

Abstract
A variety of evidence suggests that, among humans, the individual tendency to choose immediate rewards (“Now”) over larger, delayed rewards (“Later”), or Now bias, varies with frontal dopamine (DA) levels. As cyclic elevations in estradiol (E+) modulate other frontal DA-dependent behaviors, we tested ovarian cycle effects on Now bias, and whether any such effects are E+ mediated. To do so, we quantified Now/Later choice behavior in naturally cycling adult females (n = 87; ages 18–40 years) during both the menstrual phase (MP; cycle day 1–2; low E+), and the follicular phase (FP; cycle day 11–12; high E+). Now bias decreased an average of 3.6% from MP to FP (p = 0.006). Measures of salivary E+ levels at each visit were available in a subsample of participants (n = 34). Participants with a verified E+ rise from MP to FP showed significantly greater decreases in Now bias at mid-cycle (n = 23) than those without a rise (n = 11; p = 0.03); Now bias decreased an average of 10.2% in the E+ rise group but increased an average of 7.9% in the no E+ rise group. The change in Now bias from MP to FP inversely correlated with the change in E+ (ρ = −0.39; p = 0.023), an effect driven by individuals with putatively lower frontal DA based on genotype at the Val158Met polymorphism in the COMT gene. This is the first demonstration that intertemporal choice varies across the ovarian cycle, with Now bias declining at mid-cycle, when fertility peaks. Moreover, our data suggest that the interacting effects of estradiol and frontal DA mediate this cycle effect on decision making.
Keywords: COMT, decision-making, delay discounting, dopamine, estrogen, impulsivity
Introduction
All animals, including humans, discount delayed rewards (Mazur, 1987; Rachlin, 2000), a tendency variously known as delay discounting (DD) or temporal discounting. While some degree of DD is normal, an excessive bias toward selecting immediate over larger delayed rewards, or “Now bias,” is associated with multiple clinical conditions, including substance abuse (Becker and Murphy, 1988; Reynolds, 2006; Perry and Carroll, 2008; Rogers et al., 2010), attention deficit hyperactivity disorder (Barkley et al., 2001; Sonuga-Barke et al., 2008; Paloyelis et al., 2010), and pathological gambling (Alessi and Petry, 2003; Leeman and Potenza, 2012). Thus, understanding the biological bases of such Now bias may have wide-reaching impact. Evidence from both humans and animals indicates that the individual DD tendency is heritable (Anokhin et al., 2011; Mitchell, 2011). However, animal studies also indicate that Now bias can be pharmacologically modulated, particularly by dopamine (DA; Dalley et al., 2008; Doya, 2008; Winstanley, 2011).
Accumulating evidence in humans also suggests that DA is a key regulator of Now bias. First, genetic variations in the DA system are associated with individual differences in Now bias (Boettiger et al., 2007; Eisenberg et al., 2007; Paloyelis et al., 2010; Smith and Boettiger, 2012), particularly variations in the gene encoding the catechol-O-methyltransferase (COMT) enzyme, which regulates tonic frontal DA (Karoum et al., 1994; Gogos et al., 1998; Slifstein et al., 2008; Käenmäki et al., 2010; Wu et al., 2012); putative lower tonic frontal DA is associated with greater Now bias among adults (Boettiger et al., 2007; Smith and Boettiger, 2012). Second, human pharmacology studies suggest that DA modulates Now bias, albeit with inconsistent results (de Wit et al., 2002; Acheson and de Wit, 2008; Hamidovic et al., 2008; de Wit, 2009; Pine et al., 2010), although these studies have not accounted for intrinsic variations in DA signaling that could interact with pharmacological effects. For example, we recently found that genetic variation in COMT modulates the effect of an acute DA manipulation of Now bias (Kelm and Boettiger, 2013). Recent data showing that COMT genotype interacts with cyclic estradiol (E+) changes to affect working memory (Jacobs and D'Esposito, 2011), which is also frontal DA dependent, suggests that cyclic variation in E+ could also modulate Now bias.
Data showing less DD among females relative to males (Bobova et al., 2009; Peper et al., 2013) support this idea, but no studies have investigated whether cyclic E+ fluctuations are associated with changes in Now bias. We hypothesized that if E+ modulates Now bias, naturally cycling females should demonstrate reduced Now bias from the menstrual phase (MP) to the follicular phase (FP). Moreover, such cycle effects should be modulated by COMT genotype. To test these ideas, we measured Now bias in a DD task among naturally cycling females (ages 18–40 years), during both the MP (cycle days 1–2; putative low E+) and the FP (cycle days 11–12; putative high E+). We also determined the COMT genotype of each subject and were able to measure free salivary E+ levels during each visit from a subset of participants.
Materials and Methods
Sample characteristics.
Participants (n = 91) were recruited from The University of North Carolina at Chapel Hill (UNC) and the surrounding community. Participants were healthy females 18–40 years old reporting no use of hormonal birth control and regular menstrual cycles of ∼28 d. Based on self-report, participants also had no known past or present neurological or psychiatric diagnoses (which would include premenstrual dysphoric disorder), including no history of substance use disorders, and no current use of psychoactive medications or other psychoactive substances aside from moderate use of caffeine, nicotine, or alcohol. All subjects were native English speakers and had at least a high-school education. Participants gave written informed consent, as approved by the UNC Office of Human Research Ethics. Subjects participated in two sessions, one on cycle days 1–2 (MP) and another on cycle days 11–12 (FP), in a counterbalanced, within-subject design. The menstrual cycle phase was based on participant self-report. All recruited participants self-reported regular cycles of ∼28 d during study screening; however, each participant's cycle was not explicitly tracked across multiple cycles. We did not exclude women who experience symptoms of premenstrual syndrome (PMS), although concerns on this point are somewhat mitigated by the fact that PMS symptoms are present during the luteal phase (LP), beginning 1–2 weeks before menses and completely resolving at menses onset (Rubinow and Schmidt, 2006), and we did not test women during the symptomatic phase of the cycle. In addition to the behavioral testing (see Delay discounting task), during session 1 (n = 45 MP, n = 46 FP) we collected information on participants' age, years of education, trait impulsiveness [Barratt Impulsiveness Scale-11 (BIS); Patton et al., 1995], orientation toward the future (Future Time Perspective Inventory; Wallace, 1956), and locus of control (Rotter's Locus of Control Scale; Rotter, 1966).
COMT genotyping.
COMTVal158Met (rs4680) genotyping was performed on DNA extracted from saliva samples (DNA Genotek) using TaqMan technology (Life Technologies), as previously described (Boettiger et al., 2007; Smith and Boettiger, 2012).
Delay discounting task.
During each testing session, participants completed a DD task, which has been described in detail previously (Altamirano et al., 2011; Smith and Boettiger, 2012). Subjects were given task instructions, completed a short practice, and then completed eight blocks of 42 trials each during each test session. In each session, subjects made a series of choices between smaller, sooner (“Now”) and larger, later (“Later”) hypothetical monetary rewards. Each trial began with an instruction cue, followed by two options. In each trial, the Later option was one of five amounts ($2, $5, $10, $20, or $100) at one of five future delays (1 week, 2 weeks, 1 month, 3 months, or 6 months), and the Now option was an amount discounted by 70, 85, 90, or 95% from the Later amount, available “TODAY.” The instruction cue was determined by trial type. There were four trial types: WANT, DON′T WANT, SOONER, and LARGER, with the latter two conditions considered together as control (CON) trials. Accuracy in these control conditions verifies that individuals are adhering to task instructions, while comparison of reaction times (RTs) among the CON, WANT, and DON′T WANT trial types indicates whether additional cognitive processes are being engaged in the WANT and DON′T WANT conditions, relative to the simple objective comparison needed for CON trials. A pattern of RTs that deviates from CON < WANT < DON′T WANT indicates that the task is not being performed as instructed, and resulted in exclusion from all analyses (see below). Trial types were pseudorandomly ordered and weighted, with one-half WANT trials and one-sixth each of the other trial types. The WANT condition was most frequent, as choice in that condition was our primary interest here. Moreover, this weighting also promotes a prepotent tendency to select the preferred option, requiring inhibition in the DON′T WANT condition. Participants indicated their preferred option on WANT trials, their nonpreferred option on the DON′T WANT trials, and the side with the sooner time or larger amount of money for SOONER and LARGER (CON) trials, respectively. The Later amount, delay time, percentage discount, and left/right position were pseudorandomly selected for each trial. The RT for each response was also collected. Four subjects were excluded from all analyses based on deviation from the expected RT pattern across trial types (see above) in one or more sessions, leaving 87 participants. Of these, 44 participants were first tested in the MP, and 43 participants were first tested in the FP.
Our primary index of Now bias was the proportion of Now choices made in the WANT condition, the impulsive choice ratio (ICR). From the DON′T WANT trials, we determined the inferred ICR (iICR) as a function of delay time, and calculated the average of the absolute value of the difference between ICR and iICR at each delay. This value provides a measure of response consistency, termed “motor mismatch,” with larger values indicating less controlled response selection (Mitchell et al., 2007). Change in ICR from the MP to FP was calculated as a simple subtraction of the ICR in the FP session from the ICR in the MP session.
