

Abstract
Siderophores are organic compounds with low molecular masses that are produced by microorganisms and plants growing under low iron conditions. The primary function of these compounds is to chelate the ferric iron [Fe(III)] from different terrestrial and aquatic habitats and thereby make it available for microbial and plant cells. Siderophores have received much attention in recent years because of their potential roles and applications in various areas of environmental research. Their significance in these applications is because siderophores have the ability to bind a variety of metals in addition to iron, and they have a wide range of chemical structures and specific properties. For instance, siderophores function as biocontrols, biosensors, and bioremediation and chelation agents, in addition to their important role in weathering soil minerals and enhancing plant growth. The aim of this literature review is to outline and discuss the important roles and functions of siderophores in different environmental habitats and emphasize the significant roles that these small organic molecules could play in applied environmental processes.
Introduction
Iron (Fe) is an essential element for the growth of almost all living microorganisms because it acts as a catalyst in enzymatic processes, oxygen metabolism, electron transfer, and DNA and RNA syntheses (Aguado-Santacruz et al., 2012). Fe is also essential for biofilm formation because it regulates surface motility and stabilizes the polysaccharide matrix (Weinberg, 2004; Chhibber et al., 2013). Under iron-deficient growth conditions, the microbial surface hydrophobicity decreases which alters the surface protein composition and leads to limitation of biofilm formation (Simões et al., 2007). Thus, because of the low bioavailability of Fe in the environment, microorganisms have developed specific uptake strategies such as production of siderophores. Siderophores are metal-chelating agents with low molecular masses (200–2000 Da) that are produced by microorganisms and plants, especially under Fe-limiting conditions (Schwyn and Neilands, 1987). Marine organisms such as phytoplankton (Trick et al., 1983) and cyanobacteria (Armstrong and Van Baalen, 1979) can also produce siderophores. The role of siderophores is primarily to scavenge Fe, but they also form complexes with other essential elements (i.e. Mo, Mn, Co and Ni) in the environment and make them available for microbial cells (Bellenger et al., 2008; Braud et al., 2009a,b). Siderophores are divided into three main families depending on the characteristic functional group, i.e. hydroxamates, catecholates and carboxylates. More than 500 different types of siderophores are known, of which 270 have been structurally characterized (Boukhalfa et al., 2003). Hydroxamate siderophores have a 1:1 stability constant with Fe(III) that nears that of the Fe(III)-EDTA complex (1030), whereas catecholates and carboxylates can form 1:1 complexes with stability constants near that of Fe(III)-EDDHA (1040) (Robert and Chenu, 1992). The formation of Fe(III)–siderophore complexes are affected by pH because of the competition for the free siderophore ligands between free protons and Fe (Albrecht-Gary and Crumbliss, 1998). In nature, Fe has to compete not only against free protons for the siderophore binding sites but also against other metal ions such as divalent cations, including Cd2+, Cu2+, Ni2+, Pb2+ and Zn2+ (Albrecht-Gary and Crumbliss, 1998); trivalent cations, such as Mn3+, Co3+ and Al3+; and actinides, such as Th4+, U4+ and Pu4+ (Peterson et al., 2004). There are several studies that have shown that siderophores have an impact on the mobility of these metal ions in the environment (e.g. Renshaw et al., 2002; Dahlheimer et al., 2007; Schalk et al., 2011). Siderophores are not only contributing in plant and microorganism nutrition but also in other environmental applications. In this review, microbial and plant siderophores will be discussed with a primary focus on the roles, functions and applications of siderophores in different areas of environmental research.
Microbial siderophores
Microorganisms produce a wide range of siderophores (Fig. 1). Most of the bacterial siderophores are catecholates (i.e. enterobactin), and some are carboxylates (i.e. rhizobactin) and hydroxamates (i.e. ferrioxamine B) (Matzanke, 1991). However, there are also certain types of bacterial siderophores that contain a mix of the main functional groups (i.e. pyoverdine) (Cornelis, 2010). One of the most common fungal siderophores is hydroxamates belonging to the ferrichrome family (i.e. ferrichrome), which is further divided into five groups, depending on the side chain of the hydroxamate functional group (Renshaw et al., 2002; Winkelmann, 2007).
Figure 1.
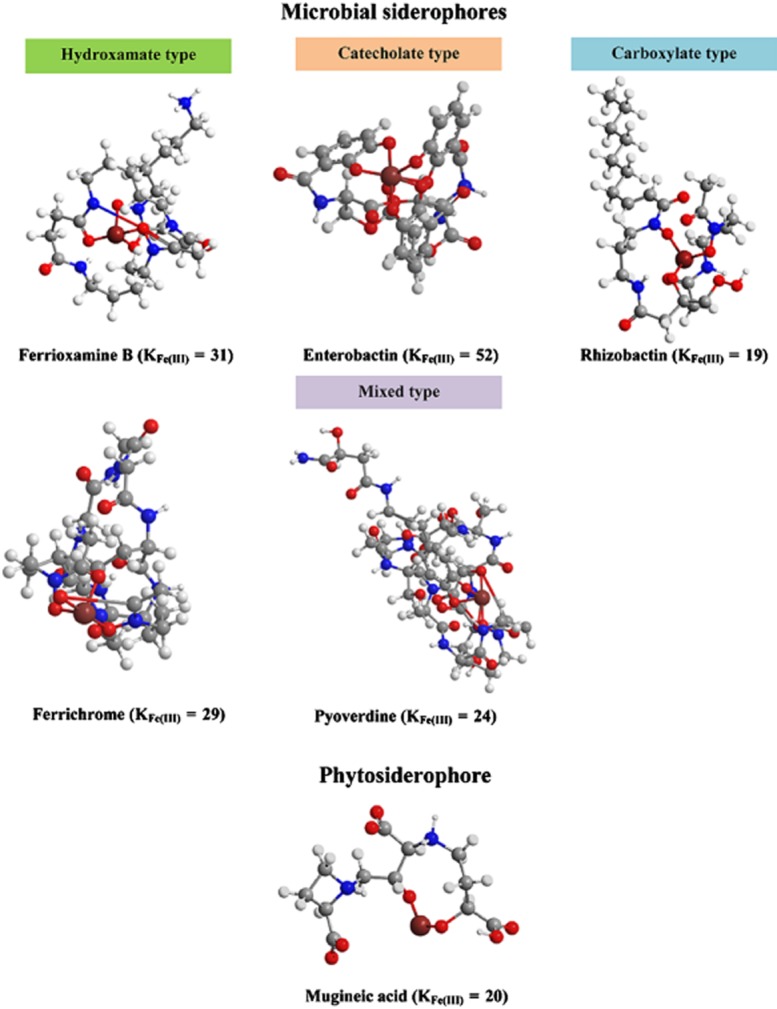
Representative examples of microbial siderophores and phytosiderophores with the stability constant of each type. Microbial siderophores consist of four main types. Hydroxamates produced by both bacteria (ferrioxamine B) and fungi (ferrichrome). Catecholate (enterobactin), carboxylate (rhizobactin) and mixed type (pyoverdine) produced by bacteria. The most common phytosiderophore is mugineic acid. All the chemical structures were drawn using ChemDraw Standard 13.0 software (PerkinElmer, Waltham, MA, USA).
Microorganisms use different siderophore-mediated Fe transport systems. For bacteria, the transport systems vary between gram-positive bacteria and gram-negative bacteria. Gram-negative bacteria (e.g. Escherichia sp.) have TonB-dependent outer membrane receptors that recognize the Fe(III)–siderophore complexes at the cell surface (Krewulak and Vogel, 2008). Once the Fe(III)–siderophore binds to the outer membrane receptor, the complex crosses the membrane through an energy-dependent system consisting of the outer membrane receptor proteins, periplasmic binding proteins and inner membrane transport proteins (Matzanke, 1991). Thereafter, the Fe(III)–siderophore complex that is bound by a high-affinity periplasmic binding protein, which accompanied the complex to the cytoplasmic membrane, is released into the periplasmic space (Noinaj et al., 2010). Then the complex is transported across the cytoplasmic membrane by an ATP-binding cassette (ABC) transport system, allowing it to reach the cytoplasm. Eventually, Fe(II) is released from the Fe(III)–siderophore complex via reduction of Fe(III). In contrast, in gram-positive bacteria (e.g. Bacillus sp.), which lack the outer membrane, the outer membrane receptors are completely absent. Therefore, the Fe(III)–siderophore complexes are bound by periplasmic siderophore binding proteins that are anchored to the cell membrane because of the lack of a periplasmic space (Fukushima et al., 2013). Then the Fe(III)–siderophore complexes are further transported by ATP-dependent transporters to the cytoplasm according to the ABC transport systems in the same way as the gram-negative bacteria (Braun and Hantke, 2011).
