

Abstract
In this study, we developed a gene disruption system for Thermococcus barophilus using simvastatin for positive selection and 5-fluoroorotic acid (5-FOA) for negative selection or counterselection to obtain markerless deletion mutants using single- and double-crossover events. Disruption plasmids carrying flanking regions of each targeted gene were constructed and introduced by transformation into wild-type T. barophilus MP cells. Initially, a pyrF deletion mutant was obtained as a starting point for the construction of further markerless mutants. A deletion of the hisB gene was also constructed in the UBOCC-3256 (ΔpyrF) background, generating a strain (UBOCC-3260) that was auxotrophic for histidine. A functional pyrF or hisB allele from T. barophilus was inserted into the chromosome of UBOCC-3256 (ΔpyrF) or UBOCC-3260 (ΔpyrF ΔhisB), allowing homologous complementation of these mutants. The piezophilic genetic tools developed in this study provide a way to construct strains with multiple genetic backgrounds that will allow further genetic studies for hyperthermophilic piezophilic archaea.
INTRODUCTION
Since the discovery of deep-sea hydrothermal vents, many mesophilic, thermophilic, and hyperthermophilic Bacteria and Archaea have been described. However, only a few thermopiezophilic organisms have been described so far, mainly belonging to the domain Archaea: Thermococcus barophilus (1), Palaeococcus pacificus (2), Palaeococcus ferrophilus (3), Marinitoga piezophila (4), Methanopyrus kandleri (5), and Pyrococcus yayanosii (6, 7). P. yayanosii is the first and only known obligate piezophilic hyperthermophilic archaeon isolated from a deep-sea hydrothermal vent. The genomes of M. kandleri, T. barophilus, P. yayanosii, and M. piezophila are now available (8–11), but the development of genetic tools for the above species is lagging.
T. barophilus strain MP was the first true hyperthermophilic piezophilic archaeon isolated, in 1993 (1); it grows in rich medium from 48°C to 100°C, with an optimum at 85°C, and within a pressure range of 0.1 to 85 MPa, with an optimum of 40 MPa (1). T. barophilus is an obligate piezophile for temperatures over 95°C.
Genetic manipulations in nonpiezophilic members of the Thermococcales, such as Thermococcus kodakarensis and Pyrococcus furiosus, which are phylogenetically related to T. barophilus, have been described (12–14). For some archaeal genetic systems, such as T. kodakarensis, many selectable markers have been described, allowing selection according to prototrophic markers (arginine/citrulline, tryptophan, agmatine, or uracil) or antibiotics (simvastatin or mevinolin) (15); simvastatin or mevinolin can be used to screen for transformed cells. Such cells can overexpress the P. furiosus hmg gene (hmgPf), which encodes 3-hydroxy-3-methylglutaryl coenzyme A (HMG-CoA) reductase, a pivotal enzyme for archaeal membrane lipid biosynthesis (16), so that the hmgPf gene can then be used as a positive selection marker (17). Among prototrophic selective markers, arginine/citrulline-based selection is used when strains are able to transform aspartate into arginine using citrulline supplied by the culture medium and thus achieve arginine prototrophy (15). Agmatine, a polyamine produced by decarboxylation of arginine, is used as a marker for positive selection for agmatine prototrophs in strains from which the argD gene encoding arginine decarboxylase has been deleted (18). Agmatine auxotrophy is lethal, even in rich medium, which facilitates the isolation of transformants overnight on rich medium, in contrast to the several days required when transformants are selected on defined medium. As for other types of prototrophic selection, tryptophan-based selection is limited to defined medium and plasmid introduction into a specific strain from which the trpE gene, encoding the large subunit of anthranilate synthase, has previously been deleted (12).
The uracil marker is used in all model organisms for genetics in Archaea (17). The pyrEF genes enable complementation of the uracil auxotrophy, and 5-fluoroorotic acid (5-FOA) can be used as a counterselection marker; the uracil marker is functional for both positive selection and counterselection (negative selection) for constructing markerless deletion mutants. A similar counterselective strategy is also offered by 6-methyl purine (6-MP) (18), which must be paired with a positive selection marker because it cannot be used for positive selection itself. All these markers have at least one disadvantage, such as limitation to use in defined medium, supplement requirement, spontaneous resistance providing a high background, limited host range, and/or no consecutive positive selection and counterselection (15).
Thus, the development of genetic manipulations of the deep-sea hydrothermal vent strain T. barophilus MP, proposed in this study, was derived from what we knew about gene deletion systems established in the shallow hydrothermal vent species T. kodakarensis. A suicide vector was constructed to be used as a tool for gene disruption by homologous recombination in T. barophilus. We showed that 1 kb can be efficiently integrated directly into the chromosome by using circular DNA fragments. A strategy was developed to generate deletion mutants by PCR amplification and vector cloning to select marker replacement events with subsequent disruption, or “pop-out,” of the selected marker. The genetic manipulations in T. barophilus established here rely on uracil auxotrophy and simvastatin resistance as selectable markers, using the pop-in/pop-out method. We constructed multiple gene mutants and tested the possibility of complementation in trans.
MATERIALS AND METHODS
Media and growth conditions.