In addition to ICR, we also quantified the degree of impulsive choice using the q-exponential discount function based on Tsallis' statistics (Takahashi et al., 2008; Takahashi, 2009), as follows:
where D represents delay time, and kq and q are measures of impulsivity and of inconsistency across delay times, respectively. To estimate kq and q, we conducted nonlinear curve fitting with the Levenberg–Marquardt algorithm implemented in MATLAB (MathWorks, Curve Fitting Toolbox, Custom Equation option). Discounted Value was calculated as the cumulative selected/maximum dollar ratio at each delay, D. In addition to estimating kq and q, we can also quantify the degree to which impatience declines as a function of delay time [decreasing impatience (DI)], as proposed by Prelec (2004) and implemented within the q-exponential discount function (Takahashi, 2011), as follows:
where D, kq, and q are as defined for Equation 1. We excluded n = 37 subjects from these analyses based on an inadequate fit by the q-exponential model (defined as an adjusted R2 value of <0.2 for one or both sessions). For the subjects from whom we had E+ data, our sample was reduced to n = 10 and therefore was underpowered for any E+ change analyses.
Salivary estradiol quantification.
To confirm and quantify E+ changes from the MP to the FP in our participants, we obtained permission to collect saliva samples at each visit and were able to collect these data from 34 of our 87 participants. There were no significant differences between the participants from whom we obtained salivary E+ measures and those we did not, in terms of ICR (p = 0.61), demographics (minimum p = 0.46), or psychometric measures (minimum p = 0.09). The use of salivary E+ was based first on the noninvasive and less stressful nature of this collection method (relative to serum collection). Given data showing that cortisol, a physiological mediator of stress responses, can impact discounting behavior (Takahashi, 2004; Takahashi et al., 2010; Kawamura et al., 2013), avoiding the expected stress associated with an actual or anticipated blood draw in some subjects, served to minimize unwanted variance in our data. Second, empirical data support the correlation between salivary and serum E+ levels (Lu et al., 1999). Three saliva samples were collected during each session from participants via passive drool into 15 ml tubes over a period of ∼1 h while they completed questionnaires and/or during task breaks. Samples were pooled and stored at −20°C until analysis. Upon thawing, saliva samples were centrifuged at 1500 × g for 15 min to separate mucous material and pellet out any potential contaminants. Cleared saliva was then decanted into a separate tube. We quantified salivary estradiol via an enzyme immunoassay kit (Salimetrics). On each plate, pooled samples were tested in duplicate, with samples from both visits for each subject tested on the same plate. Optical densities were measured using a μQuant microplate spectrophotometer (BioTek) and, following kit procedure, were transformed into picogram per milliliter E+ values based on plate-specific four-parameter sigmoid minus curves derived from standard E+ samples. E+ concentrations for each sample were averaged across plates. Only individuals with higher average E+ levels in their FP sample than in their MP sample were classified as showing a positive change in E+ (ΔE+). The proportional change in E+ value used in our correlation analyses reflect the difference in E+ between the FP and MP samples divided by MP E+ (ΔE+ = (FPE+ − MPE+)/MPE+).
Statistical analysis.
As the ICR, kq, q, and DI data in this sample were not normally distributed, the Wilcoxon signed-rank test was used for comparisons across sessions. Paired t tests and repeated-measures ANOVAs were used to test for the effect of cycle phase on other DD task measures as well as on arcsine-root transformed ICR values. Group comparisons of ICR change were made using unpaired t tests. Pearson's r or Spearman's ρ were used for correlation analyses, as indicated. We used bootstrapping where indicated to eliminate concerns about violations of parametric assumptions, and 95% CIs are reported. Where sphericity was violated, we applied the Greenhouse-Geisser correction.
Results
Sample characteristics
Eighty-seven healthy females from the Chapel Hill/Durham, NC, area volunteered and were paid for their participation (Table 1). In the sample as a whole, we observed a positive correlation (r(85) = 0.22; p = 0.040) between Now bias measured in session 1 and the nonplanning subscale of the BIS, a trait measure of impulsivity, which is consistent with some previous findings (Mitchell et al., 2005; de Wit et al., 2007).
Table 1.
Demographic data
| Total sample (n = 87) | MP first (n = 44) | FP first (n = 43) | t(85), p | |
|---|---|---|---|---|
| Age (years) | 24 ± 5 | |||
| Education (years) | 16 ± 2 | |||
| Psychometric measures | ||||
| BIS total score | 60.2 ± 11.7 | 60.6 ± 11.6 | 59.8 ± 12.0 | 0.32, 0.75 |
| BIS attention | 16.4 ± 4.1 | 16.6 ± 4.1 | 16.3 ± 4.2 | 0.35, 0.73 |
| BIS motor | 21.6 ± 4.2 | 22.0 ± 3.9 | 21.3 ± 4.6 | 0.80, 0.43 |
| BIS planning | 22.2 ± 5.4 | 22.1 ± 5.4 | 22.3 ± 5.5 | −0.20, 0.84 |
| FTPI | ||||
| Mean extension | 6.9 ± 5.2 | 7.2 ± 6.0 | 6.7 ± 4.3 | 0.46, 0.64 |
| Maximum extension | 23.7 ± 18.2 | 25.6 ± 20.7 | 21.7 ± 15.3 | 0.98, 0.33 |
| LOC | 11.0 ± 4.0 | 11.1 ± 4.6 | 11.0 ± 3.3 | 0.13, 0.89 |
| Genetics (%) | ||||
| COMT Val158Met genotype | ||||
| Met/Met | 19.5 | |||
| Met/Val | 44.8 | |||
| Val/Val | 35.6 |
Values are reported as the mean ± SD, unless otherwise indicated. FTPI, Future Time Perspective Inventory; LOC, Rotter's Locus of Control Scale.
Impulsive choice declines from early to mid-cycle
We predicted a decline in our measure of Now bias, ICR (see Materials and Methods) from the MP to the FP, which was indeed observed. Across the participant group as a whole, we found a significant effect of cycle phase on ICR, with higher ICRs in the MP (median, 0.72) relative to the FP (median, 0.66) using either a Wilcoxon signed-rank test on the raw ICR values (z = −2.77; p = 0.006; r = −0.21), or a paired-samples t test on arcsine-root transformed ICR data (t(86) = 2.13; p = 0.036; Fig. 1A). Although we counterbalanced session order, we confirmed that this cycle effect remained significant after covarying for session order in a repeated-measures ANOVA (F(1,85) = 4.65; p = 0.034; η2 = 0.049).
Figure 1.

Impatient choice declines from early to mid-cycle. A, Now bias declines from early to mid-cycle. The plot depicts an index of Now bias, ICR, measured during the MP (cycle days 1–2) and the FP (cycle days 11–12) within subjects. ICR values were significantly reduced at mid-cycle (FP) relative to the MP (*t(86) = 2.13; p = 0.036). B, Decreasing Impatience, DIq is plotted as a function of delay time. DIq is elevated in the MP relative to the FP (F(1,49) = 5.56; p = 0.022), and, while DIq declines as a function of delay in both the MP and FP, DIq declines more steeply in the MP. Values reflect the mean ± SEM.
This DD task includes objective choice CON trials; accuracy in these trials did not differ significantly between the MP and FP sessions (t(86) = −0.66; p = 0.51). The task also includes a control condition (DON′T WANT) in which participants are instructed to select the monetary reward option that they do not prefer. Comparing ICR in the WANT trials to inferred ICR in the DON′T WANT trials provides a measure of response consistency. We observed a significant effect of cycle on response consistency or motor mismatch (see Materials and Methods), with greater mismatch in the FP (0.13 ± 0.08) relative to the MP (0.10 ± 0.06; t(86) = −2.63; p = 0.010). Importantly, our observed cycle effect on Now bias remained significant after covarying for changes in response consistency in a repeated-measures ANOVA (F(1,85) = 6.02; p = 0.016; η2 = 0.066), indicating that a drop in response consistency cannot explain the decrease in Now bias at mid-cycle. We observed no cycle effects on RT in the objective choice (CON) trials (t(86) = 1.28; p = 0.21), subjective choice trials (WANT; t(86) = −0.01; p = 0.99), or DON′T WANT trials (t(86) = −0.27; p = 0.79).