Fungi have four different mechanisms for siderophore-mediated Fe transport systems (Van der Helm and Winkelmann, 1994): (i) In the shuttle mechanism, the Fe(III)–siderophore complex is transported across the cell membrane, where the Fe(III) is released from the ligand and reduced by the reductive enzymes, whereas the free siderophore is then recycled. This mechanism is, for example, used for transporting ferrichrome in some fungal species such as Ustilago maydis (Ardon et al., 1998). (ii) In the taxicab mechanism, the Fe(III) from the extracellular siderophore is transferred across the cell membrane to intracellular ligands. This mechanism is used by Rhodotorula species (Winkelmann and Huschka, 1987). (iii) In the hydrolytic mechanism, the whole Fe(III)–siderophore complex is transported into the cell and is subjected to several reductive and degradative processes to release the Fe(III). The Fe(III) is reduced to Fe(II) inside the cell and the siderophore is excreted again. This mechanism is used in the uptake of Fe(III)–triacetylfusarinine complexes by Mycelia sterilia (Adjimani and Emery, 1988). (iv) In the reductive mechanism, the Fe(III)–siderophore complex is not transported across the cell membrane. The reduction of Fe(III) to Fe(II) occurs at the cell membrane and then the reduced Fe is taken up by the cell. This mechanism is used by Ustilago sphaerogena in the uptake of Fe(III) from ferrichrome (Ecker and Emery, 1983).
Plant siderophores
Fe is an essential micronutrient for plant growth (Kobayashi and Nishizawa, 2012). Under conditions of Fe deficiency, graminaceous plants (e.g. barley and wheat) have developed an efficient strategy for acquiring Fe from insoluble sources (Kraemer et al., 2006). These plants secrete Fe(III)-chelating compounds called phytosiderophores that form specific strong complexes with Fe(III) (Ma, 2005). The phytosiderophores are hexadentate ligands that coordinate Fe(III) with their amino and carboxyl groups (Singh et al., 2011). When the phytosiderophore is released to the rhizosphere, it chelates Fe from the soil by forming Fe(III)–phytosiderophore complexes that can be subsequently transported across the root plasma membrane (Römheld and Marschner, 1986; Dell'mour et al., 2012). In comparison with the molecular mass of microbial siderophores, which is ranged between 200 and 2000 Da, phytosiderophores are ranged between 500 and 1000 Da (Neilands, 1981).
Mugineic acid (MA) is the most common and the first identified phytosiderophore (Takemoto et al., 1978) (Fig. 1). The stability constant of the MA-Fe(III) complex is K = 1020 (Raymond et al., 1984), which is low compared with the stability constant of microbial siderophores such as ferrichrome (K = 1029), ferrioxamine B (K = 1031) and enterobactin (K = 1052) (Schwarzenbach and Schwarzenbach, 1963; Harris et al., 1979). Other types of phytosiderophores have also been isolated from gramineous plants such as avenic acid A from oats (Avena sativa) and distichonic acid from beer barley (Hordeum vulgate) (Nomoto et al., 1981). In general, several studies showed that plant species such as barley, rye and wheat, which produce a high concentration of phytosiderophores, are more resistant to Fe deficiency than other species such as maize, sorghum and rice, which produce a lower concentration of phytosiderophores (e.g. Masuda et al., 2009; Kobayashi et al., 2010).
Role of siderophores in nature
Soil mineral weathering
In soils, the microbial communities that colonize mineral surfaces differ from the inhabitants of the surrounding soil (Certini et al., 2004). Microbial attachment to mineral surfaces leads to the formation of a microenvironment that protects the microorganisms against environmental stresses (Liermann et al., 2000; Ojeda et al., 2006). In these microenvironments, mineral nutrients can be chelated directly from the soil minerals or shared amongst the surrounding microorganisms (Roberts Rogers and Bennett, 2004). Siderophores produced by soil microorganisms can promote the mineral dissolution of insoluble phases (Kraemer, 2004; Shirvani and Nourbakhsh, 2010). There are different mechanisms suggested for siderophore-promoted Fe dissolution (see Holmén and Casey, 1996). The general mechanism is that the Fe(III)–siderophore complex is formed at the mineral surface and is then transferred into the surrounding soil solution and becomes available for uptake by microorganisms or plants (Kalinowski et al., 2000; Kraemer, 2004).
The impact of siderophores on soil mineral weathering can be more effective compared with that of low molecular mass organic acids (LMMOAs) because siderophores form more stable complexes with Fe. Siderophores form 1:1 complexes with Fe(III), with constants ranging between K = 1030 and K = 1052 (Matzanke, 1991), while the constants of oxalic and citric acids with Fe(III) are K = 108 and 1012 respectively (Perrin, 1979). However, when both LMMOAs and siderophores are present, they may function synergistically, which results in a higher mineral dissolution rate compared with siderophore only (Reichard et al., 2007).
Because of the importance of microbial siderophores in weathering and soil formation, the role of siderophores in the dissolution of Fe minerals has been investigated intensively. For instance, Watteau and Berthelin (1994) reported that hydroxamate siderophores produced by Suillus granulatus have a high efficiency in the dissolution of goethite. Significant quantities (10−9 mol m−2 h−1) of Fe were mobilized in the presence of Suillus sp. because of its continuous production of siderophores. In addition, fungal siderophores such as ferrichrome and ferricrocin belonging to the ferrichrome family contributed in changing the surface structure of biotite and increasing its dissolution in podzolic forest soil (Sokolova et al., 2010). The dissolution of Fe from hornblende has also been observed to be higher in the presence of siderophore-producing actinobacteria (Streptomyces and Arthrobacter) compared with the dissolution of Fe by a synthetic siderophore (desferrioxamine B) (Kalinowski et al., 2000).
Some studies have also shown that phytosiderophores can play a significant role in mineral-weathering processes by accelerating the dissolution of some Fe-containing minerals such as ferrihydrate and goethite (Hiradate and Inoue, 2010). Reichard and colleagues (2005) reported that the maximum dissolution rate of goethite (1.7 nmol m−2 h−1) in the presence of phytosiderophores (2′-deoxymugineic acid) was found at pH 6. Recently, genetic engineering applications have allowed us to observe that the expression of Fe–phytosiderophore transporter genes in barley enhances its ability to dissolve Fe from soil minerals (Gómez-Galera et al., 2012).
Biogeochemical cycling of Fe in the ocean
The biogeochemical cycling of trace metals in the oceans has become a subject of great concern. Of all trace metals present in the marine waters, Fe have received the most attention because it is an essential micronutrient for marine organisms, in addition to its low concentration in the ocean controls the productivity and community structure of phytoplankton (Gledhill and Buck, 2012). Marine bacteria produce most of the organic Fe chelators present in seawater and thereby play a significant role in the biogeochemical cycling of Fe in the ocean (Boyd et al., 2007). Those bacteria compete with phytoplankton for Fe by producing different types of siderophores that have a great impact on the Fe abundance and solubility in the marine environment (Cordero et al., 2012). Marine siderophores include a hydroxyl–carboxylate functional group, provided either by citrate (i.e. snychobactins, petrobactin, aerobactin and marinobactins) or by b-hydroxyaspartate (i.e. aquachelins, loihichelins and alterobactin) (Martinez and Butler, 2007; Hider and Kong, 2010).
In the surface water, siderophores participate in the photochemical cycling of Fe by forming Fe(III)–siderophore complexes that increase Fe availability for phytoplankton (Barbeau et al., 2001; Hunter and Boyd, 2007). Ferrioxamine G was found to be widely distributed in surface waters throughout the Atlantic Ocean, whereas ferrioxamine E had a more varied distribution within depths. These findings suggest that marine siderophores are important in enhancing the Fe abundance and availability in the water column of the Atlantic Ocean, thus playing an important role in the biogeochemical cycle of Fe (Mawji et al., 2008; Amin et al., 2012).
Biotechnological applications of siderophores
Enhancing growth and pathogen biocontrol of plants
It is known that microbial siderophores provide plants with Fe nutrition to enhance their growth when the bioavailability of Fe is low (Crowley, 2006), whereas the exact mechanism is fairly unknown. Two possible mechanisms were suggested by which plants could obtain Fe from microbial siderophores: (i) Microbial siderophores with high redox potential can be reduced to donate Fe(II) to the transport system of the plant. In this mechanism, it has been hypothesized that the microbial Fe(III)–siderophores are transported to the apoplast of the plant root where siderophore reduction may be occur (Mengel, 1995). Consequently, Fe(II) is trapped in the apoplast, which leads to high Fe concentrations in the root (Kosegarten et al., 1999). (ii) Microbial siderophores can chelate Fe from soils and then do a ligand exchange with phytosiderophores (Masalha et al., 2000). This mechanism depends on several parameters, i.e. the stability constants and concentrations of both microbial and phytosiderophores, and the pH and redox conditions of the root environment (Crowley, 2006).
