

Abstract
A single nucleotide polymorphism (SNP) rs1122608 on chromosome 19p13.2 and in the BRG1/SMARCA4 gene was previously associated with coronary artery disease (CAD). CAD and ischemic stroke are both associated with atherosclerosis. Thus, we tested the hypothesis that rs1122608 is associated with ischemic stroke. Further studies were used to identify the most likely mechanism by which rs1122608 regulates atherosclerosis. For case–control association studies, two independent Chinese Han GeneID cohorts were used, including a Central cohort with 1,075 cases and 2,685 controls and the Northern cohort with 1,208 cases and 824 controls. eQTL and real-time RT-PCR analyses were used to identify the potential candidate gene(s) affected by rs1122608. The minor allele T of SNP rs1122608 showed significant association with a decreased risk of ischemic stroke in the Central GeneID cohort (adjusted Padj = 2.1 × 10−4, OR 0.61). The association was replicated in an independent Northern GeneID cohort (Padj = 6.00 × 10−3, OR 0.69). The association became more significant in the combined population (Padj = 7.86 × 10−5, OR 0.73). Allele T of SNP rs1122608 also showed significant association with a decreased total cholesterol level (Padj = 0.013). Allele T of rs1122608 was associated with an increased expression level of SFRS3 encoding an mRNA splicing regulator, but not with the expression of BRG1/SMARCA4 or LDLR (located 36 kb from rs1122608). Increased expression of SFSR3 may decrease IL-1β expression and secretion, resulting in reduced risk of atherosclerosis and stroke. This is the first study that demonstrates that rs1122608 confers protection against ischemic stroke and implicates splicing factor SFSR3 in the disease process.
Introduction
Stroke is a common cause of death and morbidity worldwide (Dichgans 2007), but more than 60 % of stroke-related deaths occur in developing countries (Sacco et al. 2006). In China, there are 7.5 million stroke survivors and nearly 2.5 million new stroke cases are diagnosed each year (Johnston et al. 2009; Liu et al. 2011). A recent study showed that the total age-adjusted incidence of first-ever stroke in China did not differ from developed countries (Liu et al. 2007). Therefore, stroke is also a major health-care burden in China.
About 87 % of stroke cases are caused by ischemia (Donnan et al. 2008). Ischemic stroke can be caused by stenosis of large extracranial or intracranial arteries of the brain, small-vessel disease and cardioembolism (Markus 2011). The risk of ischemic stroke is influenced by genetic factors, environmental factors, and their interactions. The heritability of stroke subtypes is 40.3 % for large-vessel stroke, 32.6 % for cardioembolic stroke and 16.1 % for small-vessel stroke (Bevan et al. 2012). Genetic variants in a number of genes have been associated with ischemic stroke using either a genome-wide association study (GWAS) or a candidate case–control association study, including MTHFR, PAL-1, TNF, APOA, APOE, ACE, Factor V Leiden, ALOX5AP, Angiopoietin-1, CRP, CYP4AII, CYP4F2, CYPIIB2, DDAH1, NOS3, PCSK9, GP1BA, GPIIIa, IL-6, LTC4S2, MTHFR, NPY, PDE4D, SGK1, VKORC1, NINJ2, PRKCH, PITX2, ZFHX3, and the pro-thrombin gene (Casas et al. 2004; Ariyaratnam et al. 2007; Bevan et al. 2012). However, these risk loci may explain only a small portion of the total heritability for stroke, thus many more new loci or genetic variants need to be identified. Identification of a majority of stroke susceptibility genes may ultimately facilitate early diagnosis or risk prediction for stroke.
Ischemic stroke is caused by the thrombus or embolus that interrupts blood supply to the brain. Atherosclerosis in the brain is the major cause for the formation of thrombi and is estimated to cause 70–80 % of ischemic strokes or 60 % of all strokes (Markus 2011). Atherosclerosis in the coronary arteries causes coronary artery disease (CAD), and is considered as one of the most common causes in both ischemic stroke and CAD (Traylor et al. 2012). Ischemic stroke also shares some risk factors with CAD, such as smoking and hypertension (Cheng et al. 2012). Based on this rationale, we previously reported that a single nucleotide polymorphism (SNP) rs11206510 on chromosome 1p32 identified as a risk variant for CAD by GWAS was also associated with risk of ischemic stroke using the GeneID cohort for the Chinese population (Xu et al. 2010). The GeneID is one of the largest GeneBanks for cardiovascular and cerebrovascular diseases in China with more than 80,000 study subjects (Shi et al. 2009; Li et al. 2010; Xu et al. 2010; Cheng et al. 2011; Wang et al. 2011). In this study, we used the same GeneID population to assess whether SNP rs1122608 identified as a risk variant for CAD by GWAS was also associated with stroke.
SNP rs1122608 is located in the 30th intron of the SMARCA4 gene (also commonly referred to as BRG1). Interestingly, BRG1/SMARCA4 is located about 36 kb from the LDLR gene. In 2009, GWAS identified the association between SNP rs1122608 and myocardial infarction (MI) (Kathiresan et al. 2009). In 2010, a separate study showed that rs1122608 was also associated with CAD (Martinelli et al. 2010). BRG1/SMARCA4 is located on chromosome 19p13.2, contains 36 exons, and encodes the catalytic subunit of the SWI/SNF chromatin-remodeling complex, which influences transcriptional regulation by disrupting histone-DNA contacts in an ATP-dependent manner (Medina 2005). A recent study showed that overexpression or knockdown of BRG1/SMARCA4 affected H2S-induced inhibition of vascular smooth muscle cell proliferation (Li et al. 2013). In this study, we studied two independent case–control ischemic stroke cohorts with a total of 5,792 Chinese Han subjects (2,283 ischemic stroke cases and 3,509 controls from the GeneID database; cohort I with 1,075 cases and 2,685 controls from Central China; cohort II with 1,208 cases and 824 controls from Northern China). Allelic and genotypic association analyses were used to determine whether SNP rs1122608 was associated with ischemic stroke. We also analyzed whether SNP rs1122608 was associated with various lipid concentrations (risk factors for atherosclerosis). Analysis of expression quantitative trait loci (eQTL) was also carried out to identify the downstream gene(s) associated with rs1122608.
Materials and methods
Study subjects
All subjects were selected from GeneID, which is an ongoing Chinese population database that aims to collect DNA samples and clinical information for cardiovascular and cerebrovascular diseases national wide and to identify risk genes related to these diseases (Shi et al. 2009). The study was approved by local institutional review boards on human subjects and conformed to guidelines set forth by the Declaration of Helsinki. Written informed consent was obtained from subjects following instructions approved by the institutional review boards.
