ± 1.42 ng/mL, n = 5, mean ± SD, Figure 4). In line with previous findings demonstrating that sHLA-G plasma levels were significantly higher in AML-M4 and AML-M5 acute leukemia subtypes [25], we detected higher amounts of sHLA-G in sera of AML-M4 patients as compared to those of other AML subtypes (2.8 ± 1.28 ng/mL n = 5 versus 1.2 ± 0.93 ng/mL, n = 10 mean ± SD; data not shown). In parallel, we detected HLA-G5 in 10 out of 18 patients with a mean value of positive samples of 2.33 ± 1.84 ng/mL (mean ± SD), and similar to sHLA-G, no differences were found between sera from patients with HLA-G+ and HLA-G− blasts (2.33 ± 1.84 ng/mL, n = 7 versus 2.35 ± 2.5 ng/mL, n = 3; mean ± SD; Figure 4). Overall the levels of sHLA-G observed in our cohort of AML patients were lower compared to a previous work [25]. This discrepancy can be due to the fact that in our study we measured sHLA-G serum levels whereas Gros et al. [25] reported results from plasma samples. It has been indeed demonstrated that the levels of sHLA-G (shed HLA-G1 and HLA-G5) are significantly higher in plasma treated with EDTA as compared to those in plasma treated with heparin or in serum [62].
Figure 4.
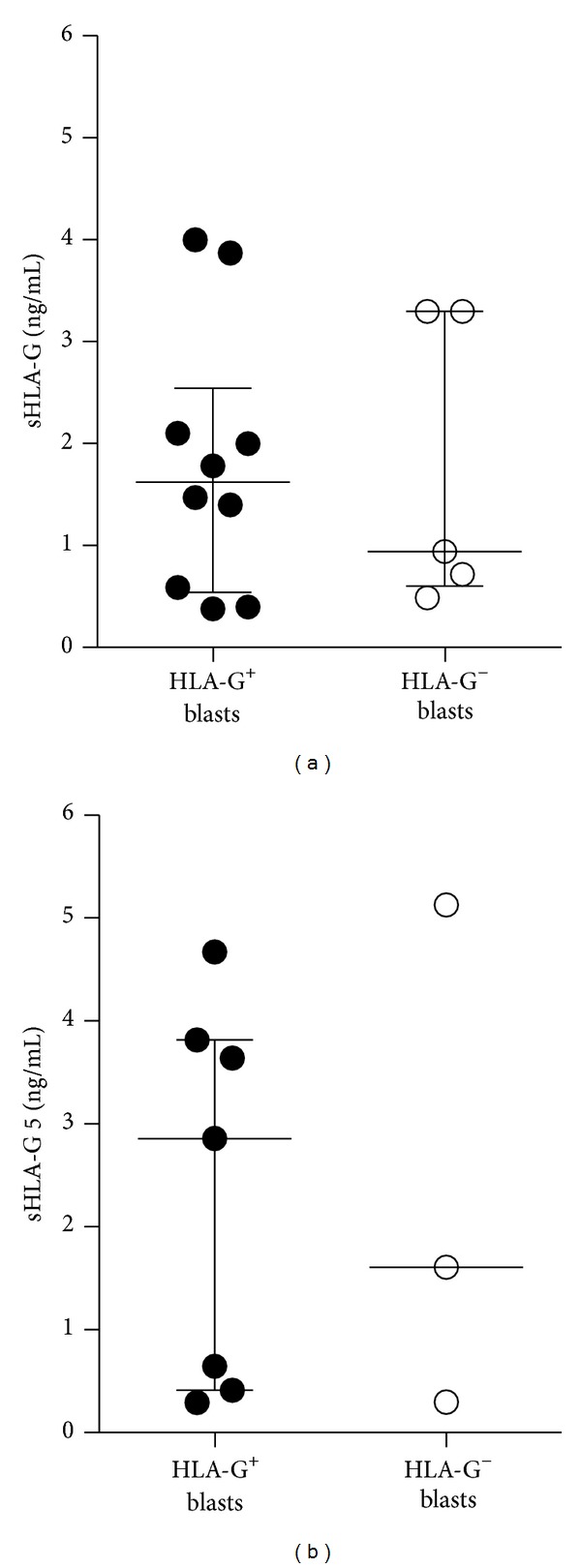
Soluble HLA-G levels in the sera of AML patients. The concentration of (a) sHLA-G (shed HLA-G1 and HLA-G5) and (b) HLA-G5 in the sera of AML patients was evaluated using specific sandwich ELISA. Each dot represents single AML patient. Lines indicate the median of serum levels (ng/mL) of positive samples in patients with HLA-G+ or HLA-G− blasts.