Siderophores have been suggested to be an environmentally friendly alternative to hazardous pesticides (Schenk et al., 2012). It has been known for more than three decades that different Pseudomonas species can improve plant growth by producing siderophores (pyoverdines) and/or by protecting them from pathogens, and thus this group of bacteria was classified as plant growth-promoting bacteria (Kloepper et al., 1980; Gamalero and Glick, 2011). In addition to pseudomonads, other bacteria such as Azadirachta indica which produce ferrioxamines could contribute into plant Fe nutrition and promote the root and shoot growth (Siebner-Freibach et al., 2003; Verma et al., 2011). Mycorrhizal fungi can also be used as biofertilizers to enhance plant growth. Mycorrhizal sorghum plants were shown to take up higher concentrations of Fe than nonmycorrhizal plants (Caris et al., 1998). It is suggested that the ectomycorrhizal fungi associations in plant nutrition depend on fungal siderophores (Van Schöll et al., 2008). Recently, the plant growth-promoting activities of fungi were investigated, and siderophores produced by Aspergillus niger, Penicillium citrinum and Trichoderma harzianum were found to increase the shoot and root lengths of chickpeas (Cicer arietinum) (Yadav et al., 2011).
Furthermore, the significant role of siderophores in the biological control mechanism has also been demonstrated by Kloepper and colleagues (1980). This mechanism depends on the role of siderophores as competitors for Fe in order to reduce the Fe availability for the phytopathogens (Beneduzi et al., 2012). There are several studies regarding the role of siderophores in the biological control of plant pathogens. Pyoverdine siderophores produced by pseudomonads were found to control the wilt diseases of potato caused by Fusarium oxysporum (Schippers et al., 1987), in addition to being involved in the biocontrol of Gaeumannomyces graminis, which causes a deficiency of wheat and barley growth (Voisard et al., 1989). Furthermore, pyoverdines were also observed to suppress the phytopathogens in peanuts and maize (Pal et al., 2001). There are other bacterial species besides pseudomonads that can be used as biocontrol agents. For example, siderophores produced by Bacillus subtilis had a significant role in the biocontrol of F. oxysporum, which causes the Fusarium wilt of pepper (Yu et al., 2011). Also, siderophores produced by A. indica had a high affinity to chelate Fe(III) from soil and thereby negatively affect the growth of several fungal pathogens (Verma et al., 2011).
Biocontrol of fish pathogens
Siderophores play an important role in disease control of fish by limiting Fe that is necessary in virulence and bacterial interactions (Li and Chi, 2004). The pathogenic bacterium use two different ways in infecting the fish host: (i) by production of harmful enzymes such as proteases and cholesterol acyl transferase to resist the defense mechanisms of the host (Ellis, 1999) and (ii) by production of transferrin in order to compete with the host for Fe and suppress its growth (Yano, 1996). The biocontrol mechanism mainly depend on the competition between the transferrin produced by pathogens and the siderophore produced by biocontrol agents in forming complexes with Fe, and the siderophore is always the winner because of its much higher Fe stability constants (Gram et al., 1999). Siderophore-producing bacteria (Pseudomonas fluorescens) can inhibit the growth of several fish pathogens (Vibrio anguillarum, Vibrio ordalii, Aeromonas salmonicida, Lactococcus garvieae, Streptococcus iniae, Flavobacterium psychrophilum and c ruckeri) and is therefore used as probiotics in fish farming (Gram et al., 2001; Brunt et al., 2007; Dimitroglou et al., 2011). Furthermore, Bacillus sp. has also been recommended to be used as a biocontrol agent in fish intestines and culture water (Sugita et al., 1998). For instance, Bacillus sp. strain NM 12 produces siderophores with a wide antibacterial spectrum that inhibited the growth of 62.5% of 363 intestinal bacteria identified from several coastal fishes (Li and Chi, 2004). In addition, siderophores produced by Bacillus cereus has also been found to inhibit the growth of the fish pathogen Aeromonas hydrophila (Lalloo et al., 2010). Recently it was found that several siderophore-producing bacteria isolated from the intestinal tracts of fishes inhibit fish pathogens such as Aliivibrio logei, Vibrio ichthyoenteri, V. anguillarum, Vibrio splendidus and A. salmonicida (Sugita et al., 2012).
Microbial ecology and taxonomy
Siderotyping is defined as the characterization of microbial strains according to the siderophore types they produce (Neilands, 1981; Bosne and Levy Frebault, 1992). There are two different methods for siderotyping, the analytical and the biological methods (Meyer et al., 2002). The analytical methods using high-performance liquid chromatography (HPLC) coupled with mass spectrometry are based on the physicochemical properties of siderophores. However, the biological methods are based on the direct measurement of siderophore-mediated Fe in the microbial cells or using a molecular biology method based on the recognition of specific DNA sequences related to siderophores (Bach et al., 2000).
Pseudomonas, produce mainly over 50 different pyoverdine siderophores (Boukhalfa and Crumbliss, 2002), in addition to a wide variety of other siderophore types (Cornelis, 2010). The peptide chain of the fluorescent pyoverdine varies among the different species and this variability can be easily used to determine the relatedness of these species (Chincholkar et al., 2005). Siderotyping application has been previously investigated on 400 strains of fluorescent and non-fluorescent Pseudomonas spp. and these strains were grouped into 28 taxa including 15 well-defined species depending on the different siderophore types (Meyer et al., 2002). Sixty-eight fluorescent Pseudomonas strains were also identified using mass spectrometry analysis of their pyoverdines, and thus siderotyping was recommended as a helpful method for studying microbial diversity and taxonomy (Meyer et al., 2008; Meyer, 2010). Other studies have found that siderophore production can be used as a chemotaxonomic marker for the identification of other types of bacteria such as Burkholderia spp. and Mycobacterium spp. based on the variation of the chemical structures of ornibactins and mycobactin respectively (Mokracka et al., 2004; Bultreys et al., 2006). Those findings suggest that siderotyping could become a powerful tool in environmental research because it can provide a quick and unambiguous identification of microbes at the species level (Meyer et al., 2002).
Bioremediation of environmental pollutants
Metals
Metals play a vital role in the development of human civilizations (Jonhson et al., 2002), but the manufacturing industry, sludge applications, nuclear power stations and mining have led to metal pollution (Wasi et al., 2013). Siderophores are extremely effective in solubilizing and increasing the mobility of a wide range of metals such as Cd, Cu, Ni, Pb, Zn, and the actinides Th(IV), U(IV) and Pu(IV) (Schalk et al., 2011). This ability of siderophores mainly depends on their ligand functionalities, by which means siderophores may have a strong affinity or selectivity for a particular metal other than Fe with regards to the stability constants of this metal–siderophore complex (Hernlem et al., 1999). Thereby, siderophores become a useful tool in bioremediation, which is a cost-effective and environmentally friendly technique (Rajkumar et al., 2010).
During the last years, there had been an increasing interest in investigating the potential of using siderophores in metal bioremediation. Neubauer and colleagues (2000) showed that siderophores such as desferrioxamine B could chelate Co(III) better than Fe(III) in high pH conditions. Azotobacter vinelandii, which produced siderophores, i.e. azotochelin and azotobactin, had the ability to use those siderophores for both Mo and V acquisition (Wichard et al., 2009). Moreover, the siderophore pyochelin produced by Pseudomonas aeruginosa could chelate a wide range of metals, i.e. Ag+, Al3+, Cd2+, Co2+, Cr2+, Cu2+, Eu3+, Ga3+, Hg2+, Mn2+, Ni2+, Pb2+, Sn2+, Tb3+, Tl+ and Zn2+; however, the uptake process did not appear to assimilate any metal other than Fe3+ (Braud et al., 2009a). Hong and colleagues (1996) reported that siderophores produced by Fusarium solani contributed in solubilizing Cu and Zn in vitro. Siderophores also played a significant role in mobilizing metals from mine waste material or metal-contaminated soils. Several metals (i.e. Fe, Ni and Co) were mobilized from waste material (acid-leached ore) of a former uranium mine in the presence of siderophores produced by P. fluorescens (Edberg et al., 2010). It has been shown that the siderophores produced by Agrobacterium radiobacter removed approximately 54% of the As from a metal-contaminated soil (Wang et al., 2011). Recently, it has been found that pyoverdines mobilized U(VI), Np(V) and other metals from uranium mine waste (Behrends et al., 2012).
Not only microbial siderophores contribute to metal bioremediation, but also there are many studies that have demonstrated that phytosiderophores are efficient in mobilizing metals in soil (e.g. Rajkumar et al., 2009; 2010; 2012). Phytosiderophores have a high affinity for complexation with several metals in the following order (Cd2+ > Ni2+ > Pb2+ > Sn2+ > AsO4−2 > AsO2−1 > Mn2+ > Co2+ > Cu2+ > Fe+3) and very weakly binds Al3+ and Cr3+ (Ruggiero et al., 1999). Studies in both uncontaminated and contaminated soils have shown that phytosiderophores are more efficient in mobilizing Fe, Cu, Zn, Ni and Cd from the soil in comparison with synthetic chelators and microbial siderophores (i.e. Awad and Römheld, 2000; Singh et al., 2008).