In the present study, a total of 5,792 subjects, including 2,283 ischemic stroke patients/cases and 3,509 non-stroke controls, were analyzed. For selection of stroke cases, only ischemic stroke patients were selected. The study subjects were from two geographically different study cohorts: Cohort I (China-Central) and Cohort II (China-Northern). Cohort China-Central consisted of 1,075 ischemic stroke cases and 2,685 controls, and was enrolled from hospitals in Central China, whereas Cohort China-Northern included 1,208 ischemic stroke cases and 824 comparable controls enrolled in hospitals from Northern China. For real-time qRT-PCR analysis, RNA samples from 127 randomly selected individuals from the control populations described above were used. All subjects were self-reported to be of Chinese Han origin.
We followed the World Health Organization criteria for the diagnosis of ischemic stroke. We also classified subjects based on a medical history of stroke, stroke signs by neurological examinations, and cerebral ischemia by CT or MRI images. We excluded patients with a relevant brain stem or subcritical hemispheric lesion with a diameter of <1.5 cm based on data from CT or MRI examinations. We also excluded the patients with arterial fibrillation associated with cardiologic embolic stroke. Comparable non-stroke controls were subjects free of stroke, lack of a medical history of stroke at the time of evaluation, and/or those who showed normal brain CT/MRI during examinations.
For each study cohort, non-stroke controls were matched by the same geographical area to minimize potential latent ancestral biases. Other clinical data including age, gender, smoking history, hypertension, and serum lipid concentrations were obtained from medical records. Hypertension was defined as a systolic blood pressure of ≥140 mm Hg or a diastolic blood pressure of ≥90 mm Hg. Total cholesterol (TC), high-density lipoprotein (HDL) cholesterol, and triglyceride (TG) concentrations were analyzed on an Olympus AU2700 Clinical Analyzer after overnight fasting, and low-density lipoprotein (LDL) cholesterol concentrations were calculated using the standard Friedewald formula.
Isolation of genomic DNA and SNP genotyping
Genomic DNA was extracted from peripheral blood samples using Wizard Genomic DNA Purification Kit (Promega, USA).
A total of 25 μL of PCR reaction mixture was prepared and contained a final concentration of 1.5 mM Mg2+, 0.2 mM dNTPs, 0.5 μM each primer (forward primer, 5′-TTT GTC ACA GTG CTC TCA TTT C-3′; reverse primer, 5′-CAG ATG AGA ACC CAG GTG TG-3′), 25 ng of human genomic DNA template, 5 μmol/LSYTO 9 green fluorescent and 0.15 U of Taq DNA polymerase (TIANGEN, China). PCR was performed on an ABI 9700 System with a thermal profile of 95 °C for 3 min, 40 cycles of 95 °C for 10 s, 58 °C for 10 s and 72 °C for 15 s, and a final step of 72 °C for 10 min. PCR amplicons harboring SNP rs1122608 were directly genotyped using high-resolution melting analysis (HRM) on our Rotor-Gene 6000 System (Corbett Life Science, Australia) under the standard protocols with minor modifications (Xu et al. 2010). During each run of HRM, we included three positive control DNA samples with known genotypes of GG, TG, TT, and one negative control of ddH2O. The success rate of HRM genotyping was 95.2 %. To validate the accuracy of HRM genotyping data, ten samples for each genotype of SNP rs1122608 were randomly selected for direct Sanger sequencing. The primers for sequencing include a forward primer of 5′-CGC CCC TGC CTG CTC ACT TC-3′ and a reverse primer of 5′-GCC CAG CCT CCA AGT GCA GTT T-3′. All sequencing results were completely consistent with genotypes determined by HRM analysis.
Expression quantitative loci (eQTL) analysis
In order to find the downstream target gene(s) of SNP rs1122608 that may be involved in the development of stroke, we searched the eQTL database at Center for Statistical Genetics of University of Michigan (http://www.sph.umich.edu/csg/liang/imputation/). The identified candidate eQTL was verified in the Chinese population using a panel of Chinese subjects with different genotypes of SNP rs1122608.
Total RNA samples were extracted from leukocytes from 127 individuals with different genotypes in the Chinese Han population using TRIzol (Invitrogen, USA), precipitated with isopropanol, and treated with RQ1 RNase-Free DNase (Promega, USA) to eliminate DNA contamination. The 127 study subjects were selected randomly from the mixed controls from the two study Cohorts. We used 1 μg of total RNA samples for reverse transcription with superscript II reverse transcriptase (Invitrogen, USA) and oligo (dT) (Kathiresan et al. 2009). The cDNA products were then used for real-time qPCR analysis with the SYBR Green Master Kit (Bio-Rad) and a 7900 HT Fast Real-Time PCR system (ABI).
The sequences for real-time qPCR primers are 5′-GCT CAT GGC TGA AGA TGA GG-3′ (forward primer) and 5′-CAG GCG TCT GTC CTT CTG C-3′ (reverse primer) for BRG1/SMARCA4, 5′-TAG AAT GGG CTG GTG TTG GGA GAC T-3′ (forward primer) and 5′-CTG GCT GCA GGT GTC GGG AT-3′ (reverse primer) for LDLR, 5′-TGA TGC AGT CCG AGA GCT AGA TG-3′ (forward primer) and 5′-GGA TCG GGA CGG CTT GTG-3′(reverse primer) for SFRS3, and 5′-AAG GTG AAG GTC GGA GTC AAC-3′ (forward primer) and 5′-GGG GTC ATT GAT GGC AAC AAT A-3′ (reverse primer) for GAPDH. GAPDH served as an internal standard. The RQ for the calibrator was set to 1. The calibrator was a mixed cDNA samples pooled from ten randomly selected individuals. The data were analyzed using RQ = 2−ΔΔCq (ΔΔCq is for the individual, ΔCq is for the calibrator) as described by us previously (Wang et al. 2011).
For qRT-PCR analysis, RQ values were transformed into standard scores (z scores) using SPSS v17.0. The scores below 2.0 and above −2.0 were considered to be effective values (Wang et al. 2011).
Statistical analyses
Statistical power analysis of study populations was conducted using program PS (Power and Sample size Calculations, version 3.0.43). Hardy–Weinberg linkage equilibrium was tested in each control group of two study cohorts using PLINK version 1.07 (Purcell et al. 2007).
Pearson χ2 tests and unpaired student t tests were performed for categorical traits (ischemic stroke) and continues traits (lipid levels), respectively, as implemented in program SPSS v17.0 (IBM Company). For allelic association analysis, 2 × 2 Pearson χ2 contingence tables were used to compare differences of frequencies of SNP rs1122608 minor allele T between the stroke group and the control group. Odds ratios (ORs) and corresponding 95 % confidential intervals (95 % CI) were also calculated. Genotypic association analysis was performed using 2 × 3 Pearson χ2 contingence tables. Multiple logistic regression analysis was used to adjust covariates such as sex, age, hypertension, smoking history, LDL-C, HDL-C, TC and TG levels.
To determine whether ORs differed between the two study cohorts, we performed a heterogeneity test using Comprehensive Meta Analysis version 2 (Borenstein et al. 2005).
For the association between lipid levels and SNP rs1122608, statistical analysis program SPSS (version 17.0) was used. Genotypic association was assessed by linear regression under additive, dominant and recessive genetic models.
