

Abstract
Fruit set is initiated only after fertilization and is tightly regulated primarily by gibberellins (GAs) and auxins. The application of either of these hormones induces parthenocarpy, fruit set without fertilization, but the molecular mechanism underlying this induction is poorly understood. In the present study, we have shown that the parthenocarpic fruits induced by GA application at pre-bloom result from the interaction of GA with auxin signaling. The transcriptional levels of the putative negative regulators of fruit set initiation, including Vitis auxin/indole-3-acetic acid transcription factor 9 (VvIAA9), Vitis auxin response factor 7 (VvARF7), and VvARF8 were monitored during inflorescence development in seeded diploid ‘Tamnara’ grapevines with or without GA application. Without GA application, VvIAA9, VvARF7, and VvARF8 were expressed at a relatively high level before full bloom, but decreased thereafter following pollination. After GA application at 14 days before full bloom (DBF); however, the expression levels of VvIAA9 and VvARF7 declined at 5 DBF prior to pollination. The effects of GA application on auxin levels or auxin signaling were also analyzed by monitoring the expression patterns of auxin biosynthesis genes and auxin-responsive genes with or without GA application. Transcription levels of the auxin biosynthesis genes Vitis anthranilate synthase β subunit (VvASB1-like), Vitis YUCCA2 (VvYUC2), and VvYUC6 were not significantly changed by GA application. However, the expressions of Vitis Gretchen Hagen3.2 (VvGH3.2) and VvGH3.3, auxin-responsive genes, were up-regulated from 2 DBF to full bloom with GA application. Furthermore, the Vitis GA signaling gene, VvDELLA was up-regulated by GA application during 12 DBF to 7 DBF, prior to down-regulation of VvIAA9 and VvARF7. These results suggest that VvIAA9 and VvARF7 are negative regulators of fruit set initiation in grapevines, and GA signaling is integrated with auxin signaling via VvDELLA during parthenocarpic fruit development in grapevines.
Introduction
Fruit set is initiated only after two sequential events, pollination and fertilization [1], concurrent with changes in the levels of endogenous plant hormones, primarily gibberellins (GAs) and auxins [2]–[5]. Application of GA or auxin can trigger fruit set even without pollination and can induce parthenocarpic fruit development [6]–[10].
Although the molecular mechanisms by which either GA or auxin mediates fruit set initiation are not clearly established, several auxin signaling genes related to parthenocarpic fruit development have been identified. The auxin/indole-3-acetic acid (Aux/IAA) transcription factor family is a known essential repressor of auxin signaling in various developmental processes, including fruit set [11]. Among the Aux/IAA (IAA) family genes, IAA9 has been regarded as a negative regulator, preventing fruit set initiation in the absence of pollination in tomato, with the silencing line, SlIAA9, showing parthenocarpic fruit development [12]. Two auxin response factors (ARF) related to parthenocarpic fruit development have also been identified in Arabidopsis and tomato [12]–[15]. The ARF family acts as a regulator of auxin-responsive genes by specifically binding to auxin response elements (AuxREs) in the promoters of auxin-responsive genes. ARFs play important roles in diverse developmental processes in embryos, hypocotyls, floral organs, and fruit [15]–[21]. The Arabidopsis fruit without fertilization (fwf) mutant produces parthenocarpic fruit as the result of the expression of truncated ARF8 [14], [15], and the same mutation in the SlARF8 in tomato induced parthenocarpy [14]. Parthenocarpy induction and pollen tube growth inhibition were also observed in a silencing line of ARF7 in tomato [13], [22]. The expression of SlARF7 was maintained at high levels before pollination and rapidly declined after pollination with increasing auxin content [1]–[3], [6], in a manner similar to its down-regulation upon exogenous auxin application [13].
GA-induced parthenocarpic fruit development has been observed in tomato mutants (pat, pat-2, and pat-3/pat-4) showing overexpression of the GA biosynthesis genes [23]–[25], and in silencing lines of DELLA in Arabidopsis [26] and tomato [27], demonstrating that GA signaling plays a role in parthenocarpic fruit development. The regulatory roles of the DELLA protein in ARF7 expression, and the partial activation of auxin signaling during parthenocarpic fruit development have been reported in the procera (pro) mutant of DELLA in tomato [28]. Additionally, auxin application and the silencing line of ARF7 induced parthenocarpy by regulating expression levels of the GA metabolic genes [22], [29], [30]. These data suggest that both GA and auxin influence fruit set initiation, and crosstalk between GA and auxin signaling plays a role in parthenocarpic fruit development.
In grapevines, GA has commonly been used to induce parthenocarpy [2], [7], [8], [31]. The effects of GA application have been studied on early ripening and berry enlargement [32]–[34], and on the appropriate application timing at the pre-bloom stage for the induction of seedless grapes [31], [35]. On a molecular level, both GA and auxin biosynthesis genes are up-regulated after pollination in grapevines [3], [36], [37]. However, although GA-mediated parthenocarpy is a highly desirable trait for table grapes, how GA induces parthenocarpic fruit development remains unclear. GA application at the pre-bloom stage in grapevines inhibited pollen tube growth and disturbed the balance of GA metabolism at near full bloom [37]. In the present study, changes in the expression levels of the fruit set related genes, VvIAA9, VvARF7, VvARF8, and VvDELLA in grapevines were monitored to determine whether GA application coordinates auxin signaling during parthenocarpic fruit development.
Materials and Methods
Plant Material and GA Application
Five-year-old grapevines of the seeded diploid cultivar ‘Tamnara’ (Vitis spp.), grown in an overhead arbor system, were used for the GA application and gene expression analysis. The cultivar used in this study was bred from a cross between ‘Campbell Early’ (V. labruscana) and ‘Himrod’ (Vitis spp.) at the National Institute of Horticultural and Herbal Science, Suwon, Republic of Korea in 1998 [38]. A GA solution (Dongbu, Seoul, Korea) at 100 ppm was applied as described by Okamoto and Miura [35] onto inflorescence clusters 14 days before full bloom (DBF), which corresponded to the stage showing eight separated leaves and a compact grouped flower, based on the E-L system of Coombe [39], and they were labeled. Clusters were harvested at 0, 1, 2, 4, 7, 9, 12, 14, 16, and 19 days after GA application. Harvested inflorescence samples were immediately frozen in liquid N2, and stored at −80°C until RNA extraction.