T. barophilus strain MP was isolated from chimney samples harvested from the Snake Pit hydrothermal site, at a depth of 3,550 m on the mid-Atlantic ridge, in July 1993 (1). T. barophilus cultures were grown under anaerobic conditions at 85°C in Thermococcales rich medium (TRM) (7) or Thermococcales amino acid (TAA) medium (A. Cario and P. Oger, unpublished data) supplemented with sulfur. The TAA medium composition is as follows: 23 g NaCl, 3.3 g PIPES [piperazine-N,N′-bis(2-ethanesulfonic acid)], 3 g MgCl2 · 6H2O, 2 g C2H3NaO2, 1 g (NH4)2SO4, 0.5 g KCl, 0.05g NaBr, 0.02 g SrCl2 · 6H2O, vitamin mixture, modified Wolfe's trace minerals, 1 ml resazurin, and 0.1 g per liter of each of 19 amino acids (except histidine). After cell transformation, the mutants were selected on TRM supplemented with 2.5 μg/ml simvastatin (Sigma) or 8 mg/ml 5-FOA (5-fluoroorotic acid hydrate; Euromedex). Auxotrophic growth assays were performed on TAA medium with the addition of uracil in the presence or absence of histidine, as appropriate. After filtration (Millipore filter, 0.45 μm), the liquid medium was dispensed anaerobically into 50-ml vials which were sealed with butyl-rubber stoppers, and the medium was reduced with 0.1 ml of a 10% (wt/vol) sterile Na2S · 9H2O solution just before inoculation. Unless stated otherwise, the experiments were carried out in triplicate in the presence of sulfur.
A pressure ranging from 0.1 MPa to 70 MPa was used to monitor the growth of mutants in TRM and TAA medium; cultivation under hydrostatic pressure was performed in sterile syringes, and cultures were incubated in a high-hydrostatic-pressure (HHP), high-temperature incubator (Top Industrie), as previously described (7).
Growth was monitored by cell counting using a Thoma chamber and photonic microscopy at a magnification of ×40 (Olympus) or using flow cytometry (CyFlow Space; Partec). Cells were fixed with 2.5% glutaraldehyde and counted by one of the two previously described methods.
The genetic manipulations, DNA preparation, and transformation protocols were conducted under atmospheric pressure (0.1 MPa). Escherichia coli strain DH5α was used for general DNA manipulation, E. coli was cultured in LB medium (liquid or solid) at 37°C, and the presence of plasmids was selected for by adding 100 μg/ml ampicillin to the medium.
Construction of suicide vectors.
The pUD plasmid was kindly provided by the Imanaka lab (19). This plasmid bears the ampicillin resistance gene and the T. kodakarensis pyrF gene with its putative promoter region; it is a replicative plasmid in E. coli but not in Thermococcales strains.
The hmgPf gene was obtained by PCR amplification on the pLC70 plasmid (20) with the primers SalI-HMG-CoA-Up and KpnI-HMG-CoA-Do (see Table S1 in the supplemental material); the resulting PCR product bears the KpnI and SalI restriction enzyme sites at its extremities. The hmgPf amplicon was cloned in the pUD plasmid digested by the restriction enzymes KpnI and SalI, and after ligation between the pUD plasmid and the hmgPf PCR-amplified gene (using T4 ligase; Promega), a plasmid named pUDH was constructed (Fig. 1).
FIG 1.
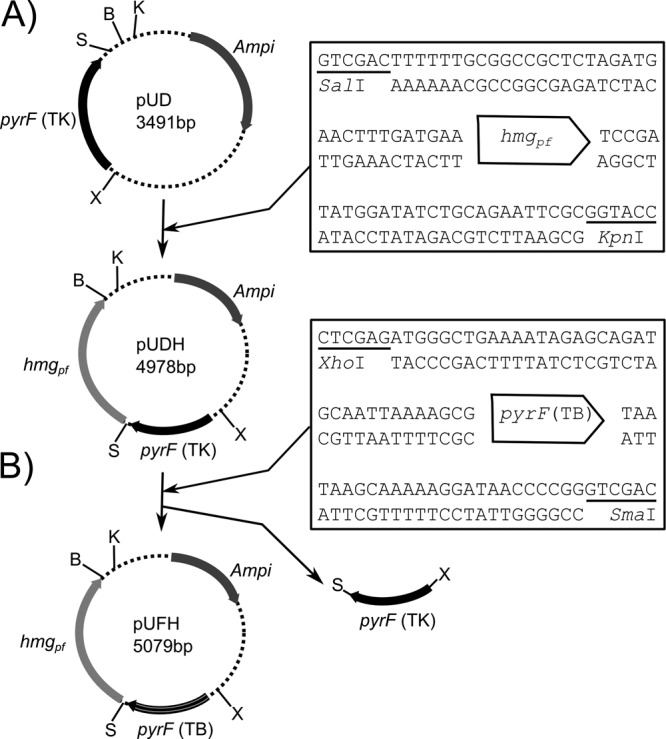
Construction of pUDH and pUFH plasmids. (A) Primers SalI-HMG-CoA-Up and KpnI-HMG-CoA-Do were used to amplify hmgPf from the vector pLC70 (20). pUD and HMG-CoA were digested by SalI (S) and BamHI (B) and were then ligated to obtain the plasmid pUDH. (B) Primers XhoI-pyrF-TB-Up and XmaISmaI-pyrF-TB-Do were used to amplify the T. barophilus pyrF gene. pUDH and pyrF(TB) were digested by XhoI (X) and SmaI (S) and were then ligated to obtain the plasmid pUFH. In pUDH and pUFH, the restriction enzyme sites BamHI (B) and KpnI (K) were conserved to enable cloning of the homologous regions in these plasmids.
In the second step of this work, the pUDH plasmid was digested by XhoI and SmaI enzymes. This digestion linearized the plasmid, allowing the excision of the T. kodakarensis pyrF gene but not its promoter region. The linearized plasmid was ligated with the PCR-amplified T. barophilus pyrF gene obtained with the primers XhoI-pyrF-TB-Up and SmaI-pyrF-TB-Do from genomic DNA, and the resulting plasmid was named pUFH (Fig. 1).
