

Abstract
The serotonergic feeding circuit in Drosophila melanogaster larvae can be used to investigate neuronal substrates of critical importance during the development of the circuit. Using the functional output of the circuit, feeding, changes in the neuronal architecture of the stomatogastric system can be visualized. Feeding behavior can be recorded by observing the rate of retraction of the mouth hooks, which receive innervation from the brain. Locomotor behavior is used as a physiological control for feeding, since larvae use their mouth hooks to traverse across an agar substrate. Changes in feeding behavior can be correlated with the axonal architecture of the neurites innervating the gut. Using immunohistochemistry it is possible to visualize and quantitate these changes. Improper handling of the larvae during behavior paradigms can alter data as they are very sensitive to manipulations. Proper imaging of the neurite architecture innervating the gut is critical for precise quantitation of number and size of varicosities as well as the extent of branch nodes. Analysis of most circuits allow only for visualization of neurite architecture or behavioral effects; however, this model allows one to correlate the functional output of the circuit with the impairments in neuronal architecture.
Keywords: Neuroscience, Issue 81, Neural Pathways, Drosophila, Microscopy, Neuroimaging, Behavior, Behavior Mechanisms, Dopamine, Immunohistochemistry, neurite, proventriculus, serotonin, varicosities, animal model
Introduction
Drosophila is an extremely powerful model system for studying neural circuit development due to rapid generation time, low experimental cost, and the ability to manipulate and control genetic and environmental factors. Neurogenesis, neuronal path finding and synaptogenesis are conserved between humans and Drosophila; therefore the mechanisms in creating, maintaining and modifying neural circuits are conserved as well.
Classical neurotransmitters, such as serotonin (5-hydroxytryptamine, or 5-HT) can serve as growth factors before adopting their roles as signaling molecules in the mature neural circuit1-3Previous studies have shown that perturbed levels of 5-HT during embryogenesis alter the connectivity of mature neurons4. Others have shown that ectopic application of 5-HT to cultured Helisoma neurons repress neurite outgrowth as well as synaptogenesis5-7. In Drosophila, developmental 5-HT levels are inversely related to varicosity number and size, as well as the degree of aborization, along the length of neurites projecting to the foregut from the CNS8.
Serotonergic neurotransmission has been shown to modulate feeding behaviors in diverse species, including Drosophila8-9. The feeding circuit in Drosophila is a relatively simple circuit that can be used as a model to correlate the functional output (feeding) with alterations in the development of the axonal projections from the brain to the foregut. Schoofs et al. have shown that Drosophila larval feeding is regulated by central pattern generators that influence the musculature10. While the specific muscular anatomy is not completely understood, it has been shown that the antennal nerve, maxillary nerve, and prothoracic accessory nerve are responsible for the muscular targets involved in the feeding behavior. Most data involving the musculature and nerve anatomy of invertebrate feeding is limited to Calliphora larvae.
The feeding rate of second instar larvae can be assessed by retraction of the cephalopharyngeal sclerites (mouth hooks), and is reproducible and high-throughput. The cephalopharyngeal plates are innervated by fibers from central 5-HT neurons via the frontal nerve. The proventriculus, or foregut, is innervated by serotonergic fibers (recurrens nerve) that fasciculate in the midgut and are responsible for contractions of the foregut (Figure 1)11-12. Changes in axonal branching, and the number and size of varicosities along the neurite length, can be quantitated using immunohistochemical techniques. Manipulating neuronal 5-HT during development, either directly or indirectly, can change the functional output of this feeding circuit, which can be assessed and correlated with changes in the morphology of the neurite architecture.
Protocol
1. Maintenance of Population Cages
Maintain population cages at 25 °C on a 12 hr light-dark cycle. As long as the control and experimental groups are exposed to the same lighting conditions, then this technique can be performed in a standard laboratory setting.
Allow females to lay eggs overnight on apple juice-agar plates.
Collect newly hatched larvae by maintaining plates with newly deposited eggs at 25 °C for 24 hr. Place a small dollop of yeast in the center of the plate to attract hatched larvae.
Collect 1st instar larvae that have migrated into the yeast paste in the center of the apple juice plate. Use a metal spatula to transfer it to a fresh apple juice plate. Additional yeast paste can be added to ensure there is adequate food until the assays are performed. Grape juice plates can also be used in place of apple juice plates.
Obtain late 2nd-early 3rd instar larvae by allowing 1st instars to age for 40-48 hr. Within this range the feeding rates are constant13. Age can be confirmed by examination of the mouth hooks as there are distinct changes in this structure with each larval molt.