Protein Identification
The Vitis homologous proteins ARF7, ARF8, YUCCA2 (YUC2), YUC6, and DELLA were identified using a BLASTP search, except for the previously identified Vitis IAA9 (VvIAA9) (HQ337788) [40] and VvGAI1 (XP_002284648) [41], [42]. Amino acid sequences for the Arabidopsis ARF, Aux/IAA, YUC, and DELLA family proteins were obtained from the National Center for Biotechnology Information (NCBI). For tomato, however, amino acid sequences of the homologues were acquired from previous genome-wide studies [43]–[45]. Using these sequences, iTAK (http://bioinfo.bti.cornell.edu/cgi-bin/itak/index.cgi), the Plant Transcription Factor Database, version 3.0 (PlnTFDB, http://plntfdb.bio.uni-potsdam.de/v3.0/), and GreenPhyl (http://www.greenphyl.org/cgi-bin/index.cgi) were screened, and protein sequences were confirmed using the NCBI. The gene sequences of VvARF7 (GSVIVT01015035001), VvARF8 (GSVIVT01035204001), VvYUC2 (GSVIVT01015388001), VvYUC6 (GSVIVT01035678001), and VvDELLA (GSVIVT01030735001) were identified using the Grape Genome Browser, version 12X (http://www.genoscope.cns.fr/externe/GenomeBrowser/Vitis/). Distinct domains for each protein were predicted using PROSITE (http://prosite.expasy.org/prosite.html), Pfam (http://pfam.janelia.org/), and previous reports of ARFs [13], [46], [47], YUCs [48], [49], and DELLAs [50]. Accession numbers for all amino acid sequences used in this study are listed in Table S1.
Phylogenetic Analysis
Protein sequence alignments were generated using ClustalW version 2.1 for multiple alignments, and phylogenetic analyses were performed using neighbor-joining algorithms of the MEGA5 program [51] with the pairwise-deletion option. One thousand replicates were used in the bootstrap analysis.
Total RNA Isolation and cDNA Synthesis
Total RNA was isolated from whole inflorescence samples including berries and pedicels, at various developmental stages according to an RNA extraction method [52] modified to remove polysaccharides and phenolic compounds. The cDNA was synthesized by reverse transcription of 0.5 µg RNA using the PrimeScript first-strand cDNA Synthesis Kit (Takara, Tokyo, Japan) with an oligo-dT primer, according to the manufacturer’s instructions.
Quantitative Reverse Transcription-polymerase Chain Reaction (qRT-PCR)
cDNA was subjected to qRT-PCR using the gene-specific forward and reverse primers shown in Table S2. Primers used in previous studies [36], [53] were used for the grapevine auxin biosynthesis gene, the putative Vitis anthranilate synthase β subunit-like (VvASB1-like), and two auxin signaling Vitis Gretchen Hagen3 (GH3) family genes. qRT-PCR was performed with the SYBR Premix Ex Taq (Takara) on a Thermal Cycler Dice Real-Time System TP800, version 4.0 (Takara), under universal thermal cycling conditions described by the manufacturer. Vitis Actin1 was used as an endogenous control for normalization of gene expression. With cycle threshold (CT) values obtained from the qRT-PCR results, the ΔCT value (CT target gene CT VvActin1) was calculated for each gene. The relative expression of each gene normalized to the ΔCT value of samples from 14 DBF was determined using the comparative CT method (2ΔΔCT). Analysis of qRT-PCR efficiency showed that all amplicons of all genes used in this study were in the optimal range of 95–105% (Figure S1).
Results
Characterization and Identification of Vitis ARF7 and ARF8
Two ARF family proteins, VvARF7 and VvARF8, were identified from several Vitis transcription factor databases. VvARF7 and VvARF8 were previously reported as VvIAA24 and VvIAA7, respectively [54]. Using several plant transcription databases and PROSITE and Pfam, the VvARF7 amino acid sequence was deduced to contain an N-terminal B3-type DNA-binding domain (DBD; amino acids 126–228), a middle region (MR), two C-terminal Aux/IAA dimerization domains, domain III (amino acids 753–788), and domain IV (amino acids 797–839), needed for typical ARF protein activities. The VvARF8 sequence was also deduced to contain an N-terminal DBD (amino acids 128–230), a MR, two C-terminal Aux/IAA dimerization domains, domain III (amino acids 721–756), and domain IV (amino acids 764–807) (Figure 1A). Furthermore, the VvARF7 sequence showed 63 and 61% identity with AtARF7 and SlARF7, respectively, and VvARF8 had 68% identity with AtARF8 and more than 71% identity with SlARF8 (Figure S2). Thus, we renamed these proteins VvARF7 and VvARF8 to comply with the nomenclature of the Arabidopsis ARFs, based on the similarity between these proteins. Phylogenetic analysis and comparison of VvARF7 and VvARF8 with Arabidopsis and tomato ARFs showed that both proteins were clustered with the transcription-activating AtARFs, AtARF5, 6, 7, 8, and 19, according to Guilfoyle and Hagen [55] (Figure 1B).
Figure 1. Protein sequence alignment and phylogenetic analysis of VvARF7 and VvARF8.

(A) Schematic diagrams of VvARF7 and VvARF8. The length of each protein is denoted. DBD, B3 DNA-binding domain; MR, middle region; III, Aux/IAA dimerization domain III; IV, Aux/IAA dimerization domain IV. (B) Phylogenetic tree of Vitis ARF7 and ARF8 with Arabidopsis and tomato ARF families. Asterisks indicate previously reported SlARF7 and SlARF8 proteins, numbered SlARF9 and SlARF4 [46], respectively, based on their locations on the chromosome.