Results
Significant ALLELIC association between SNP rs1122608 and stroke
Since SNP rs1122608 in the BRG1/SMARCA4 gene was associated with CAD, we hypothesized that it was also associated with stroke as both diseases share a similar cause, atherosclerosis. To test the hypothesis, we characterized SNP rs1122608 for its potential association with ischemic stroke in a Chinese population enrolled in a Central China city, Wuhan (Cohort China-Central). Cohort China-Central consists of 1, 075 ischemic stroke patients and 2,685 controls (Table 1). The demographical and clinical characteristics of the cases and controls are comparable (gender, smoking, age, and lipid levels) although significant differences were detected for gender, age, TC, LDL, and HDL levels (P < 0.05) (Table 1). The rate of hypertension, the most prevalent risk factor for stroke in the Chinese population, differed markedly and significantly between the two groups (71.9 % in cases and 13.8 % in controls) (Table 1).
Table 1.
Clinical and demographical characteristics of study subjects
| Characteristics | Cohort I (China-Central)
|
P value | Cohort II (China-North)
|
P value | Cohort I + Cohort II (combine)
|
P value | |||
|---|---|---|---|---|---|---|---|---|---|
| Ischemic strokes (n = 1075) | Controls (n = 2685) | Ischemic stroke (n = 1208) | Controls (n = 824) | Ischemic strokes (n = 2283) | Controls (n = 3509) | ||||
| Male, n (%) | 561 (52.2) | 1614 (60.1) | <0.001 | 651 (53.9) | 514 (62.4) | <0.001 | 1212 (53.1) | 2128 (60.6) | <0.001 |
| Hypertension, n (%) | 773 (71.9) | 371 (13.8) | <0.001 | 315 (26.1) | 158 (19.2) | 0.007 | 1088 (47.6) | 529 (15.1) | <0.001 |
| Smoking, n (%) | 207 (19.3) | 558 (20.8) | 0.29 | 389 (32.2) | 220 (26.7) | 0.006 | 596 (26.1) | 778 (22.2) | 0.007 |
| Age, years | 61.6 ± 14.5 | 57.4 ± 16.5 | <0.001 | 60.5 ± 10.8 | 58.1 ± 9.6 | 0.008 | 61.1 ± 12.7 | 57.5 ± 14.9 | <0.001 |
| Total cholesterol (TC, mmol/L) | 4.6 ± 1.3 | 4.5 ± 0.9 | 0.05 | 5.2 ± 1.6 | 4.8 ± 1.1 | 0.01 | 4.7 ± 1.4 | 4.6 ± 1.0 | 0.50 |
| LDL-C (mmol/L) | 2.7 ± 0.8 | 2.9 ± 0.9 | 0.002 | 3.0 ± 0.7 | 3.0 ± 0.5 | 0.89 | 2.8 ± 0.8 | 2.9 ± 0.8 | 0.026 |
| HDL-C (mmol/L) | 1.1 ± 0.3 | 1.2 ± 0.3 | <0.001 | 1.3 ± 0.5 | 1.2 ± 0.5 | 0.20 | 1.1 ± 0.4 | 1.2 ± 0.4 | <0.001 |
| Triglyceride (TG, mmol/L) | 1.5 ± 1.0 | 1.5 ± 1.3 | 0.38 | 1.5 ± 0.5 | 1.3 ± 0.5 | 0.004 | 1.5 ± 1.0 | 1.4 ± 1.0 | 0.14 |
For quantitative traits, data were shown as mean ± SD
Before the case–control association analysis, prior statistical power was estimated to assess whether Cohort China-Central had enough power for the proposed study. Given the type I error of 0.05 and a minor allele frequency (MAF) of 0.14 for SNP rs1122608 in the Chinese Han population (HapMap data), an OR of 0.82 for CAD (Martinelli et al. 2010), Cohort China-Central had a power of 73 % to detect the association.
SNP rs1122608 was genotyped in Cohort China-Central. Hardy–Weinberg equilibrium (HWE) tests of genotyping data indicated that Cohort China-Central did not deviate from a randomly mated population (P = 0.83). The minor allele T of SNP rs1122608 showed significant allelic association with a decreased risk of stroke (Pobs = 3.31 × 10−4, OR 0.71 for minor allele T) (Table 2). The association remained significant with an OR = of 0.61 (Padj = 2.1 × 10−4) after adjusting for covariates, including sex, age, hypertension, smoking history, LDL-C, HDL-C, TC and TG levels (Table 2).
Table 2.
Allelic association of SNP rs1122608 with stroke in the Chinese Han population
| Study population | Sample size | Phwe | Chi-square test
|
Logistic regression analysis
|
||||
|---|---|---|---|---|---|---|---|---|
| Case/control | Minor allele | Frequency | Pobs | OR (95 % CI) | Padj | OR (95 % CI) | ||
| Cohort I (China-Central) | 1,075/2,685 | 0.83 | T | 0.073/0.10 | 3.31E–04 | 0.71 (0.59–0.86) | 2.10E–04 | 0.61 (0.47–0.80) |
| Cohort II (China-North) | 1,208/824 | 0.18 | T | 0.088/0.12 | 2.63E–03 | 0.73 (0.59–0.90) | 6.00E–03 | 0.69 (0.54–0.90) |
| Combined | 2,283/3,509 | 0.36 | T | 0.081/0.10 | 3.57E–05 | 0.76 (0.67–0.87) | 7.86E–05 | 0.73 (0.62–0.85) |
Frequency frequency of the risk allele in case group/control groups, OR odds ratio; 95 % CI 95 % confidential interval
Phwe, P value for Hardy–Weinberg equilibrium (HWE) tests using PLINK version 1.07
Pobs, P value for association before adjusting for covariates by 2 × 2 contingence tables using PLINK version 1.07
Padj, P value for association after adjusting for covariates sex, age, Hyperion, smoking, LDL-C, HDL-C, TC and TG by multiple logistic regression analysis using SPSS v17.0
To validate the significant association between SNP rs1122608 and a decreased risk of stroke identified above, we enrolled the second set of cases and controls from Northern China (Cohort China-Northern). Cohort China-Northern consisted of 1,208 ischemic stroke patients and 824 controls, which are comparable in gender, smoking history, age, hypertension, and lipid levels although the significant differences were detected between the two groups except for LDL and HDL levels (Table 1). We tried our best to enroll stroke patients without hypertension in Cohort China-Northern so that the difference on the rate of hypertension in cases and controls did not differ too much as in Cohort China-Central. Cohort China-Northern also had a higher percentage of smokers than Cohort China-Central (Table 1).