Late 2nd-early 3rd instar larvae are collected for behavior assays by gently and extensively washing apple juice plates with water and collecting the larvae on a mesh filter. The larvae are then transferred to an agar plate.
All analyses are performed in parallel using control and experimental animals.
2. Behavioral Paradigm – Locomotion
Place a single 3rd instar larva on a 2% agar substrate in a 100 mm tissue culture dish and allow the larva to acclimate for 30 sec. Larvae use their cephalopharyngeal sclerites (mouth hooks) to propel their bodies across the surface of the agar substrate. This is done on a separate plate because it is more difficult to visualize the body contractions in the yeast solution as the animal is almost the same color as the yeast.
Observe and record each posterior to anterior movement over the substrate for a period of 1 min. n = 20 for each genotype. No more than 10 animals should be assayed per plate.
3. Behavior Paradigm – Feeding
Using blunt Inox #5 tweezers, carefully transfer the 3rd instar larva from the locomotor agar plate to the center of an agar-filled plate overlaid with 5 ml of 2% activated baker's yeast solution. Ensure that the solution is homogenous as the yeast will settle over time. When in the yeast solution, larvae will largely remain in place and feed, facilitating observation of the behavior. The rate of mouth hook contractions directly correlates with the amount of food ingested14.
Allow larva to acclimate for 30 sec.
Observe and record the number of mouth hook contractions for a period of 1 min. n = 20 for each genotype.
4. Larval Gut Dissections
Make a 4% EM-grade formaldehyde fixation solution in 1x phosphate-buffered saline (PBS) and place in a 3-well spot glass.
Carefully dissect out wandering late 3rd instar larval guts in a 3-well glass dish using 1x PBS solution, making sure each time that the proventriculus is left intact. With one forcep, hold the posterior end, and with the other, hold the mouth hooks. While holding the posterior end immobile, gently pull on the mouth hooks to gain access to the guts. Remove any associated tissues (salivary glands, brain, fat body, etc.), then transfer each gut to the 3-well dish containing the formaldehyde fix. Wandering 3rd instar larva are used because at this stage of development, the larva cease feeding in preparation for pupariation, and the proventriculus is cleared of yeast.
Incubate guts at 4 °C overnight in an opaque tissue culture box.
Before removing formaldehyde fixation solution from wells, remove the gastric caeca and clip the midgut ~150 µm2 from the proventriculus so that projections can clearly be viewed without impediment.
Remove formaldehyde fix from the wells and replace with 1x PBT (1x PBS, 0.1% protease-free bovine serum albumin, 0.1% Triton X-100) buffer solution. Thoroughly wash guts 6x for 10 min in 1x PBT. Place tissue samples on a mechanical rotator while doing washes.
Incubate at 4 °C for 1 hr in 10-6 M 5-HT to enhance serotonin signaling. Thoroughly wash guts 6x for 10 min in 1x PBT. Previous studies have demonstrated that this concentration of exogenous 5-HT does not affect neuronal architecture or varicosity density in immunohistochemical analyses and simply enhances the signal to noise ratio15-16.
Incubate at 4 °C overnight in anti-serotonin primary antibody (monoclonal raised in mouse or polyclonal raised in rabbit). Thoroughly wash guts 6x for 10 min in 1x PBT. Place tissue samples on a mechanical rotator while doing washes.
Incubate at 4 °C for 90 min in secondary antibody (Alexa Fluor 568 goat anti-mouse or anti-rabbit IgG; 1:400 dilution). Thoroughly wash guts 6x for 10 min in 1x PBT. Place tissue samples on a mechanical rotator while doing washes.
Incubate in 4 mM sodium carbonate for 10 min on a mechanical rotator, then mount in 4% n-propyl gallate/20 mM sodium carbonate, and view under fluorescence. Sodium carbonate is used to convert the samples to the buffer and pH used in the mountant media.
Capture images of immunostained tissue samples at 400X magnification for analysis.
5. Analysis of Neural Circuitry
Neurite fibers were quantified (number and size of varicosities and degree of branching) using Neuroleucida and Neuroexplorer. However, this can also be accomplished manually or using Simple Neurite Tracer (a free application that can be downloaded online).
Trace the individual fiber projections from the brain to the proventriculus and quantify varicosity number, branches and the number of large varicosities per unit length. The axonal fibers projecting from the brain are bundled into the recurrens nerve and are not accessible for analysis until they reach the proventriculus where they separate and fasciculate.