The homologous regions encompassing the target genes to be excised were amplified by two successive PCR amplification steps, in a process known as splicing by overlap extension (21, 22). After the first amplification, two fragments of 1 kb encompassing the targeted genes were obtained; the primers used for this first amplification were named 1Up/1Do and 2Up/2Do (see Table S1 in the supplemental material). The two fragments were used as the matrix for the second PCR amplification, by using the primers 1Up and 2Do (see Table S1 in the supplemental material), and the resulting PCR product of the second step was a DNA fragment of 2 kb composed of the two merged homologous regions. The different primers used for amplified homologous regions were ΔpyrF-1Up, ΔpyrF-1Do, ΔpyrF-2Up, and ΔpyrF-2Do for deletion of pyrF (TERMP_01290); ΔhisB-1Up, ΔhisB-1Do, ΔhisB-2Up, and ΔhisB-2Do for deletion of hisB (TERMP_00437); and ΔTERMP_00005-1Up, ΔTERMP_00005-1Do, ΔTERMP_00005-2Up, and ΔTERMP_00005-2Do for the deletion of TERMP_00005 (see Table S1 in the supplemental material). By using this approach, three homologous-region fragments were created: the ΔpyrF, ΔhisB, and ΔTERMP_00005 fragments. The fragment TERMP_01289(HR1)-TERMP_01291(HR2) was cloned into the plasmid pUDH using the restriction sites KpnI and BglII (on the amplification)/BamHI (on the plasmid), and the fragments TERMP_00436(HR1)-TERMP_00438(HR2) and TERMP_00004(HR1)-TERMP_00006(HR2) were cloned into the plasmid pUFH using the restriction sites KpnI and BglII/BamHI; we thus obtained the suicide vectors pUDH-1, pUFH-1, and pUFH-2 (Table 1).
TABLE 1.
T. barophilus plasmids used and constructed in this study
| Plasmids | Resistance | Parent plasmid | Description | Reference |
|---|---|---|---|---|
| pUD | Ampicillin | 19 | ||
| pUDH | Ampicillin, simvastatin | pUD | Cloning of HMG-CoA reductase-encoding gene in pUD | This work |
| pUFH | Ampicillin, simvastatin | pUDH | Replacement of pyrF (T. kodakarensis) by pyrF (T. barophilus) | This work |
| pUDH-1 | Ampicillin, simvastatin | pUDH | Cloning of homologous regions flanking pyrF | This work |
| pUFH-1 | Ampicillin, simvastatin | pUFH | Cloning of homologous regions flanking hisB | This work |
| pUFH-2 | Ampicillin, simvastatin | pUFH | Cloning of homologous regions flanking TERMP_00005 | This work |
| pUHH | Ampicillin, simvastatin | pUFH-2 | Replacement of pyrF by hisB | This work |
In order to complement the hisB mutant, the pyrF gene present in pUFH but not its promoter region was replaced by the T. barophilus hisB gene; the hisB gene was amplified using the primers Verif-hisB-Up and Verif-hisB-Do (see Table S1 in the supplemental material). The hisB PCR product and pUFH vector were digested by XhoI and SmaI and then ligated (using T4 ligase; Promega), and the resulting plasmid was named pUHH (Table 1).
Transformation of T. barophilus.
The CaCl2 method for Methanococcus voltae PS (23) was modified for the transformation of T. barophilus. The CaCl2 cell treatment was not a prerequisite for the transformation. Cells were cultivated in liquid TRM with sulfur for 16 h at 85°C at atmospheric pressure, and an aliquot of 1 ml of this overnight culture was introduced into 50 ml of fresh TRM without sulfur and incubated for 6 h at 85°C. Cells were harvested by centrifugation (8,000 × g, 6 min), concentrated in 1 ml of fresh TRM without sulfur, and kept on ice for 30 min under an anaerobic tent. An aliquot of 4 to 5 μg of plasmid DNA was added to 200 μl of concentrated cells, and the mixture was incubated on ice for 1 h. A heat shock at 85°C was carried out for 10 min, followed by incubation for 10 min on ice. The transformants were then used to inoculate 20 ml of fresh TRM with sulfur and incubated at 85°C for 18 h. The cells were harvested by centrifugation (8,000 × g, 6 min), resuspended in 100 μl of fresh TRM, and spread on plated TRM containing simvastatin (final concentration of 2.5 μg/ml) or 5-FOA (8 mg/ml) under anaerobic conditions. Medium solidification was achieved using Phytagel (Sigma) at a concentration of 10 g/liter, and the plates were incubated for 5 days at 85°C. The resulting Simr strains were analyzed by PCR using genomic DNA as a matrix.
Complementation of pyrF and hisB mutants.
The pUFH-2 and pUHH plasmids were introduced, respectively, into UBOCC-3256 (ΔpyrF) and UBOCC-3260 (ΔpyrF ΔhisB). Many transformants that were resistant to simvastatin were obtained, and after PCR screening, all clones had undergone single-crossover recombination (pyrF+ or hisB+) at one or the other of the homologous regions (TERMP_00004 or TERMP_00006) carried by both plasmids (data not shown).
Pop-out recombination.
After the transformation of T. barophilus by a suicide vector (Fig. 2), two steps were needed to perform the excision of the targeted gene, a pop-in (vector insertion) by one crossover event and a pop-out recombination. For this purpose, a strain with a suicide vector inserted was streaked onto TRM plates supplemented with 5-FOA at 8 mg/ml. In principle, the strains that are able to grow on this medium are obviously resistant to 5-FOA, sensitive to simvastatin, and ΔpyrF; however, we observed several false positives at this step. To address this problem, the colonies that grew on TRM–5-FOA plates were restreaked on TRM-simvastatin plates to eliminate the false positives that were still simvastatin resistant. The strains which were simvastatin sensitive and 5-FOA resistant were kept; the pop-out recombination was then obtained, and a PCR amplification was performed to check whether the pop-out recombination resulted in a targeted gene deletion or wild type (WT) genotype recurrence (Fig. 2).
FIG 2.