Petroleum hydrocarbons
Petroleum hydrocarbons in marine ecosystems are one of the major environmental problems. Microorganisms could play an important role in the remediation of petroleum hydrocarbons from the marine environment (Das and Chandran, 2011). Microbial siderophores participate in the biodegradation of petroleum hydrocarbons through an indirect mechanism, by facilitating the Fe acquisition for the degraded microorganisms under Fe-limiting conditions (Barbeau et al., 2002). Petrobactin was the first structurally characterized siderophore produced by the oil-degrading marine bacterium Marinobacter hydrocarbonoclasticus (Barbeau et al., 2002). Hickford and colleagues (2004) identified another sulfonated siderophore called ‘Petrobactin sulfonate’ isolated from the same oil-degrading marine bacterium. Few studies suggested that the use of siderophores may be a good strategy for oil spill clean-up. Gauglitz and colleagues (2012) showed recently that marine Vibrio spp. isolated from the Gulf of Mexico after the 2010 Deepwater Horizon oil spill, produce amphiphilic siderophores called ‘ochrobactins’ that were suggested to efficiently contribute in the degradation of petroleum hydrocarbons.
Nuclear fuel reprocessing
Siderophores contain anionic hydroxamate or catecholate functional groups that form hard oxodonors that strongly bind to Lewis acids, resulting in complexes with remarkably high stability constants (Harris et al., 1979). Because actinides form strong complexes with hard oxygen anions, it has been suggested that siderophores could bind actinides with a complexation constant estimated to be K = 1016 (Raymond, 1990; Jarvis and Hancock, 1991).
The Purex process has been used commercially to reprocess irradiated nuclear fuel by solvent extraction techniques and separate U and Pu for reuse from fission products such as Ti and Np (Taylor and May, 1999). During this process, U and Pu flow into the solvent and become contaminated with Np. Siderophores have been shown to allow for the selective removal of Np from the solvent phase (Taylor et al., 1998), and thus siderophores could be used in the Purex process to simplify the removing of the actinides (Renshaw et al., 2002). Desferrioxamine B forms a stable complex with U(VI), where its hydroxamate functional group is similar to acetohydroxamic acid, a ligand that has been proposed for actinide complexation (Mullen et al., 2007). Recently, Marshall and colleagues (2010) reported that low concentration of siderophores was enough to potentially influence the dissolution of spent nuclear fuel, and it seemed not to be a significant difference between using the synthetic desferrioxamine B or pyoverdine produced by P. fluorescens. Based on those studies, siderophores have been proposed for the remediation of radioactive waste and reprocessing of nuclear fuel.
Optical biosensor
A biosensor is a biomolecule coupled to an electrical device such as a transducer, amplifier or noise filter in order to increase the signal to noise ratio that allows detection of various types of responses through specifically engineered systems (Gupta et al., 2008). Pyoverdines are yellow-green water-soluble fluorescent siderophores characterized by the following properties (Barrero et al., 1993): (i) They form a strong complex with Fe(III) and have a weak or negligible affinity for Fe(II), and (ii) the Fe(III) complexes have very high stability constants (approximately K = 1032) (Kurtz and Crouch, 1991). These characteristics make pyoverdine a promising agent for the construction of optical biosensors (Pesce and Kaplan, 1990). Using the siderophore with an exceptional Fe(III)-binding constant would be an ideal choice for the molecular recognition element of the sensor that could be applied in the determination of Fe bioavailability in oceanic water or soils (Chung Chun Lam et al., 2006). The concentration of Fe present in the ocean has been determined by using a siderophore as a biosensor (Chung Chun Lam et al., 2006). In that study, they used parabactin that was produced by Paracoccus denitrificans as a biosensor by encapsulating it in sol-gel thin film on a quartz substrate. The seawater samples were analysed by a flow cell that was mounted in the sample partition of the fluorescence spectrometer. Siderophores also provide the potential for a good sensitive and selective detection system that would mimic the biological uptake process (Ellerby et al., 1992). For instance, azotobactin produced by A. vinelandii has been used as an optical biosensor for Fe(III) in a modified design that depends on the encapsulation of the azotobactin in sol-gel matrices without significant loss of its fluorescence signal (Sharma and Gohil, 2010). Additionally, the Fe(II and III) specificity for the Fe biosensor pyoverdine has been optimized by immobilizing it in three formulations of porous sol-gel glass (A, B and C), which contained various amounts of water added (Yoder and Kisaalita, 2011). In that study, the most specific and linear response for binding to Fe(II and III) was observed for pyoverdine immobilized in sol-gel C that had more water than A and B.
Bio-bleaching of pulps
The pulp and paper industry is a primary source of many environmental problems including global warming, human toxicity, ecotoxicity, photochemical oxidation, acidification, nitrification and solid wastes (Singh et al., 2008; Bajpai, 2010). The main problem of pulp and paper manufacturing results from the bleaching processes. Some pollutants are emitted into the air, while others are discharged in wastewater.
Siderophores are considered as effective agents in pulp treatment, where they can reduce 70% of the chemicals needed to bleach Kraft pulp (Bajpai, 2004), and that makes siderophores environment-friendly alternatives. Brown-rot fungi are considered to be one of the most important groups of wood-decaying microorganisms. Some studies reported the production of catecholate and hydroxamate siderophores by wood-decaying fungi (Fekete et al., 1989; Fekete, 1993). For instance, hydroxamate siderophores isolated from the brown-rot fungus (Gloephyllum trabeum) had the ability to mediate the reduction of Fe in redox cycling processes. The reduced Fe can then react with hydrogen peroxide to generate oxygen radical species that depolymerize cellulose, hemicellulose and lignocellulose. This depolymerization process was considered to be the main role of siderophores in the bio-bleaching of pulps (Xu and Goodell, 2001; Arantes and Milagres, 2007). Moreover, Milagres and colleagues (2002) showed the ability of siderophores produced by different microbial species in bio-bleaching of pulps with regard to the loss of viscosity and they found that siderophores produced by G. trabeum could degrade pulp with 10.8% loss of viscosity, whereas siderophores produced by Perenniporia medula-panis and Tinea versicolor degraded pulps with 13.6% and 14.4% loss respectively. It has also been observed that siderophores produced by Coriolus versicolor could alter the lignin structure to make it more susceptible to degradation and accordingly contribute in bio-bleaching of pulps (Wang et al., 2008).
Concluding remarks and future perspectives
The study of mineral–microbe interactions underscores the importance of microorganisms in making the earth a suitable environment for all forms of life. In recent years, it has become clear that siderophores represent central organic compounds in Fe uptake in many microorganisms and plants. Understanding the chemical structures of different siderophores and the membrane receptors involved in Fe uptake has opened new areas for research. The importance of siderophores is obvious, and they play a significant role in the environmental applications, even if there are many questions remaining to be answered. What is the specific role of microorganisms and plants in the selectivity of metal uptake by siderophores? Why do microorganisms secrete more than one type of siderophores to meet their mineral nutritional needs? What is the relative importance of the different siderophore structures involved in environmental applications? Can modified genetic methods such as labelled DNA be useful tools for the direct detection of siderophore functional genes in the environment?
More research is focusing on finding effective ways to use siderophores in bioremediation and biocontrol, which should enhance their application in the environment. Siderophore variability and their structural and functional characteristics in relation to microbial communities must be vigorously investigated to improve the role of siderophores in environmental applications. The relationship between siderophores and microbial structure, in environments with low Fe bioavailability, i.e. oceans and some soil conditions, are still not clearly known. Combining metagenomics with detailed chemical analysis will reveal important information that could be used to improve the current environmental applications and develop new applications for siderophores.
Acknowledgments
We would like to thank the reviewers for their valuable comments and suggestions. We thank Hildred Crill, Stockholm University, for her helpful suggestions in improving the writing of the manuscript.
Conflict of interest
None declared.
Funding Information
The present article was supported by grants from The Swedish Research Council for Environment, Agricultural Sciences and Spatial Planning (FORMAS), Sweden and the Faculty of Science, Stockholm University, Sweden.