To define whether variations in the 3′UTR of HLA-G are associated with HLA-G expression, we analyzed 8 polymorphic sites at the HLA-G 3′UTR segment, including the 14 bp Ins/Del (rs1704), +3003 C/T (rs1707), +3010 C/G (rs1710), +3027 A/C (rs17179101), +3035 C/T (rs17179108), +3142 C/G (rs1063320), +3187 A/G (rs9380142), and +3196 C/G (rs1610696), previously described [34, 35]. The frequency of the 14 bp genotypes was similar in AML patients and healthy donors (Table 3). Since the 14 bp Ins is in strong linkage disequilibrium with the G in position +3142, we classified patients and controls according to the presence of the 14 bp Ins/Del and the +3142 C/G polymorphisms as InsG/InsG, DelC/DelC, DelC/InsG, and DelG/X. The relative frequencies of these genotypes in healthy donors were for InsG/InsG 23%, for DelC/DelC 22%, for DelC/InsG 32%, and for DelG/X 23% (Table 3). Interestingly, in AML patients we found a higher frequency of DelG/X genotype as compared to that observed in healthy donors (32% and 23%, resp.; Table 3). In line with these results, the frequency of UTR-3 haplotype (14 bp Del, +3003T, +3010C, +3027C, +3035C, +3142G, +3187A, and +3196C) was highly represented in AML patients than in healthy donors (16% and 9%, resp.; Table 3). Polymorphisms at the 3′UTR of HLA-G locus, and particularly the 14 bp Ins/Del, have been associated with different clinical conditions including autoimmunity and pathological pregnancy. So far, limited information has been published on the association of HLA-G polymorphisms in tumor cells with the levels of HLA-G expression and/or clinical outcome of patients [17]. Recently, studies in small cohort of patients investigated the association of 14 bp Ins/Del with the susceptibility to cervicovaginal and breast cancer, with controversial results [52, 63, 64]. Although in the present study the variation in the 3′UTR of HLA-G was evaluated in a limited number of AML patients, results showed no specific association, with the exception of UTR-3 haplotype. A more extensive study is warranted in a large cohort of patients in order to define whether specific UTRs of HLA-G might be associated with AML or can be used as genetic risk factor for the disease susceptibility.
Table 3.
Frequency of 3′UTR haplotypes and genotypes.
| 3′UTR polymorphic sites | AML patients (n = 19) | HDs (n = 141) |
|---|---|---|
| Ins/Ins | 0.26 | 0.22 |
| Del/Del | 0.37 | 0.30 |
| Ins/Del | 0.37 | 0.47 |
| InsG/InsG | 0.26 | 0.23 |
| DelC/DelC | 0.15 | 0.22 |
| InsG/DelC | 0.26 | 0.32 |
| DelG/X | 0.32 | 0.23 |
| UTR-1 | 0.21 | 0.32 |
| UTR-2 | 0.29 | 0.3 |
| UTR-3 | 0.16 | 0.09 |
| UTR-4 | 0.13 | 0.11 |
| UTR-5 | 0.03 | 0.05 |
| UTR-6 | 0.05 | 0.02 |
| UTR-7 | 0.08 | 0.08 |
| UTR-8 | 0.05 | 0.032 |
4. Conclusions
Results from this study further improve the knowledge on the role of HLA-G in promoting tolerance. Moreover, they opem new clinical perspectives: HLA-G expression can be used as prognostic tumor biomarker to monitor disease state or as therapeutic target for improving immune responses against leukemia. The expression of HLA-G on blasts and the analysis of DC-10 and HLA-G+ CD4+ Tregs can be used to evaluate the effectiveness of anti-tumor therapies. Moreover, the analysis of HLA-G polymorphisms will allow the identification of specific HLA-G genotypes that could be associated with AML susceptibility.