As a measure of impulsive choice, ICR has several advantages. First, it has very strong internal reliability. For the present dataset, the Cronbach's α for ICR ranged between 0.977 and 0.995 across the various conditions, indicating an extremely high level of reliability. Second, ICR is an assumption-free metric, rendering it less sensitive to errors introduced by incorrect models. These strengths are of particular importance when making comparisons across conditions, as in the present study. In contrast, discounting rates (k) are derived via curve-fitting using discounting models and depend strongly on both the assumptions of the particular model and on the variability of the underlying data from which such fits are derived. Despite these caveats, recent studies have demonstrated the utility of the q-exponential discount function in parameterizing both Now versus Later preference (impulsivity; kq) and the inconsistency (q) in such Now bias across delay times in intertemporal choice tasks (Takahashi et al., 2008; Takahashi, 2009). Consistent with our ICR-based results, we found that kq decreased significantly in the FP, relative to the MP; in contrast, we did not observe a significant change in q from early-to mid-cycle (Wilcoxon sign-rank test: kq, p = 0.032; q, p = 0.188; Table 2). Perhaps not surprisingly, given these findings, kq was strongly correlated with ICR (Spearman's ρ = 0.850; 95% CI, 0.759–0.909; p < 0.001), while the correlation between ICR and q was much weaker and in the opposite direction (Spearman's ρ = −0.274; 95% CI, −0.069 to −0.446; p = 0.006). We also quantified the time decay of the discount rate (kq) across delay times using Prelec's measure of decreasing impatience (DI) (2004), as implemented within the q-exponential discount function by Takahashi (2011). A repeated-measures ANOVA (Cycle Phase × Delay Time) found significant main effects of both Cycle Phase (F(1,49) = 5.56; p = 0.022; η2 = 0.015) and Delay Time (F(1.02,50.09) = 69.7; p < 0.001; η2 = 0.412), and a significant interaction between Cycle Phase and Delay Time (F(1.02,50.14) = 4.33; p = 0.042; η2 = 0.011; Fig. 1B). Average DIq across delay times was higher in the MP (mean, 0.0229; 95% CI, 0.0183–0.0277) versus the FP visit (mean, 0.0171; 95% CI, 0.0127–0.0218; t(49) = 2.36; p = 0.022). We confirmed the effect of delay time on DIq with a Friedman's test on DIq change scores (FP − MP) across delay times (χ2 = 11.52; p = 0.021; df = 4). Post hoc paired comparisons of DIq at each delay time in the MP and FP visits showed significant decreases in DIq from the MP to the FP at all five delays (mean decreases, bootstrapped: 7 d, 0.0145, t = 2.15, p = 0.036; 14 d, 0.0083, t = 2.39, p = 0.013; 30 d, 0.0043, t = 2.57, p = 0.005; 90 d, 0.0015, t = 2.57, p = 0.012; 180 d, 0.0007, t = 2.43, p = 0.017). These data indicate that the irrationality of intertemporal choice declines from the MP to the FP, with the greatest cycle effects at the shortest delay and diminishing cycle effects with increasing delay times.
Table 2.
Estimated parameters in q exponential discounting by cycle phase
| MP | FP | z score | p value | |
|---|---|---|---|---|
| kq (impulsivity) | 0.002488 | 0.001242 | −2.148 | 0.032* |
| q (consistency) | −20.6 | −13.91 | −1.318 | 0.188 |
Median parameter values are given by cycle phase. Smaller kq indicates less impulsive intertemporal choice at D = 0. Less negative q indicates a more consistent intertemporal choice.
*p < 0.05.
The mid-cycle drop in Now bias is associated with an E+ rise
While the decrease in Now bias from the MP to the FP is consistent with a hypothetically E+-mediated effect, hormonal fluctuation across the cycle varies, even among women with regular cycles. To more directly assess the relationship between changes in E+ and changes in Now bias, we assayed salivary E+ from both sessions from a subset of participants (n = 34). On average, E+ rose (0.50 ± 0.23 pg/ml) from the MP to the FP (t(33) = −2.20; p = 0.035). However, of the 34 participants, only 23 demonstrated a detectable increase in E+ from the MP to the FP (E+ rose 1.15 ± 0.89 pg/ml; t(22) = −6.20; p < 0.001); critically, these 23 participants also showed a significant decrease in Now bias (−0.07 ± 0.03) from the MP to the FP (t(22) = −2.18; p = 0.041; Fig. 2). In contrast, in those participants without a detectable rise in E+ (n = 11; E+ decreased by 0.85 ± 1.07 pg/ml; t(10) = 2.62; p = 0.026) Now bias tended to rise from the MP to FP (0.05 ± 0.05; t(10) = 1.09; p = 0.303; Fig. 2). The cycle effect on Now bias differed significantly between the E+ rise and the no-E+ rise participants (t(32) = 2.145; p = 0.040; Fig. 2).
Figure 2.
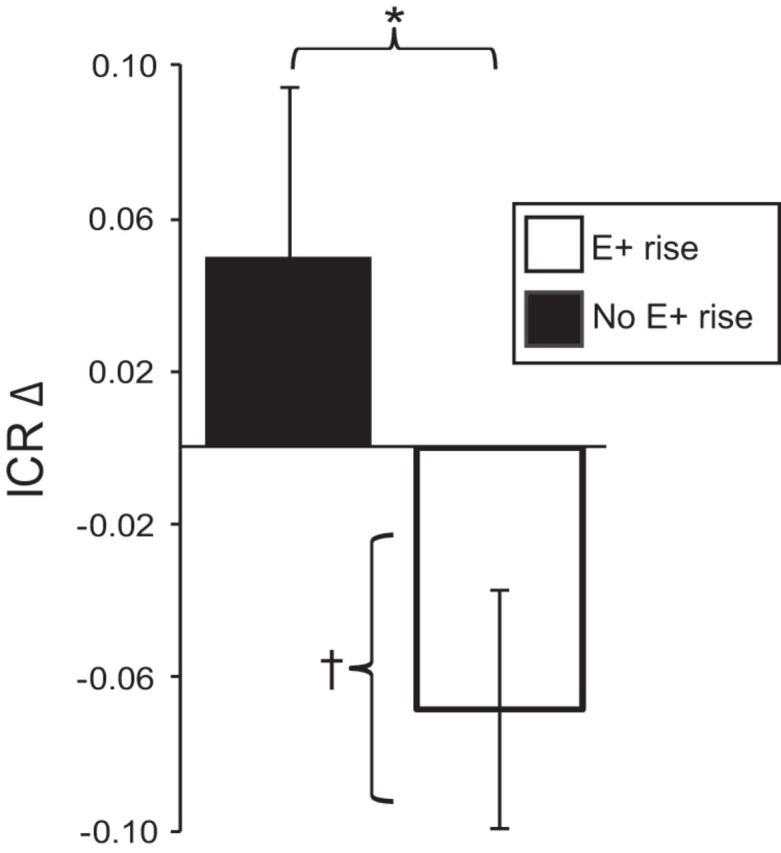
Now bias declines at mid-cycle when E+ rises at mid-cycle. The plot depicts the change in ICR from early to mid-cycle in participants with a verified rise in E+ levels at mid-cycle (E+ rise), and those without a detectable rise in E+ levels at mid-cycle (No E+ rise). ICR declines significantly at mid-cycle in the E+ rise group (†t(22) = −2.18; p = 0.041). The change in ICR from early to mid-cycle differed significantly between the E+ rise and no-E+ rise groups (*t(32) = 2.145; p = 0.040).
Changes in Now bias from the MP to FP are inversely related to changes in E+
As we observed substantial variation in E+ changes from the MP to the FP, we tested whether the magnitude of the E+ change between sessions was correlated with individual change in Now bias. To avoid concerns about violating parametric assumptions, we calculated the Spearman's ρ between the proportional change in E+ from the MP to the FP and the change in ICR from the MP to the FP, and conducted a robust regression analysis procedure using bootstrapping. We found a significant negative correlation between E+ change and ICR change (ρ(32) = −0.39; 95% CI, −0.67 to −0.06; p = 0.023; Fig. 3, solid line), suggesting a role for E+ in mediating the observed cycle effects on Now bias.
Figure 3.