Down-regulation of VvIAA9 and VvARF7 with GA Application
Using GA applied inflorescence clusters at 14 DBF, which induces parthenocarpy in the seeded diploid ‘Tamnara’ grapevines, relative transcription levels of VvIAA9, VvARF7, and VvARF8 during inflorescence development were analyzed to determine whether GA-induced parthenocarpy in grapevines is associated with auxin-related genes. Without GA application, the expression patterns of VvIAA9 and VvARF7 changed similarly until 7 DBF, except for peak expression times of VvIAA9 and VvARF7 at 2 DBF and full bloom, respectively, following which the expression levels of both genes rapidly declined (Figure 2A–B). VvARF8 also showed the highest expression at 2 DBF, with a rapid down-regulation without GA application. With GA application, VvIAA9 and VvARF7 expressions at 12 DBF remained at approximately 50 and 14% of the levels observed without GA application, respectively. Furthermore, the expression levels of VvIAA9 at 2 DBF and VvARF7 at 5 DBF were significantly lower, dropping to 50 and 37% compared to without GA application, respectively, and VvARF7 expression further reduced to 15% at full bloom, compared to without GA application (Figure 2A–B). However, VvARF8 had a fluctuating expression pattern with GA application, showing a more than 2-fold up-regulation from 14 DBF to 5 DAF, and a down-regulation over 20 and 50% at 12 DBF, and during 5 to 2 DBF, respectively, compared to without GA application (Figure 2C). These results indicate that VvIAA9, VvARF7, and VvARF8 expressions were maintained at high levels before pollination and rapidly declined after pollination, but GA application at pre-bloom, down-regulated VvIAA9 and VvARF7 without pollination.
Figure 2. Transcriptional changes in Vitis negative regulator genes for fruit set initiation.
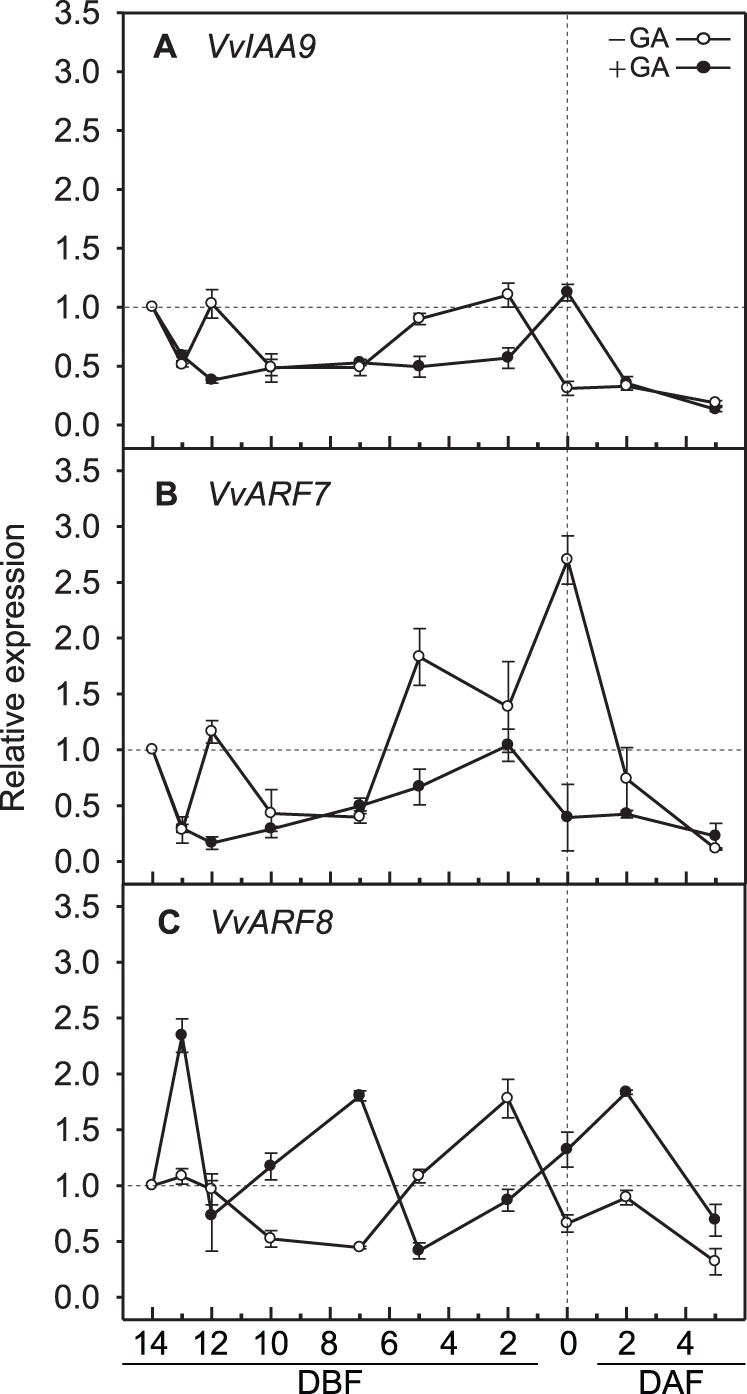
(A) VvIAA9, (B) VvARF7, and (C) VvARF8 in grapevine inflorescences with and without GA application. DAF, days after full bloom; DBF, days before full bloom. Quantitative reverse transcription-PCR was performed to measure the expression level of each gene using the comparative CT method. After normalization of each sample to the expression of the internal control VvActin1, the expression of each gene relative to the 14 DBF sample was calculated. Bars are standard errors of the means from three independent experiments.
Up-regulation of VvGH3.2 and VvGH3.3 with GA Application
To determine whether auxin or auxin signaling was affected by GA application in grapevines, the expression patterns of auxin biosynthesis and the auxin-responsive genes were analyzed during inflorescence development. Auxin biosynthesis was verified by analyzing the expression patterns of the VvASB1-like, VvYUC2, and VvYUC6 genes, which encode key enzymes in auxin biosynthesis. The expression of VvASB1-like was reported by Dauelsberg et al. [36], and VvYUC2 and VvYUC6 were found to be the closest Vitis homologue genes of Arabidopsis YUC2 (AtYUC2) and AtYUC6, and the tomato YUC2 homologue, ToFZY2, which were abundantly expressed in flowers [43], [48]. The VvYUC2 and VvYUC6 proteins shared more than 65% identity and had highly conserved FAD and NADPH binding sites with those of Arabidopsis and tomato (Figure S3). The levels of VvASB1-like transcript fluctuated and peaked at 10 DBF and full bloom, but both VvYUC2 and VvYUC6 were expressed at relatively low levels during inflorescence development without GA application, except for VvYUC2, which was up-regulated at 2 DAF (Figure 3A–C). With GA application, expression patterns of these auxin biosynthesis genes were not significantly different from those without GA application, except for the up-regulations of VvASB1-like, VvYUC2, and VvYUC6 at 7 DBF, full bloom, and 13 DBF, respectively (Figure 3A–C). These results indicate that auxin biosynthesis was not affected by GA application. However, transcription levels of the early auxin-responsive genes were significantly changed following GA application. Without GA application, the expression of VvGH3.2 and VvGH3.3 declined steadily until 2 DBF and then increased after full bloom, with higher expression of VvGH3.2 than VvGH3.3. With GA application, however, VvGH3.2 and VvGH3.3 showed significantly higher levels of transcription at full bloom, with increases of more than 26- and 5-fold, respectively (Figure 3D–E), suggesting that GA application altered auxin signaling.