Deletion pathway of pyrF and hisB genes. (A) Two suicide vectors were constructed to ligate homologous-region amplification (TERMP_01289, TERMP_01291, TERMP_00436, and TERMP_00438) with pUFH or pUDH. The plasmids used were pUDH-1 and pUFH-1. After transformation, the plasmid was integrated into the genome by a first crossover event in the homologous-region fragment. The second step was the pop-out recombination (or excision) event. There are two possibilities: a recombination between the other homologous fragments, resulting in the deletion of the targeted gene, or a recombination between the same homologous fragments of the first recombination, which gives the WT genotype. pyrF was deleted from strain UBOCC-3107 (WT), and hisB was deleted from strain UBOCC-3256 (ΔpyrF). (B) To verify the different genotype configurations, PCR amplification was performed with the primers matching the HR1 and HR2 regions: XhoI-pyrF-TB-Up and SmaI-pyrF-TB-Do for pyrF deletion and Verif-hisB-Up and Verif-hisB-Do for hisB deletion (data not shown).
DNA purification and extraction.
Plasmid DNA was extracted from an E. coli DH5α strain with a plasmid extraction kit supplied by Thermofisher. E. coli DH5α cultures (5 ml) at exponential growth phase were harvested after centrifugation (6,000 × g, 6 min, 4°C) and then used to extract DNA vectors with a final concentration of 160 to 200 ng/μl.
Genomic DNA extraction of T. barophilus was performed using a phenol-chloroform-isoamyl alcohol (PCI) extraction method, as follows. A 20-ml portion of cell culture at exponential growth phase (approximately 108 cells/ml) was centrifuged at 7,500 × g for 15 min at 4°C. The cell pellet was resuspended in 800 μl TE buffer (100 mM Tris HCl [pH 8], 50 mM NaCl, 50 mM EDTA [pH 8]). To ensure cell lysis, 100 μl of 10% SDS, 100 μl of 10% Sarkosyl, and 50 μl of proteinase K (20 mg/ml) were added, and the cell suspension was then incubated for 1 h at 55°C; 1 ml phenol-chloroform-isoamyl alcohol (PCI) (25:24:1) was added, and after centrifugation at 14,000 × g for 15 min at 4°C, the aqueous phase containing the total DNA was recovered, to which 1 ml of chloroform was then added to eliminate all phenol traces. The DNA was subsequently precipitated by the addition of 0.7 ml isopropanol for 1 h at −20°C and recovered after centrifugation (14,000 × g for 15 min at 4°C). The DNA was then washed with 75% ethanol, and after centrifugation (14,000 × g for 15 min at 4°C), it was resuspended in 200 μl 10 mM Tris-HCl (pH 8) buffer; RNase (50 mg/ml) was added to eliminate residual RNA in each sample.
The DNA was quantified by NanoDrop, and the quality of extraction was checked by electrophoresis on a 1% agarose gel containing ethidium bromide at a final concentration of 0.5 mg/ml (in a bath of 40 mM Tris [pH 8], 40 mM acetate, 1 mM EDTA [pH 8] [1× TAE]). The separation was performed at 85 V for 40 min, with a 1-kb ladder (Promega) as the size marker.
PCR conditions.
hmgPf, pyrF, hisB, and all homologous regions were amplified using Pfu polymerase (Promega). Routine tests by PCR amplification were performed using Taq Polymerase (Promega), and PCR was performed as follows: 94°C for 5 min; 30 cycles of 94°C for 60 s, 50 to 58°C for 60 s, and 72°C for 1 to 4 min; 72°C for 10 min.
RESULTS
Effects of different simvastatin and 5-FOA concentrations on the growth of T. barophilus.
To select the markers that could be used for positive-selection integration and counterselection excision from the T. barophilus genome, the sensitivity of T. barophilus to different drugs and antibiotics was assessed using 5-FOA, 6-MP, simvastatin, and mevinolin. Surprisingly, T. barophilus was insensitive to the drug 6-MP, even though it possesses a gene (TERMP_00517) (9) annotated as encoding a hypoxanthine-guanine phosphoribosyl-transferase. TERMP_00517 is a homologue of the TK0664 gene in T. kodakarensis (80% of identity), the inactivation of which results in 6-MP resistance in T. kodakarensis (18). In contrast to T. kodakarensis, the T. barophilus MP strain is auxotrophic for tryptophan, and it is not amenable to tryptophan-based positive selection, because this WT species possesses only one gene (TERMP_00157) encoding a Trp synthase-like β subunit and lacks six genes (among them trpE) encoding enzymes that transform chorismate to tryptophan (9, 24).
T. barophilus MP seems to be sensitive to 5-FOA concentrations ranging from 5 to 8 mg/ml, but only during 24 h of growth on solid rich medium and less than 10 h of growth on liquid rich medium (see Fig. S2A in the supplemental material), beyond which cell growth was observed, probably due to uracil contamination or drug thermal degradation, which gives high backgrounds, as was reported for T. kodakarensis and other Archaea (15, 17). The uracil marker is functional in T. barophilus, and a concentration as high as 8 mg/ml was chosen, at least for negative selection.
In comparison with T. kodakarensis, sensitivity to simvastatin was assessed in T. barophilus using drug concentrations ranging from 1 to 10 μg/ml. As previously described, 5 μg/ml of simvastatin was needed to observe a total inhibition of T. kodakarensis growth (13). In contrast, 2.5 μg/ml of simvastatin was sufficient to inhibit the growth of T. barophilus cells for at least 5 days of incubation on both solid and liquid media (see Fig. S2B in the supplemental material), indicating that this concentration would be suitable and sufficient for selecting Simr transformants and that simvastatin could be used for positive selection.
Construction of gene deletion plasmids.
To achieve gene disruption in T. barophilus, two plasmids, pUDH and pUFH, were designed for double-crossover events (Fig. 1) and constructed using the plasmid pUD (19) (see Materials and Methods). The pUDH plasmid contains a marker cassette containing the pyrF gene from T. kodakarensis and an hmgPf cassette (Fig. 1A), whereas the pUFH plasmid contains the same hmgPf cassette and the pyrF gene from T. barophilus (Fig. 1B). These two plasmids were replicative in E. coli and conferred ampicillin resistance but were not replicative in T. barophilus. The strategy for construction of a targeted gene excision strain is shown in Fig. 1. The pop-in/pop-out method was used in T. barophilus; by this method, integration of constructs into the genome (pop-in) is selected for by transformation to simvastatin resistance, and intramolecular recombinants that have lost the plasmid (pop-out) are counterselected using 5-FOA.