References
- Adjimani JP, Emery T. Stereochemical aspects of iron transport in Mycelia sterilia EP-76. J Bacteriol. 1988;170:1377–1379. doi: 10.1128/jb.170.3.1377-1379.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aguado-Santacruz GAA, Moreno-Gómez BA, Jiménez-Francisco BB, García-Moya EB, Preciado-Ortiz RE. Impact of the microbial siderophores and phytosiderophores on the iron assimilation by plants: a synthesis. Rev Fitotec Mex. 2012;35:9–21. [Google Scholar]
- Albrecht-Gary AM, Crumbliss AL. Coordination chemistry of siderophores: thermodynamics and kinetics of iron chelation and release. Met Ions Biol Syst. 1998;35:239–327. [PubMed] [Google Scholar]
- Amin SA, Green DH, Al Waheeb D, Gärdes A, Carrano CJ. Iron transport in the genus Marinobacter. Biometals. 2012;25:135–147. doi: 10.1007/s10534-011-9491-9. [DOI] [PubMed] [Google Scholar]
- Arantes V, Milagres AMF. The effect of a catecholate chelator as a redox agent in Fenton-based reactions on degradation of lignin-model substrates and on COD removal from effluent of an ECF kraft pulp mill. J Hazard Mater. 2007;141:273–279. doi: 10.1016/j.jhazmat.2006.06.134. [DOI] [PubMed] [Google Scholar]
- Ardon O, Nudelman R, Caris C, Libman J, Shanzer A, Chen Y, Hadar Y. Iron uptake in Ustilago maydis: tracking the iron path. J Bacteriol. 1998;180:2021–2026. doi: 10.1128/jb.180.8.2021-2026.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Armstrong JE, Van Baalen C. Iron transport in microalgae: the isolation and biological activity of a hydroxamate siderophore from the blue-green alga Agmenellum quadruplieatum. J Gen Microbiol. 1979;111:253–262. [Google Scholar]
- Awad F, Römheld V. Mobilization of heavy metals from contaminated calcareous soils by plant born, microbial and synthetic chelators and their uptake by wheat plants. J Plant Nutr. 2000;23:1847–1855. [Google Scholar]
- Bach S, Almeida de A, Carniel E. The Yersinia high-pathogenicity island is present in different members of the family Enterobacteriaceae. FEMS Microbiol Lett. 2000;183:289–294. doi: 10.1111/j.1574-6968.2000.tb08973.x. [DOI] [PubMed] [Google Scholar]
- Bajpai P. Biological bleaching of chemical pulps. Crit Rev Biotechnol. 2004;24:1–58. doi: 10.1080/07388550490465817. [DOI] [PubMed] [Google Scholar]
- Bajpai P. Environmentally Friendly Production of Pulp and Paper. New Jersey, USA: John Wiley & Sons; 2010. [Google Scholar]
- Barbeau K, Rue EL, Bruland KW, Butler A. Photochemical cycling of iron in the surface ocean mediated by microbial iron (II)-binding ligands. Nature. 2001;413:409–413. doi: 10.1038/35096545. [DOI] [PubMed] [Google Scholar]
- Barbeau K, Zhang GP, Live DH, Butler A. Petrobactin, a photoreactive siderophore produced by the oil-degrading marine bacterium Marinobacter hydrocarbonoclasticus. J Am Chem Soc. 2002;124:378–379. doi: 10.1021/ja0119088. [DOI] [PubMed] [Google Scholar]
- Barrero JM, Moreno-Bondi MC, Pérez-Conde MC, Cámara C. A biosensor for ferric ion. Talanta. 1993;40:1619–1623. doi: 10.1016/0039-9140(93)80075-3. [DOI] [PubMed] [Google Scholar]
- Behrends T, Krawczyk-Bärsch E, Arnold T. Implementation of microbial processes in the performance assessment of spent nuclear fuel repositories. Appl Geochem. 2012;27:453–462. [Google Scholar]
- Bellenger JP, Wichard T, Kustka AB, Kraepiel AML. Uptake of molybdenum and vanadium by a nitrogen-fixing soil bacterium using siderophores. Nat Geosci. 2008;1:243–246. [Google Scholar]
- Beneduzi A, Ambrosini A, Passaglia LM. Plant growth-promoting rhizobacteria (PGPR): their potential as antagonists and biocontrol agents. Genet Mol Biol. 2012;35:1044–1051. doi: 10.1590/s1415-47572012000600020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bosne S, Levy Frebault V. Mycobactin analysis as an aid for the identification of Mycobacterium fortuitum and Mycobacterium chelonae subspecies. J Clin Microbiol. 1992;30:1225–1231. doi: 10.1128/jcm.30.5.1225-1231.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boukhalfa H, Crumbliss AL. Chemical aspects of siderophores mediated iron transport. Biometals. 2002;15:325–339. doi: 10.1023/a:1020218608266. [DOI] [PubMed] [Google Scholar]
- Boukhalfa H, Lack J, Reilly SD, Hersman L, Neu MP. Siderophore production and facilitated uptake of iron and plutonium in P. putida. AIP Conf Proc. 2003;673:343–344. [Google Scholar]
- Boyd PW, Jickells T, Law CS, Blain S, Boyle EA, Buesseler KO, et al. Mesoscale iron enrichment experiments 1993–2005: synthesis and future directions. Science. 2007;315:612–617. doi: 10.1126/science.1131669. [DOI] [PubMed] [Google Scholar]
- Braud A, Jézéquel K, Bazot S, Lebeau T. Enhanced phytoextraction of an agricultural Cr-and Pb-contaminated soil by bioaugmentation with siderophore-producing bacteria. Chemosphere. 2009a;74:280–286. doi: 10.1016/j.chemosphere.2008.09.013. [DOI] [PubMed] [Google Scholar]
- Braud A, Hoegy F, Jezequel K, Lebeau T, Schalk IJ. New insights into the metal specificity of the Pseudomonas aeruginosa pyoverdine–iron uptake pathway. Environ Microbiol. 2009b;11:1079–1091. doi: 10.1111/j.1462-2920.2008.01838.x. [DOI] [PubMed] [Google Scholar]
- Braun V, Hantke K. Recent insights into iron import by bacteria. Curr Opin Chem Biol. 2011;15:328–334. doi: 10.1016/j.cbpa.2011.01.005. [DOI] [PubMed] [Google Scholar]
- Brunt J, Newaj-Fyzul A, Austin B. The development of probiotics for the control of multiple bacterial diseases of rainbow trout, Oncorhynchus mykiss (Walbaum) J Fish Dis. 2007;30:573–579. doi: 10.1111/j.1365-2761.2007.00836.x. [DOI] [PubMed] [Google Scholar]
- Bultreys A, Gheysen I, Hoffmann de E. Yersiniabactin production by Pseudomonas syringae and Escherichia coli, and description of a second yersiniabactin locus evolutionary group. Appl Environ Microbiol. 2006;72:3814–3825. doi: 10.1128/AEM.00119-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caris C, Hördt W, Hawkins HJ, Römheld V, George E. Studies of iron transport by arbuscular mycorrhizal hyphae from soil to peanut and sorghum plants. Mycorrhiza. 1998;8:35–39. [Google Scholar]
- Certini G, Campbell CD, Edwards AC. Rock fragments in soil support a different microbial community from the fine earth. Soil Biol Biochem. 2004;36:1119–1128. [Google Scholar]
- Chhibber S, Nag D, Bansal S. Inhibiting biofilm formation by Klebsiella pneumoniae B5055 using an iron antagonizing molecule and a bacteriophage. BMC Microbiol. 2013;13:174–183. doi: 10.1186/1471-2180-13-174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chincholkar SB, Rane MR, Chaudhari BL. Siderophores: their biotechnological applications. In: Varma A, Podila GK, editors. Biotechnological Application of Microbes. New Delhi, India: I.K. International; 2005. pp. 177–198. [Google Scholar]
- Chung Chun Lam CKS, Jickells TD, Richardson DJ, Russell DA. Fluorescence-based siderophore biosensor for the determination of bioavailable iron in oceanic waters. Anal Chem. 2006;78:5040–5045. doi: 10.1021/ac060223t. [DOI] [PubMed] [Google Scholar]
- Cordero OX, Ventouras LA, DeLong EF, Polz MF. Public good dynamics drive evolution of iron acquisition strategies in natural bacterioplankton populations. Proc Natl Acad Sci USA. 2012;109:20059–20064. doi: 10.1073/pnas.1213344109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cornelis P. Iron uptake and metabolism in pseudomonads. Appl Microbiol Biotechnol. 2010;86:1637–1645. doi: 10.1007/s00253-010-2550-2. [DOI] [PubMed] [Google Scholar]
- Crowley DA. Microbial siderophores in the plant rhizosphere. In: Barton LL, Abadía J, editors. Iron Nutrition in Plants and Rhizospheric Microorganisms. Netherlands: Springer; 2006. pp. 169–189. [Google Scholar]
- Dahlheimer SR, Neal CR, Fein JB. Potential mobilization of platinum group elements by siderophores in surface environments. Environ Sci Technol. 2007;41:870–875. doi: 10.1021/es0614666. [DOI] [PubMed] [Google Scholar]