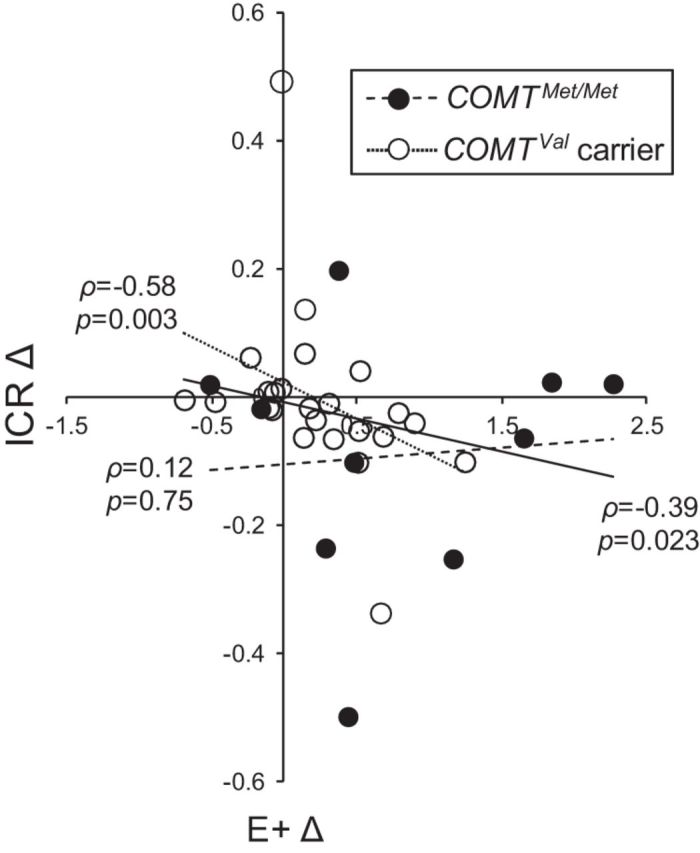
The change in ICR from early to mid-cycle is proportional to the change in E+ and is driven by putatively lower tonic frontal dopamine COMTVal carriers. In the sample as a whole, ICR decreases correlate with E+ increases (ρ = −0.39; p = 0.023, solid line). This effect is driven by COMTVal allele carriers (open circles; ρ = −0.58; p = 0.003, dotted line). Changes in E+ levels and ICR from MP to FP are not correlated in individuals with putatively high tonic frontal dopamine levels (COMTMet/Met, filled circles; ρ = 0.12; p = 0.75, dashed line).
E+ in session 1 correlates with trait impulsivity
Based on our findings that session 1 ICR positively correlated with BIS nonplanning scores, and that ICR fluctuated from the MP to the FP in tandem with E+ changes, we tested whether E+ levels in session 1 correlated with BIS nonplanning scores. We found that, indeed, each additional picogram per milliliter of E+ measured in session 1 was associated with 0.79 points lower BIS nonplanning scores (r(32) = −0.42; 95% CI, −0.63 to −0.20; p = 0.014). A similar relationship between session 1 E+ levels and total BIS scores (r(32) = −0.36; p = 0.036) was also observed.
COMT genotype modulates E+ effects on Now bias
As the effects of E+ changes from the MP to FP on working memory, which is also frontal DA dependent, are modulated by the COMT Val158Met genotype (Jacobs and D'Esposito, 2011), we tested whether the observed relationship between E+ elevation and reduced Now bias is also modulated by COMT genotype. Of the participants from whom we had E+ measures, the distribution of COMT genotypes was as follows: 10 methionine (Met)/Met, 9 Met/valine (Val), 15 Val/Val. Unlike the full sample, within the Met/Met group, we observed a statistically insignificant increase in ICR with greater E+ rises (ρ(8) = 0.12; p = 0.75; 95% CI, −0.68 to 0.70; Fig. 3, dashed line). However, in the Met/Val group, we observed a trend toward greater mid-cycle declines in ICR with greater E+ rises (ρ(7) = −0.63; p = 0.067; 95% CI, −1.0 to 0.24); Val homozygotes showed a similar negative relationship (ρ(13) = −0.29; p = 0.302; 95% CI, −0.73 to 0.34). This similarity between the latter two groups is consistent with data showing that Val carrier status predicts elevated impulsivity in females (Qian et al., 2003; Lang et al., 2007), and impaired executive function (Small et al., 2011) and hypothalamic-pituitary-adrenal axis dysfunction (Fernández-de-Las-Peñas et al., 2012) in female breast cancer survivors. As such, we considered Val allele carriers together and found significantly greater mid-cycle declines in ICR with greater mid-cycle E+ rises among COMTVal carriers (ρ(22) = −0.58; p = 0.003; 95% CI, −0.78 to −0.22; Fig. 3, dotted line), indicating that Val carriers drove the effect observed in the sample as a whole. Note that average E+ levels did not differ significantly by COMT genotype in either the MP (F(2,31) = 0.91; p = 0.412) or FP visit (F(2,31) = 0.05; p = 0.952). However, there was a trend toward COMT genotype-dependent effects on the difference in measured E+ levels between the MP and FP visits (F(2,31) = 3.05; p = 0.062), reflecting the fact that we observed the greatest rise in E+ among COMTMet/Met individuals (0.78 ± 0.91, standardized values), with substantially smaller average E+ rises in COMTVal/Met (0.31 ± 0.41) and COMTVal/Val (0.18 ± 0.47) individuals. The difference in E+ rise between the COMTMet/Met and COMTVal/Val groups approached statistical significance (t(12.28) = −1.97; p = 0.072). Thus, the apparently greater sensitivity of Now bias to changes in E+ levels among COMTVal carriers cannot be attributed to differences in baseline E+ levels or to a greater degree of E+ change among the COMTVal carriers, who together had smaller increases in E+ from the MP to the FP than COMTMet/Met individuals (0.22 ± 0.45; t(10.86) = 1.87; p = 0.088).
Measures of frontal DA neurotransmission decrease from adolescence to adulthood (Wahlstrom et al., 2010), including a rise in COMT expression with age (Tunbridge et al., 2007). Moreover, in our previous study (Smith and Boettiger, 2012) with a mixed sample of males and females, we found that age interacts with COMT genotype to predict Now bias. That study defined the late adolescent and adult groups as ages 18–21 and 22–40 years, respectively, based both on preliminary results from other studies in our laboratory showing Now bias differences between these two age groups, and a large-scale investigation of functional brain maturation, indicating that brain maturation asymptotes at approximately age 22 years (Dosenbach et al., 2010). Thus, we also conducted an exploratory examination of the correlation between E+ change and ICR change within COMTVal carriers separately within our previously defined age groups (ages 18–21 vs 22–40 years). We observed a substantially larger correlation among 22- to 40-year-old COMTVal carriers (ρ(9) = −0.77; p = 0.005; 95% CI, −0.95 to −0.26), who have the lowest predicted tonic PFC DA levels, relative to 18- to 21-year-old Val carriers (ρ(11) = −0.604; p = 0.029; 95% CI, −0.93 to −0.09), consistent with our inverted-U model (Smith and Boettiger, 2012). Within the group with the highest predicted tonic PFC DA level (18- to 21-year-old Met/Met individuals), increasing E+ levels were associated with increased ICR at mid-cycle, although the effect was not statistically significant (ρ(2) = 0.400; p = 0.60). The relationship between E+ change and ICR change was relatively flat in the 22- to 40-year-old Met/Met group (ρ(4) = −0.086; p = 0.872). While these observations are consistent with our hypothesized results, these are highly preliminary findings, and it is particularly important to note that the tests within the Met/Met age groups are grossly underpowered to detect even large effects.
Discussion
Here, we demonstrate that Now bias decreases significantly from the MP to the FP. Participants with a verified rise in E+ levels from the MP to the FP showed significantly greater decreases in Now bias at mid-cycle than those without, with Now bias decreasing significantly in the E+ rise group but not in the no-E+ rise group. The change in Now bias from the MP to the FP inversely correlated with the change in E+, an effect driven by individuals with putatively lower frontal DA levels. These data suggest that the E+ rise during the peak fertility window modulates Now bias through interactions with tonic frontal DA.
Neurocognitive effects of estrogen
Existing evidence links E+ and frontal-dependent function in animals, presumably via E+-induced enhancement of DA signaling (Xiao and Becker, 1994; Pasqualini et al., 1995; Shansky et al., 2004). E+ increases frontal dopaminergic innervation in primates (Kritzer and Kohama, 1999), increases frontal pyramidal cell spine number (Tang et al., 2004), and affects frontal-dependent task performance (Tinkler and Voytko, 2005; Hao et al., 2007). Direct evidence for the relationship between E+ and frontal DA levels in humans is lacking. PET studies suggest sex-specific differences in DA D2 receptor levels (Kaasinen et al., 2001); however, no published studies comparing the sexes have used radiotracers suited to detect frontal DA levels. Furthermore, there are no published human PET data regarding the effect of cyclic E+ changes on DA signaling. However, behavioral and fMRI data from humans indicate that E+ modulates frontal-dependent cognitive processes (Keenan et al., 2001; Rosenberg and Park, 2002; Colzato et al., 2010; Jacobs and D'Esposito, 2011; Joseph et al., 2012). Moreover, E+ effects on working memory interact with COMT genotype (Jacobs and D'Esposito, 2011). Notably, time perception depends upon the frontoparietal circuits supporting working memory (Lewis and Miall, 2006; Piras et al., 2014), and time perception has been shown to correlate with delay discounting (Takahashi, 2004; Zauberman et al., 2009; Han and Takahashi, 2012). Moreover, time perception has long been known to be sensitive to DA (Meck, 1996), with DA increases speeding up subjective time. More recently, acute E+ administration was also demonstrated to speed subjective time perception in rats (Pleil et al., 2011). Thus, reduced Now bias at mid-cycle could reflect accelerated subjective time perception consequent to frontal circuit modulation by rising E+ levels. However, elevated Now bias in the MP due to heightened responses to the more affectively salient proximate rewards during low E+ cannot be ruled out (Goldstein et al., 2005). Furthermore, E+ actions in the hippocampus (Ooishi et al., 2012; Kato et al., 2013), a structure linked to discounting behavior in both rodents (Abela and Chudasama, 2014) and humans (Peters and Büchel, 2010; Benoit et al., 2011), could also contribute to E+ modulation of Now bias.