Construction of pyrF and hisB mutants.
The new constructs pUDH and pUFH were used to clone the flanking regions of the targeted genes pyrF and hisB, respectively, and the resulting plasmids were named pUDH-1 and pUFH-1 (Fig. 2A). These plasmids were used to transform T. barophilus MP using simvastatin as the resistant marker (see Materials and Methods). The transformation efficiency was approximately 102 transformants per μg plasmid DNA. The Simr transformants containing plasmids integrated into the T. barophilus chromosome were checked by PCR, and as shown in Fig. 2B, this led to an amplification of two bands: a large one (852 bp), corresponding to the WT allele, and a small one (221 bp), corresponding to the deleted gene (Fig. 2B). After PCR verification, selected clones were spread on solid TRM with 5-FOA (8 mg/ml) and incubated for 4 days at 85°C. About 100 to 300 colonies per plate were obtained, some of which were restreaked on solid TRM containing simvastatin (2.5 μg/ml); 70 to 90% of these were Simr, which demonstrated that the pop-out event had not occurred in these cases. At least 10% of the colonies were Sims and were checked by PCR amplification of a 221-bp fragment, demonstrating that the pop-out event had occurred. (Fig. 2B). One ΔpyrF strain mutant (UBOCC-3256) (Table 2) was selected and will serve as a starting point for further genetic manipulations.
TABLE 2.
T. barophilus strains used and constructed in this study
| Strain | Genotype | Parent strain | Genome region(s) deleted from parent strain | Reference |
|---|---|---|---|---|
| UBOCC-3107 | Wild type | T. barophilus MP | 1 | |
| UBOCC-3256 | ΔpyrF | UBOCC-3107 | TERMP_01290 (nucleotides 1122862–1123491) | This work |
| UBOCC-3260 | ΔpyrF ΔhisB | UBOCC-3256 | TERMP_00437 (nucleotides 362319–362849) | This work |
| UBOCC-3262 | ΔpyrF TERMP_00006::pUFH-2 | UBOCC-3256 | This work | |
| UBOCC-3265 | ΔpyrF ΔhisB TERMP_00004::pUHH | UBOCC-3260 | This work |
In order to delete the hisB gene, the plasmid pUFH-1 was used to transform the strain UBOCC-3256 by following the strategy described above for pUDH-1. Similar frequencies of pop-in and pop-out events were obtained, and at least three Sims mutants were verified by PCR to confirm the excision of the hisB gene (data not shown). One strain, named UBOCC-3260 (Table 2), was selected for further experiments.
Characterization of mutants.
A comparison of the growth rates and yields of the T. barophilus wild type and T. barophilus strains UBOCC-3256 and UBOCC-3260 in TAA medium in the presence or absence of uracil and/or uracil plus histidine is shown in Fig. 3. Growth experiments were performed at 0.1 MPa, 40 MPa (Fig. 3), and 70 MPa (see Fig. S1 in the supplemental material). Strain UBOCC-3256 (ΔpyrF) did not grow in the absence of uracil but did grow in defined medium supplemented with uracil and showed a growth rate and yield comparable to those of the wild type (Fig. 3). Strain UBOCC-3260 (ΔpyrF ΔhisB) was not able to grow on TAA medium with or without uracil. The growth of this strain resumed when defined medium was supplemented with uracil and histidine (Fig. 3), and the growth rate and yield were similar to those of the wild-type strain. These data demonstrated that UBOCC-3256 and UBOCC-3260 are auxotrophic for uracil and for uracil plus histidine, respectively. The mutants and the WT strains grew better in a defined medium at 40 MPa (Fig. 3), which corresponds to the optimal growth pressure for T. barophilus MP (1), and they also showed growth capabilities that were comparable at 0.1 and 70 MPa (Fig. 3; also, see Fig. S1 in the supplemental material) but slightly lower than at 40 MPa (Fig. 3). The growth yield of the mutants was comparable to that of the WT when cells were grown in TRM regardless of the hydrostatic pressure applied (see Table S2 in the supplemental material). These results demonstrated that pyrF and hisB gene deletion did not show pleiotropic effects in T. barophilus.
FIG 3.

Characterization of UBOCC-3256 (ΔpyrF) and UBOCC-3260 (ΔpyrF ΔhisB) mutants at 0.1 MPa and 40 MPa. Growth assays were carried out in TAA medium at 85°C, without uracil (■), with uracil (●), and with uracil and histidine (▲). These growth experiments were carried out at hydrostatic pressures of 0.1 MPa and 40 MPa.
Complementation of the ΔpyrF and hisB strains by wild-type pyrF and hisB alleles restores uracil and histidine prototrophy.