- Das N, Chandran P. Microbial degradation of petroleum hydrocarbon contaminants: an overview. Biotechnol Res Int. 2011;11:1–13. doi: 10.4061/2011/941810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dell'mour M, Schenkeveld W, Oburger E, Fischer L, Kraemer S, Puschenreiter M, et al. Analysis of iron-phytosiderophore complexes in soil related samples: LC-ESI-MS/MS versus CE-MS. Electrophoresis. 2012;33:726–733. doi: 10.1002/elps.201100466. [DOI] [PubMed] [Google Scholar]
- Dimitroglou A, Merrifield DL, Carnevali O, Picchietti S, Avella M, Daniels C, et al. Microbial manipulations to improve fish health and production – a Mediterranean perspective. Fish Shellfish Immunol. 2011;30:1–16. doi: 10.1016/j.fsi.2010.08.009. [DOI] [PubMed] [Google Scholar]
- Ecker DJ, Emery T. Iron uptake from ferrichrome A and iron citrate in Ustilago sphaerogena. J Bacteriol. 1983;155:616–622. doi: 10.1128/jb.155.2.616-622.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Edberg F, Kalinowski BE, Holmström SJM, Holm K. Mobilization of metals from uranium mine waste: the role of pyoverdines produced by Pseudomonas fluorescens. Geobiology. 2010;8:278–292. doi: 10.1111/j.1472-4669.2010.00241.x. [DOI] [PubMed] [Google Scholar]
- Ellerby LM, Nishida CR, Nishida F, Yamanaka SA, Dunn B, Valentine JS, Zink JI. Encapsulation of proteins in transparent porous silicate-glasses prepared by the sol-gel method. Science. 1992;255:1113–1115. doi: 10.1126/science.1312257. [DOI] [PubMed] [Google Scholar]
- Ellis AE. Immunity to bacteria in fish. Fish Shellfish Immunol. 1999;9:291–308. [Google Scholar]
- Fekete FA. Assays for microbial siderophores. In: Barton LL, Hemming BC, editors. Iron Chelation in Plants and Soil Microorganisms. New York: Academic Press; 1993. pp. 399–417. [Google Scholar]
- Fekete FA, Chandhoke V, Jellison J. Iron-binding compounds produced by wood-decaying basidiomycetes. Appl Environ Microbiol. 1989;55:2720–2722. doi: 10.1128/aem.55.10.2720-2722.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fukushima T, Allred BE, Sia AK, Nichiporuk R, Andersen UN, Raymond KN. Gram-positive siderophore-shuttle with iron-exchange from Fe-siderophore to apo-siderophore by Bacillus cereus YxeB. Proc Natl Acad Sci USA. 2013;110:13821–13826. doi: 10.1073/pnas.1304235110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gamalero E, Glick BR. Mechanisms used by plant growth-promoting bacteria. In: Maheshwari DK, editor. Bacteria in Agrobiology: Plant Nutrient Management. Berlin, Heidelberg, Germany: Springer Verlag; 2011. pp. 17–46. [Google Scholar]
- Gauglitz JM, Zhou H, Butler A. A suite of citrate-derived siderophores from a marine Vibrio species isolated following the Deepwater Horizon oil spill. J Inorg Biochem. 2012;107:90–95. doi: 10.1016/j.jinorgbio.2011.10.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gledhill M, Buck KN. The organic complexation of iron in the marine environment: a review. Front Microbiol. 2012;3:1–17. doi: 10.3389/fmicb.2012.00069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gómez-Galera S, Sudhakar D, Pelacho AM, Capell T, Christou P. Constitutive expression of a barley Fe phytosiderophore transporter increases alkaline soil tolerance and results in iron partitioning between vegetative and storage tissues under stress. Plant Physiol Biochem. 2012;53:46–53. doi: 10.1016/j.plaphy.2012.01.009. [DOI] [PubMed] [Google Scholar]
- Gram L, Melchiorsen J, Spanggaard B, Huber I, Nielsen TF. Inhibition of Vibrio anguillarum by Pseudomonas ftuorescens AH2, a possible probiotic treatment of fish. Appl Environ Microbiol. 1999;65:969–973. doi: 10.1128/aem.65.3.969-973.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gram L, Lovold T, Nielsen J, Melchiorsen J, Spanggaard B. In vitro antagonism of the probiont Pseudomonas fluorescens strain AH2 against Aeromonas salmonicida does not confer protection of salmon against furunculosis. Aquaculture. 2001;199:1–11. [Google Scholar]
- Gupta V, Saharan K, Kumar L, Gupta R, Sahai V, Mittal A. Spectrophotometric ferric ion biosensor from Pseudomonas fluorescens culture. Biotechnol Bioeng. 2008;100:284–296. doi: 10.1002/bit.21754. [DOI] [PubMed] [Google Scholar]
- Harris WR, Carranao CJ, Cooper SR, Sofen SR, Avdeef AE, McArdle JV, Raymond KN. Coordination chemistry of microbial iron transport compounds. 19. Stability constants and electrochemical behavior of ferric enterobactin and model complexes. J Am Chem Soc. 1979;101:6097–6104. [Google Scholar]
- Hernlem BJ, Vane LM, Sayles GD. The application of siderophores for metal recovery and waste remediation: examination of correlations for prediction of metal affinities. Water Res. 1999;33:951–960. [Google Scholar]
- Hickford SJH, Küpper FC, Zhang G, Carrano CJ, Blunt JW, Butler A. Petrobactin sulfonate, a new siderophore produced by the marine bacterium Marinobacter hydrocarbonoclasticus. J Nat Prod. 2004;67:1897–1899. doi: 10.1021/np049823i. [DOI] [PubMed] [Google Scholar]
- Hider RC, Kong X. Chemistry and biology of siderophores. Nat Prod Rep. 27:637–657. doi: 10.1039/b906679a. [DOI] [PubMed] [Google Scholar]
- Hiradate S, Inoue K. Dissolution of iron from iron (hydr) oxides by mugineic acid. Soil Sci Plant Nutr. 1998;44:305–313. [Google Scholar]
- Holmén BA, Casey WH. Hydroxamate ligands, surface chemistry, and the mechanism of ligand-promoted dissolution of goethite [α-FeOOH(s)] Geochim Cosmochim Acta. 60:4403–4416. [Google Scholar]
- Hong JW, Park JY, Gadd GM. Pyrene degradation and copper and zinc uptake by Fusarium solani and Hypocrea lixii isolated from petrol station soil. J Appl Microbiol. 2010;108:2030–2040. doi: 10.1111/j.1365-2672.2009.04613.x. [DOI] [PubMed] [Google Scholar]
- Hunter KA, Boyd PW. Iron-binding ligands and their role in the ocean biogeochemistry of iron. Environ Chem. 2007;4:221–232. [Google Scholar]
- Jarvis N, Hancock R. Some correlations involving the stability of complexes of transuranium metal ions and ligands with negatively charged oxygen donors. Inorg Chim Acta. 1991;182:229–232. [Google Scholar]
- Jonhson GV, Lopez A, La Valle Foster N. Reduction and transport of Fe from siderophores. Plant Soil. 2002;241:27–33. [Google Scholar]
- Kalinowski BE, Liermann LJ, Brantley SL, Barnes AS, Pantano CG. X-ray photoelectron evidence for bacteria-enhanced dissolution of hornblende. Geochim Cosmochim Acta. 2000;64:1331–1343. [Google Scholar]
- Kloepper JW, Leong J, Teintze M, Schiroth MN. Enhanced plant growth by siderophores produced by plant growth promoting rhizobacteria. Nature. 1980;286:885–886. [Google Scholar]
- Kobayashi T, Nishizawa NK. Iron uptake, translocation, and regulation in higher plants. Annu Rev Plant Biol. 2012;63:131–152. doi: 10.1146/annurev-arplant-042811-105522. [DOI] [PubMed] [Google Scholar]
- Kobayashi T, Nakanishi H, Nishizawa NK. Recent insights into iron homeostasis and their application in graminaceous crops. Proc Jpn Acad Ser B Phys Biol Sci. 2010;86:900–913. doi: 10.2183/pjab.86.900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kosegarten H, Grolig F, Esch A, Glüsenkamp KH, Mengel K. Effects of NH4+, NO3− and HCO3− on apoplast pH in the outer cortex of root zones of maize, as measured by the fluorescence ratio of fluorescein boronic acid. Planta. 1999;209:444–452. doi: 10.1007/s004250050747. [DOI] [PubMed] [Google Scholar]
- Kraemer SM. Iron oxide dissolution and solubility in the presence of siderophores. Aquat Sci. 2004;66:3–18. [Google Scholar]
- Kraemer SM, Crowley DE, Kretzschmar R. Geochemical aspects of phytosiderophore promoted iron acquisition by plants. Adv Agron. 2006;91:1–46. [Google Scholar]
- Krewulak KD, Vogel HJ. Structural biology of bacterial iron uptake. Biochim Biophys Acta. 2008;1778:1781–1804. doi: 10.1016/j.bbamem.2007.07.026. [DOI] [PubMed] [Google Scholar]
- Kurtz KS, Crouch SR. Design and optimization of a flow-injection system for enzymatic determination of galactose. Anal Chim Acta. 1991;254:201–208. [Google Scholar]