Cohort China-Northern had a power of >90 % to verify the association between SNP rs1122608 and ischemic stroke identified in Cohort China-Central. Assumptions for power analysis for Cohort China-Northern were based on the data from Cohort China-Central (MAF of 0.10, OR of 0.61). The genotyping data in Cohort China-Northern did not deviate from Hardy–Weinberg equilibrium (P = 0.18). Significant association was identified between SNP rs1122608 and a decreased risk of ischemic stroke in Cohort China-Northern with a Pobs value of 2.63 × 10−3 and an OR of 0.73 (Table 2). After adjusting for covariates of sex, age, hypertension, smoking history, LDL-C, HDL-C, TC and TG levels, the association remained significant (Padj = 6.00 × 10−3, OR 0.69) (Table 2).
Comprehensive Meta Analysis (Version 2) found that the ORs did not differ between Cohort China-Central and Cohort China-Northern (heterogeneity P = 0.847, I-squared = 0), thus we combined the two Cohorts for further analysis. When combined, the association between SNP rs1122608 and a decreased risk of stroke became more significant (Pobs = 3.57 × 10−5, OR = 0.76; Padj = 7.86 × 10−5, OR = 0.73). Together, the data from two independent populations strongly suggest that the minor allele T of SNP rs1122608 plays a significant protective role in stroke in the Chinese Han population.
Significant genotypic association between SNP rs1122608 and stroke
Association analysis for genotypic frequencies may pinpoint potential genetic models for the action for a genetic variant in contrast to allelic association analysis. Thus, we assessed the association between the frequencies of different genotypes of SNP rs1122608 in ischemic stroke cases and controls. We analyzed the genotypic association under three common genetic models: an additive, a dominant, or a recessive model, and the results were summarized in Table 3.
Table 3.
Genotypic association of SNP rs1122608 with ischemic stroke under three different genetic models
| Study population | Modela | Genotypes (TT/TG/GG)
|
Chi-square test
|
Logistic regression analysis
|
|||
|---|---|---|---|---|---|---|---|
| Cases | Controls | Pobs | OR (95 % CI) | Padj | OR (95 % CI) | ||
| Cohort I (China-Central) | Additive | 10/136/929 | 25/485/2,175 | 2.83E–04 | n.a | 2.78E–04 | 0.62 (0.48–0.80) |
| Dominant | 146/929 | 510/2,175 | 7.76E–05 | 0.67 (0.55–0.82) | 1.60E–04 | 0.58 (0.44–0.77) | |
| Recessive | 10/1,065 | 25/2,660 | 1.00 | 1.00 (0.48–2.09) | 0.42 | 0.66 (0.24–1.79) | |
| Cohort II (China-North) | Additive | 6/200/1,002 | 7/178/639 | 8.98E–03 | n.a | 5.00E–03 | 0.68 (0.52–0.89) |
| Dominant | 206/1,002 | 185/639 | 2.44E–03 | 0.71 (0.57–0.88) | 4.00E–03 | 0.66 (0.50–0.87) | |
| Recessive | 6/1,202 | 7/817 | 0.33 | 0.58 (0.19–1.72) | 0.72 | 0.78 (0.21–2.92) | |
| Combined | Additive | 16/336/1,931 | 32/663/2,814 | 1.24E–04 | n.a | 1.47E–04 | 0.75 (0.63–0.86) |
| Dominant | 352/1,931 | 695/2,814 | 2.23E–05 | 0.74 (0.64–0.85) | 2.71E–04 | 0.73 (0.62–0.87) | |
| Recessive | 16/2,267 | 32/3,477 | 0.39 | 0.77 (0.42–1.40) | 0.70 | 0.52 (0.26–1.05) | |
OR odds ratio, 95 % CI 95 % confidential interval
Pobs, P value for association before adjusting for covariates by 2 × 2 contingence tables using PLINK version 1.07
Padj, P value for association after adjusting for covariates sex, age, Hyperion, smoking, LDL-C, HDL-C, TC and TG by multiple logistic regression analysis using SPSS v17.0
Additive model = TT/TG/GG; dominant model = TT + TG/GG; recessive model = TT/TG + GG
Significant genotypic association was identified between SNP rs1122608 and a decreased risk of ischemic stroke under an additive model or a dominant model, but not under a recessive model in Cohort China-Central before and after adjusting for covariates of sex, age, hypertension, smoking history, LDL-C, HDL-C, TC and TG levels (Pobs = 2.83 × 10−4, Padj = 2.78 × 10−4 under an additive model; Pobs = 7.76 × 10−5, Padj = 1.60 × 10−4 under a dominant model) (Table 3). The finding was confirmed in Cohort China-Northern (Pobs = 8.98 × 10−3, Padj = 5.00 × 10−3 under an additive model; Pobs = 2.44 × 10−3, Padj = 4.00 × 10−3 under a dominant model) (Table 3). The association became more significant in the combined Cohort China-Central and Cohort China-Northern population (Pobs = 1.24 × 10−4, Padj = 1.47 × 10−4 under an additive model; Pobs = 2.23 × 10−5, Padj = 2.71 × 10−4 under a dominant model) (Table 3).
Assessment of association between SNP rs1122608 and serum lipids levels
Since lipid levels are associated with atherosclerosis and some SNPs are associated with both lipid levels and atherosclerosis, we also used linear regression to analyze the potential association between SNP rs1122608 and lipid levels, including the plasma concentrations of TC, LDL-C, HDL-C and TG. Significant association was found for TC under an additive model and a recessive model in Cohort China-Central after adjusting for age, gender and hypertension (Padj = 0.04 and 1.87 × 10−3, respectively, Table 4). The association was confirmed in Cohort China-Northern (Padj = 0.02 and 0.04, respectively, Table 4) and in the combined population (Padj = 8.24 × 10−3 and 2.55 × 10−4, respectively, Table 4). No significant association was identified between SNP rs1122608 and LDL-C, HDL-C and TG levels at any genetic model (Padj > 0.05, Table 4). The mean serum total cholesterol levels were 4.66 ± 1.12 mmol/L for the GG genotype, 4.59 ± 0.94 mmol/L with for the GT genotype, and 3.84 ± 1.18 mmol/L with for the TT genotype. Thus, the minor allele T of SNP rs1122608 was associated significantly with a decreased level of TC concentrations.
Table 4.