Acknowledgments
The authors thank Dr.ssa Roberta Rizzo for providing reagents for measuring soluble HLA-G, Dr. Giovanni Povoleri for technical help, and Dr.ssa Katharina Fleishhauer and Dr. Luca Vago for clinical patients' information. This work was supported by AIRC (Associazione Italiana per la Ricerca sul Cancro): Project AIRC-2010, IG 8978 to S.G. Dr. Giada Amodio conducted this study as partial fulfillment of her Ph.D. in Molecular Medicine, Program in Basic and Applied Immunology, San Raffaele University, Milan, Italy.
Conflict of Interests
The authors declare that there is no conflict of interests regarding the publication of this paper.
Authors' Contribution
Grazia Locafaro and Giada Amodio contributed equally to this work. Grazia Locafaro and Giada Amodio performed the experiments and the analysis of the data and contributed to the preparation of the paper; Daniela Tomasoni performed some of the experiments; Cristina Tresoldi and Fabio Ciceri provided the clinical samples; Silvia Gregori conceived the scientific idea, supervised the project, and wrote the paper.
References
- 1.Rouas-Freiss N, Gonçalves RM-B, Menier C, Dausset J, Carosella ED. Direct evidence to support the role of HLA-G in protecting the fetus from maternal uterine natural killer cytolysis. Proceedings of the National Academy of Sciences of the United States of America. 1997;94(21):11520–11525. doi: 10.1073/pnas.94.21.11520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Hunt JS, Petroff MG, McIntire RH, Ober C. HLA-G and immune tolerance in pregnancy. The FASEB Journal. 2005;19(7):681–693. doi: 10.1096/fj.04-2078rev. [DOI] [PubMed] [Google Scholar]
- 3.Gonzalez A, Rebmann V, LeMaoult J, Horn PA, Carosella ED, Alegre E. The immunosuppressive molecule HLA-G and its clinical implications. Critical Reviews in Clinical Laboratory Sciences. 2012;49(3):63–84. doi: 10.3109/10408363.2012.677947. [DOI] [PubMed] [Google Scholar]
- 4.Fujii T, Ishitani A, Geraghty DE. A soluble form of the HLA-G antigen is encoded by a messenger ribonucleic acid containing intron 4. Journal of Immunology. 1994;153(12):5516–5524. [PubMed] [Google Scholar]
- 5.Ishitani A, Geraghty DE. Alternative splicing of HLA-G transcripts yields proteins with primary structures resembling both class I and class II antigens. Proceedings of the National Academy of Sciences of the United States of America. 1992;89(9):3947–3951. doi: 10.1073/pnas.89.9.3947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Riteau B, Rouas-Freiss N, Menier C, Paul P, Dausset J, Carosella ED. HLA-G2, -G3, and -G4 isoforms expressed as nonmature cell surface glycoproteins inhibit NK and antigen-specific CTL cytolysis. Journal of Immunology. 2001;166(8):5018–5026. doi: 10.4049/jimmunol.166.8.5018. [DOI] [PubMed] [Google Scholar]
- 7.Lila N, Rouas-Freiss N, Dausset J, Carpentier A, Carosella ED. Soluble HLA-G protein secreted by allo-specific CD4+ T cells suppresses the allo-proliferative response: a CD4+ T cell regulatory mechanism. Proceedings of the National Academy of Sciences of the United States of America. 2001;98(21):12150–12155. doi: 10.1073/pnas.201407398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Le Rond S, Le Maoult J, Créput C, et al. Alloreactive CD4+ and CD8+ T cells express the immunotolerant HLA-G molecule in mixed lymphocyte reactions: in vivo implications in transplanted patients. European Journal of Immunology. 2004;34(3):649–660. doi: 10.1002/eji.200324266. [DOI] [PubMed] [Google Scholar]