- Lalloo R, Moonsamy G, Ramchuran S, Görgens J, Gardiner N. Competitive exclusion as a mode of action of a novel Bacillus cereus aquaculture biological agent. Lett Appl Microbiol. 2010;50:563–570. doi: 10.1111/j.1472-765X.2010.02829.x. [DOI] [PubMed] [Google Scholar]
- Li J, Chi Z. Siderophores from marine microorganisms and their applications. J Ocean Univ China. 2004;3:40–47. [Google Scholar]
- Liermann LJ, Kalinowski BE, Brantley SL, Ferry JG. Role of bacterial siderophores in dissolution of hornblende. Geochim Cosmochim Acta. 2000;64:587–602. [Google Scholar]
- Ma JF. Plant root responses to three abundant soil minerals: silicon, aluminum and iron. Crit Rev Plant Sci. 2005;24:267–281. [Google Scholar]
- Marshall MJ, Beliaev AS, Fredrickson JK. Microbiological transformations of radionuclides in the subsurface. In: Mitchell R, Gu J, editors. Environmental Microbiology. New Jersey, USA: Wiley-Blackwell; 2010. pp. 95–114. [Google Scholar]
- Martinez JS, Butler A. Marine amphiphilic siderophores: marinobactin structure, uptake, and microbial partitioning. J Inorg Biochem. 2007;101:1692–1698. doi: 10.1016/j.jinorgbio.2007.07.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Masalha J, Kosegarten H, Elmaci Ö, Mengel K. The central role of microbial activity for iron acquisition in maize and sunflower. Biol Fertil Soils. 2000;30:433–439. [Google Scholar]
- Masuda H, Usuda K, Kobayashi T, Ishimaru Y, Kakei Y, Takahashi M, et al. Overexpression of the barley nicotianamine synthase gene HvNAS1 increases iron and zinc concentrations in rice grains. Rice. 2009;2:155–166. [Google Scholar]
- Matzanke BF. Structures, coordination chemistry and functions of microbial iron chelates. In: Winkelmann G, editor. CRC Handbook of Microbial Iron Chelates. Boca Raton, FL, USA: CRC Press; 1991. pp. 15–64. [Google Scholar]
- Mawji E, Gledhill M, Milton JA, Tarran GA, Ussher S, Thompson A, et al. Hydroxamate siderophores: occurrence and importance in the Atlantic Ocean. Environ Sci Technol. 2008;42:8675–8680. doi: 10.1021/es801884r. [DOI] [PubMed] [Google Scholar]
- Mengel K. Iron availability in plant tissues – iron chlorosis on calcareous soils. In: Abadia J, editor. Iron Nutrition in Soils and Plants. Dordrecht, The Netherlands: Kluwer; 1995. pp. 389–397. [Google Scholar]
- Meyer JM. Pyoverdine siderophores as taxonomic and phylogenic markers Molecular Microbiology, Infection and Biodiversity. In: Ramos JL, Filloux A, editors. Pseudomonas. Vol. 6. London, UK: Springer; 2010. pp. 201–233. [Google Scholar]
- Meyer JM, Geoffroy VA, Baida N, Gardan L, Izard D, Lemanceau P, et al. Siderophore typing, a powerful tool for the identification of fluorescent and nonfluorescent Pseudomonas. Appl Environ Microbiol. 2002;68:2745–2753. doi: 10.1128/AEM.68.6.2745-2753.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meyer JM, Gruffaz C, Raharinosy V, Bezverbnaya I, Schäfer M, Budzikiewicz H. Siderotyping of fluorescent Pseudomonas: molecular mass determination by mass spectrometry as a powerful pyoverdine siderotyping method. Biometals. 2008;21:259–271. doi: 10.1007/s10534-007-9115-6. [DOI] [PubMed] [Google Scholar]
- Milagres AMF, Arantes V, Medeiros CL, Machuca A. Production of metal chelating compounds by white and brown-rot fungi and their comparative abilities for pulp bleaching. Enzyme Microb Technol. 2002;30:562–565. [Google Scholar]
- Mokracka J, Koczura R, Kaznowski A. Yersiniabactin and other siderophores produced by clinical isolates of Enterobacter spp. and Citrobacter spp. FEMS Immunol Med Microbiol. 2004;40:51–55. doi: 10.1016/S0928-8244(03)00276-1. [DOI] [PubMed] [Google Scholar]
- Mullen L, Gong C, Czerwinski K. Complexation of uranium (VI) with the siderophore desferroixamine B. J Radioanal Nucl Chem. 2007;273:683–688. [Google Scholar]
- Neilands JB. Iron absorption and transport in microorganisms. Annu Rev Nutr. 1981;1:27–46. doi: 10.1146/annurev.nu.01.070181.000331. [DOI] [PubMed] [Google Scholar]
- Neubauer U, Nowak B, Furrer G, Schulin R. Heavy metal sorption on clay minerals affected by the siderophore desferroixamine B. Environ Sci Technol. 2000;34:2749–2755. [Google Scholar]
- Noinaj N, Guillier M, Barnard TJ, Buchanan SK. TonB-dependent transporters: regulation, structure and function. Annu Rev Microbiol. 2010;64:43–60. doi: 10.1146/annurev.micro.112408.134247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nomoto K, Mino Y, Ishida T, Yoshioka H, Ota N, Inoue M, et al. X-ray crystal structure of the copper (II) complex of mugineic acid, a naturally occurring metal chelator of graminaceous plants. J Chem Soc, Chem Commun. 1981;7:338–339. [Google Scholar]
- Ojeda L, Keller G, Mühlenhoff U, Rutherford JC, Lill R, Winge D. Role of glutaredoxin-3 and glutaredoxin-4 in the iron regulation of the Aft1 transcriptional activator in Saccharomyces cerevisiae. J Biol Chem. 2006;281:17661–17669. doi: 10.1074/jbc.M602165200. [DOI] [PubMed] [Google Scholar]
- Pal KK, Tilak KV, Saxena AK, Dey R, Singh CS. Suppression of maize root diseases caused by Macrophomina phaseolinaFusarium moniliforme and Fusarium graminearum by plant growth promoting rhizobacteria. Microbiol Res. 2001;156:209–223. doi: 10.1078/0944-5013-00103. [DOI] [PubMed] [Google Scholar]
- Perrin DD. Stability Constants: Part B. Pergamon, Turkey: IUPAC; 1979. [Google Scholar]
- Pesce AJ, Kaplan LA. MPtodos Qubnicu Clinica. Buenos Aires, Argentina: MédicaPanamericana Ed; 1990. [Google Scholar]
- Peterson RL, Massicotte HB, Melville LH. Mycorrhizas: Anatomy and Cell Biology. Ottawa, Canada: NRC Research Press; 2004. [Google Scholar]
- Rajkumar M, Ae N, Freitas H. Endophytic bacteria and their potential to enhance heavy metal phytoextraction. Chemosphere. 2009;77:153–160. doi: 10.1016/j.chemosphere.2009.06.047. [DOI] [PubMed] [Google Scholar]
- Rajkumar M, Ae N, Prasad MNV, Freitas H. Potential of siderophore-producing bacteria for improving heavy metal phytoextraction. Trends Biotechnol. 2010;28:142–149. doi: 10.1016/j.tibtech.2009.12.002. [DOI] [PubMed] [Google Scholar]
- Rajkumar M, Sandhya S, Prasad MNV, Freitasc H. Perspectives of plant-associated microbes in heavy metal phytoremediation. Biotechnol Adv. 2012;30:1562–1574. doi: 10.1016/j.biotechadv.2012.04.011. [DOI] [PubMed] [Google Scholar]
- Raymond K. Biomimetic metal encapsulation. Coord Chem Rev. 1990;105:135–153. [Google Scholar]
- Raymond KN, Müller G, Matzanke BF. Complexation of iron by siderophores a review of their solution and structural chemistry and biological function Structural Chemistry. In: Boschke FL, editor. Topics in Current Chemistry. Vol. 123. Berlin, Heidelberg, Germany: Springer-Verlag; 1984. pp. 49–102. [Google Scholar]
- Reichard PU, Kraemer SM, Frazier SW, Kretzschmar R. Goethite dissolution in the presence of phytosiderophores: rates, mechanisms, and the synergistic effect of oxalate. Plant Soil. 2005;276:115–132. [Google Scholar]
- Reichard PU, Kretzschmar R, Kraemer SM. Dissolution mechanisms of goethite in the presence of siderophores and organic acids. Geochim Cosmochim Acta. 2007;71:5635–5650. [Google Scholar]
- Renshaw JC, Robson GD, Trinci APJ, Wiebe MG, Livens FR, Collison D, Taylor RJ. Fungal siderophores: structures, functions and applications. Mycol Res. 2002;106:1123–1142. [Google Scholar]
- Robert M, Chenu C. Interactions between soil minerals and microorganisms. In: Stotzky G, Bollag J-M, editors. Soil Biochemistry 7. New York, USA: Marcel Dekker; 1992. pp. 307–404. [Google Scholar]
- Roberts Rogers J, Bennett PC. Mineral stimulation of subsurface microorganisms: release of limiting nutrients from silicates. Chem Geol. 2004;203:91–108. [Google Scholar]