Several attempts to transform T. barophilus MP with the pLC70 plasmid (20) were unsuccessful. This shuttle vector, known to replicate and to express genes in both T. kodakarensis and E. coli, was constructed by ligating the pTN1 plasmid (25) from Thermococcus nautilus to the commercial vector pCR2.1-TOPO with addition of selectable markers (hmgPf and trpE) (20). Maintenance of the pLC70 plasmid in T. kodakarensis depends on a likely functional replication protein, Rep74, of pTN1 (25), but this plasmid could not be propagated in T. barophilus even though it carries an hmgPf gene, which confers simvastatin resistance to T. barophilus cells. Neither an E. coli/T. barophilus shuttle vector nor a plasmid replicative in T. barophilus is yet available, and these should be constructed in order to expand the genetic toolbox for this species. An alternative complementation strategy was used by introducing pUFH-2 into UBOCC-3256 (ΔpyrF) (Table 1). This plasmid carries the flanking regions of the gene TERMP_00005 and a copy of the pyrF gene from T. barophilus. The construct was integrated into the chromosome of the UBOCC-3256 strain, and transformants were selected for their resistance to simvastatin. Many clones were screened by PCR, and all had undergone single-crossover recombination (pyrF+) at one or the other of the homologous regions (data not shown). One clone was selected and named strain UBOCC-3262 (Table 2). The chromosome of this strain was checked using the primers Verif-int-comp-2Up and Verif-int-comp-2Do (see Table S1 in the supplemental material), and this showed that the plasmid pUFH-2 was inserted in TERMP_00006 gene (data not shown). We examined the growth of T. barophilus MP and its derivatives in TAA medium. The growth rate of UBOCC-3262 was comparable to that of the WT, while UBOCC-3256 could not grow in the absence of uracil (Fig. 4A).
FIG 4.

Complementation of pyrF and hisB mutations. Growth assays of strains UBOCC-3107 (WT) (●), UBOCC-3256 (ΔpyrF) (■), and UBOCC-3262 (ΔpyrF TERMP_00006::pUFH-2) (▲) (A) and of strains UBOCC-3107 (WT) (●), UBOCC-3260 (ΔpyrF-ΔhisB) (■), and UBOCC-3265 (ΔpyrF ΔhisB TERMP_00004::pUHH) (▲) (B) were carried out at 85°C in TAA medium without (A) or with (B) uracil. These growth experiments were performed at 0.1 MPa.
The T. barophilus pyrF gene of pUFH-2 plasmid was replaced by the T. barophilus hisB gene, and the resulting plasmid, pUHH (Table 1), was introduced into UBOCC-3260 (ΔpyrF ΔhisB). A number of transformants were selected for their resistance to simvastatin, and after PCR screening and growth assays (data not shown), one clone was selected and named UBOCC-3265 (Table 2). The chromosome of this strain was checked using the primers Verif-int-comp-1Up and Verif-int-comp-1Do (see Table S1 in the supplemental material), and this showed that the plasmid pUHH was inserted in the TERMP_00004 gene (data not shown). The growth rate and yield of UBOCC-3265 were comparable to those of the WT strain, while UBOCC-3260 could not grow in the absence of histidine (Fig. 4B).
These results demonstrated that pyrF or hisB gene deletion in T. barophilus can be complemented by ectopic integration of a pyrF or hisB allele from T. barophilus.
DISCUSSION
In this study, we report the successful disruption of the pyrF locus in T. barophilus, a hyperthermophilic piezophilic archaeon, and the development of a gene deletion system based on resistance against simvastatin and 5-FOA. Simvastatin was used for positive-selection transformants in nutrient-rich medium for hyperthermophilic and halophilic archaea (13, 26–29). T. barophilus appeared to be more sensitive to this drug than T. kodakarensis (13) and P. furiosus (28). The MIC for the untransformed host was low (1 μg/ml), and the sensitivity of T. barophilus to 5-FOA is comparable to that of T. kodakarensis (19). As in T. kodakarensis and P. furiosus, an effective gene disruption system has been established in T. barophilus, and the ability to use this method to generate single and multiple deletions in the same strain will help analyze and decipher the mechanisms of adaptation to HHP in this important hyperthermophilic piezophilic archaeon. Using circular DNA containing 1 kb of homologous regions, the frequency of transformation for T. barophilus is estimated at 102 transformants per μg DNA, which is comparable to that reported for T. kodakarensis (101 to 102 transformants per μg DNA) (12) but less than that observed for P. furiosus (105 transformants per μg of DNA) (30). The advantage of the method developed here to generate gene deletion in T. barophilus is that the selection and counterselection steps can be performed in rich medium. Even through no spontaneous Simr mutants were generated in T. barophilus, in contrast to what was reported for the genetic manipulation of T. kodakarensis and P. furiosus (13, 15, 30), it is necessary to enrich Simr cells in liquid medium after transformation prior to direct isolation of Simr colonies on plates.
Uracil prototrophic selection can be used for simultaneous transformation and gene deletion in T. barophilus, but the limitation of this selectable marker is the interference caused by background growth of the ΔpyrF strain on solid medium. Simvastatin-based selection is useful for positive selection in T. barophilus, whereas uracil-based negative selection or counterselection suffers from a lack of efficiency in T. barophilus. This pattern has already been observed in other hyperthermophiles: such a counterselective strategy is also available for a 6-methyl purine-based marker, which provides a reliable counterselective pressure in T. kodakarensis and P. furiosus strains from which the xgprt gene (TK0664 and PF1950), encoding a hypoxanthine-guanine phosphoribosyltransferase (20, 31), has been deleted. T. barophilus is insensitive to 6-MP despite the presence of TERMP_00517, which is orthologous to TK0664 (80% identity) and PF1950 (77% identity), in its genome.
As a proof of concept of the efficiency of these genetic tools, the hisB gene, encoding imidazole glycerol-phosphate dehydratase, was deleted in UBOCC-3256. The corresponding mutant showed impaired growth in defined medium in the absence of histidine, and the growth resumed in defined medium where histidine was supplied. Moreover, we introduced a nonreplicative plasmid carrying the pyrF allele from T. kodakarensis or T. barophilus, which was integrated into the ΔpyrF T. barophilus strain by a single crossover into the homologous genomic region of TERMP_00005, and we demonstrated that T. barophilus mutants can be complemented using this strategy until a shuttle vector that can stably replicate and express selectable phenotypes in both T. barophilus and E. coli becomes available.