Dopamine as a potential mediator of the cycle effect on decision making
The present data support the idea that E+ modulates Now bias via the DA system, especially frontal DA. This conclusion derives from the data described in the previous section coupled with our finding that the correlation between changes in E+ and Now bias from the MP to the FP varies with COMT Val158Met genotype. The Val158Met polymorphism substitutes the ancestral Val for a Met at position 158 in the COMT enzyme, resulting in a fourfold reduction in enzyme activity (Chen et al., 2004). Reduced tonic frontal DA is observed in people with the COMT 158Val/Val genotype relative to COMTMet allele carriers (Wu et al., 2012). According to a model wherein Now bias varies with frontal DA according to an inverted-U function (Mitchell et al., 2007; Altamirano et al., 2011; Smith and Boettiger, 2012; Kelm and Boettiger, 2013), these data suggest that E+ elevations alter Now bias via interaction with frontal DA levels. This idea remains to be empirically tested, but frontal-dependent tasks are generally sensitive to frontal DA according to a quadratic function (Sawaguchi and Goldman-Rakic, 1991; Vijayraghavan et al., 2007; Cools and D'Esposito, 2011).
Role of other hormones in delay discounting
Other steroid hormones are implicated in the modulation of Now bias. For example, salivary testosterone correlates with DD among healthy male students (Takahashi et al., 2006). Delay discounting has also been shown to negatively correlate with salivary cortisol levels (Takahashi, 2004) and with a genetic polymorphism associated with increased plasma cortisol levels (Kawamura et al., 2013). However, sex moderates the effect of salivary cortisol: the relationship is negative among males, but positive among females (Takahashi et al., 2010). It is worth noting that stress (associated with elevated cortisol levels) impacts frontal function in an inverted-U manner (Arnsten, 2009), with E+ exacerbating stress-induced working memory impairment in animal models (Shansky et al., 2004, 2009). In humans, females are more susceptible to stress-induced working memory impairment than are men (Schoofs et al., 2013). To date, no studies have investigated the effects of progesterone (PROG) on DD. Moreover, no study has yet examined multiple steroids, and this study is the first to control for cycle phase in females. Future work investigating the individual contributions of different steroid hormones is needed for a complete picture of the hormonal modulation of Now bias. Perhaps most importantly, to date, no hormone challenge studies have been conducted, which are required to determine causal roles in modulating Now bias.
Other potential mediators of cycle effects on Now bias
Although our finding of COMT-dependent E+ effects on Now bias suggests a mediating role for DA in the observed cycle effect on Now bias, other neurochemical systems could be involved. In particular, PROG or its neuroactive steroid (NAS) derivatives, such as allopregnanolone (ALLO), which also rise across the cycle, may contribute. Indeed, ALLO modulates cognitive function in humans (Marx et al., 2009), possibly via positive modulation of GABAA receptors (Majewska et al., 1986). Neuroimaging studies have demonstrated that increases in PROG levels and decreases in E+ levels from the FP to the LP result in changes in activity in a host of PFC, limbic, and striatal regions (Dreher et al., 2007; van Wingen et al., 2008; Ossewaarde et al., 2011; Marecková et al., 2014). Like PROG and ALLO, GABA levels also rise from the FP to LP (Epperson et al., 2002). Thus, PROG, ALLO, or GABA may alter the function of neural circuitry implicated in Now bias, and this requires further study. Imprecision in our sampling method means that we cannot rule out a role for cyclic changes in PROG, ALLO, or GABA in reducing Now bias at mid-cycle. Increased GABAergic signaling could theoretically improve cognitive control, leading to reduced Now bias. Indeed, PROG administration can improve cognitive control and reduce smoking urges (Sofuoglu et al., 2011). However, in women, ALLO impairs episodic memory (Kask et al., 2008), which shares common neural substrates with episodic prospection, a mental action that decreases Now bias via increased frontotemporal connectivity (Peters and Büchel, 2010; Benoit et al., 2011), which is weakened by PROG increases (van Wingen et al., 2008). However, enhancing GABAA signaling reportedly has no effect on Now bias (Reynolds et al., 2004; Acheson et al., 2006). Thus, although the reduced Now bias we observed at mid-cycle could theoretically reflect some contribution of PROG, a NAS, or GABA, no direct data are available, and the circumstantial evidence does not favor that interpretation.
Study limitations
Some study limitations prevent drawing strong conclusions regarding the role of specific hormones in the observed cycle effect on Now bias. First, we assayed only E+ in our subjects, so roles for other hormones cannot be ruled out. Second, we were not able to collect E+ samples from all participants, although this concern is somewhat mitigated by the fact that the subset providing E+ samples did not differ from the subset that did not. Considering the two subsets as independent samples, this fact could be seen instead as a replication of our basic behavioral finding across subsamples, strengthening that finding. Third, our method relied on self-report of cycle day, and did not document cycle duration or hormonal markers of ovulation across the cycle, which could have substantially reduced variance, particularly in the FP. We likely failed to catch the E+ peak in many participants, which could substantially diminish effect size. Indeed, among the participants for whom we quantified E+, many showed little or no rise in E+ levels from the FP to the MP visit, and the effect size was substantially greater in the participants for whom we verified a rise in E+ levels in the FP. More precise monitoring of hormone levels within individuals and targeting behavioral measurements to individually determined cycle phases are advised for future research. Moreover, while the results suggest a role for E+ in modulating Now bias, direct manipulation of E+ is required to establish a causal role.
Finally, while our finding that E+ interacts with a marker of frontal DA tone is consistent with some existing literature, our interpretation of those findings remains speculative. In particular, no direct evidence demonstrates that E+ modulates frontal DA in humans, nor that frontal DA modulates Now bias according to a U-shaped function. Future PET studies of DA signaling are needed to test our model. Regardless, our model suggests that the effects of dopaminergic medications may be optimized by taking E+ levels and COMT genotype into account. Given the widespread use of such medications, such personalization could have a broad therapeutic impact.
Footnotes
This work was supported by National Institutes of Health Grants UL1RR025747, KL2RR025746, and P60AA011605; by a University of North Carolina Lindquist Faculty Award (C.A.B.); and by National Institutes of Health Grants T32NS007431 and F31AA020132 (C.T.S.). We thank M. Parrish, M. Le, J. Drost-Lopez, C. Lang, and E. Steel for technical assistance.
The authors declare no competing financial interests.