- Römheld V, Marschner H. Evidence for a specific uptake system for iron phytosiderophores in roots of grasses. Plant Physiol. 1986;80:175–180. doi: 10.1104/pp.80.1.175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruggiero CE, Neu MP, Vanderberg LA. Plutonium and uranium interactions with siderophores of aerobic soil microbes. J Inorg Biochem. 1999;74:282. [Google Scholar]
- Schalk IJ, Hannauer M, Braud A. Minireview new roles for bacterial siderophores in metal transport and tolerance. Environ Microbiol. 2011;13:2844–2854. doi: 10.1111/j.1462-2920.2011.02556.x. [DOI] [PubMed] [Google Scholar]
- Schenk PM, Carvalhais LC, Kazan K. Unraveling plant–microbe interactions: can multi-species transcriptomics help? Trends Biotechnol. 2012;30:177–184. doi: 10.1016/j.tibtech.2011.11.002. [DOI] [PubMed] [Google Scholar]
- Schippers B, Bakker AW, Bakker PAH. Interactions of deleterious and beneficial rhizosphere microorganisms and the effect of cropping practices. Ann Rev Phytopathol. 1987;25:339–358. [Google Scholar]
- Schwarzenbach G, Schwarzenbach K. Hydroxamate complexes. I. The stabilities of the iron (III) complexes of simple hydroxamic acids and desferriferrioxamine B. Helv Chim Acta. 1963;46:1390–1400. [Google Scholar]
- Schwyn B, Neilands JB. Universal chemical assay for the detection and determination of siderophores. Anal Biochem. 1987;160:47–56. doi: 10.1016/0003-2697(87)90612-9. [DOI] [PubMed] [Google Scholar]
- Sharma M, Gohil NK. Optical features of the fluorophore azotobactin: Applications for iron sensing in biological fluids. Eng Life Sci. 2010;10:304–310. [Google Scholar]
- Shirvani M, Nourbakhsh F. Desferrioxamine-B adsorption to and iron dissolution from palygorskite and sepiolite. Appl Clay Sci. 2010;48:393–397. [Google Scholar]
- Siebner-Freibach H, Hadar Y, Chen Y. Siderophores sorbed on Ca-montmorillonite as an iron source for plants. Plant Soil. 2003;251:115–124. [Google Scholar]
- Simões LC, Simões M, Vieira MJ. Biofilm interactions between distinct bacterial genera isolated from drinking water. Appl Environ Microbiol. 2007;73:6192–6200. doi: 10.1128/AEM.00837-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh G, Ahuja N, Batish M, Capalash N, Sharma P. Biobleaching of wheat straw-rich soda pulp with alkalophilic laccase from γ-proteobacterium JB: Optimization of process parameters using response surface methodology. Bioresource Technol. 2008;99:7472–7479. doi: 10.1016/j.biortech.2008.02.023. [DOI] [PubMed] [Google Scholar]
- Singh JS, Pandey VC, Singh DP. Efficient soil microorganisms: a new dimension for sustainable agriculture and environmental development. Agr Ecosys Environ. 2011;140:339–353. [Google Scholar]
- Sokolova TA, Tolpeshta II, Topunova IV. Biotite weathering in podzolic soil under conditions of a model field experiment. Eurasian Soil Sci. 2010;43:1150–1158. [Google Scholar]
- Sugita H, Hirose Y, Matsuo N, Deguchi Y. Production of the antibacterial substance by Bacillus sp strain NM 12, an intestinal bacterium of Japanese coastal fish. Aquaculture. 1998;165:269–280. [Google Scholar]
- Sugita H, Mizuki H, Itoi S. Diversity of siderophore-producing bacteria isolated from the intestinal tracts of fish along the Japanese coast. Aquac Res. 2012;43:481–488. [Google Scholar]
- Takemoto T, Nomoto K, Fushiya S, Ouchi R, Kusano G, Hikino H, et al. Structure of mugineic acid, a new amino acid possessing an iron-chelating activity from roots washings of water-cultured Hordeum vulgare L. Proc Jpn Acad Ser B. 1978;54:469–473. [Google Scholar]
- Taylor RJ, May I. The reduction of actinide ions by hydroxamic acids. Czech J Phys. 1999;49:617–621. [Google Scholar]
- Taylor RJ, May I, Wallwork AL, Denniss IS, Hill NJ, Galkin BY, et al. The applications of formo-and aceto-hydroxamic acids in nuclear fuel reprocessing. J Alloy Compd. 1998;271–273:534–537. [Google Scholar]
- Trick CG, Andersen RJ, Gillam A, Harrison PJ. Prorocentrin: an extracellular siderophore produced by the marine dinoflagellate Prorocentrum minimum. Science. 1983;219:306–308. doi: 10.1126/science.219.4582.306. [DOI] [PubMed] [Google Scholar]
- Van der Helm D, Winkelmann G. Hydroxamates and polycarboxylates as iron transport agents (siderophores) in fungi. In: Winkelmann G, Winge D, editors. Metal Ions in Fungi. New York, USA: Marcel Dekker; 1994. pp. 39–98. [Google Scholar]
- Van Schöll L, Kuyper TW, Smits MM, Landeweert R, Hoffland E, Breemen van N. Rock-eating mycorrhizas: their role in plant nutrition and biogeochemical cycles. Plant Soil. 2008;303:35–47. [Google Scholar]
- Verma VC, Singh SK, Prakash S. Bio-control and plant growth promotion potential of siderophore producing endophytic Streptomyces from Azadirachta indica A. Juss. J Basic Microbiol. 2011;51:550–556. doi: 10.1002/jobm.201000155. [DOI] [PubMed] [Google Scholar]
- Voisard C, Keel C, Haas D, Defago G. Cyanide production by Pseudomonas fluorescens helps suppress black root rot of tobacco under gnotobiotic conditions. EMBO J. 1989;8:351–358. doi: 10.1002/j.1460-2075.1989.tb03384.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang L, Yan W, Chen J, Huang F, Gao P. Function of the iron-binding chelator produced by Coriolus versicolor in lignin biodegradation. Sci China C Life Sci. 2008;51:214–221. doi: 10.1007/s11427-008-0033-9. [DOI] [PubMed] [Google Scholar]
- Wang Q, Xiong D, Zhao P, Yu X, Tu B, Wang G. Effect of applying an arsenic-resistant and plant growth-promoting rhizobacterium to enhance soil arsenic phytoremediation by Populus deltoides LH05–17. J Appl Microbiol. 2011;111:1065–1074. doi: 10.1111/j.1365-2672.2011.05142.x. [DOI] [PubMed] [Google Scholar]
- Wasi S, Tabrez S, Ahmad M. Toxicological effects of major environmental pollutants: an overview. Environ Monit Assess. 2013;185:2585–2593. doi: 10.1007/s10661-012-2732-8. [DOI] [PubMed] [Google Scholar]
- Watteau F, Berthelin J. Microbial dissolution of iron and aluminum from soil minerals – efficiency and specificity of hydroxamate siderophores compared to aliphatic acids. Eur J Soil Biol. 1994;30:1–9. [Google Scholar]
- Weinberg ED. Suppression of bacterial biofilm formation by iron limitation. Med Hypotheses. 2004;63:863–865. doi: 10.1016/j.mehy.2004.04.010. [DOI] [PubMed] [Google Scholar]
- Wichard T, Bellenger JP, Morel FM, Kraepiel AM. Role of the siderophore azotobactin in the bacterial acquisition of nitrogenase metal cofactors. Environ Sci Technol. 2009;43:7218–7224. doi: 10.1021/es8037214. [DOI] [PubMed] [Google Scholar]
- Winkelmann G. Ecology of siderophores with special reference to the fungi. Biometals. 2007;20:379–392. doi: 10.1007/s10534-006-9076-1. [DOI] [PubMed] [Google Scholar]
- Winkelmann G, Huschka HG. Molecular recognition and transport of siderophores in fungi. In: Winkelmann G, Helm van der D, Neilands JB, editors. Iron Transport in Microbes, Plants and Animals. Weinheim, Germany: VCH; 1987. pp. 317–336. [Google Scholar]
- Xu G, Goodell B. Mechanisms of wood degradation by brown-rot fungi: chelator-mediated cellulose degradation and binding of iron by cellulose. J Biotechnol. 2001;87:43–57. doi: 10.1016/s0168-1656(00)00430-2. [DOI] [PubMed] [Google Scholar]
- Yadav S, Kaushik R, Saxena AK, Arora DK. Diversity and phylogeny of plant growth-promoting bacilli from moderately acidic soil. J Basic Microbiol. 2011;51:98–106. doi: 10.1002/jobm.201000098. [DOI] [PubMed] [Google Scholar]
- Yano T. The nonspecific immune system: humoral defense. In: Iwama G, Nakanishi T, editors. The Fish Immune System: Organism, Pathogen, and Environment. San Diego, CA, USA: Academic Press; 1996. pp. 105–157. [Google Scholar]
- Yoder MF, Kisaalita WS. Iron specificity of a biosensor based on fluorescent pyoverdin immobilized in sol-gel glass. J Biol Eng. 2011;5:4–16. doi: 10.1186/1754-1611-5-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu X, Ai C, Xin L, Zhou G. The siderophore-producing bacterium, Bacillus subtilis CAS15, has a biocontrol effect on Fusarium wilt and promotes the growth of pepper. Eur J Soil Biol. 2011;47:138–145. [Google Scholar]