Representative Results
The serotonergic feeding circuit in the Drosophila larva can serve as an extremely effective model to observe the influence of particular factors on nervous system development. By quantitating feeding rate, it is possible to link the axonal architecture of the feeding circuit with its functional output (Figure 1). The locomotor assay is used as a physiological control for the retractions of the mouth hooks, since larvae use their mouth hooks to propel themselves across the surface of the agar. There should be no difference in the locomotor responses between control and mutant genotypes if the mutations only affect the feeding circuit8 (Figure 2A). If significant differences do occur, it is possible the larval behavior was compromised by improper handling. If the larvae stop during the assay to attempt to burrow through the agar substrate, they may be too old, and are likely transitioning to wandering instars. It is also possible the agar substrate may be too hard, thus making it difficult for larval mouth hooks to grab the agar substrate; this can be addressed by wetting the agar surface.
This assay can be used to assess whether Drosophila strains with neuronal anatomic defects affect development of the serotonergic feeding circuit. The mutant ellipsoid body open (ebo3) has a structural defect in the ellipsoid body of the central complex. Comparison with the wild-type parental Canton-S strain, CSwu, reveals that these anatomical defects during brain development result in depressed feeding while locomotion is unaffected (Figure 2B).
The anatomical defects in ebo3 mutants appear to change the development of the neurite architecture of the gut. Figure 3 shows the changes in fiber architecture in the ebo3 larvae compared with CSwu; these larvae display an increase in branching as well as an increase in the number of both small and large varicosities along the neurite length. Note the branch nodes (arrows), varicosities (arrowheads), and large varicosities (asterisks). Figure 4 represents the quantitation of these images.
Proper quantitation of the axonal architecture requires that the images be extremely clear. Figure 5A represents an image appropriate for analysis. Images of poorer quality will make it difficult to distinguish between the fiber and varicosities (Figure 5B). When photographing the fiber architecture, avoid taking images that include the projections at the anterior of the proventriculus, since the fibers are tightly bundled and are separating from each other and may appear as though they are branched. More posterior fibers within the midgut are more branched because they fasciculate once they are in within this tissue. Quantitation of branch and varicosity number, and size of varicosities, can be analyzed manually or via a program designed for the purpose of studying neurite morphology, such as Neuroleucida. As long as the proventriculus is not damaged during the immunohistochemical protocol, and the image is in focus, the preparations will be acceptable for imaging and analysis. If the fiber architecture is clearly distinguishable from background, and if individual varicosities can be identified along the neurite length, the preparation is appropriate for analysis. Also, if individual varicosities can be identified from the rest of the fiber, this is also another indicator of a quality image for analysis. All fibers are analyzed with the exception of those that out of the range of focus (in some cases the fibers will curve between multiple planes of focus).
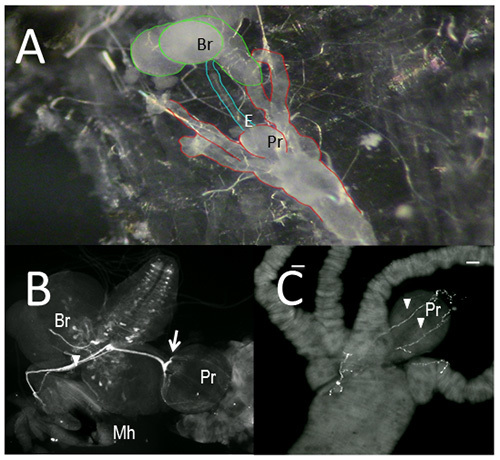 Figure 1. The larval feeding circuit. Fillet a of 3rd instar larva showing brain and gut tissues (A). Gut tissues dissected from 3rd instar larvae were immunostained with an antibody raised against Drosophila neuronal tryptophan hydroxylase (DTRH, B) or 5-HT (C).
A,B. E, esophagus; Mh, mouth hooks; Pr, proventriculus; Br, brain (note the pattern of 5-HT neurons). Arrowhead designates the frontal nerve; arrow, the recurrens nerve. C. proventriculus showing axonal fibers (arrowheads). Scale bar = 20 µm. Click here to view larger image.
Figure 1. The larval feeding circuit. Fillet a of 3rd instar larva showing brain and gut tissues (A). Gut tissues dissected from 3rd instar larvae were immunostained with an antibody raised against Drosophila neuronal tryptophan hydroxylase (DTRH, B) or 5-HT (C).