References
- Abela AR, Chudasama Y. Noradrenergic alpha2A-receptor stimulation in the ventral hippocampus reduces impulsive decision-making. Psychopharmacology (Berl) 2014;231:521–531. doi: 10.1007/s00213-013-3262-y. [DOI] [PubMed] [Google Scholar]
- Acheson A, de Wit H. Bupropion improves attention but does not affect impulsive behavior in healthy young adults. Exp Clin Psychopharmacol. 2008;16:113–123. doi: 10.1037/1064-1297.16.2.113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Acheson A, Reynolds B, Richards JB, de Wit H. Diazepam impairs behavioral inhibition but not delay discounting or risk taking in healthy adults. Exp Clin Psychopharmacol. 2006;14:190–198. doi: 10.1037/1064-1297.14.2.190. [DOI] [PubMed] [Google Scholar]
- Alessi SM, Petry NM. Pathological gambling severity is associated with impulsivity in a delay discounting procedure. Behav Processes. 2003;64:345–354. doi: 10.1016/S0376-6357(03)00150-5. [DOI] [PubMed] [Google Scholar]
- Altamirano LJ, Fields HL, D'Esposito M, Boettiger CA. Interaction between family history of alcoholism and Locus of Control in the opioid regulation of impulsive responding under the influence of alcohol. Alcohol Clin Exp Res. 2011;35:1905–1914. doi: 10.1111/j.1530-0277.2011.01535.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anokhin AP, Golosheykin S, Grant JD, Heath AC. Heritability of delay discounting in adolescence: a longitudinal twin study. Behav Genet. 2011;41:175–183. doi: 10.1007/s10519-010-9384-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arnsten AF. Stress signalling pathways that impair prefrontal cortex structure and function. Nat Rev Neurosci. 2009;10:410–422. doi: 10.1038/nrn2648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barkley RA, Edwards G, Laneri M, Fletcher K, Metevia L. Executive functioning, temporal discounting, and sense of time in adolescents with attention deficit hyperactivity disorder (ADHD) and oppositional defiant disorder (ODD) J Abnorm Child Psychol. 2001;29:541–556. doi: 10.1023/A:1012233310098. [DOI] [PubMed] [Google Scholar]
- Becker GS, Murphy KM. A theory of rational addiction. J Polit Econ. 1988;96:675–700. doi: 10.1086/261558. [DOI] [Google Scholar]
- Benoit RG, Gilbert SJ, Burgess PW. A neural mechanism mediating the impact of episodic prospection on farsighted decisions. J Neurosci. 2011;31:6771–6779. doi: 10.1523/JNEUROSCI.6559-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bobova L, Finn PR, Rickert ME, Lucas J. Disinhibitory psychopathology and delay discounting in alcohol dependence: personality and cognitive correlates. Exp Clin Psychopharmacol. 2009;17:51–61. doi: 10.1037/a0014503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boettiger CA, Mitchell JM, Tavares VC, Robertson M, Joslyn G, D'Esposito M, Fields HL. Immediate reward bias in humans: fronto-parietal networks and a role for the catechol-O-methyltransferase 158(Val/Val) genotype. J Neurosci. 2007;27:14383–14391. doi: 10.1523/JNEUROSCI.2551-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen J, Lipska BK, Halim N, Ma QD, Matsumoto M, Melhem S, Kolachana BS, Hyde TM, Herman MM, Apud J, Egan MF, Kleinman JE, Weinberger DR. Functional analysis of genetic variation in catechol-O-methyltransferase (COMT): effects on mRNA, protein, and enzyme activity in postmortem human brain. Am J Hum Genet. 2004;75:807–821. doi: 10.1086/425589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Colzato LS, Hertsig G, van den Wildenberg WP, Hommel B. Estrogen modulates inhibitory control in healthy human females: evidence from the stop-signal paradigm. Neuroscience. 2010;167:709–715. doi: 10.1016/j.neuroscience.2010.02.029. [DOI] [PubMed] [Google Scholar]
- Cools R, D'Esposito M. Inverted-U-shaped dopamine actions on human working memory and cognitive control. Biol Psychiatry. 2011;69:e113–125. doi: 10.1016/j.biopsych.2011.03.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dalley JW, Mar AC, Economidou D, Robbins TW. Neurobehavioral mechanisms of impulsivity: fronto-striatal systems and functional neurochemistry. Pharmacol Biochem Behav. 2008;90:250–260. doi: 10.1016/j.pbb.2007.12.021. [DOI] [PubMed] [Google Scholar]
- de Wit H. Impulsivity as a determinant and consequence of drug use: a review of underlying processes. Addict Biol. 2009;14:22–31. doi: 10.1111/j.1369-1600.2008.00129.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Wit H, Enggasser JL, Richards JB. Acute administration of d-amphetamine decreases impulsivity in healthy volunteers. Neuropsychopharmacology. 2002;27:813–825. doi: 10.1016/S0893-133X(02)00343-3. [DOI] [PubMed] [Google Scholar]
- de Wit H, Flory JD, Acheson A, McCloskey M, Manuck SB. IQ and nonplanning impulsivity are independently associated with delay discounting in middle-aged adults. Pers Individ Dif. 2007;42:111–121. doi: 10.1016/j.paid.2006.06.026. [DOI] [Google Scholar]
- Dosenbach NU, Nardos B, Cohen AL, Fair DA, Power JD, Church JA, Nelson SM, Wig GS, Vogel AC, Lessov-Schlaggar CN, Barnes KA, Dubis JW, Feczko E, Coalson RS, Pruett JR, Jr, Barch DM, Petersen SE, Schlaggar BL. Prediction of individual brain maturity using fMRI. Science. 2010;329:1358–1361. doi: 10.1126/science.1194144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doya K. Modulators of decision making. Nat Neurosci. 2008;11:410–416. doi: 10.1038/nn2077. [DOI] [PubMed] [Google Scholar]
- Dreher JC, Schmidt PJ, Kohn P, Furman D, Rubinow D, Berman KF. Menstrual cycle phase modulates reward-related neural function in women. Proc Natl Acad Sci U S A. 2007;104:2465–2470. doi: 10.1073/pnas.0605569104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eisenberg DT, Mackillop J, Modi M, Beauchemin J, Dang D, Lisman SA, Lum JK, Wilson DS. Examining impulsivity as an endophenotype using a behavioral approach: a DRD2 TaqI A and DRD4 48-bp VNTR association study. Behav Brain Funct. 2007;3:2. doi: 10.1186/1744-9081-3-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Epperson CN, Haga K, Mason GF, Sellers E, Gueorguieva R, Zhang W, Weiss E, Rothman DL, Krystal JH. Cortical gamma-aminobutyric acid levels across the menstrual cycle in healthy women and those with premenstrual dysphoric disorder: a proton magnetic resonance spectroscopy study. Arch Gen Psychiatry. 2002;59:851–858. doi: 10.1001/archpsyc.59.9.851. [DOI] [PubMed] [Google Scholar]
- Fernández-de-Las-Peñas C, Cantarero-Villanueva I, Fernández-Lao C, Ambite-Quesada S, Díaz-Rodríguez L, Rivas-Martínez I, del Moral-Avila R, Arroyo-Morales M. Influence of catechol-o-methyltransferase genotype (Val158Met) on endocrine, sympathetic nervous and mucosal immune systems in breast cancer survivors. Breast. 2012;21:199–203. doi: 10.1016/j.breast.2011.09.012. [DOI] [PubMed] [Google Scholar]
- Gogos JA, Morgan M, Luine V, Santha M, Ogawa S, Pfaff D, Karayiorgou M. Catechol-O-methyltransferase-deficient mice exhibit sexually dimorphic changes in catecholamine levels and behavior. Proc Natl Acad Sci U S A. 1998;95:9991–9996. doi: 10.1073/pnas.95.17.9991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldstein JM, Jerram M, Poldrack R, Ahern T, Kennedy DN, Seidman LJ, Makris N. Hormonal cycle modulates arousal circuitry in women using functional magnetic resonance imaging. J Neurosci. 2005;25:9309–9316. doi: 10.1523/JNEUROSCI.2239-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hamidovic A, Kang UJ, de Wit H. Effects of low to moderate acute doses of pramipexole on impulsivity and cognition in healthy volunteers. J Clin Psychopharmacol. 2008;28:45–51. doi: 10.1097/jcp.0b013e3181602fab. [DOI] [PubMed] [Google Scholar]
- Han R, Takahashi T. Psychophysics of time perception and valuation in temporal discounting of gain and loss. Physica A. 2012;391:6568–6576. doi: 10.1016/j.physa.2012.07.012. [DOI] [Google Scholar]
- Hao J, Rapp PR, Janssen WG, Lou W, Lasley BL, Hof PR, Morrison JH. Interactive effects of age and estrogen on cognition and pyramidal neurons in monkey prefrontal cortex. Proc Natl Acad Sci U S A. 2007;104:11465–11470. doi: 10.1073/pnas.0704757104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jacobs E, D'Esposito M. Estrogen shapes dopamine-dependent cognitive processes: implications for women's health. J Neurosci. 2011;31:5286–5293. doi: 10.1523/JNEUROSCI.6394-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Joseph JE, Swearingen JE, Corbly CR, Curry TE, Jr, Kelly TH. Influence of estradiol on functional brain organization for working memory. Neuroimage. 2012;59:2923–2931. doi: 10.1016/j.neuroimage.2011.09.067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaasinen V, Någren K, Hietala J, Farde L, Rinne JO. Sex differences in extrastriatal dopamine d(2)-like receptors in the human brain. Am J Psychiatry. 2001;158:308–311. doi: 10.1176/appi.ajp.158.2.308. [DOI] [PubMed] [Google Scholar]