Genotypic association between rs1122608 and serum lipids levels under three genetic models
| Lipid | Cohort |
Padj
|
Effect (â)
|
||||
|---|---|---|---|---|---|---|---|
| Additivea | Dominanta | Recessivea | Additivea | Dominanta | Recessivea | ||
| T C | Cohort I (China-Central) | 0.04 | 0.16 | 1.87E–03 | −0.10 (−0.20 to −0.003) | −0.07 (−0.18 to 0.03) | −0.67 (−1.10 to −0.25) |
| Cohort II (China-North) | 0.02 | 0.12 | 0.04 | −0.24 (−0.44 to −0.04) | −0.73 (−1.66 to 0.20) | −0.23 (−0.44 to −0.02) | |
| Combined | 8.24E–03 | 0.05 | 2.55E–04 | −0.12 (−0.20 to −0.03) | −0.09 (−0.19 to 7.88E–05) | −0.73 (−1.12 to −0.34) | |
| LDL-C | Cohort I (China-Central) | 0.38 | 0.42 | 0.59 | 0.05 (−0.06 to 0.16) | 0.05 (−0.07 to 0.16) | 0.13 (−0.34 to 0.60) |
| Cohort II (China-North) | 0.12 | 0.15 | 0.31 | −0.12 (−0.26 to 0.03) | −0.11 (−0.27 to 0.04) | −0.49 (−1.42 to 0.45) | |
| Combined | 0.95 | 0.97 | 0.87 | 0.12 (−3.30 to 3.55) | 0.07 (−3.59 to 3.73) | 1.32 (−14.63 to 17.26) | |
| HDL-C | Cohort I (China-Central) | 0.15 | 0.36 | 0.01 | 0.03 (0.07 to 0.01) | 0.02 (−0.02 to 0.06) | 0.22 (0.05–0.38) |
| Cohort II (China-North) | 0.91 | 0.86 | 0.82 | 0.005 (−0.08 to 0.09) | 0.008 (−0.08 to 0.10) | −0.05 (−0.43 to 0.34) | |
| Combined | 0.12 | 0.21 | 0.15 | 0.03 (−0.01 to 0.07) | 0.03 (−0.01 to 0.07) | 0.12 (−0.04 to 0.29) | |
| TG | Cohort I (China-Central) | 0.56 | 0.86 | 0.07 | 0.03 (−0.07 to 0.17) | 0.01 (−0.10 to 0.12) | −0.44 (−0.03 to 0.91) |
| Cohort II (China-North) | 0.72 | 0.96 | 0.14 | 0.02 (−0.07 to 0.10) | 0.002 (−0.09 to 0.09) | 0.29 (−0.10 to 0.67) | |
| Combined | 0.65 | 0.95 | 0.02 | 1.75 (−5.77 to 9.26) | −0.26 (−8.31 to 7.79) | 39.86 (6.10–73.60) | |
Padj, P value obtained from multiple linear modeling after adjustment for age, gender, and hypertension using SPSS v17.0
β, effect size related to the minor allele, using SPSS v17.0
Concentration unit, mmol/L for LDL-C, HDL-C, TC and TG, using SPSS v17.0
Additive model = TT/TG/GG; dominant model = TT + TG/GG; recessive model = TT/TG + GG
eQTL and real-time RT-PCR analyses identified significant association between SNP rs1122608 and the expression level of the SFRS3 gene
Single nucleotide ploymorphism rs1122608 is located within the 30th intron of BRG1/SMARCA4; therefore, we used real-time qRT-PCR analysis to assess whether SNP rs1122608 was associated with the expression level of BRG1/SMARCA4. We analyzed 127 total RNA samples isolated from leukocytes, and the data showed that there was no significant association between SNP rs1122608 and the expression level of BRG1/SMARCA4 (P = 0.633) (Fig. 1).
Fig. 1.

Assessment of the relationship between the risk allele T of SNP rs1122608 and the expression level of the BRG1/SMARCA4 mRNA (top panel) or the LDLR mRNA (bottom panel) assuming a dominant model
The next gene located closest to SNP rs1122608 is the LDLR gene, which is >36 kb downstream from BRG1 and plays a key role in atherosclerosis. Real-time RT-PCR analysis did not identify significant association between SNP rs1122608 and the expression level of LDLR (P = 0.513) (Fig. 1).
We then analyzed potential trans-eQTLs for SNP rs1122608. The eQTL analysis is a strategy for discovering the complex patterns correlating genotypic data of genetic variants to global microarray gene expression data from a population of individuals (Curtis et al. 2013). By searching the University of Michigan Center for Statistical Genetics eQTL database (http://www.sph.umich.edu/csg/liang/imputation/), we found that the common risk allele G of SNP rs1122608 was negatively associated with the expression level of the SFRS3 gene (Affymetrix HG U133 Plus 2.0 probe:232392_at, effect = −0.344, P = 0.00041) (Dixon et al. 2007). No other eQTL was identified. On the other hand, the data from the Genevar database showed no association between SNP rs1122608 and the expression level of the SFRS3 gene (P = 0.22–0.98) (http://www.sanger.ac.uk/resources/software/genevar/). Therefore, we carried out our own real-time qRT-PCR analysis to test whether SNP rs1122608 was associated with the expression level of SFRS3 in the Chinese population. Real-time qRT-PCR analysis showed that the expression levels of the SFRS3 mRNA were significantly different among the individuals with three different genotypes (P = 0.001) (Fig. 2). The expression level of the SFRS3 mRNA was significantly lower in 98 individuals with the GG genotype than that in 29 individuals with GT and TT genotypes (P = 0.01) (Fig. 2). Similarly, individuals with the GG or GT genotypes had significantly lower expression of the SFRS3 mRNA than those with the TT genotype (P = 0.0001) (Fig. 2). Together, these results suggest that the minor allele T of SNP rs1122608 is associated with an increased expression of SFRS3.
Fig. 2.
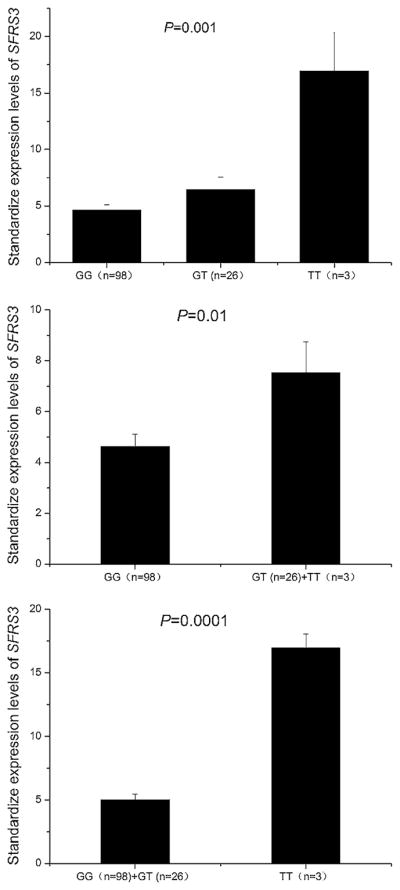
Significant association of the risk allele T of rs1122608 with decreased expression of SFRS3 mRNA assuming an additive model (top panel), a dominant model (middle panel) and a recessive model (bottom panel)
Discussion
In this study, we report a novel, highly significant association between SNP rs1122608 in the BRG1/SMARCA4 gene on chromosome 19p13 and a decreased risk of stroke. Several pieces of evidence strongly support this finding. Significant allelic association between SNP rs1122608 and a decreased risk of stroke was first identified in a Central Chinese population from GeneID with 1,075 ischemic stroke patients and 2,685 controls before and after adjusting for important covariates for stroke (Padj = 2.1 × 10−4) (Table 2). The minor allele T of SNP rs1122608 played a protective role in ischemic stroke (OR 0.61). Significant genotypic association was also identified under an additive or dominant inheritance model (Table 3). The novel association between SNP rs1122608 and a decreased risk of stroke was replicated in the second large Northern China population from GeneID with 1,208 ischemic stroke patients and 824 controls for both allelic frequencies and genotypic frequencies, and before and after adjustment for important covariates for stroke (Tables 2, 3). When the two sub-populations were combined, the P value for the association became more significant (Padj = 7.86 × 10−5, OR 0.73) (Tables 2, 3). To the best of our knowledge, this is the first time that significant association between SNP rs1122608 and a decreased risk of stroke was found.