A,B. E, esophagus; Mh, mouth hooks; Pr, proventriculus; Br, brain (note the pattern of 5-HT neurons). Arrowhead designates the frontal nerve; arrow, the recurrens nerve. C. proventriculus showing axonal fibers (arrowheads). Scale bar = 20 µm. Click here to view larger image.
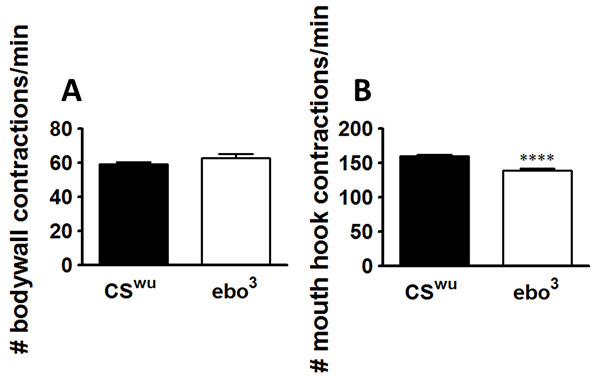 Figure 2. Anatomical defects during brain development results in depressed feeding behavior. The animals were assayed for locomotor (A) and feeding behaviors (B). Locomotion was unaffected. n = 20 for each behavioral assay, from 2-3 independent experiments. ****p<0.0001, unpaired t-test. Lines above the graph depict standard error of the mean. Click here to view larger image.
Figure 2. Anatomical defects during brain development results in depressed feeding behavior. The animals were assayed for locomotor (A) and feeding behaviors (B). Locomotion was unaffected. n = 20 for each behavioral assay, from 2-3 independent experiments. ****p<0.0001, unpaired t-test. Lines above the graph depict standard error of the mean. Click here to view larger image.
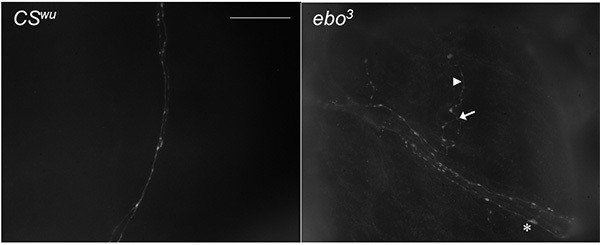 Figure 3. Anatomical defects during development of the CNS results in aberration in gut fiber architecture. Gut tissue dissected from 3rd instar larvae and immunostained with anti-5-HT. Arrow denotes branch node. Arrowhead denotes a small varicosity. Asterisk denotes large varicosity. Scale bar = 40 µm. Click here to view larger image.
Figure 3. Anatomical defects during development of the CNS results in aberration in gut fiber architecture. Gut tissue dissected from 3rd instar larvae and immunostained with anti-5-HT. Arrow denotes branch node. Arrowhead denotes a small varicosity. Asterisk denotes large varicosity. Scale bar = 40 µm. Click here to view larger image.
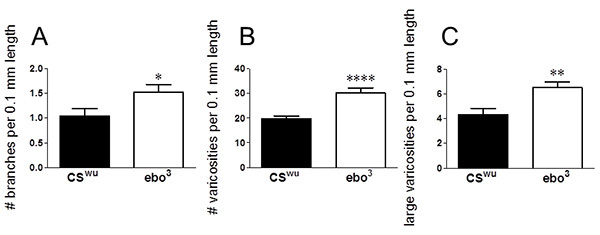 Figure 4. Anatomical defects during brain development result in aberrant gut fiber architecture. Analysis of proventricular tissue from 3rd instar larvae dissected and incubated with anti-5-HT. Neurite branching (A), number of total varicosities per 0.1 mm neurite length (B) and number of large varicosities (>1µm2) per 0.1 mm length (C). CSwu, 20 fibers from 17 guts from 2 independent experiments; ebo3, 20 fibers from 18 guts from 3 independent experiments. ****p<0.0001, **p<0.01, *p<0.05, unpaired t-test. Lines above graph depict standard error of the mean. Click here to view larger image.
Figure 4. Anatomical defects during brain development result in aberrant gut fiber architecture. Analysis of proventricular tissue from 3rd instar larvae dissected and incubated with anti-5-HT. Neurite branching (A), number of total varicosities per 0.1 mm neurite length (B) and number of large varicosities (>1µm2) per 0.1 mm length (C). CSwu, 20 fibers from 17 guts from 2 independent experiments; ebo3, 20 fibers from 18 guts from 3 independent experiments. ****p<0.0001, **p<0.01, *p<0.05, unpaired t-test. Lines above graph depict standard error of the mean. Click here to view larger image.