- Käenmäki M, Tammimäki A, Myöhänen T, Pakarinen K, Amberg C, Karayiorgou M, Gogos JA, Männistö PT. Quantitative role of COMT in dopamine clearance in the prefrontal cortex of freely moving mice. J Neurochem. 2010;114:1745–1755. doi: 10.1111/j.1471-4159.2010.06889.x. [DOI] [PubMed] [Google Scholar]
- Karoum F, Chrapusta SJ, Egan MF. 3-Methoxytyramine is the major metabolite of released dopamine in the rat frontal cortex: reassessment of the effects of antipsychotics on the dynamics of dopamine release and metabolism in the frontal cortex, nucleus accumbens, and striatum by a simple two pool model. J Neurochem. 1994;63:972–979. doi: 10.1046/j.1471-4159.1994.63030972.x. [DOI] [PubMed] [Google Scholar]
- Kask K, Backstrom T, Nilsson LG, Sundstrom-Poromaa I. Allopregnanolone impairs episodic memory in healthy women. Psychopharmacology (Berl) 2008;199:161–168. doi: 10.1007/s00213-008-1150-7. [DOI] [PubMed] [Google Scholar]
- Kato A, Hojo Y, Higo S, Komatsuzaki Y, Murakami G, Yoshino H, Uebayashi M, Kawato S. Female hippocampal estrogens have a significant correlation with cyclic fluctuation of hippocampal spines. Front Neural Circuits. 2013;7:149. doi: 10.3389/fncir.2013.00149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kawamura Y, Takahashi T, Liu X, Nishida N, Tokunaga K, Ukawa K, Noda Y, Yoshikawa A, Shimada T, Umekage T, Sasak T. DNA polymorphism in the FKBP5 gene affects impulsivity in intertemporal choice. Asia Pac Psychiatry. 2013;5:31–38. doi: 10.1111/appy.12009. [DOI] [PubMed] [Google Scholar]
- Keenan PA, Ezzat WH, Ginsburg K, Moore GJ. Prefrontal cortex as the site of estrogen's effect on cognition. Psychoneuroendocrinology. 2001;26:577–590. doi: 10.1016/S0306-4530(01)00013-0. [DOI] [PubMed] [Google Scholar]
- Kelm MK, Boettiger CA. Effects of acute dopamine precusor depletion on immediate reward selection bias and working memory depend on catechol-o-methyltransferase genotype. J Cogn Neurosci. 2013;25:2061–2071. doi: 10.1162/jocn_a_00464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kritzer MF, Kohama SG. Ovarian hormones differentially influence immunoreactivity for dopamine beta- hydroxylase, choline acetyltransferase, and serotonin in the dorsolateral prefrontal cortex of adult rhesus monkeys. J Comp Neurol. 1999;409:438–451. doi: 10.1002/(SICI)1096-9861(19990705)409:3<438::AID-CNE8>3.0.CO%3B2-5. [DOI] [PubMed] [Google Scholar]
- Lang UE, Bajbouj M, Bajbouj M, Sander T, Gallinat J. Gender-dependent association of the functional catechol-O-methyltransferase Val158Met genotype with sensation seeking personality trait. Neuropsychopharmacology. 2007;32:1950–1955. doi: 10.1038/sj.npp.1301335. [DOI] [PubMed] [Google Scholar]
- Leeman RF, Potenza MN. Similarities and differences between pathological gambling and substance use disorders: a focus on impulsivity and compulsivity. Psychopharmacology (Berl) 2012;219:469–490. doi: 10.1007/s00213-011-2550-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lewis PA, Miall RC. Remembering the time: a continuous clock. Trends Cogn Sci. 2006;10:401–406. doi: 10.1016/j.tics.2006.07.006. [DOI] [PubMed] [Google Scholar]
- Lu Y, Bentley GR, Gann PH, Hodges KR, Chatterton RT. Salivary estradiol and progesterone levels in conception and nonconception cycles in women: evaluation of a new assay for salivary estradiol. Fertil Steril. 1999;71:863–868. doi: 10.1016/S0015-0282(99)00093-X. [DOI] [PubMed] [Google Scholar]
- Majewska MD, Harrison NL, Schwartz RD, Barker JL, Paul SM. Steroid hormone metabolites are barbiturate-like modulators of the GABA receptor. Science. 1986;232:1004–1007. doi: 10.1126/science.2422758. [DOI] [PubMed] [Google Scholar]
- Marecková K, Perrin JS, Nawaz Khan I, Lawrence C, Dickie E, McQuiggan DA, Paus T. Hormonal contraceptives, menstrual cycle and brain response to faces. Soc Cogn Affect Neurosci. 2014;9:191–200. doi: 10.1093/scan/nss128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marx CE, Keefe RS, Buchanan RW, Hamer RM, Kilts JD, Bradford DW, Strauss JL, Naylor JC, Payne VM, Lieberman JA, Savitz AJ, Leimone LA, Dunn L, Porcu P, Morrow AL, Shampine LJ. Proof-of-concept trial with the neurosteroid pregnenolone targeting cognitive and negative symptoms in schizophrenia. Neuropsychopharmacology. 2009;34:1885–1903. doi: 10.1038/npp.2009.26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mazur JE. Quantitative analyses of behavior. Hillsdale, NJ: Erlbaum; 1987. An adjusting procedure for studying delayed reinforcement. [Google Scholar]
- Meck WH. Neuropharmacology of timing and time perception. Brain Res Cogn Brain Res. 1996;3:227–242. doi: 10.1016/0926-6410(96)00009-2. [DOI] [PubMed] [Google Scholar]
- Mitchell JM, Fields HL, D'Esposito M, Boettiger CA. Impulsive responding in alcoholics. Alcohol Clin Exp Res. 2005;29:2158–2169. doi: 10.1097/01.alc.0000191755.63639.4a. [DOI] [PubMed] [Google Scholar]
- Mitchell JM, Tavares VC, Fields HL, D'Esposito M, Boettiger CA. Endogenous opioid blockade and impulsive responding in alcoholics and healthy controls. Neuropsychopharmacology. 2007;32:439–449. doi: 10.1038/sj.npp.1301226. [DOI] [PubMed] [Google Scholar]
- Mitchell SH. The genetic basis of delay discounting and its genetic relationship to alcohol dependence. Behav Processes. 2011;87:10–17. doi: 10.1016/j.beproc.2011.02.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ooishi Y, Kawato S, Hojo Y, Hatanaka Y, Higo S, Murakami G, Komatsuzaki Y, Ogiue-Ikeda M, Kimoto T, Mukai H. Modulation of synaptic plasticity in the hippocampus by hippocampus-derived estrogen and androgen. J Steroid Biochem Mol Biol. 2012;131:37–51. doi: 10.1016/j.jsbmb.2011.10.004. [DOI] [PubMed] [Google Scholar]
- Ossewaarde L, van Wingen GA, Kooijman SC, Bäckström T, Fernández G, Hermans EJ. Changes in functioning of mesolimbic incentive processing circuits during the premenstrual phase. Soc Cogn Affect Neurosci. 2011;6:612–620. doi: 10.1093/scan/nsq071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paloyelis Y, Asherson P, Mehta MA, Faraone SV, Kuntsi J. DAT1 and COMT effects on delay discounting and trait impulsivity in male adolescents with attention deficit/hyperactivity disorder and healthy controls. Neuropsychopharmacology. 2010;35:2414–2426. doi: 10.1038/npp.2010.124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pasqualini C, Olivier V, Guibert B, Frain O, Leviel V. Acute stimulatory effect of estradiol on striatal dopamine synthesis. J Neurochem. 1995;65:1651–1657. doi: 10.1046/j.1471-4159.1995.65041651.x. [DOI] [PubMed] [Google Scholar]
- Patton JH, Stanford MS, Barratt ES. Factor structure of the Barratt impulsiveness scale. J Clin Psychol. 1995;51:768–774. doi: 10.1002/1097-4679(199511)51:6<768::AID-JCLP2270510607>3.0.CO%3B2-1. [DOI] [PubMed] [Google Scholar]
- Peper JS, Mandl RC, Braams BR, de Water E, Heijboer AC, Koolschijn PC, Crone EA. Delay discounting and frontostriatal fiber tracts: a combined DTI and MTR study on impulsive choices in healthy young adults. Cereb Cortex. 2013;23:1695–1702. doi: 10.1093/cercor/bhs163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perry JL, Carroll ME. The role of impulsive behavior in drug abuse. Psychopharmacology (Berl) 2008;200:1–26. doi: 10.1007/s00213-008-1173-0. [DOI] [PubMed] [Google Scholar]
- Peters J, Büchel C. Episodic future thinking reduces reward delay discounting through an enhancement of prefrontal-mediotemporal interactions. Neuron. 2010;66:138–148. doi: 10.1016/j.neuron.2010.03.026. [DOI] [PubMed] [Google Scholar]
- Pine A, Shiner T, Seymour B, Dolan RJ. Dopamine, time, and impulsivity in humans. J Neurosci. 2010;30:8888–8896. doi: 10.1523/JNEUROSCI.6028-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piras F, Piras F, Ciullo V, Danese E, Caltagirone C, Spalletta G. Time dysperception perspective for acquired brain injury. Front Neurol. 2014;4:217. doi: 10.3389/fneur.2013.00217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pleil KE, Cordes S, Meck WH, Williams CL. Rapid and acute effects of estrogen on time perception in male and female rats. Front Integr Neurosci. 2011;5:63. doi: 10.3389/fnint.2011.00063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prelec D. Decreasing impatience: a criterion for non-stationary time preference and “hyperbolic” discounting. Scand J Econ. 2004;106:511–532. doi: 10.1111/j.0347-0520.2004.00375.x. [DOI] [Google Scholar]