Figure 3. Transcriptional changes in auxin biosynthesis genes (A, B, and C) and early auxin-responsive genes (D and E).

(A) VvASB1-like, (B) VvYUC2, (C) VvYUC6, (D) VvGH3.2, and (E) VvGH3.3 in grapevine inflorescences with and without GA application. The expression level of VvGH3.3 is magnified in the insets. DAF, days after full bloom; DBF, days before full bloom. Quantitative reverse transcription-PCR was performed to measure the expression level of each gene using the comparative CT method. After normalization of each sample to the expression of the internal control VvActin1, the expression of each gene relative to the 14 DBF sample was calculated. Bars are standard errors of the means from three independent experiments.
Up-regulation of VvDELLA with GA Application
The GA signal was analyzed by monitoring changes in the expression pattern of VvDELLA and VvGAI1, Vitis DELLA family genes, known as key integrators of GA and other hormonal signaling pathways [41], [56]. VvDELLA shares 64 and 66% identity with AtGAI and SlDELLA, respectively, and contains highly conserved functional motifs, such as DELLA, TVHYNP, and GRAS domains (Figure S4A–B). Without GA application, transcription levels of VvDELLA remained low throughout inflorescence development, and the expression levels of VvGAI1 were continuously reduced (Figure 4). These expression patterns of VvGAI1 were not affected by GA application (Figure 4B), but VvDELLA showed different expression patterns. One day after GA application, at 13 DBF, the expression level of VvDELLA was not significantly different from that observed without GA application. However, expression in VvDELLA increased more than 6-fold, from 12 to 7 DBF on GA application, and declined rapidly thereafter (Figure 4A).
Figure 4. Transcriptional changes in two DELLA genes.
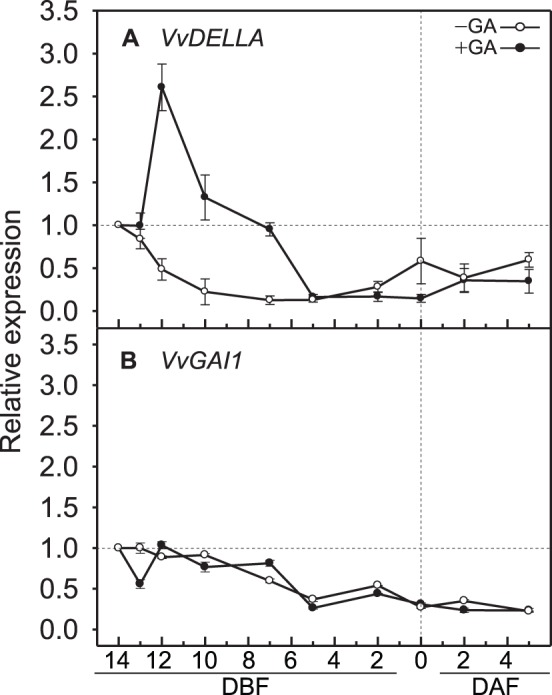
(A) VvDELLA and (B) VvGAI1 in grapevine inflorescences with and without GA application. DAF, days after full bloom; DBF, days before full bloom. Quantitative reverse transcription-PCR was performed to measure the expression level of each gene using the comparative CT method. After normalization of each sample to the expression of the internal control VvActin1, the expression of each gene relative to the 14 DBF sample was calculated. Bars are standard errors of the means from three independent experiments.
Discussion
GA Application Induces Parthenocarpy by Early Down-regulation of Negative Regulators of Fruit Set Initiation
Parthenocarpic fruit development in mutants of the IAA9, ARF7, and ARF8 genes was the result of reduced expression of these genes before pollination [5], [13]–[15]. Without GA application, VvIAA9, VvARF7, and VvARF8 were highly expressed until near full bloom, after which their expressions declined after 2 DBF or full bloom (Figure 2). These results imply that VvIAA9, VvARF7, and VvARF8 function as negative regulators of fruit set initiation, whose down-regulation leads to fruit set initiation. With GA application, however, VvIAA9 and VvARF7 were down-regulated 2 days after the application, and their transcription levels remained low until full bloom or 2 DBF, respectively (Figure 2A–B), in which expression levels of GA biosynthesis genes were down-regulated by GA application at 14 DBF [37]. These results indicate that effects of GA application at pre-bloom were not limited on transcriptional changes of these genes at 1 or 2 days after the application. An enlarged ovary and inhibition of pollen tube growth were observed, both in the pistil of grapevines with GA applied at 14 DBF [35], [37], and in ARF7-silencing lines [13], substantiating the down-regulatory effects of GA application on VvARF7 expression at full bloom. These results also suggest that GA can substitute for the effects of auxin in fruit set initiation in the absence of pollination. Although ARF8 has been reported as a negative regulator of fruit set initiation in Arabidopsis and tomato [14], [15], only truncated, and not null mutants of ARF8 showed parthenocarpic fruit initiation [15], and direct involvement of ARF8 in parthenocarpic fruit development was not observed in the present study.