SNP rs1122608 was first associated with MI by GWAS (Kathiresan et al. 2009) and later with CAD (Martinelli et al. 2010). The data from the present study indicate that SNP rs1122608 is associated with both CAD/MI and stroke by regulating atherosclerosis, the common cause for both CAD/MI and stroke. A previous study by Yang et al. (2010) attempted to assess the association between SNP rs1122608 and stroke in a Chinese population, but no significant association was identified. The contrasting results may be related to the small sample size utilized by Yang et al. (420 ischemic stroke patients and 467 controls). We performed power analysis and found that a cohort with 420 ischemic stroke patients and 467 controls would have a statistical power of 46 % assuming an OR of 0.73 derived from the present study for ischemic stroke and for rs1122608. This may explain the differences between the two studies.
In this study, we also assessed the association between SNP rs1122608 and various serum lipid concentrations. In both the GeneID Cohort China-Central and Cohort China-Northern, genotypic association between the minor allele T of rs1122608 and a decreased serum TC level was identified under an additive model or a recessive model, but no significant association was found for other lipid levels (Table 4). The reason underlying the contrasting associations with different lipid levels is not known; however, our positive association result is consistent with the data from a previous study showing significant association of a SNP in the BRG1/SMARCA4 gene with TC levels in a Doetinchem Cohort and in a meta-analysis of seven GWAS. We analyzed a linkage disequilibrium (LD) block for a 20-kb genomic region around SNP rs1122608 using software Haploview 4.2 for the genotyping data for the Chinese samples in the HapMap database. SNPs rs1122608 and rs7275 are in the same LD (D′ = 93 %; r2 = 0.86). These data are consistent with the similar findings in our study and the study by Lu et al. (2010).
To identify a potential mechanism by which SNP rs1122608 regulates stroke, we examined potential genes whose expression levers were associated with the variant. SNP rs1122608 is located within the BRG1/SMARCA4 gene and >36 kb from the LDLR gene, but we found that SNP rs1122608 was not associated the expression levels of either BRG1/SMARCA4 or LDLR. Interestingly, by searching publically available eQTL databases and follow-up real-time qRT-PCR analysis, we found that the minor allele T of SNP rs1122608, which protects against stroke, was associated with an increased expression of SFRS3 (Fig. 2). The SFRS3 gene is on chromosome 6p22.3 and encodes the serine/arginine-rich splicing factor 3 which is involved in RNA splicing. Knockdown of the SFRS3 expression was shown to increase the expression of interleukin-1β (IL-1β) mRNA and lead to increased IL-1β secretion (Valcarcel et al. 2011). Knockout mice lacking both apoE and IL-1β showed a decreased size of atherosclerotic lesions at the aortic sinus and the percentage of the atherosclerotic area to total area (Kirii et al. 2003). A recent study showed that exogenous administration of IL-1β to spontaneously hypertensive rats increased the incidence of stroke. Similarly, IL-1β injection to stroke-prone spontaneously hypertensive rats accelerated the onset of stroke (Chiba et al. 2012). Our results showed that the protective allele T of SNP rs1122608 was associated with an increased expression of SFRS3. Increased SFRS3 is expected to decrease the expression and secretion of IL-1β. An allele T-associated decrease of IL-1β expression and secretion could decrease the risk of atherosclerosis and stroke, thus the T allele plays a protective role against stroke.
SNP rs1122608 and SFRS3 are located on different chromosomes, 19p13 and 6p22.3, respectively. The expression level of SFRS3 is, therefore, considered as a trans-eQTL for SNP rs1122608. Trans-eQTLs are common for many SNPs, but the underlying molecular mechanisms are not clear. Similarly, the molecular mechanism by which SNP rs1122608 regulates the expression level of the SFRS3 mRNA is unknown. There are two possibilities. First, although we found that SNP rs1122608 was not associated with the expression level of BRG1 (Fig. 1), other SNP(s) in LD with rs1122608 on chromosome 19p13 may affect the expression of BRG1, a factor known to regulate transcriptional regulation by disrupting histone-DNA contacts. Altered expression of BRG1 may affect the expression of SFRS3. Second, SNP rs1122608 may affect the expression of another gene on 19p13 by long-range regulation, which in turn regulates the expression of SFRS3. Future studies are needed to distinguish these two different hypotheses.
In conclusion, the data in the present study, for the first time, identified SNP rs1122608 as a significant protective variant for stroke. Our results also suggest that the minor allele T of SNP rs1122608 decreases risk of atherosclerosis and stroke by increasing the expression levels of SFRS3 in the Chinese Han population, which leads to decreased expression and secretion of IL-1β and decreased risk of atherosclerosis and stroke. Finally, our study, for the first time, implicates a novel mRNA splicing regulator/mechanism into the development of stroke.
Acknowledgments
This work was supported by grants from the National Basic Research Program of China (973 Program: 2013CB531101 and 2012CB517800), the “Innovative Development of New Drugs” Key Scientific Project (2011ZX09307-001-09), and NIH R01 HL094498.
Contributor Information
Xin Xiong, Key Laboratory of Molecular Biophysics of the Ministry of Education, College of Life Science and Technology and Center for Human Genome Research, Cardio-X Institute, Huazhong University of Science and Technology, Wuhan, China.
Chengqi Xu, Key Laboratory of Molecular Biophysics of the Ministry of Education, College of Life Science and Technology and Center for Human Genome Research, Cardio-X Institute, Huazhong University of Science and Technology, Wuhan, China.
Yuting Zhang, Key Laboratory of Molecular Biophysics of the Ministry of Education, College of Life Science and Technology and Center for Human Genome Research, Cardio-X Institute, Huazhong University of Science and Technology, Wuhan, China.
Xiuchun Li, Key Laboratory of Molecular Biophysics of the Ministry of Education, College of Life Science and Technology and Center for Human Genome Research, Cardio-X Institute, Huazhong University of Science and Technology, Wuhan, China.
Binbin Wang, National Research Institute for Family Planning, Beijing, China.
Fan Wang, Key Laboratory of Molecular Biophysics of the Ministry of Education, College of Life Science and Technology and Center for Human Genome Research, Cardio-X Institute, Huazhong University of Science and Technology, Wuhan, China.
Qin Yang, Key Laboratory of Molecular Biophysics of the Ministry of Education, College of Life Science and Technology and Center for Human Genome Research, Cardio-X Institute, Huazhong University of Science and Technology, Wuhan, China.