- 9.Carosella ED, Gregori S, Rouas-Freiss N, Lemaoult J, Menier C, Favier B. The role of HLA-G in immunity and hematopoiesis. Cellular and Molecular Life Sciences. 2011;68(3):353–368. doi: 10.1007/s00018-010-0579-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.LeMaoult J, Krawice-Radanne I, Dausset J, Carosella ED. HLA-G1-expressing antigen-presenting cells induce immunosuppressive CD4+ T cells. Proceedings of the National Academy of Sciences of the United States of America. 2004;101(18):7064–7069. doi: 10.1073/pnas.0401922101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Naji A, Durrbach A, Carosella ED, Rouas-Freiss N. Soluble HLA-G and HLA-G1 expressing antigen-presenting cells inhibit T-cell alloproliferation through ILT-2/ILT-4/FasL-mediated pathways. Human Immunology. 2007;68(4):233–239. doi: 10.1016/j.humimm.2006.10.017. [DOI] [PubMed] [Google Scholar]
- 12.Gregori S. Dendritic cells in networks of immunological tolerance. Tissue Antigens. 2011;77(2):89–99. doi: 10.1111/j.1399-0039.2010.01615.x. [DOI] [PubMed] [Google Scholar]
- 13.Gregori S, Tomasoni D, Pacciani V, et al. Differentiation of type 1 T regulatory cells (Tr1) by tolerogenic DC-10 requires the IL-10-dependent ILT4/HLA-G pathway. Blood. 2010;116(6):935–944. doi: 10.1182/blood-2009-07-234872. [DOI] [PubMed] [Google Scholar]
- 14.Amodio G, Gregori S. Human tolerogenic DC-10: perspectives for clinical applications. Transplantation Research. 2012;1(1, article 14) doi: 10.1186/2047-1440-1-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Amodio G, Mugione A, Sanchez AM, et al. HLA-G expressing DC-10 and CD4(+) T cells accumulate in human decidua during pregnancy. Human Immunology. 2013;74(4):406–411. doi: 10.1016/j.humimm.2012.11.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Curigliano G, Criscitiello C, Gelao L, Goldhirsch A. Molecular pathways: human leukocyte antigen G, (HLA-G) Clinical Cancer Research. 2013;19(20):5564–5571. doi: 10.1158/1078-0432.CCR-12-3697. [DOI] [PubMed] [Google Scholar]
- 17.Amiot L, Ferrone S, Grosse-Wilde H, Seliger B. Biology of HLA-G in cancer: a candidate molecule for therapeutic intervention? Cellular and Molecular Life Sciences. 2011;68(3):417–431. doi: 10.1007/s00018-010-0583-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Rouas-Freiss N, Moreau P, Menier C, Carosella ED. HLA-G in cancer: a way to turn off the immune system. Seminars in Cancer Biology. 2003;13(5):325–336. doi: 10.1016/s1044-579x(03)00023-3. [DOI] [PubMed] [Google Scholar]
- 19.Rouas-Freiss N, Moreau P, Menier C, LeMaoult J, Carosella ED. Expression of tolerogenic HLA-G molecules in cancer prevents antitumor responses. Seminars in Cancer Biology. 2007;17(6):413–421. doi: 10.1016/j.semcancer.2007.07.003. [DOI] [PubMed] [Google Scholar]
- 20.Poláková K, Kŕčová M, Kuba D, Russ G. Analysis of HLA-G expression in malignant hematopoetic cells from leukemia patients. Leukemia Research. 2003;27(7):643–648. doi: 10.1016/s0145-2126(02)00228-x. [DOI] [PubMed] [Google Scholar]
- 21.Mizuno S, Emi N, Kasai M, Ishitani A, Saito H. Aberrant expression of HLA-G antigen in interferon γ-stimulated acute myelogenous leukaemia. British Journal of Haematology. 2000;111(1):280–282. doi: 10.1046/j.1365-2141.2000.02345.x. [DOI] [PubMed] [Google Scholar]
- 22.Nückel H, Rebmann V, Dürig J, Dührsen U, Grosse-Wilde H. HLA-G expression is associated with an unfavorable outcome and immunodeficiency in chronic lymphocytic leukemia. Blood. 2005;105(4):1694–1698. doi: 10.1182/blood-2004-08-3335. [DOI] [PubMed] [Google Scholar]
- 23.Rebmann V, Nückel H, Dührsen U, Grosse-Wilde H. HLA-G in B-chronic lymphocytic leukaemia: clinical relevance and functional implications. Seminars in Cancer Biology. 2007;17(6):430–435. doi: 10.1016/j.semcancer.2007.06.011. [DOI] [PubMed] [Google Scholar]