The nutritional markers targeted in this study provide nutrition selection, and the resulting mutants were auxotrophic regardless of the culture hydrostatic pressure. These genetic tools developed for T. barophilus will help to study the adaptation of T. barophilus to deep-sea hydrothermal-vent conditions, notably HHP. Indeed, transcriptomic studies have highlighted HHP-responsive genes involved in hydrogen production, amino acid uptake and metabolism, sugar uptake and metabolism, and CO assimilation (24). Deletions of genes involved in these pathways in vivo are in progress to examine the roles of related enzymes in hydrostatic pressure adaptation. This will provide greater insight into the mechanisms that have evolved to allow T. barophilus to cope with HHP conditions.
Supplementary Material
ACKNOWLEDGMENTS
We thank Jacques Oberto, Tadayuki Imanaka, Haruyuki Atomi, and Thomas J. Santangelo for helpful discussions and/or for providing vectors and protocols and Mickael Beauverger for his advice and help with performing growth experiments under high hydrostatic pressure. We are indebted to Helen McCombie for helpful language improvement. We also thank Nadège Quintin for the deposition of strains described in this work in the UBO culture collection (http://www.univ-brest.fr/souchotheque/Collection+LM2E).
This work was supported by the Agence Nationale de la Recherche (ANR-10-BLAN-1725 01-Living deep). A.T. was supported by a postdoctoral fellowship from the Conseil Général 29 and from Ifremer. G.M. was supported by a Ph.D. fellowship from the Conseil Régional de Bretagne.
Footnotes
Published ahead of print 31 January 2014
Supplemental material for this article may be found at http://dx.doi.org/10.1128/AEM.00084-14.
REFERENCES
- 1.Marteinsson VT, Birrien JL, Reysenbach AL, Vernet M, Marie D, Gambacorta A, Messner P, Sleytr UB, Prieur D. 1999. Thermococcus barophilus sp. nov., a new barophilic and hyperthermophilic archaeon isolated under high hydrostatic pressure from a deep-sea hydrothermal vent. Int. J. Syst. Bacteriol. 49:351–359. 10.1099/00207713-49-2-351 [DOI] [PubMed] [Google Scholar]
- 2.Zeng X, Zhang X, Jiang L, Alain K, Jebbar M, Shao Z. 2012. Palaeococcus pacificus sp. nov., a novel archaeon from a deep-sea hydrothermal sediment. Int. J. Syst. Evol. Microbiol. 63:2155–2159. 10.1099/ijs.0.044487-0 [DOI] [PubMed] [Google Scholar]
- 3.Takai K, Sugai A, Itoh T, Horikoshi K. 2000. Palaeococcus ferrophilus gen. nov., sp. nov., a barophilic, hyperthermophilic archaeon from a deep-sea hydrothermal vent chimney. Int. J. Syst. Evol. Microbiol. 50:489–500. 10.1099/00207713-50-2-489 [DOI] [PubMed] [Google Scholar]
- 4.Alain K, Marteinsson VT, Miroshnichenko ML, Bonch-Osmolovskaya EA, Prieur D, Birrien JL. 2002. Marinitoga piezophila sp. nov., a rod-shaped, thermo-piezophilic bacterium isolated under high hydrostatic pressure from a deep-sea hydrothermal vent. Int. J. Syst. Evol. Microbiol. 52:1331–1339. 10.1099/ijs.0.02068-0 [DOI] [PubMed] [Google Scholar]
- 5.Takai K, Nakamura K, Toki T, Tsunogai U, Miyazaki M, Miyazaki J, Hirayama H, Nakagawa S, Nunoura T, Horikoshi K. 2008. Cell proliferation at 122 degrees C and isotopically heavy CH4 production by a hyperthermophilic methanogen under high-pressure cultivation. Proc. Natl. Acad. Sci. U. S. A. 105:10949–10954. 10.1073/pnas.0712334105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Birrien JL, Zeng X, Jebbar M, Cambon-Bonavita MA, Querellou J, Oger P, Bienvenu N, Xiao X, Prieur D. 2011. Pyrococcus yayanosii sp. nov., an obligate piezophilic hyperthermophilic archaeon isolated from a deep-sea hydrothermal vent. Int. J. Syst. Evol. Microbiol. 61:2827–2831. 10.1099/ijs.0.024653-0 [DOI] [PubMed] [Google Scholar]
- 7.Zeng X, Birrien JL, Fouquet Y, Cherkashov G, Jebbar M, Querellou J, Oger P, Cambon-Bonavita MA, Xiao X, Prieur D. 2009. Pyrococcus CH1, an obligate piezophilic hyperthermophile: extending the upper pressure-temperature limits for life. ISME J. 3:873–876. 10.1038/ismej.2009.21 [DOI] [PubMed] [Google Scholar]
- 8.Slesarev AI, Mezhevaya KV, Makarova KS, Polushin NN, Shcherbinina OV, Shakhova VV, Belova GI, Aravind L, Natale DA, Rogozin IB, Tatusov RL, Wolf YI, Stetter KO, Malykh AG, Koonin EV, Kozyavkin SA. 2002. The complete genome of hyperthermophile Methanopyrus kandleri AV19 and monophyly of archaeal methanogens. Proc. Natl. Acad. Sci. U. S. A. 99:4644–4649. 10.1073/pnas.032671499 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Vannier P, Marteinsson VT, Fridjonsson OH, Oger P, Jebbar M. 2011. Complete genome sequence of the hyperthermophilic, piezophilic, heterotrophic, and carboxydotrophic archaeon Thermococcus barophilus MP. J. Bacteriol. 193:1481–1482. 10.1128/JB.01490-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Jun X, Lupeng L, Minjuan X, Oger P, Fengping W, Jebbar M, Xiang X. 2011. Complete genome sequence of the obligate piezophilic hyperthermophilic archaeon Pyrococcus yayanosii CH1. J. Bacteriol. 193:4297–4298. 10.1128/JB.05345-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Lucas S, Han J, Lapidus A, Cheng JF, Goodwin LA, Pitluck S, Peters L, Mikhailova N, Teshima H, Detter JC, Han C, Tapia R, Land M, Hauser L, Kyrpides NC, Ivanova N, Pagani I, Vannier P, Oger P, Bartlett DH, Noll KM, Woyke T, Jebbar M. 2012. Complete genome sequence of the thermophilic, piezophilic, heterotrophic bacterium Marinitoga piezophila KA3. J. Bacteriol. 