Dan Wang, Key Laboratory of Molecular Biophysics of the Ministry of Education, College of Life Science and Technology and Center for Human Genome Research, Cardio-X Institute, Huazhong University of Science and Technology, Wuhan, China.
Xiaojing Wang, Key Laboratory of Molecular Biophysics of the Ministry of Education, College of Life Science and Technology and Center for Human Genome Research, Cardio-X Institute, Huazhong University of Science and Technology, Wuhan, China.
Sisi Li, Key Laboratory of Molecular Biophysics of the Ministry of Education, College of Life Science and Technology and Center for Human Genome Research, Cardio-X Institute, Huazhong University of Science and Technology, Wuhan, China.
Shanshan Chen, Key Laboratory of Molecular Biophysics of the Ministry of Education, College of Life Science and Technology and Center for Human Genome Research, Cardio-X Institute, Huazhong University of Science and Technology, Wuhan, China.
Yuanyuan Zhao, Key Laboratory of Molecular Biophysics of the Ministry of Education, College of Life Science and Technology and Center for Human Genome Research, Cardio-X Institute, Huazhong University of Science and Technology, Wuhan, China.
Dan Yin, Key Laboratory of Molecular Biophysics of the Ministry of Education, College of Life Science and Technology and Center for Human Genome Research, Cardio-X Institute, Huazhong University of Science and Technology, Wuhan, China.
Yufeng Huang, Key Laboratory of Molecular Biophysics of the Ministry of Education, College of Life Science and Technology and Center for Human Genome Research, Cardio-X Institute, Huazhong University of Science and Technology, Wuhan, China.
Xuan Zhu, Key Laboratory of Molecular Biophysics of the Ministry of Education, College of Life Science and Technology and Center for Human Genome Research, Cardio-X Institute, Huazhong University of Science and Technology, Wuhan, China.
Li Wang, Key Laboratory of Molecular Biophysics of the Ministry of Education, College of Life Science and Technology and Center for Human Genome Research, Cardio-X Institute, Huazhong University of Science and Technology, Wuhan, China.
Longfei Wang, Key Laboratory of Molecular Biophysics of the Ministry of Education, College of Life Science and Technology and Center for Human Genome Research, Cardio-X Institute, Huazhong University of Science and Technology, Wuhan, China.
Le Chang, Key Laboratory of Molecular Biophysics of the Ministry of Education, College of Life Science and Technology and Center for Human Genome Research, Cardio-X Institute, Huazhong University of Science and Technology, Wuhan, China.
Chaoping Xu, Key Laboratory of Molecular Biophysics of the Ministry of Education, College of Life Science and Technology and Center for Human Genome Research, Cardio-X Institute, Huazhong University of Science and Technology, Wuhan, China.
Hui Li, Key Laboratory of Molecular Biophysics of the Ministry of Education, College of Life Science and Technology and Center for Human Genome Research, Cardio-X Institute, Huazhong University of Science and Technology, Wuhan, China.
Tie Ke, Key Laboratory of Molecular Biophysics of the Ministry of Education, College of Life Science and Technology and Center for Human Genome Research, Cardio-X Institute, Huazhong University of Science and Technology, Wuhan, China.
Xiang Ren, Key Laboratory of Molecular Biophysics of the Ministry of Education, College of Life Science and Technology and Center for Human Genome Research, Cardio-X Institute, Huazhong University of Science and Technology, Wuhan, China.
Yanxia Wu, The First Hospital of Wuhan City, Wuhan, China.
Rongfeng Zhang, First Affiliated Hospital of Dalian Medical University, Dalian, China.
Tangchun Wu, Department of Occupational and Environmental Health, State Key Laboratory of Environmental Health for Incubation, School of Public Health, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, China.
Yunlong Xia, First Affiliated Hospital of Dalian Medical University, Dalian, China.
Yanzong Yang, First Affiliated Hospital of Dalian Medical University, Dalian, China.
Xu Ma, National Research Institute for Family Planning, Beijing, China.
Xin Tu, Email: xtu@mail.hust.edu.cn, Key Laboratory of Molecular Biophysics of the Ministry of Education, College of Life Science and Technology and Center for Human Genome Research, Cardio-X Institute, Huazhong University of Science and Technology, Wuhan, China.
Qing K. Wang, Email: wangq2@ccf.org, qkwang@mail.hust.edu.cn, Key Laboratory of Molecular Biophysics of the Ministry of Education, College of Life Science and Technology and Center for Human Genome Research, Cardio-X Institute, Huazhong University of Science and Technology, Wuhan, China. Department of Molecular Cardiology, Center for Cardiovascular Genetics, Lerner Research Institute, Cleveland Clinic, Cleveland, OH 44195, USA
References
- Ariyaratnam R, Casas JP, Whittaker J, Smeeth L, Hingorani AD, Sharma P. Genetics of ischaemic stroke among persons of non-European descent: a meta-analysis of eight genes involving approximately 32,500 individuals. PLoS Med. 2007;4:e131. doi: 10.1371/journal.pmed.0040131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bevan S, Traylor M, Adib-Samii P, Malik R, Paul NL, Jackson C, et al. Genetic heritability of ischemic stroke and the contribution of previously reported candidate gene and genomewide associations. Stroke. 2012;43:3161–3167. doi: 10.1161/STROKEAHA.112.665760. [DOI] [PubMed] [Google Scholar]
- Borenstein MHL, Higgins J, Rothstein H. Comprehensive meta-analysis Version 2. Biostat 2005 [Google Scholar]
- Casas JP, Hingorani AD, Bautista LE, Sharma P. Meta-analysis of genetic studies in ischemic stroke: thirty-two genes involving approximately 18,000 cases and 58,000 controls. Arch Neurol. 2004;61:1652–1661. doi: 10.1001/archneur.61.11.1652. [DOI] [PubMed] [Google Scholar]