To verify whether reduced transcription of VvIAA9 and VvARF7 with GA application was due to an increase in auxin levels, transcriptional changes were monitored in the auxin biosynthesis genes, VvASB1-like, VvYUC2, and VvYUC6, and two early auxin-responsive gene, VvGH3.2 and VvGH3.3. VvASB1-like encodes the first step enzyme of auxin biosynthesis [57], and two Vitis YUC genes; VvYUC2 and VvYUC6 genes, encode the final step enzymes [58]. An accumulation of auxin, resulting from the up-regulation of these genes, has been reported in Arabidopsis [49], [57]–[59]. The up-regulation of VvASB1-like upon pollination has also been reported in grapevines [36]. Furthermore, auxin or auxin signaling mediated induction of the GH3 gene family, which encode the early auxin-responsive IAA-amino synthetases, have been reported in Arabidopsis [60], [61] and tomato [13], [22]. In grapevines, six Vitis GH3 family genes have been identified, and both VvGH3.2, the most abundantly expressed Vitis GH3 gene in flower and VvGH3.3, the closest homolog of AtGH3.6 in Vitis [53], showed auxin-inducible expression patterns. With GA application, there was only a slight fluctuation of the expression patterns of VvASB1-like, VvYUC2, and VvYUC6 (Figure 3A–C), however VvGH3.2 and VvGH3.3 were up-regulated more than 3-fold between 2 DBF and full bloom (Figure 3D–E), when VvIAA9 and VvARF7 were down-regulated. With GA application, VvGH3.2 was expressed 4-fold higher at full bloom, compared to without GA application at 2 DAF (Figure 3D), indicating that the up-regulation of VvGH3.2 at full bloom was due to the GA application rather than to ovary development. These results are in agreement with the observed up-regulation of the SlGH3-like gene expression in a silencing line of SlARF7 [13], and also support the idea that GA application down-regulates VvARF7, with partial activation of auxin signaling during parthenocarpic fruit development. However, the possibility of GA-mediated accumulation of auxin could not be excluded, since the peak expression time of VvYUC2 correlated with the highest expression of VvGH3.2 at full bloom with GA application (Figure 3B, D).
Integration of GA and Auxin Signaling during Parthenocarpic Fruit Development via VvDELLA
The crosstalk between GA and auxin in fruit set initiation has been demonstrated by the effects of auxin on GA biosynthesis in Arabidopsis and tomato [29], [30], and the partial involvement of GA signaling in silencing lines of ARF7 in tomato [13], [22]. Using the pro tomato mutant, a loss-of-function mutant of SlDELLA, Carrera et al. [28] demonstrated that GA affected a component of auxin signaling by down-regulating the negative regulators of fruit set initiation; SlIAA9 and SlARF7. Thus, GA and auxin may activate each other in the signaling pathway to a certain extent, and this integration appears to be associated with parthenocarpy induction. The activation of auxin signaling by GA application observed in this study was consistent with the partial auxin signaling activation in silencing lines of SlARF7 [13]. Furthermore, with GA application, the prior up-regulation of VvDELLA decreased of expression levels of VvIAA9 and VvARF7, suggesting that GA application induced parthenocarpic fruits by reducing VvIAA9 and VvARF7 via VvDELLA (Figures 2A–B; 4A). The up-regulation of VvDELLA indicated that its transcription was under a GA-mediated feedback regulation, as observed in SlDELLA [28]. However, VvGAI1, another Vitis DELLA family gene, identified previously as a floral induction related Vitis DELLA family gene [41], did not show transcriptional changes, regardless of GA application, as did the Arabidopsis DELLA family genes. These differential transcriptional regulations by GA application between VvDELLA and VvGAI1 may have originated from the differences in the amino acid sequences of the Poly S/T/N motif, the transcriptional regulatory region of DELLA proteins [56], [62], [63]. A comparison of the amino acid sequences of the Poly S/T/N motif showed that VvDELLA and SlDELLA shared more conserved amino acids than VvGAI1 and the amino acid sequences of VvGAI1 in this motif were similar to those of AtGAI and AtRGA (Figure S4C–D).
Based on the results presented in this study and previous data from de Jong et al. [22] and Carrera et al. [28], we propose a model for fruit set initiation that is mediated by VvARF7, VvIAA9, and VvDELLA in grapevines with or without GA application. Elevated GA and auxin upon pollination initiate fruit set by down-regulating VvIAA9 and VvARF7, negative regulators of fruit set initiation (Figure 5A). With GA application, however, the consecutive transcriptional changes in VvDELLA, VvIAA9, and VvARF7; i.e., the up-regulation of VvDELLA followed by the down-regulation of VvIAA9 and VvARF7 during inflorescence development (Figures 2A–B; 4D), and the early reduction of VvIAA9 and VvARF7 may replace the effects of auxin and initiate parthenocarpic fruit development (Figure 5B).
Figure 5. Proposed model for GA and auxin crosstalk in grapevines during fruit set initiation.

The possible functions of VvARF7, VvIAA9, and VvDELLA are also included, based on de Jong et al. [6] and Carrera et al. [28]. (A) Pollination-mediated fruit set initiation. The dashed line with 26S indicates the regulation of protein levels by 26 s proteasome-mediated degradation, and the arrow indicates activation at the protein level. (B) Putative hormone signaling in fruit set initiation by GA application on grapevine inflorescences. Pathways inactivated by GA application are indicated in gray.
In this study, we reviewed the complex expression patterns of fruit set initiation related genes in grapevines and have shown that GA application at pre-bloom down-regulates VvIAA9 and VvARF7 before pollination and activates some auxin signaling via VvDELLA during parthenocarpic fruit development. This is the first report to detail the molecular mechanism of fruit set initiation in grapevines, and contributes to improving fruit productivity by providing information to increase fruit set initiation of other important crops. The integration of GA and auxin signaling, including the roles of VvIAA9, VvARF7, and VvDELLA should be further investigated to broaden our understanding of the molecular mechanisms underlying GA-mediated induction of parthenocarpic fruit development in viticulture.
Supporting Information
qRT-PCR efficiency plots for GA metabolic genes. Mean quantification cycle (CT) values obtained from 10-fold serial dilution series of each gene plotted against the logarithm of the cDNA template concentration. The amplification efficiency (E) was calculated by E = [10(−1/S) −1]×100, where S = the slope of the linear regression line.
(PDF)
Protein alignment and phylogenetic analysis of VvARF7 and VvARF8. (A) Comparison of the AtARF7, SlARF7, and VvARF7 amino acid sequences. (B) Comparison of the AtARF8, SlARF8, and VvARF8 amino acid sequences. The B3 DNA-binding domain is denoted with an open box. The Aux/IAA dimerization domains III and IV are underlined with solid and dashed lines, respectively. Identical and similar amino acids are shaded in black and gray, respectively.