194:5974–5975. 10.1128/JB.01430-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Sato T, Fukui T, Atomi H, Imanaka T. 2005. Improved and versatile transformation system allowing multiple genetic manipulations of the hyperthermophilic archaeon Thermococcus kodakaraensis. Appl. Environ. Microbiol. 71:3889–3899. 10.1128/AEM.71.7.3889-3899.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Matsumi R, Manabe K, Fukui T, Atomi H, Imanaka T. 2007. Disruption of a sugar transporter gene cluster in a hyperthermophilic archaeon using a host-marker system based on antibiotic resistance. J. Bacteriol. 189:2683–2691. 10.1128/JB.01692-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Farkas J, Stirrett K, Lipscomb GL, Nixon W, Scott RA, Adams MWW, Westpheling J. 2012. Recombinogenic properties of Pyrococcus furiosus strain COM1 enable rapid selection of targeted mutants. Appl. Environ. Microbiol. 78:4669–4676. 10.1128/AEM.00936-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hileman TH, Santangelo TJ. 2012. Genetics techniques for Thermococcus kodakarensis. Front. Microbiol. 3:195. 10.3389/fmicb.2012.00195 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Cabrera JA, Bolds J, Shields PE, Havel CM, Watson A. 1986. Isoprenoid synthesis in Halobacterium halobium. J. Biol. Chem. 261:3578–3583 [PubMed] [Google Scholar]
- 17.Leigh JA, Albers S-V, Atomi H, Allers T. 2011. Model organisms for genetics in the domain Archaea: methanogens, halophiles, Thermococcales and Sulfolobales. FEMS Microbiol. Rev. 35:577–608. 10.1111/j.1574-6976.2011.00265.x [DOI] [PubMed] [Google Scholar]
- 18.Santangelo TJ, Cubonová L, Reeve JN, Cubonova L. 2010. Thermococcus kodakarensis genetics: TK1827-encoded beta-glycosidase, new positive-selection protocol, and targeted and repetitive deletion technology. Appl. Environ. Microbiol. 76:1044–1052. 10.1128/AEM.02497-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Sato T, Fukui T, Atomi H, Imanaka T. 2003. Targeted gene disruption by homologous recombination in the hyperthermophilic archaeon Thermococcus kodakaraensis KOD1. J. Bacteriol. 185:210–220. 10.1128/JB.185.1.210-220.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Santangelo TJ, Cubonova L, Reeve JN. 2008. Shuttle vector expression in Thermococcus kodakaraensis: contributions of cis elements to protein synthesis in a hyperthermophilic archaeon. Appl. Environ. Microbiol. 74:3099–3104. 10.1128/AEM.00305-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Higuchi Krummel B, Saiki R. 1988. A general method of in vitro preparation and specific mutagenesis of DNA fragments: study of protein and DNA interactions. Nucleic Acids Res. 16:7351–7367. 10.1093/nar/16.15.7351 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Horton RM, Cai ZL, Ho SN, Pease LR. 1990. Gene splicing by overlap extension: tailor-made genes using the polymerase chain reaction. Biotechniques 8:528–535 [PubMed] [Google Scholar]
- 23.Bertani G, Baresi L. 1987. Genetic transformation in the methanogen Methanococcus voltae PS. J. Bacteriol. 169:2730–2738 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Vannier P. 2012. Ph.D. thesis Université de Bretagne Occidentale, Brest, France [Google Scholar]
- 25.Soler N, Justome A, Quevillon-Cheruel S, Lorieux F, Le Cam E, Marguet E, Forterre P. 2007. The rolling-circle plasmid pTN1 from the hyperthermophilic archaeon Thermococcus nautilus. Mol. Microbiol. 66:357–370. 10.1111/j.1365-2958.2007.05912.x [DOI] [PubMed] [Google Scholar]
- 26.Zheng T, Huang Q, Zhang C, Ni J, She Q, Shen Y. 2012. Development of a simvastatin selection marker for a hyperthermophilic acidophile, Sulfolobus islandicus. Appl. Environ. Microbiol. 78:568–574. 10.1128/AEM.06095-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Zhang C, Whitaker RJ. 2012. A broadly applicable gene knockout system for the thermoacidophilic archaeon Sulfolobus islandicus based on simvastatin selection. Microbiology 158:1513–1522. 10.1099/mic.0.058289-0 [DOI] [PubMed] [Google Scholar]
- 28.Waege I, Schmid G, Thumann S, Thomm M, Hausner W. 2010. Shuttle vector-based transformation system for Pyrococcus furiosus. Appl. Environ. Microbiol. 76:3308–3313. 10.1128/AEM.01951-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Wendoloski D, Ferrer C, Dyall-Smith ML. 2001. A new simvastatin (mevinolin)-resistance marker from Haloarcula hispanica and a new Haloferax volcanii strain cured of plasmid pHV2. Microbiology 147:959–964 [DOI] [PubMed] [Google Scholar]
- 30.Lipscomb GL, Stirrett K, Schut GJ, Yang F, Jenney FE, Scott RA, Adams MWW, Westpheling J. 2011. Natural competence in the hyperthermophilic archaeon Pyrococcus furiosus facilitates genetic manipulation: construction of markerless deletions of genes encoding the two cytoplasmic hydrogenases. Appl. Environ. Microbiol. 77:2232–2238. 10.1128/AEM.02624-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Kreuzer M, Schmutzler K, Waege I, Thomm M, Hausner W. 2013. Genetic engineering of Pyrococcus furiosus to use chitin as a carbon source. BMC Biotechnol. 13:9. 10.1186/1472-6750-13-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.