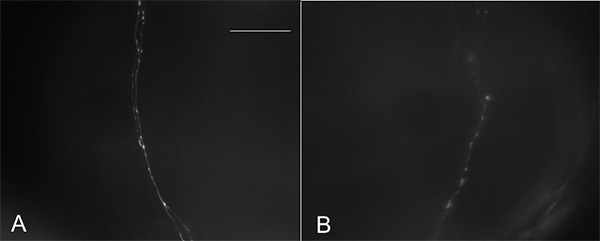 Figure 5. Quality of images is important for proper quantitation of gut fiber architecture. Gut tissues dissected from CSwu 3rd instar larvae and immunostained with anti-5-HT. (A). Good quality image. (B). Poor quality image. Scale bar = 40 µm. Click here to view larger image.
Figure 5. Quality of images is important for proper quantitation of gut fiber architecture. Gut tissues dissected from CSwu 3rd instar larvae and immunostained with anti-5-HT. (A). Good quality image. (B). Poor quality image. Scale bar = 40 µm. Click here to view larger image.
Discussion
Aberrant development of the serotonergic stomatogastric circuit, which occurs during late embryogenesis, will affect its mature function. Changes in the neurite architecture innervating the gut can be correlated with the functional output of the circuit, which is feeding rate (measured by mouth hook contractions in a yeast solution) (Figure 1). The use of the UAS-Gal4 bipartite system in Drosophila makes it possible to specifically target up- or down-regulated expression of a given transcript to a specific tissue; changes in expression of a given protein can be precisely quantitated given the appropriate tools. This technique can be used to elucidate the brain regions, and even neuronal subsets necessary for development of a specific neural circuit.
The locomotor assay is used to confirm that the larvae are not otherwise physiologically compromised; therefore, larvae with 40 body wall contractions or less should be excluded from analyses (Figure 2A). This may occur either because the genotype causes physical abnormalities, or because individual animals have been injured during handling. In addition, the temperature of the room in which the assay is performed needs to be controlled, since cooler or warmer temperatures can affect the data. It is recommended to perform this behavior assay between 24-26 °C. On days when the temperature of the testing room is cooler, the agar has a tendency to harden; therefore, rewetting of the agar plate will be required after each locomotive assay is completed to ensure the agar is sufficiently soft. It is important to keep the agar plate for locomotion moist (not wet) in order to allow the larvae to travel across the surface and to prevent them from burrowing. Cooler temperatures also impact the performance of the larvae on the agar substrate, as larvae tend to travel towards preferential temperatures (24-26 °C)17-18. No more than 10 animals should be assayed per plate, and discard any plate with punctures in the agar substrate.
When performing the feeding assay, it is important to be aware that the yeast suspension will settle over time, Swirling of the yeast on the plate ensures the yeast stays homogenous throughout the assay. Reduced visibility of the mouth hooks can occur if the yeast solution becomes too concentrated. Healthy larvae placed in the yeast media contract their mouth hooks approximately 150-170 times per min; reduced feeding can be as low as 120, and the upper range limit is 210.
When performing the immunohistochemical protocol, it is wise to immunostain both control and experimental tissue samples in parallel to ensure similar quality of tissue samples. Immunofluorescence of the neurite architecture innervating the gut can be enhanced by incubating tissue samples in primary antibody overnight at 4 °C. The incubation of antibodies at 4 °C improves the signal to noise ratio (Figure 5). The quality of the images is very important, as this is critical for precise quantitation of the fiber architecture (Figure 3) to reveal changes in branching and varicosity number and size (Figure 4). The quality of the tissue samples can be destroyed if the rotator used during washing periods heats up at any point from constant use. While imaging the tissue samples be sure not to leave samples under the microscope light for too long as this will reduce the immunofluorescence of the samples, not only the one currently under focus but surrounding samples as well. Analysis of the fiber architecture can be easily completed with the aid of software, but still can also be accomplished manually. Classification of small and large varicosities refer to area of the varicosities, varicosities measuring greater than 1 µm2 are classified as large varicosities.
Disclosures
We have nothing to disclose.
Acknowledgments
The authors would like to acknowledge the President’s Research Fund from Saint Louis University awarded to W.S.N.