(PDF)
Protein alignment and phylogenetic analysis of VvYUC2 and VvYUC6. (A) Comparison of the AtYUC2, AtYUC6, ToFZY2, VvYUC2, and VvYUC6 amino acid sequences. The FAD and NADPH binding sites are underlined. Identical and similar amino acids are shaded in black and gray, respectively. (B) A phylogenetic tree comparing VvYUC2 and VvYUC6 with Arabidopsis and tomato YUCCA families.
(PDF)
Protein sequence alignment and phylogenetic analysis of VvDELLA. (A) Comparison of the AtGAI, SlDELL, and VvDELLA amino acid sequences. The DELLA and TVHYNP motifs for GA signal perception, the poly S/T/N motif for regulation of DELLA expression and the NLS motif for nuclear localization are denoted with open boxes. The LHR and VHIID domains for dimerization of DELLA protein are indicated with solid and dashed lines, respectively. The PFYRE and SAW domains that interact with the GA receptor are denoted with dotted lines. The DELLA and GRAS domains are indicated with light and dark gray arrows, respectively. Identical and similar amino acids are shaded in black and gray, respectively. (B) A phylogenetic tree of VvDELLA with five Arabidopsis DELLAs and one SlDELLA. Comparison of the Poly S/T/N motifs (C) between VvDELLA with SlDELLA and (D) between VvGAI1 with AtGAI.
(PDF)
TIGR, SGN, or GenBank accession numbers of the proteins used for the phylogenetic analysis.
(DOCX)
Primers for the qRT-PCR used in this study.
(DOCX)
Acknowledgments
This work was supported by a grant from the Next-Generation BioGreen 21 Program (PJ008213), Rural Development Administration, Suwon, Republic of Korea.
Funding Statement
This work was supported by a grant from the Next-Generation BioGreen 21 Program (PJ008213), Rural Development Administration, Suwon, Republic of Korea. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Gillaspy G, Ben-David H, Gruissem W (1993) Fruits: A developmental perspective. Plant Cell 5: 1439–1451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Coombe BG (1960) Relationship of growth and development to changes in sugars, auxins, and gibberellins in fruit of seeded and seedless varieties of Vitis vinifera . Plant Physiol 35: 241–250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Kuhn N, Arce-Johnson P (2012) Pollination: A key event controlling the expression of genes related to phytohormone biosynthesis during grapevine berry formation. Plant Signal Behav 7: 7–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Ozga JA, Reinecke DM (2003) Hormonal interactions in fruit development. J Plant Growth Regul 22: 73–81. [Google Scholar]
- 5. Wang J, Horiuchi S, Ogata T, Matsui H (1993) Relation between the formation of parthenocarpic berries and endogenous plant hormone contents in seedless grape cultivars. J Jpn Soc Hort Sci 62: 9–14. [Google Scholar]
- 6. de Jong M, Mariani C, Vriezen WH (2009) The role of auxin and gibberellin in tomato fruit set. J Exp Bot 60: 1523–1532. [DOI] [PubMed] [Google Scholar]
- 7. Gustafson FG (1936) Inducement of fruit development by growth-promoting chemicals. Proc Natl Acad Sci USA 22: 628–636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Schwabe WW, Mills JJ (1981) Hormones and parthenocarpic fruit set: A literature survey. Hort Abstr 51: 661–698. [Google Scholar]
- 9. Vivian-Smith A, Koltunow AM (1999) Genetic analysis of growth-regulator-induced parthenocarpy in Arabidopsis . Plant Physiol 121: 437–451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Wittwer SH, Bukovac MJ, Sell HM, Weller LE (1957) Some effects of gibberellin on flowering and fruit setting. Plant Physiol 32: 39–41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Ulmasov T, Murfett J, Hagen G, Guilfoyle TJ (1997) Aux/IAA proteins repress expression of reporter genes containing natural and highly active synthetic auxin response elements. Plant Cell 9: 1963–1971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Wang H, Jones B, Li Z, Frasse P, Delalande C, et al. (2005) The tomato Aux/IAA transcription factor IAA9 is involved in fruit development and leaf morphogenesis. Plant Cell 17: 2676–2692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. de Jong M, Wolters-Arts M, Feron R, Mariani C, Vriezen WH (2009) The Solanum lycopersicum AUXIN RESPONSE FACTOR 7 (SlARF7) regulates auxin signaling during tomato fruit set and development. Plant J 57: 160–170. [DOI] [PubMed] [Google Scholar]
- 14. Goetz M, Hooper LC, Johnson SD, Rodrigues JCM, Vivian-Smith A, et al. (2007) Expression of aberrant forms of AUXIN RESPONSE FACTOR8 stimulates parthenocarpy in Arabidopsis and tomato. Plant Physiol 145: 351–366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Goetz M, Vivian-Smith A, Johnson SD, Koltunow AM (2006) AUXIN RESPONSE FACTOR 8 is a negative regulator of fruit initiation in Arabidopsis . Plant Cell 18: 1873–1886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Finet C, Fourquin C, Vinauger M, Berne-Dedieu A, Chambrier P, et al. (2010) Parallel structural evolution of auxin response factors in the angiosperms. Plant J 63: 952–959. [DOI] [PubMed] [Google Scholar]
- 17. Hardtke CS, Berleth T (1998) The Arabidopsis gene MONOPTEROS encodes a transcription factor mediating embryo axis formation and vascular development. EMBO J 17: 1405–1411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Harper RM, Stowe-Evans EL, Luesse DR, Muto H, Tatematsu K, et al. (2000) The NPH4 locus encodes the auxin response factor ARF7, a conditional regulator of differential growth in aerial Arabidopsis tissue. Plant Cell 12: 757–770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Li H, Johnson P, Stepanova A, Alonso JM, Ecker JR (2004) Convergence of signaling pathways in the control of differential cell growth in Arabidopsis . Dev Cell 7: 193–204. [DOI] [PubMed] [Google Scholar]