- Qian Q, Wang Y, Zhou R, Li J, Wang B, Glatt S, Faraone SV. Family-based and case-control association studies of catechol-O-methyltransferase in attention deficit hyperactivity disorder suggest genetic sexual dimorphism. Am J Med Genet B Neuropsychiatr Genet. 2003;118B:103–109. doi: 10.1002/ajmg.b.10064. [DOI] [PubMed] [Google Scholar]
- Rachlin H. The science of self-control. Cambridge, MA: Harvard UP; 2000. [Google Scholar]
- Reynolds B. A review of delay-discounting research with humans: relations to drug use and gambling. Behav Pharmacol. 2006;17:651–667. doi: 10.1097/FBP.0b013e3280115f99. [DOI] [PubMed] [Google Scholar]
- Reynolds B, Richards JB, Dassinger M, de Wit H. Therapeutic doses of diazepam do not alter impulsive behavior in humans. Pharmacol Biochem Behav. 2004;79:17–24. doi: 10.1016/j.pbb.2004.06.011. [DOI] [PubMed] [Google Scholar]
- Rogers RD, Moeller FG, Swann AC, Clark L. Recent research on impulsivity in individuals with drug use and mental health disorders: implications for alcoholism. Alcohol Clin Exp Res. 2010;34:1319–1333. doi: 10.1111/j.1530-0277.2010.01216.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosenberg L, Park S. Verbal and spatial functions across the menstrual cycle in healthy young women. Psychoneuroendocrinology. 2002;27:835–841. doi: 10.1016/S0306-4530(01)00083-X. [DOI] [PubMed] [Google Scholar]
- Rotter JB. Generalized expectancies for internal versus external control of reinforcement. Psychol Monogr. 1966;80:1–28. [PubMed] [Google Scholar]
- Rubinow DR, Schmidt PJ. Gonadal steroid regulation of mood: the lessons of premenstrual syndrome. Front Neuroendocrinol. 2006;27:210–216. doi: 10.1016/j.yfrne.2006.02.003. [DOI] [PubMed] [Google Scholar]
- Sawaguchi T, Goldman-Rakic PS. D1 dopamine receptors in prefrontal cortex: involvement in working memory. Science. 1991;251:947–950. doi: 10.1126/science.1825731. [DOI] [PubMed] [Google Scholar]
- Schoofs D, Pabst S, Brand M, Wolf OT. Working memory is differentially affected by stress in men and women. Behav Brain Res. 2013;241:144–153. doi: 10.1016/j.bbr.2012.12.004. [DOI] [PubMed] [Google Scholar]
- Shansky RM, Glavis-Bloom C, Lerman D, McRae P, Benson C, Miller K, Cosand L, Horvath TL, Arnsten AF. Estrogen mediates sex differences in stress-induced prefrontal cortex dysfunction. Mol Psychiatry. 2004;9:531–538. doi: 10.1038/sj.mp.4001435. [DOI] [PubMed] [Google Scholar]
- Shansky RM, Bender G, Arnsten AF. Estrogen prevents norepinephrine alpha-2a receptor reversal of stress-induced working memory impairment. Stress. 2009;12:457–463. doi: 10.1080/10253890802520988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Slifstein M, Kolachana B, Simpson EH, Tabares P, Cheng B, Duvall M, Frankle WG, Weinberger DR, Laruelle M, Abi-Dargham A. COMT genotype predicts cortical-limbic D1 receptor availability measured with [11C]NNC112 and PET. Mol Psychiatry. 2008;13:821–827. doi: 10.1038/mp.2008.19. [DOI] [PubMed] [Google Scholar]
- Small BJ, Rawson KS, Walsh E, Jim HS, Hughes TF, Iser L, Andrykowski MA, Jacobsen PB. Catechol-O-methyltransferase genotype modulates cancer treatment-related cognitive deficits in breast cancer survivors. Cancer. 2011;117:1369–1376. doi: 10.1002/cncr.25685. [DOI] [PubMed] [Google Scholar]
- Smith CT, Boettiger CA. Age modulates the effect of COMT genotype on delay discounting behavior. Psychopharmacology (Berl) 2012;222:609–617. doi: 10.1007/s00213-012-2653-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sofuoglu M, Mouratidis M, Mooney M. Progesterone improves cognitive performance and attenuates smoking urges in abstinent smokers. Psychoneuroendocrinology. 2011;36:123–132. doi: 10.1016/j.psyneuen.2010.07.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sonuga-Barke EJ, Sergeant JA, Nigg J, Willcutt E. Executive dysfunction and delay aversion in attention deficit hyperactivity disorder: nosologic and diagnostic implications. Child Adolesc Psychiatr Clin N Am. 2008;17:367–384. ix. doi: 10.1016/j.chc.2007.11.008. [DOI] [PubMed] [Google Scholar]
- Takahashi T. Cortisol levels and time-discounting of monetary gain in humans. Neuroreport. 2004;15:2145–2147. doi: 10.1097/00001756-200409150-00029. [DOI] [PubMed] [Google Scholar]
- Takahashi T. Theoretical frameworks for neuroeconomics of intertemporal choice. J Neurosci Psychol Econ. 2009;2:75. [Google Scholar]
- Takahashi T. A neuroeconomic theory of rational addiction and nonlinear time-perception. Neuro Endocrinol Lett. 2011;32:221–225. [PubMed] [Google Scholar]
- Takahashi T, Sakaguchi K, Oki M, Homma S, Hasegawa T. Testosterone levels and discounting delayed monetary gains and losses in male humans. Neuro Endocrinol Lett. 2006;27:439–444. [PubMed] [Google Scholar]
- Takahashi T, Oono H, Radford MH. Psychophysics of time perception and intertemporal choice models. Physica A. 2008;387:2066–2074. doi: 10.1016/j.physa.2007.11.047. [DOI] [Google Scholar]
- Takahashi T, Shinada M, Inukai K, Tanida S, Takahashi C, Mifune N, Takagishi H, Horita Y, Hashimoto H, Yokota K, Kameda T, Yamagishi T. Stress hormones predict hyperbolic time-discount rates six months later in adults. Neuro Endocrinol Lett. 2010;31:616–621. [PubMed] [Google Scholar]
- Tang Y, Janssen WG, Hao J, Roberts JA, McKay H, Lasley B, Allen PB, Greengard P, Rapp PR, Kordower JH, Hof PR, Morrison JH. Estrogen replacement increases spinophilin-immunoreactive spine number in the prefrontal cortex of female rhesus monkeys. Cereb Cortex. 2004;14:215–223. doi: 10.1093/cercor/bhg121. [DOI] [PubMed] [Google Scholar]
- Tinkler GP, Voytko ML. Estrogen modulates cognitive and cholinergic processes in surgically menopausal monkeys. Prog Neuropsychopharmacol Biol Psychiatry. 2005;29:423–431. doi: 10.1016/j.pnpbp.2004.12.016. [DOI] [PubMed] [Google Scholar]
- Tunbridge EM, Weickert CS, Kleinman JE, Herman MM, Chen J, Kolachana BS, Harrison PJ, Weinberger DR. Catechol-o-methyltransferase enzyme activity and protein expression in human prefrontal cortex across the postnatal lifespan. Cereb Cortex. 2007;17:1206–1212. doi: 10.1093/cercor/bhl032. [DOI] [PubMed] [Google Scholar]
- van Wingen GA, van Broekhoven F, Verkes RJ, Petersson KM, Bäckström T, Buitelaar JK, Fernández G. Progesterone selectively increases amygdala reactivity in women. Mol Psychiatry. 2008;13:325–333. doi: 10.1038/sj.mp.4002030. [DOI] [PubMed] [Google Scholar]
- Vijayraghavan S, Wang M, Birnbaum SG, Williams GV, Arnsten AF. Inverted-U dopamine D1 receptor actions on prefrontal neurons engaged in working memory. Nat Neurosci. 2007;10:376–384. doi: 10.1038/nn1846. [DOI] [PubMed] [Google Scholar]
- Wahlstrom D, Collins P, White T, Luciana M. Developmental changes in dopamine neurotransmission in adolescence: behavioral implications and issues in assessment. Brain Cogn. 2010;72:146–159. doi: 10.1016/j.bandc.2009.10.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wallace M. Future time perspective in schizophrenia. J Abnorm Psychol. 1956;52:240–245. doi: 10.1037/h0039899. [DOI] [PubMed] [Google Scholar]
- Winstanley CA. The utility of rat models of impulsivity in developing pharmacotherapies for impulse control disorders. Br J Pharmacol. 2011;164:1301–1321. doi: 10.1111/j.1476-5381.2011.01323.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu K, O'Keeffe D, Politis M, O'Keeffe GC, Robbins TW, Bose SK, Brooks DJ, Piccini P, Barker RA. The catechol-O-methyltransferase Val(158)Met polymorphism modulates fronto-cortical dopamine turnover in early Parkinson's disease: a PET study. Brain. 2012;135:2449–2457. doi: 10.1093/brain/aws157. [DOI] [PubMed] [Google Scholar]
- Xiao L, Becker JB. Quantitative microdialysis determination of extracellular striatal dopamine concentration in male and female rats: effects of estrous cycle and gonadectomy. Neurosci Lett. 1994;180:155–158. doi: 10.1016/0304-3940(94)90510-X. [DOI] [PubMed] [Google Scholar]
- Zauberman G, Kim BK, Malkoc SA, Bettman JR. Discounting time and time discounting: subjective time perception and intertemporal preferences. J Marketing Res. 2009;46:543–556. doi: 10.1509/jmkr.46.4.543. [DOI] [Google Scholar]