- 24.Yan W-H, Lin A, Chen B-G, et al. Unfavourable clinical implications for HLA-G expression in acute myeloid leukaemia. Journal of Cellular and Molecular Medicine. 2008;12(3):889–898. doi: 10.1111/j.1582-4934.2008.00175.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Gros F, Sebti Y, de Guibert S, et al. Soluble HLA-G molecules are increased during acute leukemia, especially in subtypes affecting monocytic and lymphoid lineages. Neoplasia. 2006;8(3):223–230. doi: 10.1593/neo.05703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Polakova K, Zeleznikova T, Russ G. HLA-G5 in the blood of leukemia patients and healthy individuals. Leukemia Research. 2013;37(2):139–145. doi: 10.1016/j.leukres.2012.10.019. [DOI] [PubMed] [Google Scholar]
- 27.Donadi EA, Castelli EC, Arnaiz-Villena A, Roger M, Rey D, Moreau P. Implications of the polymorphism of HLA-G on its function, regulation, evolution and disease association. Cellular and Molecular Life Sciences. 2011;68(3):369–395. doi: 10.1007/s00018-010-0580-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Hiby SE, King A, Sharkey A, Loke YW. Molecular studies of trophoblast HLA-G: polymorphism, isoforms, imprinting and expression in preimplantation embryo. Tissue Antigens. 1999;53(1):1–13. doi: 10.1034/j.1399-0039.1999.530101.x. [DOI] [PubMed] [Google Scholar]
- 29.O’Brien M, McCarthy T, Jenkins D, et al. Altered HLA-G transcription in pre-eclampsia is associated with allele specific inheritance: possible role of the HLA-G gene in susceptibility to the disease. Cellular and Molecular Life Sciences. 2001;58(12-13):1943–1949. doi: 10.1007/PL00000828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Rousseau P, Le Discorde M, Mouillot G, Marcou C, Carosella ED, Moreau P. The 14 bp deletion-insertion polymorphism in the 3′ UT region of the HLA-G gene influences HLA-G mRNA stability. Human Immunology. 2003;64(11):1005–1010. doi: 10.1016/j.humimm.2003.08.347. [DOI] [PubMed] [Google Scholar]
- 31.Tan Z, Randall G, Fan J, et al. Allele-specific targeting of microRNAs to HLA-G and risk of asthma. The American Journal of Human Genetics. 2007;81(4):829–834. doi: 10.1086/521200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Veit TD, Chies JAB. Tolerance versus immune response: MicroRNAs as important elements in the regulation of the HLA-G gene expression. Transplant Immunology. 2009;20(4):229–231. doi: 10.1016/j.trim.2008.11.001. [DOI] [PubMed] [Google Scholar]
- 33.Yie S-M, Li L-H, Xiao R, Librach CL. A single base-pair mutation in the 3′-untranslated region of HLA-G mRNA is associated with pre-eclampsia. Molecular Human Reproduction. 2008;14(11):649–653. doi: 10.1093/molehr/gan059. [DOI] [PubMed] [Google Scholar]
- 34.Alvarez M, Piedade J, Balseiro S, Ribas G, Regateiro F. HLA-G 3′-UTR SNP and 14-bp deletion polymorphisms in Portuguese and Guinea-Bissau populations. International Journal of Immunogenetics. 2009;36(6):361–366. doi: 10.1111/j.1744-313X.2009.00875.x. [DOI] [PubMed] [Google Scholar]
- 35.Castelli EC, Mendes-Junior CT, Deghaide NHS, et al. The genetic structure of 3′untranslated region of the HLA-G gene: polymorphisms and haplotypes. Genes and Immunity. 2010;11(2):134–141. doi: 10.1038/gene.2009.74. [DOI] [PubMed] [Google Scholar]
- 36.Rizzo R, Hviid TVF, Govoni M, et al. HLA-G genotype and HLA-G expression in systemic lupus erythematosus: HLA-G as a putative susceptibility gene in systemic lupus erythematosus. Tissue Antigens. 2008;71(6):520–529. doi: 10.1111/j.1399-0039.2008.01037.x. [DOI] [PubMed] [Google Scholar]