- 20. Li J, Dai X, Zhao Y (2006) A role for auxin response factor 19 in auxin and ethylene signaling in Arabidopsis . Plant Physiol 140: 899–908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Schruff MC, Spielman M, Tiwari S, Adams S, Fenby N, et al. (2006) The AUXIN RESPONSE FACTOR 2 gene of Arabidopsis links auxin signaling, cell division, and the size of seeds and other organs. Development 133: 251–261. [DOI] [PubMed] [Google Scholar]
- 22. de Jong M, Wolters-Arts M, García-Martínez JL, Mariani C, Vriezen WH (2011) The Solanum lycopersicum AUXIN RESPONSE FACTOR 7 (SlARF7) mediates cross-talk between auxin and gibberellin signalling during tomato fruit set and development. J Exp Bot 62: 617–626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Fos M, Nuez F, García-Martínez JL (2000) The gene pat-2, which induces natural parthenocarpy, alters the gibberellin content in unpollinated tomato ovaries. Plant Physiol 122: 471–479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Fos M, Proano K, Nuez F, García-Martínez JL (2001) Role of gibberellins in parthenocarpic fruit development induced by the genetic system pat-3/pat-4 in tomato. Physiol Plant 111: 545–550. [DOI] [PubMed] [Google Scholar]
- 25. Olimpieri I, Siligato F, Caccia R, Mariotti L, Ceccarelli N, et al. (2007) Tomato fruit set driven by pollination or by the parthenocarpic fruit allele are mediated by transcriptionally regulated gibberellin biosynthesis. Planta 226: 877–888. [DOI] [PubMed] [Google Scholar]
- 26. Fuentes S, Ljung K, Sorefan K, Alvey E, Harberd NP, et al. (2012) Fruit growth in Arabidopsis occurs via DELLA-dependent and DELLA-independent gibberellin responses. Plant Cell 24: 3982–3996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Marti C, Orzaez D, Ellul P, Moreno V, Carbonell J, et al. (2007) Silencing of DELLA induces facultative parthenocarpy in tomato fruits. Plant J 52: 865–876. [DOI] [PubMed] [Google Scholar]
- 28. Carrera E, Ruiz-Rivero O, Peres LEP, Atares A, García-Martínez JL (2012) Characterization of the procera tomato mutant shows novel functions of the SlDELLA protein in the control of flower morphology, cell division and expansion, and the auxin-signaling pathway during fruit-set and development. Plant Physiol 160: 1581–1596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Dorcey E, Urbez C, Blazquez MA, Carbonell J, Perez-Amador MA (2009) Fertilization-dependent auxin response in ovules triggers fruit development through the modulation of gibberellin metabolism in Arabidopsis . Plant J 58: 318–332. [DOI] [PubMed] [Google Scholar]
- 30. Serrani JC, Ruiz-Rivero O, Fos M, García-Martínez JL (2008) Auxin-induced fruit-set in tomato is mediated in part by gibberellins. Plant J 56: 922–934. [DOI] [PubMed] [Google Scholar]
- 31. Lu J, Lamikanra O, Leong S (1997) Induction of seedlessness in ‘Triumph’ muscadine grape (Vitis rotundifolia Michx) by applying gibberellic acid. HortScience 32: 89–90. [Google Scholar]
- 32. Casanova L, Casanova R, Moret A, Agusti A (2009) The application of gibberellic acid increases berry size of ‘Emperatriz’ seedless grape. Span J Agr Res 7: 919–927. [Google Scholar]
- 33. Dokoozlian NK, Peacock WL (2001) Gibberellic acid applied at bloom reduces fruit set and improves size of ‘Crimson Seedless’ table grapes. HortScience 36: 706–709. [Google Scholar]
- 34. Iwahori S, Weaver RJ, Pool RM (1968) Gibberellin-like activity in berries of seeded and seedless Tokay grapes. Plant Physiol 43: 333–337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Okamoto G, Miura K (2005) Effect of pre-bloom GA application on pollen tube growth in cv. Delaware grape pistils. Vitis 44: 157–159. [Google Scholar]
- 36. Dauelsberg P, Matus JT, Poupin MJ, Leiva-Ampuero A, Godoy F, et al. (2011) Effect of pollination and fertilization on the expression of genes related to floral transition, hormone synthesis, and berry development in grapevine. J Plant Physiol 168: 1667–1674. [DOI] [PubMed] [Google Scholar]
- 37. Jung CJ, Hur YY, Jung SM, Noh JH, Do GR, et al. (2014) Transcriptional changes of gibberellin oxidase genes in grapevines with or without gibberellin application during inflorescence development. J Plant Res 127: 359–371. [DOI] [PubMed] [Google Scholar]
- 38. Park KS, Yun HK, Suh HS, Jeong SB, Cho HM (2004) Breeding of early season grape cultivar ‘Tamnara’ (Vitis hybrid) with high quality and disease resistance. Kor J Hort Sci Technol 22: 458–461. [Google Scholar]
- 39. Coombe BG (1995) Adoption of a system for identifying grapevine growth stages. Aust J Grape Wine Res 1: 100–110. [Google Scholar]
- 40. Fujita K, Horiuchi H, Takato H, Kohno M, Suzuki S (2012) Auxin-responsive grape Aux/IAA9 regulates transgenic Arabidopsis plant growth. Mol Biol Rep 39: 7823–7829. [DOI] [PubMed] [Google Scholar]
- 41. Boss PK, Thomas MR (2002) Association of dwarfism and floral induction with a grape ‘green revolution’ mutation. Nature 416: 847–850. [DOI] [PubMed] [Google Scholar]
- 42. Zhong GY, Yang Y (2012) Characterization of grape Gibberellin Insensitive1 mutant alleles in transgenic Arabidopsis . Transgenic Res 21: 725–741. [DOI] [PubMed] [Google Scholar]
- 43. Expósito-Rodríguez M, Borges AA, Borges-Pérez A, Pérez JA (2011) Gene structure and spatiotemporal expression profile of tomato genes encoding YUCCA-like flavin monooxygenases: The ToFZY gene family. Plant Physiol Biochem 49: 782–791. [DOI] [PubMed] [Google Scholar]
- 44. Kumar R, Agarwal P, Tyagi AK, Sharma AK (2012) Genome-wide investigation and expression analysis suggest diverse roles of auxin-responsive GH3 genes during development and response to different stimuli in tomato (Solanum lycopersicum). Mol Genet Genom 287: 221–235. [DOI] [PubMed] [Google Scholar]
- 45. Wu J, Peng Z, Liu SY, He Y, Cheng L, et al. (2012) Genome-wide analysis of Aux/IAA gene family in Solanaceae species using tomato as a model. Mol Genet Genom 287: 295–311. [DOI] [PubMed] [Google Scholar]
- 46. Kumar R, Tyagi AK, Sharma AK (2011) Genome-wide analysis of auxin response factor (ARF) gene family from tomato and analysis of their role in flower and fruit development. Mol Genet Genom 285: 245–260. [DOI] [PubMed] [Google Scholar]