- Cheng X, Shi L, Nie S, Wang F, Li X, Xu C, et al. The same chromosome 9p21.3 locus is associated with type 2 diabetes and coronary artery disease in a Chinese Han population. Diabetes. 2011;60:680–684. doi: 10.2337/db10-0185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheng YC, Anderson CD, Bione S, Keene K, Maguire JM, Nalls M, et al. Are myocardial infarction–associated single-nucleotide polymorphisms associated with ischemic stroke? Stroke. 2012;43:980–986. doi: 10.1161/STROKEAHA.111.632075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chiba T, Itoh T, Tabuchi M, Nakazawa T, Satou T. Interleukin-1β accelerates the onset of stroke in stroke-prone spontaneously hypertensive rats. Mediators Inflamm. 2012;2012:1–11. doi: 10.1155/2012/701976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Curtis RE, Kim S, Woolford JL, Jr, Xu W, Xing EP. Structured association analysis leads to insight into Saccharomyces cerevisiae gene regulation by finding multiple contributing eQTL hotspots associated with functional gene modules. BMC Genomics. 2013;14:196. doi: 10.1186/1471-2164-14-196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dichgans M. Genetics of ischaemic stroke. Lancet Neurol. 2007;6:149–161. doi: 10.1016/S1474-4422(07)70028-5. [DOI] [PubMed] [Google Scholar]
- Dixon AL, Liang L, Moffatt MF, Chen W, Heath S, Wong KC, et al. A genome-wide association study of global gene expression. Nat Genet. 2007;39:1202–1207. doi: 10.1038/ng2109. [DOI] [PubMed] [Google Scholar]
- Donnan GA, Fisher M, Macleod M, Davis SM. Stroke. Lancet. 2008;371:1612–1623. doi: 10.1016/S0140-6736(08)60694-7. [DOI] [PubMed] [Google Scholar]
- Johnston SC, Mendis S, Mathers CD. Global variation in stroke burden and mortality: estimates from monitoring, surveillance, and modelling. Lancet Neurol. 2009;8:345–354. doi: 10.1016/S1474-4422(09)70023-7. [DOI] [PubMed] [Google Scholar]
- Kathiresan S, Voight BF, Purcell S, Musunuru K, Ardissino D, Mannucci PM, et al. Genome-wide association of early-onset myocardial infarction with single nucleotide polymorphisms and copy number variants. Nat Genet. 2009;41:334–341. doi: 10.1038/ng.327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kirii H, Niwa T, Yamada Y, Wada H, Saito K, Iwakura Y, et al. Lack of interleukin-1beta decreases the severity of atherosclerosis in ApoE-deficient mice. Arterioscler Thromb Vasc Biol. 2003;23:656–660. doi: 10.1161/01.ATV.0000064374.15232.C3. [DOI] [PubMed] [Google Scholar]
- Li C, Wang F, Yang Y, Fu F, Xu C, Shi L, et al. Significant association of SNP rs2106261 in the ZFHX3 gene with atrial fibrillation in a Chinese Han GeneID population. Hum Genet. 2010;129:239–246. doi: 10.1007/s00439-010-0912-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li L, Liu D, Bu D, Chen S, Wu J, Tang C, et al. Brg1-dependent epigenetic control of vascular smooth muscle cell proliferation by hydrogen sulfide. Biochim Biophys Acta. 2013;1833:1347–1355. doi: 10.1016/j.bbamcr.2013.03.002. [DOI] [PubMed] [Google Scholar]
- Liu M, Wu B, Wang WZ, Lee LM, Zhang SH, Kong LZ. Stroke in China: epidemiology, prevention, and management strategies. Lancet Neurol. 2007;6:456–464. doi: 10.1016/S1474-4422(07)70004-2. [DOI] [PubMed] [Google Scholar]
- Liu L, Wang D, Wong KSL, Wang Y. Stroke and stroke care in China: huge burden, significant workload, and a national priority. Stroke. 2011;42:3651–3654. doi: 10.1161/STROKEAHA.111.635755. [DOI] [PubMed] [Google Scholar]
- Lu Y, Feskens EJM, Boer JMA, Imholz S, Verschuren WMM, Wijmenga C, et al. Exploring genetic determinants of plasma total cholesterol levels and their predictive value in a longitudinal study. Atherosclerosis. 2010;213:200–205. doi: 10.1016/j.atherosclerosis.2010.08.053. [DOI] [PubMed] [Google Scholar]
- Markus HS. Stroke genetics. Hum Mol Genet. 2011;20:R124–R131. doi: 10.1093/hmg/ddr345. [DOI] [PubMed] [Google Scholar]
- Martinelli N, Girelli D, Lunghi B, Pinotti M, Marchetti G, Malerba G, et al. Polymorphisms at LDLR locus may be associated with coronary artery disease through modulation of coagulation factor VIII activity and independently from lipid profile. Blood. 2010;116:5688–5697. doi: 10.1182/blood-2010-03-277079. [DOI] [PubMed] [Google Scholar]
- Medina PP. Transcriptional targets of the chromatin-remodelling factor SMARCA4/BRG1 in lung cancer cells. Hum Mol Genet. 2005;14:973–982. doi: 10.1093/hmg/ddi091. [DOI] [PubMed] [Google Scholar]
- Purcell S, Neale B, Todd-Brown K, Thomas L, Ferreira MAR, Bender D, et al. PLINK: a tool set for whole-genome association and population-based linkage analyses. Am J Hum Genet. 2007;81:559–575. doi: 10.1086/519795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sacco RL, Adams R, Albers G, Alberts MJ, Benavente O, Furie K, et al. Guidelines for prevention of stroke in patients with ischemic stroke or transient ischemic attack: a statement for healthcare professionals from the American Heart Association/American Stroke Association Council on Stroke: co-sponsored by the Council on Cardiovascular Radiology and Intervention: the American Academy of Neurology affirms the value of this guideline. Stroke. 2006;37:577–617. doi: 10.1161/01.STR.0000199147.30016.74. [DOI] [PubMed] [Google Scholar]
- Shi L, Li C, Wang C, Xia Y, Wu G, Wang F, et al. Assessment of association of rs2200733 on chromosome 4q25 with atrial fibrillation and ischemic stroke in a Chinese Han population. Hum Genet. 2009;126:843–849. doi: 10.1007/s00439-009-0737-3. [DOI] [PubMed] [Google Scholar]
- Traylor M, Farrall M, Holliday EG, Sudlow C, Hopewell JC, Cheng Y-C, et al. Genetic risk factors for ischaemic stroke and its subtypes (the METASTROKE Collaboration): a meta-analysis of genome-wide association studies. Lancet Neurol. 2012;11:951–962. doi: 10.1016/S1474-4422(12)70234-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Valcarcel J, Moura-Alves P, Neves-Costa A, Raquel H, Pacheco TR, D’Almeida B, et al. An shRNA-based screen of splicing regulators identifies SFRS3 as a negative regulator of IL-1β secretion. PLoS ONE. 2011;6:e19829. doi: 10.1371/journal.pone.0019829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang F, Xu C-Q, He Q, Cai J-P, Li X-C, Wang D, et al. Genome-wide association identifies a susceptibility locus for coronary artery disease in the Chinese Han population. Nat Genet. 2011;43:345–349. doi: 10.1038/ng.783. [DOI] [PubMed] [Google Scholar]
- Xu C, Wang F, Wang B, Li X, Li C, Wang D, et al. Minor allele C of chromosome 1p32 single nucleotide polymorphism rs11206510 confers risk of ischemic stroke in the Chinese Han population. Stroke. 2010;41:1587–1592. doi: 10.1161/STROKEAHA.110.583096. [DOI] [PubMed] [Google Scholar]
- Yang XC, Zhang Q, Li SJ, Wan XH, Zhong GZ, Hu WL, et al. Association study between three polymorphisms and myocardial infarction and ischemic stroke in Chinese Han population. Thromb Res. 2010;126:292–294. doi: 10.1016/j.thromres.2010.01.030. [DOI] [PubMed] [Google Scholar]