References
- Weiss E, Maness P, Lauder J. Why do neurotransmitters act like growth factors? Perspect Dev Neurobiol. 5:323–335. [PubMed] [Google Scholar]
- Herlenius E, Lagercrantz H. Neurotransmitters and neuromodulators during early human development. Early Hum. Dev. 65:21–37. doi: 10.1016/s0378-3782(01)00189-x. [DOI] [PubMed] [Google Scholar]
- Budnik V, Wu C, White K. Altered branching of serotonin-containing neurons in Drosophila mutants unable to synthesize serotonin and dopamine. J. Neurosci. 1989;9:2866–2877. doi: 10.1523/JNEUROSCI.09-08-02866.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sodhi M, Sanders-Bush E. Serotonin and brain development. International Review of Neurobiol. 2004;59:111–174. doi: 10.1016/S0074-7742(04)59006-2. [DOI] [PubMed] [Google Scholar]
- Goldberg J, Kater S. Expression and function of the neurotransmitter serotonin during development of the Helisoma nervous system. Dev. Biol. 1989;131:483–495. doi: 10.1016/s0012-1606(89)80019-3. [DOI] [PubMed] [Google Scholar]
- Goldberg J. Serotonin regulation of neurite outgrowth in identified neurons from mature and embryonic Helisoma triyolvis. Perspect Dev Neurobiol. 1998;5:373–387. [PubMed] [Google Scholar]
- Haydon P, McCobb P, Kater S. Serotonin selectively inhibits growth cone motility and synaptogenesis of specific identified neurons. Sci. 1984;226:561–564. doi: 10.1126/science.6093252. [DOI] [PubMed] [Google Scholar]
- Neckameyer WS. A trophic role for serotonin in the development of a simple feeding circuit. Dev. Neurosci. 32:217–237. doi: 10.1159/000304888. [DOI] [PubMed] [Google Scholar]
- De Vry J, Schreiber R. Effects of selected serotonin 5-HT 1 and 5-HT 2 receptor agonists on feeding behavior: possible mechanisms of action. Neurosci. Biobehav. Rev. 2000;24:341–353. doi: 10.1016/s0149-7634(99)00083-4. [DOI] [PubMed] [Google Scholar]
- Schoofs A, Niederegger S, van Ooyen A, Heinzel H, Spieß R. The brain can eat: Establishing the existence of a central pattern generator for feeding in third instar larvae of Drosophila virilis and Drosophila melanogaster. J. Insect Physiol. 2010;56:695–705. doi: 10.1016/j.jinsphys.2009.12.008. [DOI] [PubMed] [Google Scholar]
- Spieß R, Schoofs A, Heinzel H. Anatomy of the stomatogastric nervous system associated with the foregut in Drosophila melanogaster and Calliphora vicin third instar larvae. J. Morphol. 2008;269:272–282. doi: 10.1002/jmor.10581. [DOI] [PubMed] [Google Scholar]
- Neckameyer WS, Bhatt P. Neurotrophic actions of dopamine on the development of a serotonergic feeding circuit in Drosophila melanogaster. Biomed Cent NeuroSci. 2012;13:26. doi: 10.1186/1471-2202-13-26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sewall D, Burnet B, Connolly K. Genetic analysis of larval feeding behavior in Drosophila melanogaste. Genet. Res. 1975;24:163–173. doi: 10.1017/s0016672300015196. [DOI] [PubMed] [Google Scholar]
- Joshi A, Mueller L. Evolution of higher feeding rate in Drosophila due to density-dependent natural selection. Evolution. 1988;42:1090–1093. doi: 10.1111/j.1558-5646.1988.tb02527.x. [DOI] [PubMed] [Google Scholar]
- Budnik V, Wu C, White K. Altered branching of serotonin-containing neurons in Drosophila mutants unable to synthesize serotonin and dopamine. J. Neurosci. 1989;9:2866–2877. doi: 10.1523/JNEUROSCI.09-08-02866.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sykes P, Condron B. Development and sensitivity to serotonin of Drosophila varicosities in the central nervous system. Dev. Biol. 2005;286:207–216. doi: 10.1016/j.ydbio.2005.07.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garrity PA, Goodman MB, Samuel AD, Sengupta P. Running hot and cold: behavioral strategies, neural circuits, and the molecular machinery for thermotaxis inC. elegansand Drosophila. Genes Dev. 2010;24:2365–2382. doi: 10.1101/gad.1953710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McKemy DD. Temperature sensing across species. Pflugers Archives. 2007;454:777–791. doi: 10.1007/s00424-006-0199-6. [DOI] [PMC free article] [PubMed] [Google Scholar]