- 37.Baricordi OR, Govoni M, Rizzo R, Trotta F. In rheumatoid arthritis, a polymorphism in the HLA-G gene concurs in the clinical response to methotrexate treatment. Annals of the Rheumatic Diseases. 2007;66(8):1125–1126. doi: 10.1136/ard.2006.064022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Fainardi E, Rizzo R, Melchiorri L, et al. Presence of detectable levels of soluble HLA-G molecules in CSF of relapsing-remitting multiple sclerosis: relationship with CSF soluble HLA-I and IL-10 concentrations and MRI findings. Journal of Neuroimmunology. 2003;142(1-2):149–158. doi: 10.1016/s0165-5728(03)00266-2. [DOI] [PubMed] [Google Scholar]
- 39.Hviid TV, Hylenius S, Hoegh AM, Kruse C, Christiansen OB. HLA-G polymorphisms in couples with recurrent spontaneous abortions. Tissue Antigens. 2002;60(2):122–132. doi: 10.1034/j.1399-0039.2002.600202.x. [DOI] [PubMed] [Google Scholar]
- 40.Tripathi P, Abbas A, Naik S, Agrawal S. Role of 14-bp deletion in the HLA-G gene in the maintenance of pregnancy. Tissue Antigens. 2004;64(6):706–710. doi: 10.1111/j.1399-0039.2004.00308.x. [DOI] [PubMed] [Google Scholar]
- 41.Lin A, Yan WH, Dai MZ, et al. Maternal human leukocyte antigen-G polymorphism is not associated with pre-eclampsia in a Chinese Han population. Tissue Antigens. 2006;68(4):311–316. doi: 10.1111/j.1399-0039.2006.00667.x. [DOI] [PubMed] [Google Scholar]
- 42.Yan WH, Lin A, Chen XJ, et al. Association of the maternal 14-bp insertion polymorphism in the HLA-G gene in women with recurrent spontaneous abortions. Tissue Antigens. 2006;68(6):521–523. doi: 10.1111/j.1399-0039.2006.00723.x. [DOI] [PubMed] [Google Scholar]
- 43.Iversen A-C, Nguyen OTD, Tømmerdal LF, et al. The HLA-G 14bp gene polymorphism and decidual HLA-G 14bp gene expression in pre-eclamptic and normal pregnancies. Journal of Reproductive Immunology. 2008;78(2):158–165. doi: 10.1016/j.jri.2008.03.001. [DOI] [PubMed] [Google Scholar]
- 44.Hylenius S, Andersen A-MN, Melbye M, Hviid TVF. Association between HLA-G genotype and risk of pre-eclampsia: a case-control study using family triads. Molecular Human Reproduction. 2004;10(4):237–246. doi: 10.1093/molehr/gah035. [DOI] [PubMed] [Google Scholar]
- 45.Bermingham J, Jenkins D, McCarthy T, O’Brien M. Genetic analysis of insulin-like growth factor II and HLA-G in pre-eclampsia. Biochemical Society Transactions. 2000;28(2):215–219. doi: 10.1042/bst0280215. [DOI] [PubMed] [Google Scholar]
- 46.La Nasa G, Littera R, Locatelli F, et al. The human leucocyte antigen-G 14-basepair polymorphism correlates with graft-versus-host disease in unrelated bone marrow transplantation for thalassaemia. British Journal of Haematology. 2007;139(2):284–288. doi: 10.1111/j.1365-2141.2007.06779.x. [DOI] [PubMed] [Google Scholar]
- 47.Sizzano F, Testi M, Zito L, et al. Genotypes and haplotypes in the 3′ untranslated region of the HLA-G gene and their association with clinical outcome of hematopoietic stem cell transplantation for beta-thalassemia. Tissue Antigens. 2012;79(5):326–332. doi: 10.1111/j.1399-0039.2012.01862.x. [DOI] [PubMed] [Google Scholar]
- 48.Dong D-D, Hong Yang HY, Ke Li KL, et al. Human leukocyte antigen-G (HLA-G) expression in cervical lesions: association with cancer progression, HPV 16/18 infection, and host immune response. Reproductive Sciences. 2010;17(8):718–723. doi: 10.1177/1933719110369183. [DOI] [PubMed] [Google Scholar]
- 49.Ferguson R, Ramanakumar AV, Koushik A, Coutlée F, Franco E, Roger M. Human leukocyte antigen G polymorphism is associated with an increased risk of invasive cancer of the uterine cervix. International Journal of Cancer. 2012;131(3):E312–E319. doi: 10.1002/ijc.27356. [DOI] [PubMed] [Google Scholar]