- 47. Ulmasov T, Hagen G, Guilfoyle TJ (1999) Dimerization and DNA binding of auxin response factors. Plant J 19: 309–319. [DOI] [PubMed] [Google Scholar]
- 48. Cheng Y, Dai X, Zhao Y (2006) Auxin biosynthesis by the YUCCA flavin monooxygenases controls the formation of floral organs and vascular tissues in Arabidopsis . Gene Dev 20: 1790–1799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Kim JI, Murphy AS, Baek D, Lee SW, Yun DJ, et al. (2011) YUCCA6 over-expression demonstrates auxin function in delaying leaf senescence in Arabidopsis thaliana . J Exp Bot 62: 3981–3992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Bassel GW, Zielinska E, Mullen RT, Bewley JD (2004) Down-regulation of DELLA genes is not essential for germination of tomato, soybean, and Arabidopsis seeds. Plant Physiol 136: 2782–2789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Tamura K, Peterson D, Peterson N, Stecher G, Nei M, et al. (2011) MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol 28: 2731–2739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Chang S, Puryear J, Cairney J (1993) A simple and efficient method for isolating RNA from pine trees. Plant Mol Biol Rep 11: 113–116. [Google Scholar]
- 53. Böttcher C, Boss PK, Davies C (2011) Acyl substrate preferences of an IAA-amido synthetase account for variations in grape (Vitis vinifera L.) berry ripening caused by different auxinic compounds indicating the importance of auxin conjugation in plant development. J Exp Bot 62: 4267–4280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Çakir B, Kiliçkaya O, Olcay AC (2013) Genome-wide analysis of Aux/IAA genes in Vitis vinifera: Cloning and expression profiling of a grape Aux/IAA gene in response to phytohormone and abiotic stresses. Acta Physiol Plant 35: 365–377. [Google Scholar]
- 55. Guilfoyle TJ, Hagen G (2007) Auxin response factors. Curr Opin Plant Biol 10: 453–460. [DOI] [PubMed] [Google Scholar]
- 56. Itoh H, Ueguchi-Tanaka M, Sato Y, Ashikari M, Matsuoka M (2002) The gibberellin signaling pathway is regulated by the appearance and disappearance of SLENDER RICE1 in nuclei. Plant Cell 14: 57–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Stepanova AN, Hoyt JM, Hamilton AA, Alonso JM (2005) A link between ethylene and auxin uncovered by the characterization of two root-specific ethylene-insensitive mutants in Arabidopsis . Plant Cell 17: 2230–2242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Mashiguchi K, Tanaka K, Sakai T, Sugawara S, Kawaide H, et al. (2011) The main auxin biosynthesis pathway in Arabidopsis . Proc Natl Acad Sci USA 108: 18512–18517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Mano Y, Nemoto K (2012) The pathway of auxin biosynthesis in plants. J Exp Bot 63: 2853–2872. [DOI] [PubMed] [Google Scholar]
- 60. Nakazawa M, Yabe N, Ichikawa T, Yamamoto YY, Yoshizumi T, et al. (2001) DFL1, an auxin-responsive GH3 gene homologue, negatively regulates shoot cell elongation and lateral root formation, and positively regulates the light response of hypocotyl length. Plant J 25: 213–221. [DOI] [PubMed] [Google Scholar]
- 61. Staswick PE, Serban B, Rowe M, Tiryaki I, Maldonado MT, et al. (2005) Characterization of an Arabidopsis enzyme family that conjugates amino acids to indole-3-acetic acid. Plant Cell 17: 616–627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Sheerin DJ, Buchanan J, Kirk C, Harvey D, Sun X, et al. (2011) Inter- and intra-molecular interactions of Arabidopsis thaliana DELLA protein RGL1. Biochem J 435: 629–639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Sun TP (2008) Gibberellin metabolism, perception, and signaling pathways in Arabidopsis . Arabidopsis Book 6: e0103. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
qRT-PCR efficiency plots for GA metabolic genes. Mean quantification cycle (CT) values obtained from 10-fold serial dilution series of each gene plotted against the logarithm of the cDNA template concentration. The amplification efficiency (E) was calculated by E = [10(−1/S) −1]×100, where S = the slope of the linear regression line.
(PDF)
Protein alignment and phylogenetic analysis of VvARF7 and VvARF8. (A) Comparison of the AtARF7, SlARF7, and VvARF7 amino acid sequences. (B) Comparison of the AtARF8, SlARF8, and VvARF8 amino acid sequences. The B3 DNA-binding domain is denoted with an open box. The Aux/IAA dimerization domains III and IV are underlined with solid and dashed lines, respectively. Identical and similar amino acids are shaded in black and gray, respectively.
(PDF)
Protein alignment and phylogenetic analysis of VvYUC2 and VvYUC6. (A) Comparison of the AtYUC2, AtYUC6, ToFZY2, VvYUC2, and VvYUC6 amino acid sequences. The FAD and NADPH binding sites are underlined. Identical and similar amino acids are shaded in black and gray, respectively. (B) A phylogenetic tree comparing VvYUC2 and VvYUC6 with Arabidopsis and tomato YUCCA families.
(PDF)
Protein sequence alignment and phylogenetic analysis of VvDELLA. (A) Comparison of the AtGAI, SlDELL, and VvDELLA amino acid sequences. The DELLA and TVHYNP motifs for GA signal perception, the poly S/T/N motif for regulation of DELLA expression and the NLS motif for nuclear localization are denoted with open boxes. The LHR and VHIID domains for dimerization of DELLA protein are indicated with solid and dashed lines, respectively. The PFYRE and SAW domains that interact with the GA receptor are denoted with dotted lines. The DELLA and GRAS domains are indicated with light and dark gray arrows, respectively. Identical and similar amino acids are shaded in black and gray, respectively. (B) A phylogenetic tree of VvDELLA with five Arabidopsis DELLAs and one SlDELLA. Comparison of the Poly S/T/N motifs (C) between VvDELLA with SlDELLA and (D) between VvGAI1 with AtGAI.
(PDF)
TIGR, SGN, or GenBank accession numbers of the proteins used for the phylogenetic analysis.
(DOCX)
Primers for the qRT-PCR used in this study.
(DOCX)