- 50.Ferguson R, Ramanakumar AV, Richardson H, et al. Human leukocyte antigen (HLA)-E and HLA-G polymorphisms in human papillomavirus infection susceptibility and persistence. Human Immunology. 2011;72(4):337–341. doi: 10.1016/j.humimm.2011.01.010. [DOI] [PubMed] [Google Scholar]
- 51.He X, Dong D-D, Yie S-M, et al. HLA-G expression in human breast cancer: implications for diagnosis and prognosis, and effect on allocytotoxic lymphocyte response after hormone treatment in vitro. Annals of Surgical Oncology. 2010;17(5):1459–1469. doi: 10.1245/s10434-009-0891-9. [DOI] [PubMed] [Google Scholar]
- 52.Silva ID, Muniz YC, Sousa MC, et al. HLA-G 3'UTR polymorphisms in high grade and invasive cervico-vaginal cancer. Human Immunology. 2013;74(4):452–458. doi: 10.1016/j.humimm.2012.11.025. [DOI] [PubMed] [Google Scholar]
- 53.Mitelman F, editor. ISCN: Guidelines for Cancer Cytogenetics. Supplement to an International System for Human Cytogenetic Nomenclature. Basel, Switzerland: S. Karger AG; 1991. [Google Scholar]
- 54.Lacombe F, Durrieu F, Briais A, et al. Flow cytometry CD45 gating for immunophenotyping of acute myeloid leukemia. Leukemia. 1997;11(11):1878–1886. doi: 10.1038/sj.leu.2400847. [DOI] [PubMed] [Google Scholar]
- 55.Rizzo R, Hviid TVF, Stignani M, et al. The HLA-G genotype is associated with IL-10 levels in activated PBMCs. Immunogenetics. 2005;57(3-4):172–181. doi: 10.1007/s00251-005-0788-0. [DOI] [PubMed] [Google Scholar]
- 56.Rizzo R, Mapp CE, Melchiorri L, et al. Defective production of soluble HLA-G molecules by peripheral blood monocytes in patients with asthma. Journal of Allergy and Clinical Immunology. 2005;115(3):508–513. doi: 10.1016/j.jaci.2004.11.031. [DOI] [PubMed] [Google Scholar]
- 57.Shimizu Y, Geraghty DE, Koller BH, Orr HT, DeMars R. Transfer and expression of three cloned human non-HLA-A,B,C class I major histocompatibility complex genes in mutant lymphoblastoid cells. Proceedings of the National Academy of Sciences of the United States of America. 1988;85(1):227–231. doi: 10.1073/pnas.85.1.227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Guo QY, Chen BG, Ruan YY, Lin A, Yan WH. HLA-G expression is irrelevant to prognosis in patients with acute myeloid leukemia. Leukemia Research. 2011;35(10):1350–1354. doi: 10.1016/j.leukres.2011.05.036. [DOI] [PubMed] [Google Scholar]
- 59.Yan WH, Fan LA, Yang JQ, Xu LD, Ge Y, Yao FJ. HLA-G polymorphism in a Chinese Han population with recurrent spontaneous abortion. International Journal of Immunogenetics. 2006;33(1):55–58. doi: 10.1111/j.1744-313X.2006.00567.x. [DOI] [PubMed] [Google Scholar]
- 60.Feger U, Tolosa E, Huang Y-H, et al. HLA-G expression defines a novel regulatory T-cell subset present in human peripheral blood and sites of inflammation. Blood. 2007;110(2):568–577. doi: 10.1182/blood-2006-11-057125. [DOI] [PubMed] [Google Scholar]
- 61.Huang Y-H, Zozulya AL, Weidenfeller C, et al. Specific central nervous system recruitment of HLA-G+ regulatory T cells in multiple sclerosis. Annals of Neurology. 2009;66(2):171–183. doi: 10.1002/ana.21705. [DOI] [PubMed] [Google Scholar]
- 62.Poláková K, Bandžuchová E, Russ G. Impact of blood processing on estimation of soluble HLA-G. Neoplasma. 2011;58(4):337–342. doi: 10.4149/neo_2011_04_337. [DOI] [PubMed] [Google Scholar]
- 63.Rolfsen GB, Castelli EC, Donadi EA, Duarte RA, Soares CP. HLA-G polymorphism and breast cancer. International Journal of Immunogenetics. 2013 doi: 10.1111/iji.12092. [DOI] [PubMed] [Google Scholar]
- 64.Eskandari-Nasab E, Hashemi M, Hasani SS, Omrani M, Taheri M, Mashhadi MA. Association between HLA-G 3′UTR 14-bp ins/del polymorphism and susceptibility to breast cancer. Cancer Biomarkers. 2013;13(4):253–259. doi: 10.3233/CBM-130364. [DOI] [PubMed] [Google Scholar]